Introduction
Tumors are insular masses of proliferating cancer
cells that contain complex tissues of multiple distinct cell types
that interact with each other (1).
Neoplastic cells recruit and incorporate adjacent or distant
healthy host cells, such as bone marrow-derived cells (BMDCs) for
support and essential nutrients (2). Neoplastic cells also recruit
fibroblasts, mesenchymal cells, macrophages, endothelial cells,
pericytes, hematopoietic cells and extracellular matrix (3), all of which together with other
components constitute the tumor microenvironment (TME) (1). Tumor host cells exchange cytokines
(such as IL-6, GM-CSF and IL-4), extracellular matrix proteins,
enzymes and vesicles in distinct sizes, which promote tumor
development, progression and metastases (4,5).
Nevertheless, how recruited stromal cells respond to the TME as
well as tumor cells has constituted a matter of investigation in
recent years only and, remains poorly understood.
Fibroblasts are present in various tumors. These
fibroblasts are termed cancer-associated fibroblasts (CAFs) and, in
many cases, constitute the majority of the cell population of the
tumor stroma (1). CAF includes two
distinct cell types: i) one cell type is similar to fibroblasts and
forms the structural supporting epithelial tissues, and ii) the
other type, myofibroblasts, has biological roles and properties
that are markedly different from those of tissue-derived
fibroblasts. Myofibroblasts are identified by their expression of
α-smooth muscle actin (α-SMA). CAFs have several origins. While
some studies showed that the main progenitor of CAFs appears to be
the local residing fibroblasts (6),
other authors suggest that CAFs originate from the bone marrow and
are derived from mesenchymal stem cells (MSCs) (7–11).
Bone marrow-derived MSCs also comprise a large portion of the tumor
stroma, and can differentiate into a variety of other cell types,
such as fibroblasts and endothelial cells. Irrespective of their
origin, CAFs promote tumor cell growth, invasion and metastasis,
and are therefore well recognized as an important prognostic
indicator in various types of carcinomas (4–13).
Furthermore, CAFs, but not normal fibroblasts, can stimulate
prostatic epithelial proliferation, immortalization and malignant
transformation (14,15), even without apparent mutations in
the epithelial cells themselves (16,17).
Early evidence has indicated that CAFs are different from normal
fibroblasts with regard to their structure, autocrine growth factor
signaling pathway, proliferation and migratory behavior (18–20).
However, genetic alteration of tumor stromal cells (CAFs) under the
influence of the TME, which is known to cause tumor development and
progression, has not received much attention. Although several
studies have demonstrated the association of certain CAF mutations
with carcinomas (21–23), these mutations were only associated
with the initiation of epithelial cell carcinomas (19,24)
and their role in their own malignant transformation remains to be
elucidated. Stromal cells recruited to the TME are termed
'non-tumor' cells and are known to play a role in tumor progression
or suppression. These stromal cells are considered normal and never
undergo malignant transformation (2,25–27).
However, the malignant transformation of the tumor stromal cells,
particularly CAFs and their origin, MSCs, in vitro and in
vivo has been previously reported (28–34).
Therefore, this remains a controversial topic that requires further
investigation.
The fluorescent protein gene tracer technique is
useful for visualization of the TME. The use of transgenic nude
mice with ubiquitous fluorescent protein expression as hosts for
human tumors makes it convenient to observe the interactions
between tumor and host cells (35–38).
Suetsugu et al (39)
successively passaged human tumor specimens in transgenic nude mice
ubiquitously expressing red fluorescent protein (RFP), green
fluorescent protein (GFP) and cyan fluorescent protein (CFP). They
identified that stromal components, such as CAFs, tumor-associated
macrophages (TAMs) and blood vessels expressing RFP, GFP and CFP,
coexist in the transplanted tumor and demonstrated its serial
transplantability (39). The
transplant-ability of tumor-associated stromal cells was also
previously demonstrated by other authors (40). However, the outcome of these stromal
cells is crucial. In the present study, the dual-color fluorescent
protein tracer technique in a mouse model with RFP-expressing tumor
cells and GFP-expressing nude mice was used to observe the
malignant transformation of the host derived GFP-expressing stromal
cells. It was confirmed that these transformed stromal cells were
derived from the bone marrow. Previously, we reported the host
macrophage carcinomas of abdominal xenograft tumors (41). In the present study, we also
reported the malignant transformation of bone marrow-derived
stromal fibroblasts via the transgenic nude mouse model with
ubiquitous GFP expression and BMDCs expressing GFP only.
Materials and methods
Cell culture and transfection with the
RFP gene
The highly invasive glioma stem cell (GSC) line SU3
was previously established in our laboratory (42,43).
SU3 cells were transfected with the RFP gene using a
pLenO-RIP plasmid vector, which carries a puromycin-resistant gene
according to the manufacturer's instructions (Invabio
Biotechnology, Shanghai, China). RFP-transfected SU3 cells
(SU3-RFP) cells were amplified in Dulbecco's modified Eagle's
medium (DMEM) containing 10% fetal bovine serum (FBS) (both from
HyClone, Logan, UT, USA), 100 IU/ml penicillin, 100 µg/ml
streptomycin and 10 µg/ml puromycin to select the transduced
cells. SU3-RFP stem/progenitor cells were cultured in stem cell
medium, i.e., DMEM/F12 (Gibco, Carlsbad, CA, USA), containing 20
ng/ml basic fibroblast growth factor (bFGF; PeproTech, Rocky Hill,
NJ, USA), 20 ng/ml epidermal growth factor (EGF; Invitrogen Life
Technologies, Carlsbad, CA, USA), B27 supplement (50X), 2 mmol/l
L-glutamine (100X), MEM vitamin solution (100X) and 100 mM sodium
pyruvate (100X) (all from Gibco). Immunohistochemical staining with
monoclonal antibodies against CD133 (Miltenyi, Bergisch Gladbach,
Germany), nestin (BD Biosciences, San Jose, CA, USA), GFAP (Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA) and β-tubulin-III
(BD Biosciences) was performed to detect the expression of the
markers and determine the cell type of the differentiated
cells.
Transgenic GFP nude and chimeric
mice
Transgenic C57BL/6-GFP and BALB/c athymic nude
(nu/nu) mice were purchased from the Model Animal Center of Nanjing
University (Nanjing, China). C57BL/6-GFP mice expressed GFP under
the control of the CAG promoter (CMV-IE enhancer chicken β-actin
promoter and rabbit β-globin genomic DNA) (36,44).
Findings of a previous study (36)
revealed that GFP nude mice were generated by crossing BALB/c nude
mice with C57BL/6-GFP mice. Continuous back-crossing for 10
generations was then performed to purify the background to BALB/c.
To establish chimeric mice, the bone marrow of BALB/c athymic nude
(nu/nu) mice was subjected to 6 Gy of radiation to destroy the bone
marrow, which was then rebuilt from GFP nude mice. Only bone
marrow-derived cells expressed GFP in this chimeric mouse model.
The animals were maintained under the specific pathogen-free
environment at the Animal Center of Soochow University. The animal
studies were conducted in accordance with the approved facilities
of the Chinese Experimental Animal.
Establishment of the xenograft tumor and
subculturing
Six-week-old GFP nude mice received an injection of
1×106 SU3-RFP cells in a total volume of 50 µl in
the armpit of the right forelimb via a Hamilton syringe. Whole-body
images of the tumor-bearing mice were obtained using the In Vivo
Imaging System FX Pro (Kodak, Rochester, NY, USA) under excitation
at 470 and 535 nm after induction of anesthesia. The mice were then
sacrificed and a section of the transplanted tumor tissues was
fixed with 4% paraformaldehyde, dehydrated in 20% and then 30%
sugar, and used to make 5-µm frozen sections. The sections
were stained with hematoxylin and eosin (H&E) or observed
directly under a fluorescence microscope after staining with
4′,6-diamino-2-phenylindole (DAPI; KeyGen, Nanjing, China).
Confocal laser-scanning microscopy was performed to monitor the
mutual interactions between tumor and host cells. The other section
of the tissues was harvested and washed with phosphate-buffered
saline (PBS) containing penicillin and streptomycin, three times.
The section was then minced with fine scissors into small
fragments, and cultured in culture medium containing 10% FBS. The
medium was replaced with fresh culture medium the following day.
Normal cell culture conditions were maintained for the subsequent
days.
Identification and cloning of the
GFP-positive cells
After several passages, the GFP-positive cells
multiplied persistently until a stable percentage of the total
cells was reached. Adherent cells were digested in a single-cell
suspension using the Accutase Cell Dissociation Reagent (Gibco),
and sorted using a cell sorter (MoFlo XDP; Beckman Coulter, USA)
into three or four groups: cells emitting red and green
fluorescence, both red and green fluorescence, and/or cells without
fluorescence. Cells emitting green fluorescence were collected and
cultured in fresh medium containing 10% FBS. A single cell was
obtained using a capillary pipette and cultured in 96-well
plates.
