Introduction
Chronic lymphocytic leukemia (CLL) is the most
common form of leukemia in adults, particularly in Western
countries (1). However, the
incidence of this disease is gradually increasing in China
(2). CLL is characterized by an
accumulation of abnormal B cells, resulting from the dysregulation
of proliferation and apoptosis rates (3,4).
Despite the advance in pathobiologic research and the development
of effective treatment regimens, CLL is still largely an incurable
disease (5).
It has been reported that p53 is inactivated in
10–15% of CLL patients decreasing cell apoptosis and accelerating
disease development (6).
Additionally, the anti-apoptotic protein Bcl-2 family is
overexpressed, while pro-apoptotic proteins such as Bax and Bcl-xL
are underexpressed in CLL cells (1,7). Bcl-2
upregulation increases the Bcl-2/Bax ratio, which further inhibits
the caspase-dependent apoptosis of CLL cells (8–10).
Therefore, the abnormal expression of apoptosis-related genes has
restricted the application of chemotherapeutic or immunotherapeutic
medicine and weakened the therapeutic outcome in CLL patients.
Recently, the introduction and subsequent approval
of targeted kinase inhibitors (idelalisib and ibrutinib) has
altered the standard of care for CLL patients and have acquired
outstanding efficacy (11,12). In addition, other novel
small-molecule inhibitors, such as venetoclax (ABT-199), an
inhibitor of Bcl-2 currently in clinical trials, have the potential
to improve therapy for CLL patients (12). Therefore, it is essential to develop
novel agents to target the abnormal activation of genes for CLL
treatment.
Plumbagin, a natural compound from Plumbago
zeylinica, has been shown to function as an anti-bacterial,
anti-atherosclerotic as well as an anticancer agent (13,14).
It also exhibits pro-apoptotic activities in different tumor cells
and animal models both in vitro and in vivo (15). In the present study, we explored the
possible anticancer activity of plumbagin in CLL cells by analyzing
its effects on cell viability, cell cycle regulation, proliferation
and apoptosis, as well as the expression of apoptosis-related
signaling molecules. Our data showed that plumbagin reduced CLL
cell survival by downregulating Bcl-2, but upregulating the Bax
level, suggesting that plumbagin may be considered as a promising
agent for the treatment of CLL.
Materials and methods
Patients and samples
CLL samples were obtained from the Affiliated
Hospital of Xuzhou Medical College according to the diagnostic
criteria for CLL between September 2013 and October 2015, while 6
healthy volunteers served as a normal control group. Peripheral
blood mononuclear cells (PBMCs) were isolated from heparinized
blood obtained from 35 CLL patients.
All procedures performed in the study involving
human participants were in accordance with the ethical standards of
the Institutional and/or National Research Committee and with the
1964 Helsinki Declaration and its later amendments or comparable
ethical standards.
Cell lines and reagents
CLL cell lines, HG3 and MEC-1, a kind gift from
Anders Rosén at Linköping University, were cultured in RPMI-1640
medium supplemented with 10% (v/v) fetal bovine serum (FBS) (Gibco,
Grand Island, NY, USA) at 37°C in a 5% CO2 incubator
(Thermo Fisher Scientific, Waltham, MA, USA). The Bcl-2 and Bax
antibodies were purchased from Cell Signaling Technology (Danvers,
MA, USA) and anti-actin was purchased from Santa Cruz Biotechnology
(Santa Cruz, CA, USA). Goat anti-rabbit IgG and rabbit anti-mouse
IgG were obtained from Sigma-Aldrich (St. Louis, MO, USA). Cell
Counting Kit-8 (CCK-8) was purchased from Dojindo Molecular
Technologies (Xiongben, Japan). The Cell-Light EdU
Apollo® 567 In Vitro Imaging kit was from Ruibo
Biotechnology (Guangzhou, China). Annexin V and 7-AAD
double-labeled apoptosis detection kit and propidium iodide (PI)
were obtained from eBiosience (San Diego, CA, USA). Plumbagin was
purchased from Sigma-Aldrich.
Quantitative real-time PCR
Extracted RNA from the CLL samples was
reverse-transcripted to cDNA which was used for quantification of
mRNA expression of Bcl-2 and Bax by real-time PCR with GADPH as
internal control. Amplification was performed in triplicate on
LightCycler® R480 II (Roche Life Science) in a total
volume of 20 μl. The primers for Bcl-2, Bax and GADPH were
as follows: Bcl-2 (F), 5′-ACGACTTCTCCCGCCGCTA-3′ and (R),
5′-ACCCCACCGAACTCAAAGAAG-3′; Bax (F), 5′-AGAGGATGATTGCCGCCGT-3′ and
(R), 5′-CAACCACCCTGGTCTTGGATC-3′; GAPDH (F),
5′-TGAAGGTCGGAGTCAACGGATT-3 and (R), 5′-CCTGGAAGATGGTGATGGGATT-3′.
qPCR reaction was performed according to the SYBR-Green qPCR
SuperMix manuals. The relative mRNA expression of target genes was
calculated by the comparative Ct method which was performed using
the following formula: Relative expression = 2−ΔΔCt.
CCK-8 analysis of cell viability
Three thousand cells in 100 μl of medium were
seeded into 96-well plates with three replicates. The cells were
incubutated with 0.625, 1.25, 2.5, 5, 10 and 20 μM of
plumbagin or fludarabine for 48 h, or incubated with 10 μM
of plumbagin or fludarabine for 0, 20, 40, 60 and 80 h, and then
CCK-8 reagent (5 μl) was added into each well to incubate
for an additional 4 h. The cells were exposed to measure the
absorbance at 450 nm by a microplate reader (Wellscan MK-3;
Labsystems, Dargon, Finland).
5-Ethynyl-2′-deoxyuridine
(EdU)-incorporation for detecting cell proliferation analysis
EdU-incorporation is a method for labeling DNA in
vivo during DNA replication. The HG3 and MEC-1 cells were
pretreated with 0, 2.5, 5 and 10 μM of plumbagin for 48 h,
and then incubated with EdU for an additional 3 h. After fixation,
permeabilization and staining according to the kit manual, the
cells were observed and the number of EdU-positive cells was
calculated under a microscope.
Cell cycle analysis
The HG3 and MEC-1 cells following the same
treatments with plumbagin were collected and fixed in 70% ethanol
on ice for 10 min, rinsed with phosphate-buffered saline (PBS) and
incubated with 100 mg/ml RNase A (0.25 mg/ml) for 15 min. After
washing with PBS for two times, the cells were further incubated
with 50 μg/ml PI at room temperature for 10 min, and then
the cells were subjected to cell cycle analysis.
Analysis of cell apoptosis
The HG3 and MEC-1 cells following the same
treatments with plumbagin were collected and then resuspended in 1×
binding buffer at 1–5×106/ml, and incubated with Annexin
V-APC and 7-AAD for 10 min at room temperature for analysis of
apoptosis by flow cytometry. Early apoptotic cells were labeled
with Annexin V and late apoptotic cells were double labeled with
Annexin V and 7-AAD.
Immunoblotting
The HG3 and MEC-1 cells following the same
treatments with plumbagin were collected and proteins were
extracted for western blotting. Equal amount of protein lysates
were subjected to 10–12% SDS-PAGE, and then transferred to a
0.45-μm pore size polyvinylidene fluoride (PVDF) membrane
(Millipore, Billerica, MA, USA). After blocking with 5% non-fat
milk, the membrane was probed with primary antibodies at 4°C
overnight and secondary antibodies at room temperature for 1 h.
Bound antibodies were detected using Pierce ECL Plus Western
Blotting Substrate (Thermo Fisher Scientific) and visualized by
ImageQuant LAS 4000 (GE Healthcare, Fairfield, CT, USA).
Statistical analysis
The results are representative of experiments
repeated at least three times and quantitative data are expressed
as means ± SEM. Student's t-test and ANOVA test were used to
analyze the difference between groups. P<0.05 was considered
statistically significant, and P<0.01 as very significant. All
statistical analyses were performed using GraphPad Prism 6.0
(GraphPad Software, San Diego, CA, USA).