Growth characteristics of the
GFP-positive cells
Proliferation assay
Cell proliferation was evaluated using soft agar
colony assay and Cell Counting Kit-8 (CCK-8; Dojindo Laboratories,
Mashikimachi, Kamimashiki Gun, Kumamoto, Japan) according to the
manufacturer's instructions. For the CCK-8 assay, the cells were
seeded at 1×103 cells/well in 96-well plates and
cultured in 100 µl medium. SU3-RFP, NIH-3T3 and blank
control (only medium and CCK-8 solution) were set at the same time.
CCK-8 solution (10 µl/well) was added and then incubated at
37°C for 2 h. The optical density (OD) at 450 nm was recorded by a
microplate teader (Tecan Infinite 200 PRO; Salzburg, Austria). Each
experiment was performed three times in 6-well plates.
Cell cycle assay
For the cell cycle assay, the cells were harvested
in the logarithmic phase, washed with ice-cold PBS, and fixed with
70% ethanol and stored at −20°C for at least 24 h. Prior to
analysis, the fixed cells were centrifuged for 10 min at 2,000 g
and resuspended in PBS incubated with 0.5 mg/ml RNase A (Sigma
R-4875; St. Louis, MO, USA) and 25 µg/ml propidium iodide
(PI; KeyGen Biotech, Nanjing, China) for 30 min at room temperature
in the dark. SU3-RFP and mouse peripheral blood lymphocytes (LYM)
were used as the control. The cells were counted using flow
cytometry (Cytomics FC 500; Beckman Coulter) and analyzed using CXP
software (Beckman Coulter).
Soft agar colony assay
For the soft agar colony assay, the cells
(1×103/well) were suspended in 2X DMEM containing 20%
FBS, mixed with the same volume of 0.7% low-melting agarose (Lonza,
Allendale, NJ, USA), and seeded over a layer of 0.5% agar in a
6-well plate. After 15 days of incubation, colonies with >20
cells were counted and photographed using an inverted microscope
(Olympus CKX31; Tokyo, Japan).
Genetic characteristics of the
GFP-positive cells
RT-PCR, karyotype analysis and DNA
content
GFP expression of the transformed cells was detected
using PCR as previously described (41). GFP transgenic mouse tail was used as
the positive control and the mouse NIH-3T3 cell line was used as
the negative control, and specific human and mouse primer pairs
were used to identify the cell species. Primers of the GFP
gene were: 5′-GCCACAAGTTCAGCGTGTCCG and 5′-GTTGGGGTCTTTGCTCAGGGCG
(566 bp); RFP, 5′-AGGTTCTTAGCGGGTTTCTTG and 5′-CTTCCCTGA
GGGCTTCACAT (312 bp); human-specific β-action, 5′-ACA
TCCGCAAAGACCTATAC and 5′-GCCATGCCAATCTCA TCTTG (346 bp);
mouse-specific β-action, 5′-CTTTGCAGC TCCTTCGTTG and
5′-TGGTAACAATGCCATGTTCA (278 bp). Karyotyping of these cells was
performed as previously described (45). Cell DNA content and the cell cycle
were analyzed using flow cytometry. For the DNA content assay, the
cell density of the sample and control cells after staining with PI
were adjusted until they were the same and sample flow was strictly
controlled. Proper maintenance and careful adjustment of the
instrument were ensured prior to analysis. The mouse blood
lymphocytes from the same strain were extracted as the control
cells.
Immunofluorescence analysis
The immunofluorescence analysis was performed as
previously described (42).
Briefly, frozen sections were produced as described earlier. Tissue
sections were stained with antibodies against α-SMA (ab5694, 1:100;
Abcam, Cambridge, USA), S100A4 (ab27957, 1:100), FAP-α (ab53066),
CD44 (ab25340, 1:200), CD105 (ab107595) and Ki67 (ab16667, ab15580,
1:50). Cy3-labeled goat anti-rabbit IgG and Alexa Fluor 488-labeled
goat anti-rabbit IgG (Beyotime, Shanghai, China) were used as
secondary anti-bodies. Expression of the aforementioned cell
surface markers was detected with a fluorescence microscope
(Olympus IX51; Tokyo, Japan), and images were captured and merged
using the software provided.
Tumorigenicity in nude mice
To verify the tumorigenicity of the GFP-positive
cells, BALB/c nude mice (n=10) and normal mice (n=10) aged 5 weeks,
were injected with 1×107 and 1×106
GFP-positive cells in the armpit of the right forelimb,
respec-tively. Tumor-bearing mice were sacrificed 25 days later,
and the transplanted tumor tissues were sectioned and stained with
H&E and DAPI. Images were captured and merged using a
phase-contrast and a fluorescence microscope.
Statistical analysis
Each experiment was performed as least three times,
and data are presented as the mean ± SD where applicable.
Differences were evaluated using the Student's t-tests for
two-group comparisons. P<0.05 was considered to indicate a
statistically significant result. Each group was analyzed using the
GraphPad Prism 5.0 software.
Results
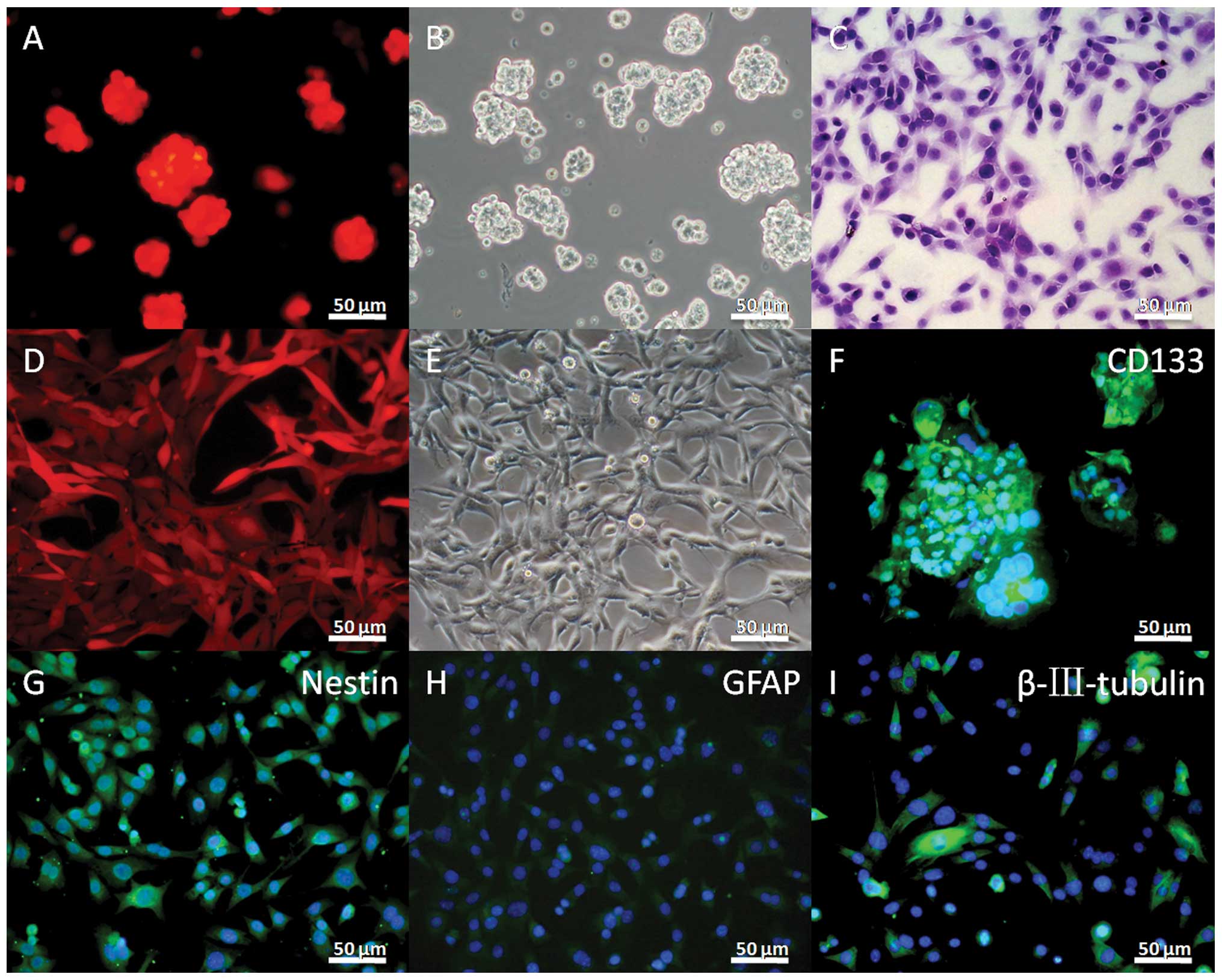
RFP-expressing SU3 cell line
The SU3-RFP showed almost 100% expression of RFP
(Fig. 1A and D). The in
vitro growth characteristics of the stem and adherent cells
were similar to those of the SU3 cells without the RFP vector
(Fig. 1B, C and E). As with the SU3
cell lines previously reported (43), the SU3-RFP cells also showed a high
expression of CD133 and nestin in the stem cells when cultured in
stem cell medium (Fig. 1F and G).