Results
Increased Bcl-2/Bax ratio in CLL
patients
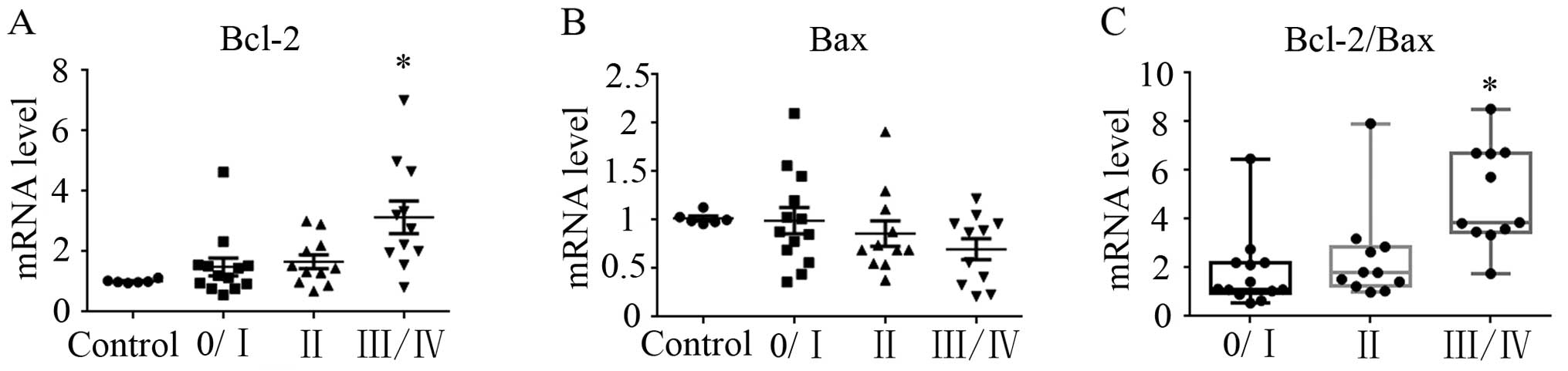
The mRNA expression of Bcl-2 and Bax was determined
in 35 CLL patients and 6 healthy donors. Bcl-2 exhibited an
increasing trend in Rai II–IV patients compared with the control
group, particularly in Rai III/IV patients (P<0.05; Fig. 1A). By contrast, Bax showed a
decreasing trend in the CLL patients, but without statistical
significance (Fig. 1B). However,
the expression ratio of Bcl-2/Bax was found to be significantly
increased (3.78-fold) in the Rai III/IV patients compared with this
ratio in the control group (P<0.05; Fig. 1C).
Plumbagin decreases cell viability and
inhibits cell proliferation
To investigate the potential growth inhibition of
plumbagin in CLL cells, the HG3 and MEC-1 cell lines were cultured
with 0.625, 1.25, 2.5, 5, 10 and 20 μM of plumbagin for 48
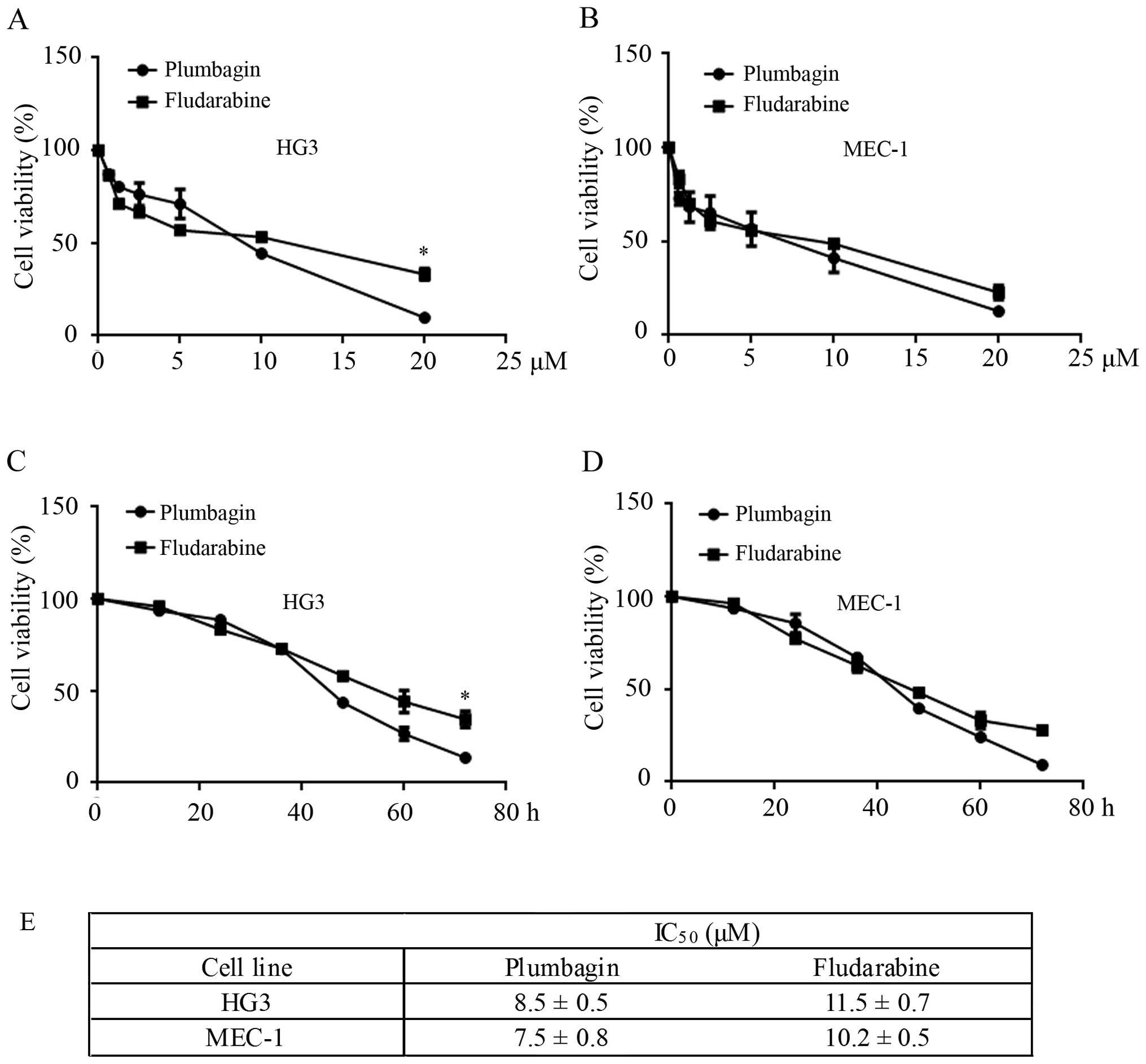
h, and the cell viability was determined by CCK-8 assay. As shown
in Fig. 2A and E, HG3 cells treated
with plumbagin and fludarabine presented decreased cell viability
in a dose-dependent manner, while the cells treated with plumbagin
showed a lower IC50 value (8.5±0.5 μM) compared
with fludarabine (11.5±0.7 μM) (P<0.05). Similarly, under
the same conditions, MEC-1 cells were more susceptible to plumbagin
(IC50=7.5±0.8 μM) compared to fludarabine
(IC50=10.2±0.5 μM) (Fig. 2B and E). Next, HG3 and MEC-1 cells
were incubated with 10 μM of plumbagin or fludarabine for 0,
20, 40, 60 and 80 h. Almost all CLL cells showed growth retardation
at 72 h when treated with plumbagin, while 34.5±0.7% (P<0.05)
and 28±0.3% (P<0.05) of HG3 and MEC-1 cells presented strong
cell viability after the same treatment with fludarabine (Fig. 2C and D).