GFAP- and β-tubulin III-positive cells were observed with 10% FBS
supplementation (Fig. 1H and
I).
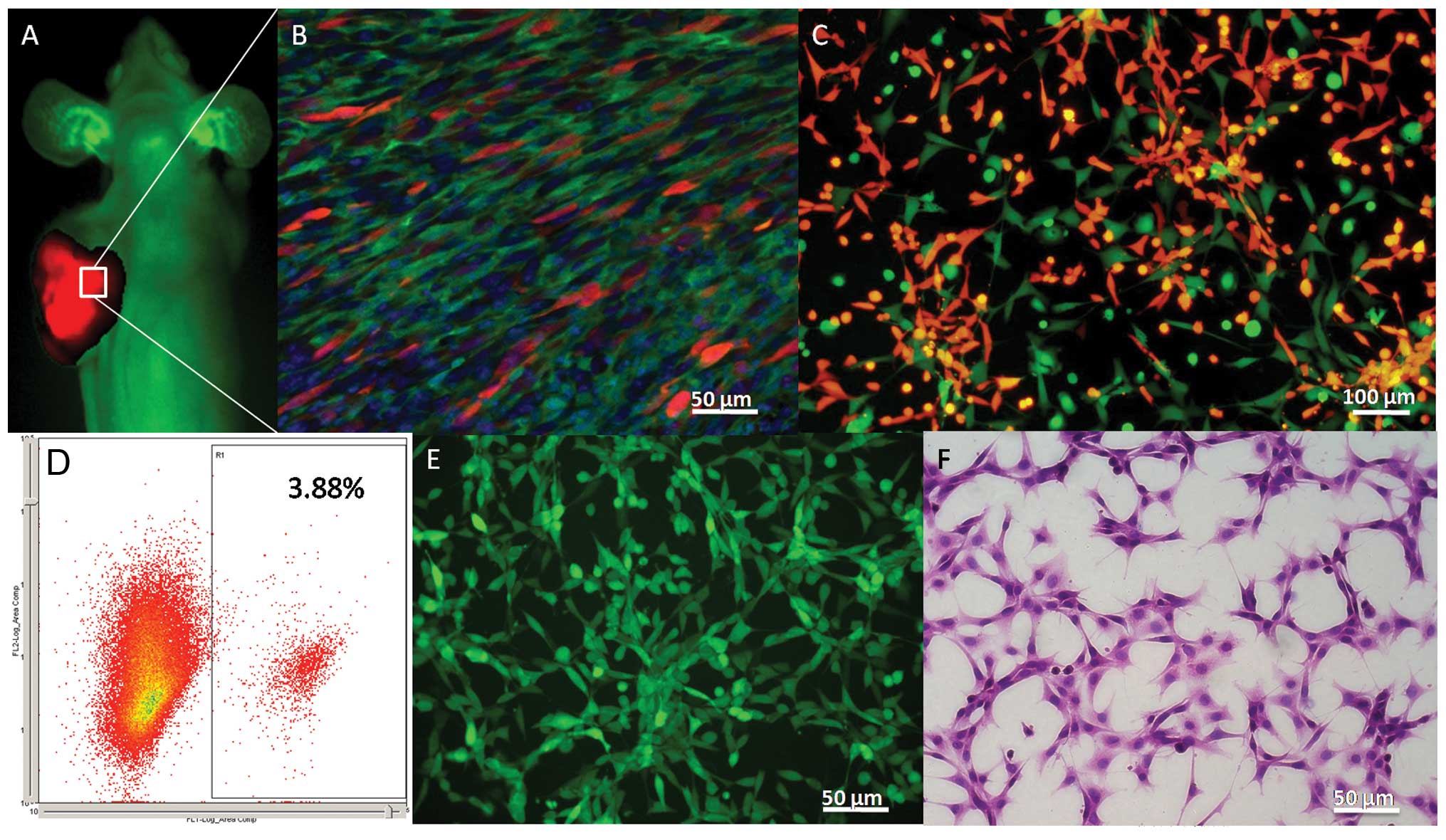
GFP-positive host stroma cells
As previously reported (44), transgenic EGFP nude mice
ubiquitously expressed green fluorescence, except for the hair and
red blood cells under blue light (Fig.
2A). The transplanted tumor showed bright red fluorescence
under excitation with green light (Fig.
2A). On the frozen DAPI-stained sections, GFP-positive host
stromal cells showed marked proliferation and accounted for a large
proportion of the total cells (Fig.
2B). After subculturing, the RFP-positive SU3RFP and
GFP-positive host stromal cells were easily distinguished under the
fluorescent microscope (Fig. 2C),
and the latter accounted for 3.88% of the total cells in the flow
cytometric analysis (Fig. 2D), less
than the proportion of transplanted tumor tissues. GFP-positive
host stromal cells were sorted by flow cytometry and termed
SU3-RFP-induced host subcutaneous transformed cells (ihSTCs), which
were maintained in continual cultures, similar to immortalized
cells (Fig. 2E). These
spindle-shaped cells, which showed strong nuclear staining and had
an unlimited capacity for proliferation, had similar
characteristics to malignant tumor cells (Fig. 2F).
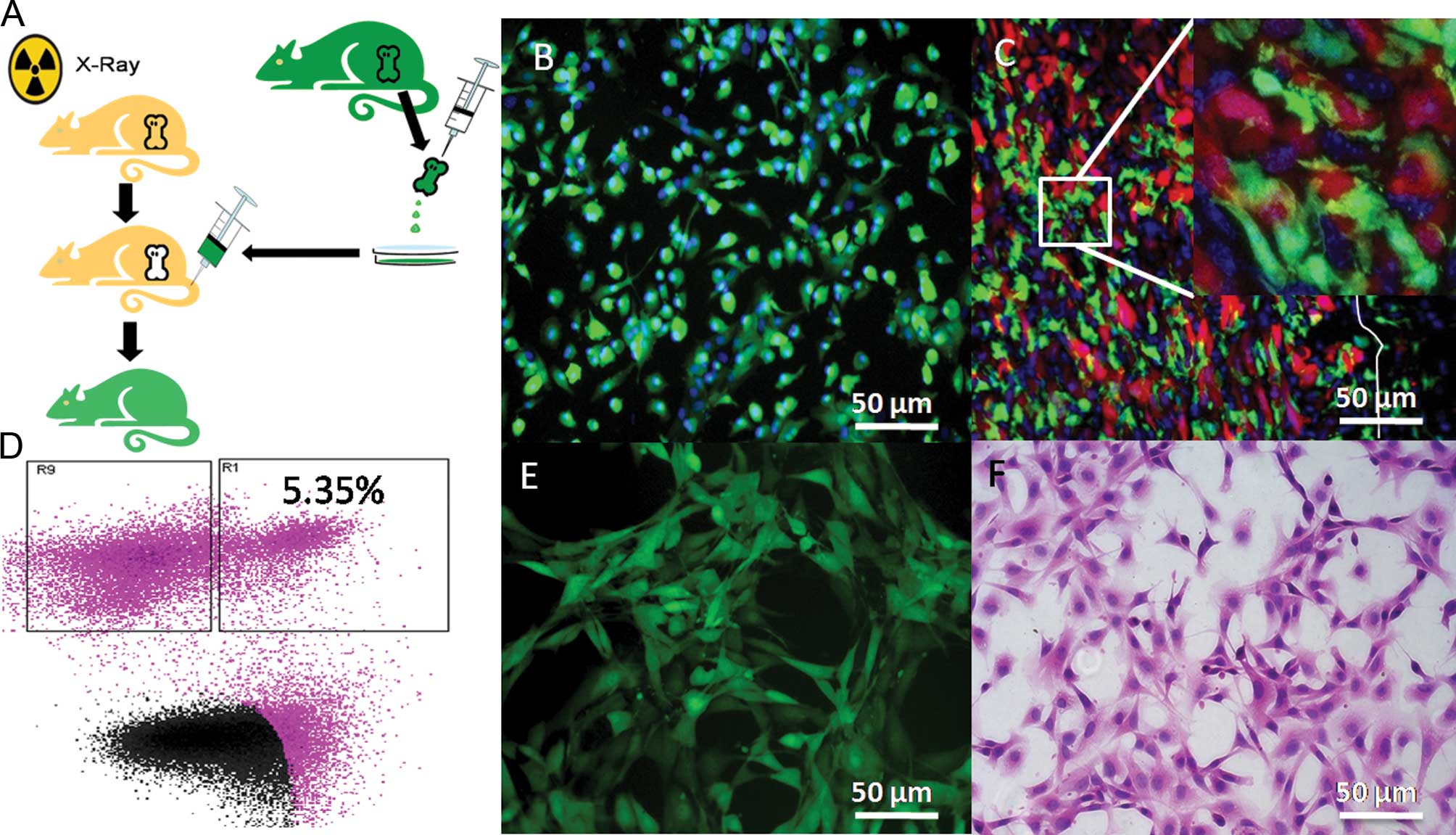
In the chimeric mice, the bone marrow system was
rebuilt (Fig. 3A), with almost all
the bone marrow-derived cells expressing GFP (Fig. 3B). In the xenograft tumors,
interactions between SU3-RFP cells and bone marrow-derived
GFP-positive cells were clearly observed under a confocal
laser-scanning microscope (Fig.