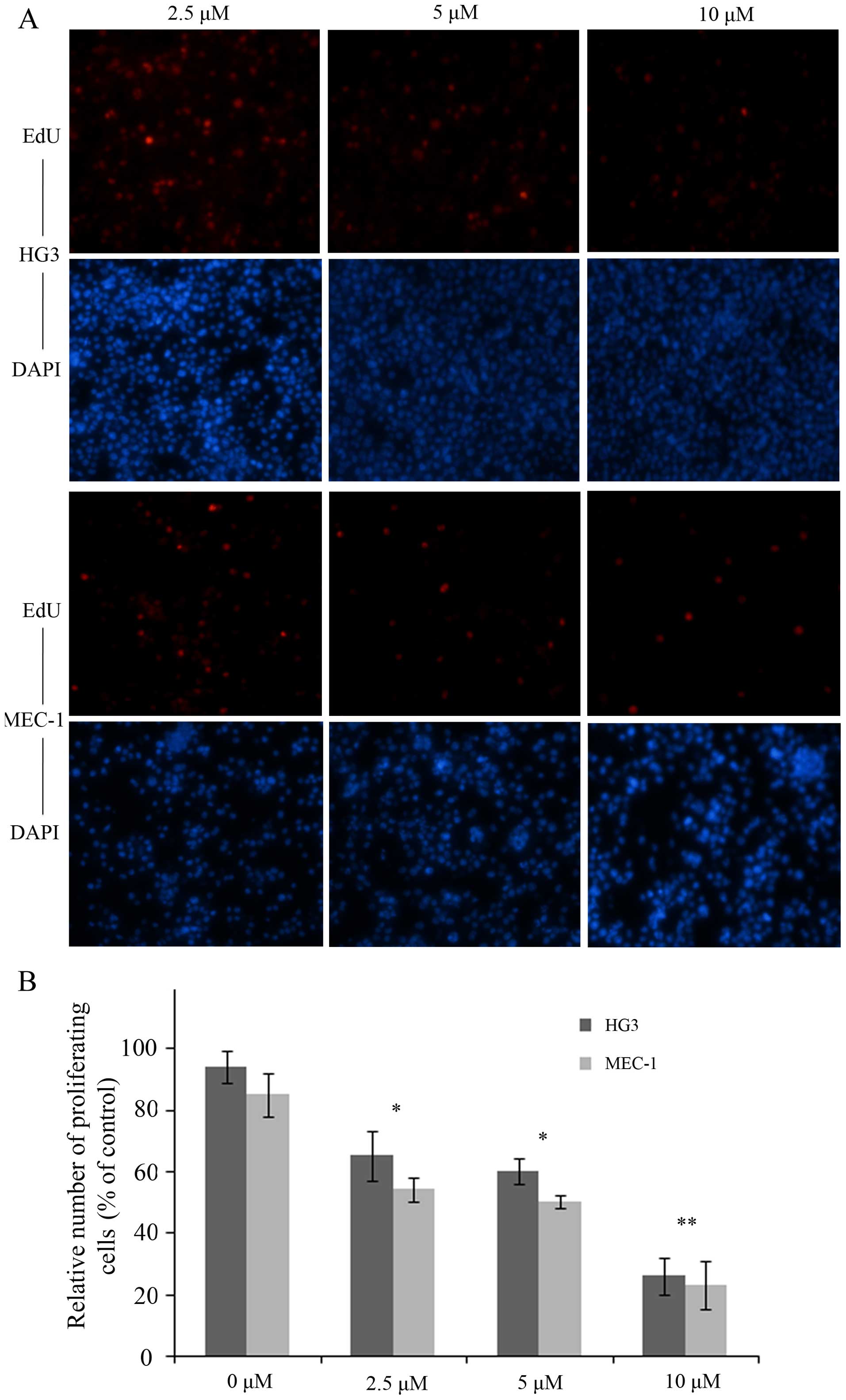
To assess the effect of plumbagin on cell
proliferation, we performed EdU incorporation assay with HG3 and
MEC-1 cells in the presence or absence of plumbagin. As expected,
the numbers of EdU-positive cells distributed as 68, 62 and 30% in
HG3 cells vs. 58, 54 and 28% in MEC-1 cells at 2.5, 5 and 10
μM of plumbagin, were significantly decreased in a
dose-dependent manner (P<0.05, P<0.05, P<0.01; Fig. 3).