3C). The GFP-positive cells were sorted by flow cytometry and
accounted for 5.35% of the total cells (Fig. 3D), with their growth characteristics
being similar to ihSTCs in vitro (Fig. 3E and F). The cells were termed
SU3-RFP-induced host subcutaneous transformed, bone marrow-derived
cells (ihSTBMCs).
Growth characteristics of ihSTCs and
ihSTBMCs
In the primary cultured cells, RFP-expressing tumors
cells, GFP-expressing host cells, and the small number of GFP-RFP
co-expressing cells were easily distinguishable. After short-term
subculturing, the cells were digested and sorted using a flow
cytometer. The sorted GFP-expressing cells were spindle-like,
polygonal or squamous and showed loss of cell contact inhibition
under the fluorescence and phase-contrast microscope with H&E
staining (Figs. 2F and 3F).
The CCK-8 assay showed that ihSTCs and ihSTBMCs grew
rapidly, faster than the SU3-RFP and NIH-3T3 cells (Fig. 4A). This result confirmed that the
proliferative abilities of the ihSTCs and ihSTBMCs were higher than
those of the original tumor cells.
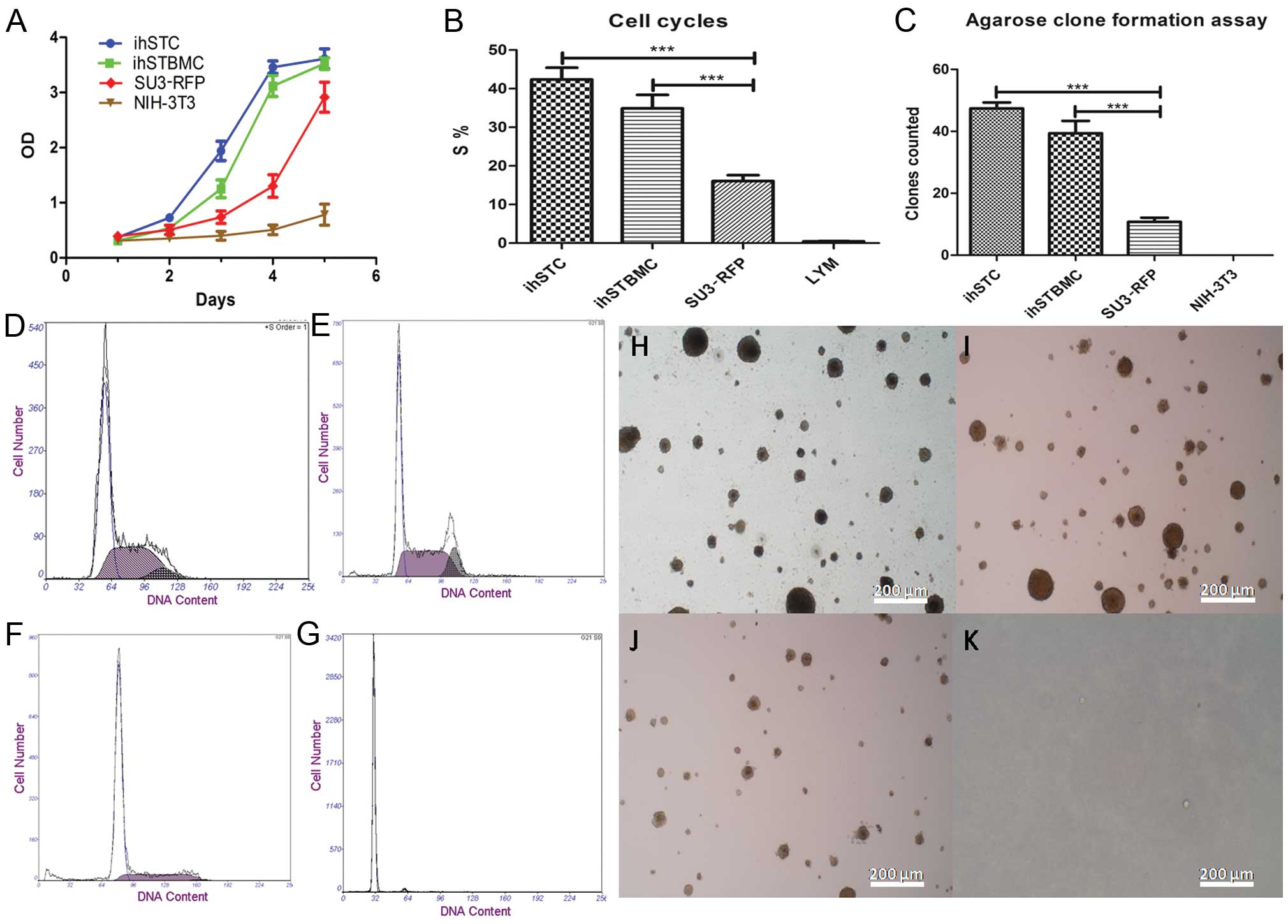 | Figure 4In vitro growth
characteristics of ihSTCs and ihSTBMCs. (A) Proliferation curve of
ihSTCs and ihSTBMCs, SU3-RFP cells and mouse embryonic fibroblast
cell NIH-3T3 (control). (B and D–G) Cell cycles of ihSTCs and
ihSTBMCs, SU3-RFP and NIH-3T3 cells (control); the S-phase cell
fractions of ihSTCs and ihSTBMCs, SU-3RFP and NIH-3T3 cells are
42.33% (D), 34.91% (E), 16.06% (F) and 0.44% (G), respectively. (C
and H–K) According to the double-layer agarose clone formation test
for cell proliferation ability, the colony-forming efficiency of
ihSTCs and ihSTBMCs, SU3-RFP and NIH-3T3 cells was 47.37‰ (H),
39.38‰ (I), 10.75‰ (J) and 0‰ (K), respectively. (H–K) Scale bar,
50 µm. |
Results of the cell cycle assay showed that a
markedly higher number of ihSTCs and ihSTBMCs were in the S phase
(Fig. 4B): 42.33% (Fig. 4D) and 34.91% (Fig. 4E), respectively, in the S phase,
higher than the number of SU3-RFP (16.06%, Fig. 4F) and LYM (0.44%, Fig. 4G) cells. Thus, ihSTCs and ihSTBMCs
showed an abnormally high DNA synthesis capacity, much higher than
that of the SU3-RFP cells (P<0.0001).
The agar colony assay showed that the colony
formation of ihSTCs (47.37‰) and ihSTBMCs (39.38‰) was
significantly enhanced compared to the SU3RFP (10.75‰) and NIH3T3
(0‰) cells (Fig. 4C and H–K)
(P<0.0001).