Plumbagin induces cell cycle arrest in
the G0/G1 or S phase
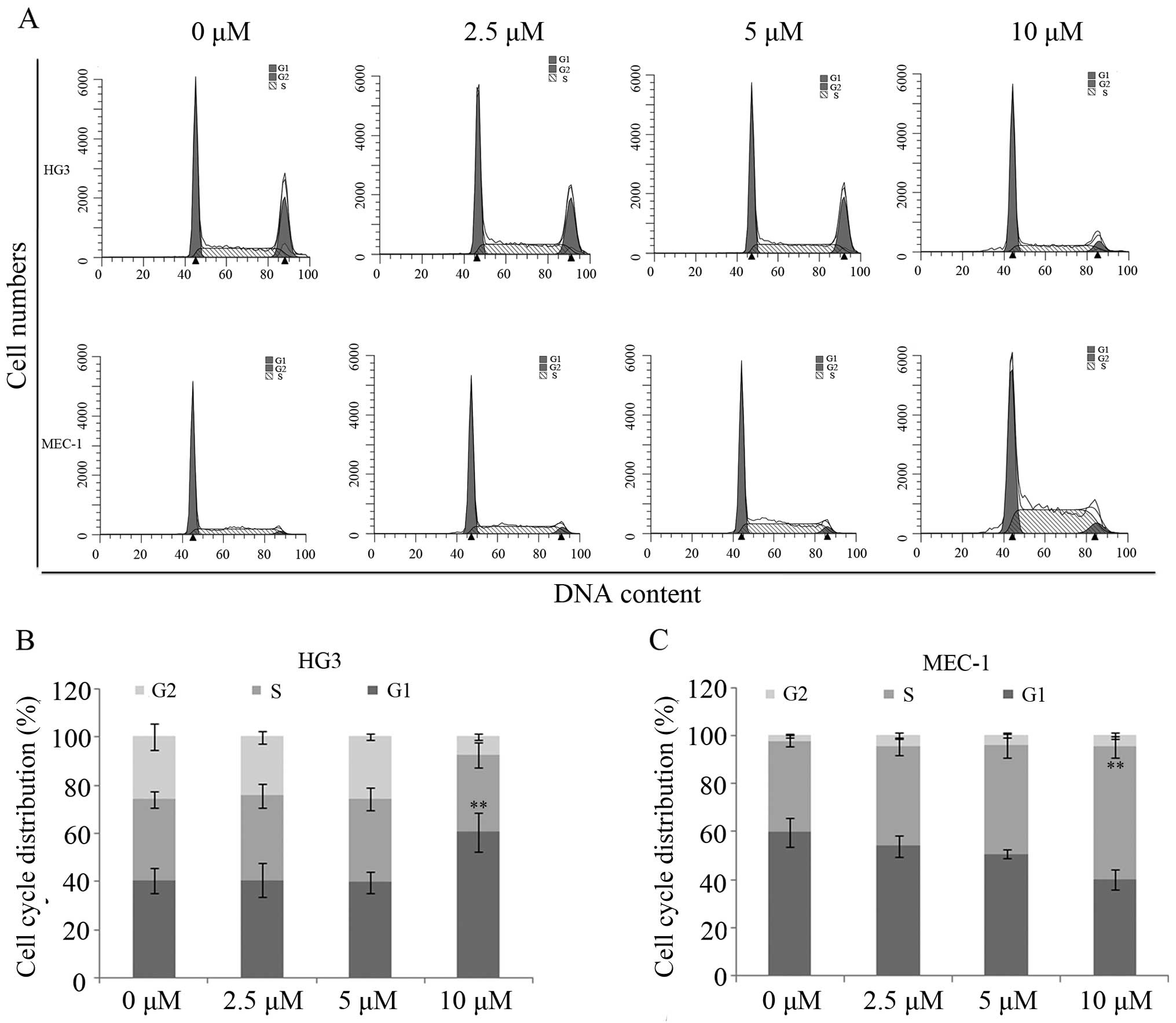
To investigate the role of plumbagin in cell cycle
progression, HG3 and MEC-1 cells treated with plumbagin were
collected and stained with PI for cell cycle analysis via flow
cytometry. The distribution of the cell cycle in the HG3 cells
exhibited no obverse changes (38–41% in G1, 19–27% in S and 35–40%
in G2/M phases) when treated with 0, 2.5 and 5 μM of
plumbagin. However, ~60% of the HG3 cells were arrested in G0/G1
phase following treatment with 10 μM of plumbagin
(P<0.01; Fig. 4A and B).
Notably, MEC-1 cells were blocked at the S phase following the same
treatment, and showed an increase from 33 to 53% compared with the
control when treated with 10 μM of plumbagin (P<0.01;
Fig. 4A and C).