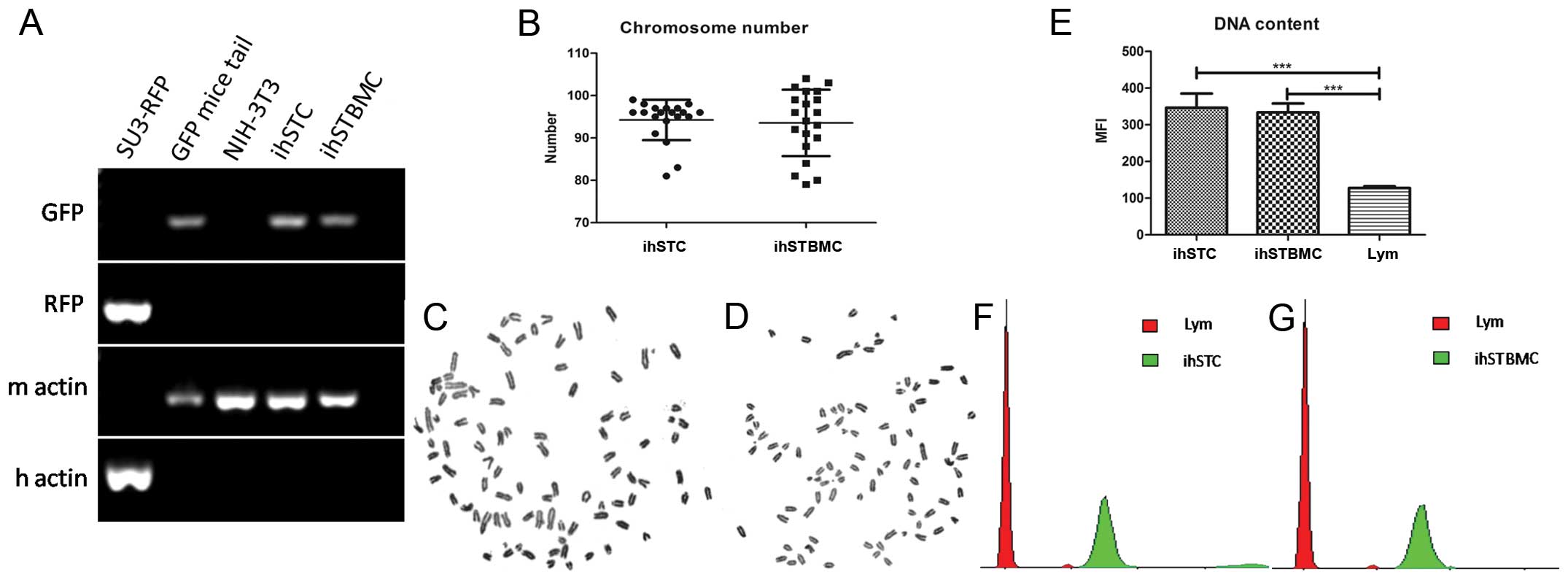
Murine origin of ihSTCs and ihSTBMCs
Under the fluore-scent microscope, ihSTCs and
ihSTBMCs emitted green fluorescence, which proved that the cell
lines were host-derived (Figs. 2E
and 3E). The origin of the two cell
lines was also identified by mRNA analysis using mouse- and
human-specific primers. Mouse β-actin was amplified but human
β-actin was not amplified in ihSTCs and ihSTBMCs (Fig. 5A). Moreover, the GFP gene was
expressed in ihSTCs and ihSTBMCs, as well as the control GFP mouse
tail, but not in the SU3-RFP or NIH-3T3 cells (Fig. 5A). Karyotype analysis also proved
that the chromosomes of the two cell lines were telo centric
(Fig. 5C and D), which is
characteristic of all mouse chromosomes. Additionally, the average
number of chromosomes of ihSTCs and ihSTBMCs was 94.25±4.66 (n=20)
and 93.55±7.65 (n=20), respectively (Fig. 5B–D), which shows the pentaploid
characteristics of these cells. The DNA content assay further
validated the results of the karyo-type analysis, showing that the
mean fluorescence intensity (MFI) of ihSTCs and ihSTBMCs was 2.71-
and 2.61-fold (Fig. 5E and F, green
peaks) that of the mouse lymphocytes, respectively (Fig. 5E and F, red peaks).
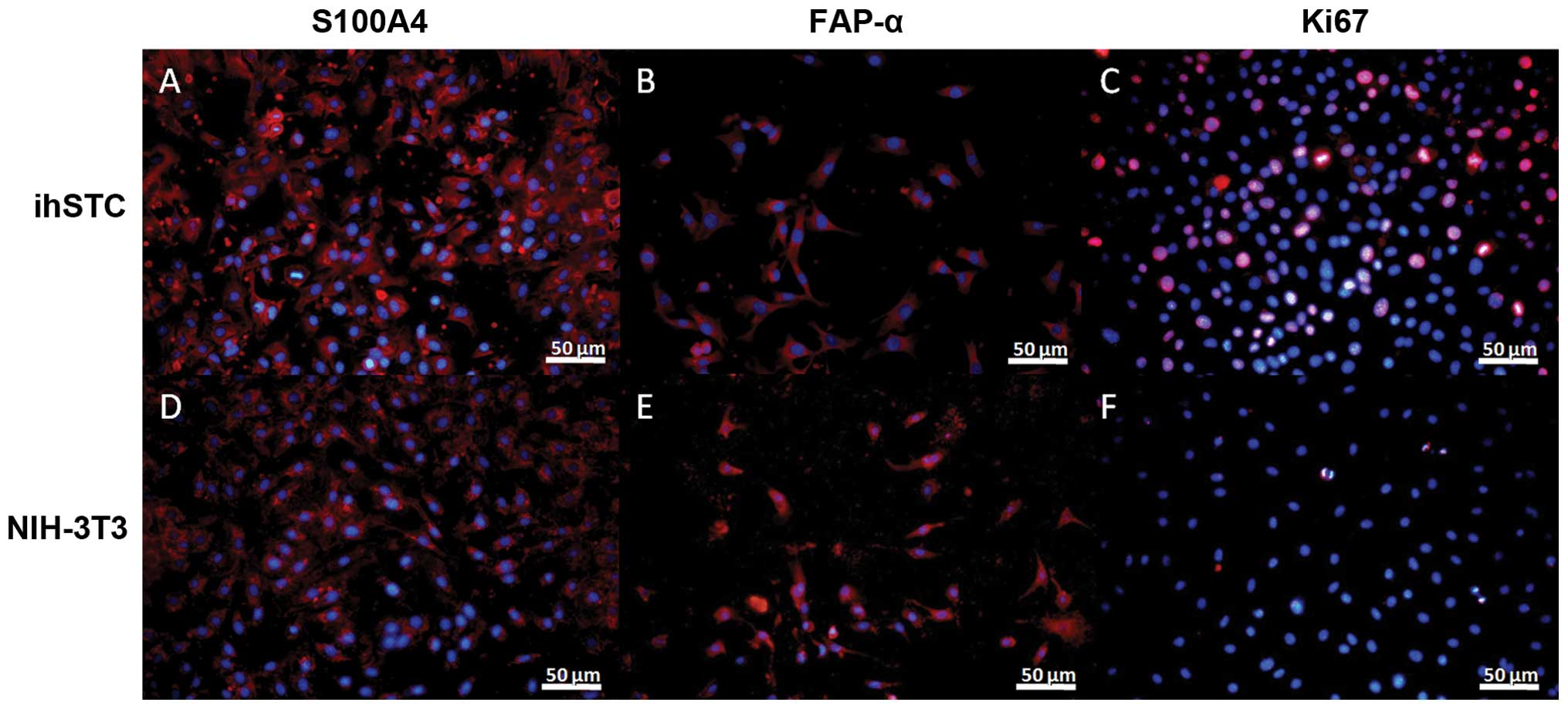
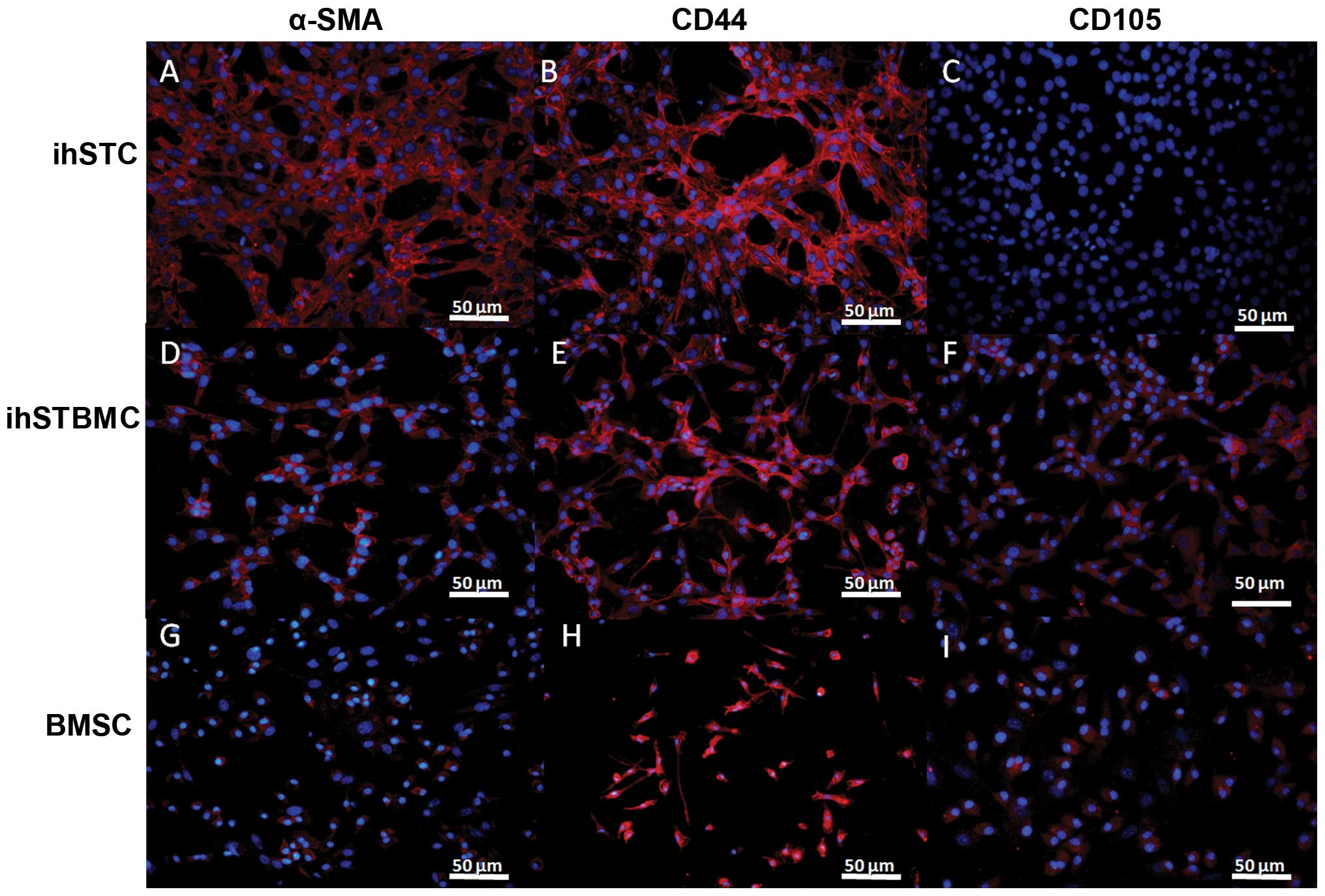
The immunofluorescence assay showed that ihSTCs were
not only positive for S100A4 (Fig.
6A), FAP-α (Fig. 6B) and α-SMA
(Fig. 7A), but also for CD44
(Fig. 7B). Additinoally, ihSTBMCs
were positive for CD44 and CD105, as well α-SMA. However, the two
cell types were negative for the glioma stem cell markers CD133,
nestin, GFAP and β-tubulin III (data not shown). Ki-67 was used as
a marker of cell proliferation. S100A4, FAP-α and α-SMA are known
as biomarkers of activated fibroblasts, and CD44 and CD105 are
common mesenchymal stromal cell markers. Based on these findings
and the GFP expression in these cells, ihSTCs and ihSTBMCs appear
to be the source of rapidly proliferating host-derived fibroblasts
and mesenchymal cells, respectively. Furthermore, transformed
fibroblasts may originate from bone marrow-derived mesenchymal stem
cells.
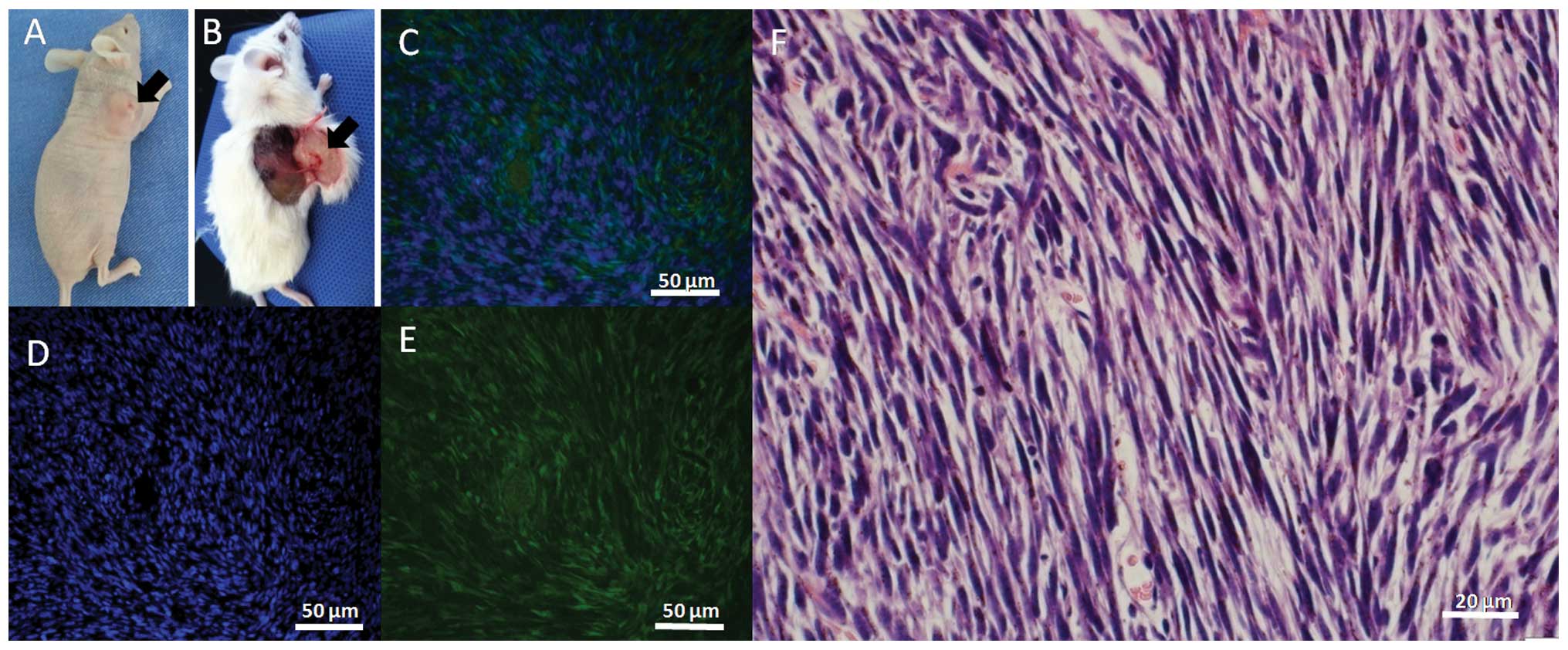
Enhanced tumorigenicity in normal and
immunosuppressed mice
The two GFP-positive cell lines established showed
100% tumorigenicity (n=10) in nude mice (Fig. 8A), 100% tumorigenicity in normal
mice (5/5) at a dose of 1×107 cells/mice and 40%
tumorigenicity (2/5) at a dose of 1×106 GFP-positive
cells/mouse (Fig. 8B). However, the
tumorigenicity of SU3-RFP was only 20% (1/5) in normal mice at a
dose of 1×107 cells and 0% (0/5) at a dose of
1×106 cells. These results confirmed that the
tumorigenicity of the two GFP-positive cell lines derived from the
host mice was stronger than that of the human glioma stem cell line
SU3-RFP. Frozen DAPI-stained sections showed that the transplanted
tumor tissue was positive for GFP (Fig.
8C–E) and showed fibrosarcoma-like changes with H&E
staining (Fig. 8F).
Discussion
The use of fluorescent proteins for imaging the TME
by color-coded cancer and stromal cells is highly useful (46), as it provides for non-invasive,
dynamic and real-time visualization. We, as well as other
investigators, have cultivated GFP athymic nude mice by crossing
C57BL/6J-GFP mice with BALB/c nude mice (35–37).
The transgenic GFP nude mice are crucial for studying tumor-host
interactions. Suetsugu et al ((47) demonstrated that GFP-expressing host
CAFs are recruited by the host tumor and proliferate, based on
fluorescence imaging and suggested the important role of these
cells in tumor progression. Previous findings have also shown that
CAFs within tumors are derived from bone marrow-derived mesenchymal
stem cells (8,48). To determine the role of BMDCS, in
the present study, we established a chimeric mouse model in which
only the BMDCs expressed GFP. First, we destroyed the bone marrow
system of BALB/c nude mice, and then we replaced it with whole bone
marrow cells of GFP nude mice via tail vein injection. The BMDCs of
these chimeric mice eventually expressed GFP. In the frozen tumor
sections, a large number of bone marrow-derived GFP-expressing
cells were found to extensively accumulate in the tumor area.
However, almost none of the GFP cells accumulated in the adjacent
non-tumor area in nude mice. Therefore, this mouse model seems
ideal for studying the interactions between tumor cells and
BMDCs.
Using this transgenic GFP nude mouse model, in the
present study, we cloned a host-derived cell line expressing GFP
from the subcutaneous transplantation tumor, and we demonstrated
that it was a fibroblast cell line positive for the characteristic
biomarkers. The application of the chimeric model also proved that
some of the tumor stroma fibroblasts, if not all, were recruited
from the bone marrow. To confirm the murine origin of the
GFP-expressing cells, mouse-specific mRNA level and karyotype
analyses were performed. Only mouse-specific β-actin was amplified
and telocentric chromosomes were observed. Based on this evidence,
it appears that the two GFP cell lines were derived from the host
mouse. De novo expression of α-SMA is the most commonly used
marker for CAFs, while other widely used CAF markers are
fibroblast-specific protein 1 (FSP1; or S100A4) and fibroblast
activation protein (FAP). However, due to their heterogeneity, not
all markers can be identified on CAFs. In the present study, we
identified all the CAF markers (α-SMA, S100A4 and FAP-α) on ihSTCs,
but only α-SMA expression was identified in ihSTBMCs. However, the
markers of mesenchymal cells, CD44 and CD105, were shown to be
expressed on ihSTBMCs. Considering that ihSTBMCs are bone
marrow-derived cells, this is reliable evidence to prove that tumor
stroma fibroblasts originate from the bone marrow and are derived
from MSCs.
The malignant potential of the two cell lines were
then demonstrated in vitro and in vivo. The CCK-8
assay did not show any significant differences in the proliferation
rates of ihSTCs and ihSTBMCs. However, the proliferation rates of
the two cell lines were higher than those of SU3-RFP and NIH-3T3.
Flow cytometric analysis of the cell cycle assay showed a marked
increase in the number of ihSTCs and ihSTBMCs in the S phase, and
this corresponded with the results of the CCK-8 assay.