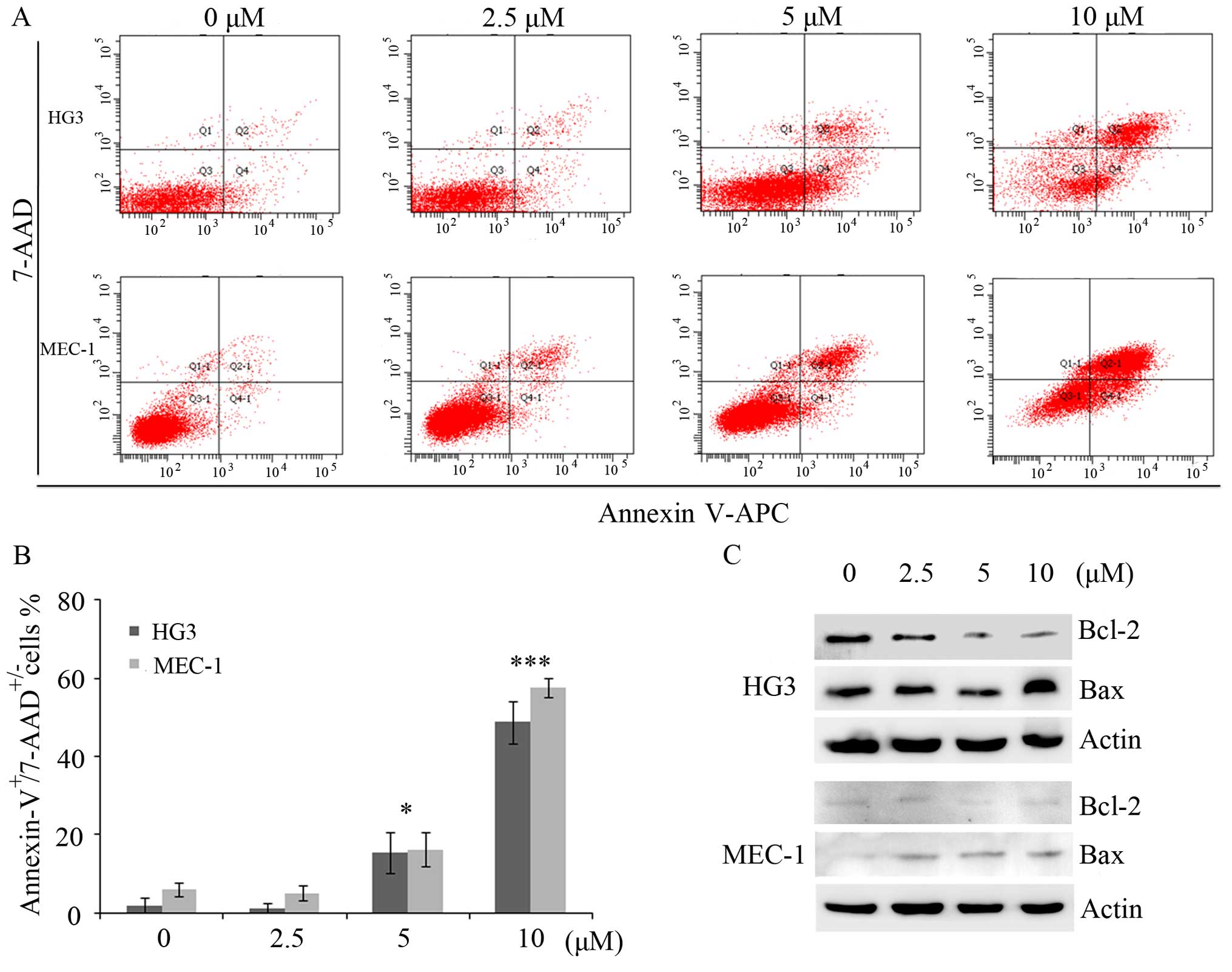
Plumbagin promotes cell apoptosis
To assess whether plumbagin induced these
growth-suppressive effects on CLL cells by apoptosis, we collected
HG3 and MEC-1 cells treated with plumbagin for double-staining
using Annexin V-APC and 7-AAD. As expected, plumbagin significantly
induced cell apoptosis in a dose-dependent manner, both in the HG3
and MEC-1 cell lines. In addition, statistical analysis showed that
the number of apoptotic cells treated with 5 or 10 μM of
plumbagin was 18±0.7 or 50±0.5% in the HG3 cells, and 19±0.5 or
58±0.2% in the MEC-1 cells (P<0.05; P<0.001; Fig. 5A and B). Furthermore, western
blotting showed that plumbagin treatment notably decreased the
expression of Bcl-2 and increased Bax in the HG3 cells. Similarly,
the expression of Bcl-2 was slightly decreased, while the Bax level
was strongly elevated even following treatment with a dose of 2.5
μM of plumbagin in the MEC-1 cells (Fig. 5C).