Anchorage-independent growth, which is generally recognized as a
hallmark of oncogenic transformation, was examined. The
colony-forming efficiency of ihSTCs and ihSTBMCs was higher than
that of SU3-RFP and NIH-3T3 cells. The most useful evidence of
malignant trans-formation is the tumorigenicity of the two cell
lines in vivo. As shown in the present results, when ihSTCs
and ihSTBMCs were injected into athymic nude mice, a 100%
tumorigenesis rate (20/20) was observed with even as few as
1×106 cells (10/10). Furthermore, ihSTCs and ihSTBMCs
showed stronger tumorigenicity than SU3-RFP cells in normal BALB/c
mice. The morphological features of fusocellular sarcomas were
evident in the H&E-stained sections. This finding is further
evidence of its fibroblast-like phenotype.
Stromal cells within a tumor can be truly malignant
and have been reported as early as 1981 (28), although it remains under
investigation (30–34,49–51).
Since it is almost impossible to distinguish between the tumor and
stroma, in vitro studies involving direct or indirect
co-culturing have been performed. Using the fluorescent tracing
mouse model, we have demonstrated that stroma fibroblasts undergo
malignant transformation, although whether this process occurred
in vitro or in vivo remains to be determined, since
the process of tumor growth in vivo occurs in approximately
30 days and the transplanted tumors were subcultured in
vitro for approximately one week for sorting. As known,
transplanted tumors in animals have a more complicated
microenvironment. Additionally, the malignant transformation of
MSCs and mouse embryonic fibroblasts was not observed with direct
continual culture or co-culture with SU3-RFP via Transwell for over
2 months until apoptosis in vitro (data not shown).
Therefore, we speculated that the process of malignant
transformation was initiated in vivo. In addition, to prove
that the viral vector containing the RFP gene transfected
into SU3 cells did not exert any effect, SU3 cells that did not
contain the RFP gene were transplanted into GFP nude mice,
which yielded GFP-positive cells, similar to the results for
SU3-RFP (data not shown).
The mechanisms involved in the transformation,
however, are unclear and remain to be investigated. Goldenberg
et al, one of the first investigators to study the malignant
transformation of stromal cells, recently suggested that cell
fusion of tumor and stromal cells is involved in malignancies
(52). Furthermore, recent findings
suggested that certain functional human genes may be involved in
the malignant transformation of stromal cells (49,50).
This view is also supported by other investigators (53–55).
However, as shown above, two of the transformed cells cloned in the
present study are of mouse origin, and only mouse chromosomes were
observed. However, as reported by Pathak et al (53) and Jacobsen et al (54), it is possible that the tumor cells
first fused with the stroma cells, led to loss of the tumor cell
chromosomes. It has been argued that stimulation with cytokines
such as IL-6, GM-CSF and IL-4 is a potential mechanism underlying
the malignant transformation of stroma cells (32,56).
However, all the experimental results are derived from in
vitro co-culture systems, which may not completely represent
the in vivo TME. Recent findings suggested that exosomes and
the microRNAs secreted by breast cancer cells induce
non-tumorigenic epithelial cells to form tumors (5). Microvesicles of cancer cells (gliomas)
can contribute to the horizontal propagation of oncogenes, such as
EGFRvIII, as previously reported (57), but whether these oncogenes can be
passed on from cancer to stromal cells remains unknown.
In conclusion, in the present study, the bone
marrow-derived tumor stromal cells ihSTCs and ihSTBMCs were found
to be of murine MSC origin and showed a rapid growth rate, high
cloning efficiency, high DNA content, expression of fibroblasts
markers and high tumorigenicity. Taken together, these results
demonstrate that the two cell lines are malignant transformed
fibroblasts that originated from the bone marrow and were recruited
by mesenchymal cells.
Acknowledgments
The present study was funded by the National Natural
Scientific Foundation of China (nos. 81172400, 81272799, 81302196,
81302180 and 81472739).
References
|
1
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tarin D: Role of the host stroma in cancer
and its therapeutic significance. Cancer Metastasis Rev.
32:553–566. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Casazza A, Di Conza G, Wenes M,
Finisguerra V, Deschoemaeker S and Mazzone M: Tumor stroma: A
complexity dictated by the hypoxic tumor microenvironment.
Oncogene. 33:1743–1754. 2014. View Article : Google Scholar
|
|
4
|
De Wever O, Demetter P, Mareel M and
Bracke M: Stromal myofibroblasts are drivers of invasive cancer
growth. Int J Cancer. 123:2229–2238. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Melo SA, Sugimoto H, O'Connell JT, Kato N,
Villanueva A, Vidal A, Qiu L, Vitkin E, Perelman LT, Melo CA, et
al: Cancer exosomes perform cell-independent microRNA biogenesis
and promote tumorigenesis. Cancer Cell. 26:707–721. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Räsänen K and Vaheri A: Activation of
fibroblasts in cancer stroma. Exp Cell Res. 316:2713–2722. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Worthley DL, Ruszkiewicz A, Davies R,
Moore S, Nivison-Smith I, Bik To L, Browett P, Western R, Durrant
S, So J, et al: Human gastrointestinal neoplasia-associated
myofibroblasts can develop from BMDCS following allogeneic stem
cell transplantation. Stem Cells. 27:1463–1468. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Quante M, Tu SP, Tomita H, Gonda T, Wang
SS, Takashi S, Baik GH, Shibata W, Diprete B, Betz KS, et al: Bone
marrow-derived myofibroblasts contribute to the mesenchymal stem
cell niche and promote tumor growth. Cancer Cell. 19:257–272. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bergfeld SA and DeClerck YA: Bone
marrow-derived mesenchymal stem cells and the tumor
microenvironment. Cancer Metastasis Rev. 29:249–261. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hanahan D and Coussens LM: Accessories to
the crime: Functions of cells recruited to the tumor
microenvironment. Cancer Cell. 21:309–322. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mishra PJ, Mishra PJ, Humeniuk R, Medina
DJ, Alexe G, Mesirov JP, Ganesan S, Glod JW and Banerjee D:
Carcinoma-associated fibroblast-like differentiation of human
mesenchymal stem cells. Cancer Res. 68:4331–4339. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kalluri R and Zeisberg M: Fibroblasts in
cancer. Nat Rev Cancer. 6:392–401. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ostman A and Augsten M: Cancer-associated
fibroblasts and tumor growth - bystanders turning into key players.
Curr Opin Genet Dev. 19:67–73. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hayward SW, Wang Y, Cao M, Hom YK, Zhang
B, Grossfeld GD, Sudilovsky D and Cunha GR: Malignant
transformation in a nontumorigenic human prostatic epithelial cell
line. Cancer Res. 61:8135–8142. 2001.PubMed/NCBI
|
|
15
|
Olumi AF, Grossfeld GD, Hayward SW,
Carroll PR, Tlsty TD and Cunha GR: Carcinoma-associated fibroblasts
direct tumor progression of initiated human prostatic epithelium.
Cancer Res. 59:5002–5011. 1999.PubMed/NCBI
|
|
16
|
Bhowmick NA, Chytil A, Plieth D, Gorska
AE, Dumont N, Shappell S, Washington MK, Neilson EG and Moses HL:
TGF-beta signaling in fibroblasts modulates the oncogenic potential
of adjacent epithelia. Science. 303:848–851. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kuperwasser C, Chavarria T, Wu M, Magrane
G, Gray JW, Carey L, Richardson A and Weinberg RA: Reconstruction
of functionally normal and malignant human breast tissues in mice.
Proc Natl Acad Sci USA. 101:4966–4971. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bhowmick NA, Neilson EG and Moses HL:
Stromal fibroblasts in cancer initiation and progression. Nature.
432:332–337. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Russell PJ, Bennett S and Stricker P:
Growth factor involvement in progression of prostate cancer. Clin
Chem. 44:705–723. 1998.PubMed/NCBI
|
|
20
|
Tlsty TD and Hein PW: Know thy neighbor:
Stromal cells can contribute oncogenic signals. Curr Opin Genet
Dev. 11:54–59. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hu M, Yao J, Cai L, Bachman KE, van den
Brûle F, Velculescu V and Polyak K: Distinct epigenetic changes in
the stromal cells of breast cancers. Nat Genet. 37:899–905. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Paterson RF, Ulbright TM, MacLennan GT,
Zhang S, Pan CX, Sweeney CJ, Moore CR, Foster RS, Koch MO, Eble JN,
et al: Molecular genetic alterations in the
laser-capture-microdissected stroma adjacent to bladder carcinoma.
Cancer. 98:1830–1836. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Moinfar F, Man YG, Arnould L, Bratthauer
GL, Ratschek M and Tavassoli FA: Concurrent and independent genetic
alterations in the stromal and epithelial cells of mammary
carcinoma: Implications for tumorigenesis. Cancer Res.
60:2562–2566. 2000.PubMed/NCBI
|
|
24
|
Houghton J, Li H, Fan X, Liu Y, Liu JH,
Rao VP, Poutahidis T, Taylor CL, Jackson EA, Hewes C, et al:
Mutations in bone marrow-derived stromal stem cells unmask latent
malignancy. Stem Cells Dev. 19:1153–1166. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bernardo ME, Zaffaroni N, Novara F, Cometa
AM, Avanzini MA, Moretta A, Montagna D, Maccario R, Villa R,
Daidone MG, et al: Human bone marrow derived mesenchymal stem cells
do not undergo transformation after long-term in vitro culture and
do not exhibit telomere maintenance mechanisms. Cancer Res.
67:9142–9149. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gou S, Wang C, Liu T, Wu H, Xiong J, Zhou
F and Zhao G: Spontaneous differentiation of murine bone
marrow-derived mesenchymal stem cells into adipocytes without
malignant transformation after long-term culture. Cells Tissues
Organs. 191:185–192. 2010. View Article : Google Scholar
|
|
27
|
Luetzkendorf J, Nerger K, Hering J, Moegel
A, Hoffmann K, Hoefers C, Mueller-Tidow C and Mueller LP:
Cryopreservation does not alter main characteristics of Good
Manufacturing Process-grade human multipotent mesenchymal stromal
cells including immunomodulating potential and lack of malignant
transformation. Cytotherapy. 17:186–198. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Goldenberg DM and Pavia RA: Malignant
potential of murine stromal cells after transplantation of human
tumors into nude mice. Science. 212:65–67. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sparrow S, Jones M, Billington S and Stace
B: The in vivo malignant transformation of mouse fibroblasts in the
presence of human tumour xenografts. Br J Cancer. 53:793–797. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Røsland GV, Svendsen A, Torsvik A, Sobala
E, McCormack E, Immervoll H, Mysliwietz J, Tonn JC, Goldbrunner R,
Lønning PE, et al: Long-term cultures of bone marrow-derived human
mesenchymal stem cells frequently undergo spontaneous malignant
transformation. Cancer Res. 69:5331–5339. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu J, Zhang Y, Bai L, Cui X and Zhu J:
Rat bone marrow mesenchymal stem cells undergo malignant
transformation via indirect co-cultured with tumour cells. Cell
Biochem Funct. 30:650–656. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cui X, Liu J, Bai L, Tian J and Zhu J:
Interleukin-6 induces malignant transformation of rat mesenchymal
stem cells in association with enhanced signaling of signal
transducer and activator of transcription 3. Cancer Sci. 105:64–71.
2014. View Article : Google Scholar
|
|
33
|
Serrano-Heras G, Domínguez-Berzosa C,
Collantes E, Guadalajara H, García-Olmo D and García-Olmo DC:
NIH-3T3 fibroblasts cultured with plasma from colorectal cancer
patients generate poorly differentiated carcinomas in mice. Cancer
Lett. 316:85–90. 2012. View Article : Google Scholar
|
|
34
|
Yu F, Hsieh WS, Petersson F, Yang H, Li Y,
Li C, Low SW, Liu J, Yan Y, Wang DY, et al: Malignant cells derived
from 3T3 fibroblast feeder layer in cell culture for nasopharyngeal
carcinoma. Exp Cell Res. 322:193–201. 2014. View Article : Google Scholar
|
|
35
|
Yang M, Reynoso J, Jiang P, Li L, Moossa
AR and Hoffman RM: Transgenic nude mouse with ubiquitous green
fluorescent protein expression as a host for human tumors. Cancer
Res. 64:8651–8656. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dong J, Dai XL, Lu ZH, Fei XF, Chen H,
Zhang QB, Zhao YD, Wang ZM, Wang AD, Lan Q, et al: Incubation and
application of transgenic green fluorescent nude mice in
visualization studies on glioma tissue remodeling. Chin Med J.
125:4349–4354. 2012.PubMed/NCBI
|
|
37
|
Iyer S, Arindkar S, Mishra A, Manglani K,
Kumar JM, Majumdar SS, Upadhyay P and Nagarajan P: Development and
evaluation of transgenic nude mice expressing ubiquitous green
fluorescent protein. Mol Imaging Biol. 17:471–478. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ricard C and Debarbieux FC: Six-color
intravital two-photon imaging of brain tumors and their dynamic
microenvironment. Front Cell Neurosci. 8:572014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Suetsugu A, Katz M, Fleming J, Truty M,
Thomas R, Moriwaki H, Bouvet M, Saji S and Hoffman RM: Multi-color
palette of fluorescent proteins for imaging the tumor
microenvironment of orthotopic tumorgraft mouse models of clinical
pancreatic cancer specimens. J Cell Biochem. 113:2290–2295. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Duda DG, Fukumura D, Munn LL, Booth MF,
Brown EB, Huang P, Seed B and Jain RK: Differential
transplantability of tumor-associated stromal cells. Cancer Res.
64:5920–5924. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang A, Dai X, Cui B, Fei X, Chen Y, Zhang
J, Zhang Q, Zhao Y, Wang Z, Chen H, et al: Experimental research of
host macrophage canceration induced by glioma stem progenitor
cells. Mol Med Rep. 11:2435–2442. 2015.
|
|
42
|
Huang Q, Zhang QB, Dong J, Wu YY, Shen YT,
Zhao YD, Zhu YD, Diao Y, Wang AD and Lan Q: Glioma stem cells are
more aggressive in recurrent tumors with malignant progression than
in the primary tumor, and both can be maintained long-term in
vitro. BMC Cancer. 8:3042008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wan Y, Fei XF, Wang ZM, Jiang DY, Chen HC,
Yang J, Shi L and Huang Q: Expression of miR-125b in the new,
highly invasive glioma stem cell and progenitor cell line SU3. Chin
J Cancer. 31:207–214. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Okabe M, Ikawa M, Kominami K, Nakanishi T
and Nishimune Y: 'Green mice' as a source of ubiquitous green
cells. FEBS Lett. 407:313–319. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Seabright M: A rapid banding technique for
human chromosomes. Lancet. 2:971–972. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bouvet M and Hoffman RM: Tumor imaging
technologies in mouse models. Methods Mol Biol. 1267:321–348. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Suetsugu A, Osawa Y, Nagaki M, Saji S,
Moriwaki H, Bouvet M and Hoffman RM: Imaging the recruitment of
cancer-associated fibroblasts by liver-metastatic colon cancer. J
Cell Biochem. 112:949–953. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhu L, Cheng X, Ding Y, Shi J, Jin H, Wang
H, Wu Y, Ye J, Lu Y, Wang TC, et al: Bone marrow-derived
myofibroblasts promote colon tumorigenesis through the
IL-6/JAK2/STAT3 pathway. Cancer Lett. 343:80–89. 2014. View Article : Google Scholar
|
|
49
|
Goldenberg DM, Rooney RJ, Loo M, Liu D and
Chang CH: In-vivo fusion of human cancer and hamster stromal cells
permanently transduces and transcribes human DNA. PLoS One.
9:e1079272014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Goldenberg DM, Gold DV, Loo M, Liu D,
Chang CH and Jaffe ES: Horizontal transmission of malignancy:
In-vivo fusion of human lymphomas with hamster stroma produces
tumors retaining human genes and lymphoid pathology. PLoS One.
8:e553242013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
He X, Li B, Shao Y, Zhao N, Hsu Y, Zhang Z
and Zhu L: Cell fusion between gastric epithelial cells and
mesenchymal stem cells results in epithelial-to-mesenchymal
transition and malignant transformation. BMC Cancer. 15:242015.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Goldenberg DM, Zagzag D, Heselmeyer-Haddad
KM, Berroa Garcia LY, Ried T, Loo M, Chang CH and Gold DV:
Horizontal transmission and retention of malignancy, as well as
functional human genes, after spontaneous fusion of human
glioblastoma and hamster host cells in vivo. Int J Cancer.
131:49–58. 2012. View Article : Google Scholar :
|
|
53
|
Pathak S, Nemeth MA, Multani AS, Thalmann
GN, von Eschenbach AC and Chung LW: Can cancer cells transform
normal host cells into malignant cells? Br J Cancer. 76:1134–1138.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Jacobsen BM, Harrell JC, Jedlicka P,
Borges VF, Varella-Garcia M and Horwitz KB: Spontaneous fusion
with, and transformation of mouse stroma by, malignant human breast
cancer epithelium. Cancer Res. 66:8274–8279. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Rappa G, Mercapide J and Lorico A:
Spontaneous formation of tumorigenic hybrids between breast cancer
and multipotent stromal cells is a source of tumor heterogeneity.
Am J Pathol. 180:2504–2515. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhou XG, Yang Y, Yang JS, Zhou J, Fang TL,
Dai WD and Chen ZR: Granulocyte-macrophage colony-stimulating
factor and interleukin 4 induce the malignant transformation of the
bone marrow-derived human adult mesenchymal stem cells. Chin Med J.
124:729–733. 2011.PubMed/NCBI
|
|
57
|
Al-Nedawi K, Meehan B, Micallef J, Lhotak
V, May L, Guha A and Rak J: Intercellular transfer of the oncogenic
receptor EGFRvIII by microvesicles derived from tumour cells. Nat
Cell Biol. 10:619–624. 2008. View Article : Google Scholar : PubMed/NCBI
|