Discussion
Chronic lymphocytic leukemia (CLL) is the most
common human leukemia, representing 30% of all cases (16). CLL cells show higher expression of
Bcl-2 (17), higher activity of the
PI3K/Akt pathway (18) as well as
constitutive activation of NF-κB than normal lymphocytes (19). Approximately 10–15% of CLL patients
reveal structural aberrations or point mutations in locus 17p13,
containing TP53 (6). These
activated proteins or abnormal genes result in the defective
apoptosis of CLL cells in response to traditional therapeutic
medicine. Thus, a personalized therapeutic approach based on
genetic and molecular status could be preferable to the
comprehensive treatment of CLL. Accordingly, several agents
targeting B cell receptor signaling pathway kinases have entered
clinical trials such as idelalisib and ibrutinib, separately
targeting phosphoinositide 3-kinase (PI3K) or Bruton's tyrosine
kinase (BTK). Moreover, oblimersen sodium (G3139) specific for the
Bcl-2 mRNA sequence and obatoclax (GX15-070) or ABT-199
antagonistic for Bcl-2 protein have been applied in clinical phase
I/II study. Other targeting agents have also been assessed for use
in CLL therapy such as histone deacetylase (HDAC) inhibitors,
cyclin-dependent kinase inhibitors and proteasome inhibitors
(11,20).
Plumbagin, a potential anticancer agent, has been
reported to regulate cell proliferation and apoptosis in the
development of cancer (21,22). As reported, plumbagin induced colon
cancer cell death via a Bak-dependent pathway (23), and promoted tongue squamous cell
carcinoma cell arrest at the G2/M phase and apoptosis via p38 MAPK-
and PI3K/Akt/mTOR-mediated pathways (24). Plumbagin also inhibited prostate
xenograft development via targeting PKCε and Stat3 (25). In the present study, we found that
plumbagin inhibited CLL cell viability with a lower dose compared
to fludarabine, and inhibited cell proliferation in a
dose-dependent manner. As reported, plumbagin showed anticancer
activity in human osteosarcoma (MG-63) cells via the inhibition of
S phase checkpoints (20).
Consistently, the present study showed that plumbagin caused a
marked accumulation of MEC-1 cells in the S phase. Notably, most of
the HG3 cells, another CLL cell line, were blocked at the G0/G1
phase following the same plumbagin treatment. These results suggest
that plumbagin regulated various cell cycle-related proteins, and
finally induced cell cycle arrest.
It has been reported that the overexpression of
anti-apoptotic proteins such as Bcl-2, Bag-1 and Mcl-1 results in
CLL cell accumulation or apoptotic resistance (26,27).
In agreement with previous results, the majority of the Chinese CLL
patients in the present study showed a high Bcl-2 mRNA, but a
slightly low Bax level as determined by qPCR. Further statistical
analysis also showed an increasing trend for the Bcl-2/Bax ratio in
the CLL patients, particularly for those in Rai III/IV phase,
suggesting that the Bcl-2/Bax ratio is associated with the
development of CLL. Fortunately, after long-time incubation with
plumbagin, both CLL cell lines presented an increased apoptotic
trend in a dose-dependent manner by downregulation of Bcl-2 and
upregulation of Bax. These results suggest that plumbagin induced
CLL cell apoptosis probably via a decrease in the ratio of
Bcl-2/Bax. Nevertheless, the targeted proteins or regulatory
mechanism of plumbagin in regards to Bcl-2 or Bax is unclear,
deserving further study.
In summary, our results showed that plumbagin
effectively decreased CLL cell viability, inhibited cell
proliferation and blocked cell cycle progression using a lower dose
compared to fludarabine. In addition, we found that plumbagin
markedly induced CLL cell apoptosis by reducing Bcl-2, but by
increasing the Bax level. These results suggest that plumbagin
decreases the ratio of Bcl-2/Bax, thereby killing CLL cells.
Acknowledgments
We are grateful to Dr Anders Rosén at Linköping
University for generously providing the HG3 and MEC-1 cell lines.
The present study was supported by grants from the National Natural
Science Foundation of China (nos. 81400127, 81201264, 81200376 and
81302034), the Jiangsu Special Grant of Clinical Science
(BL2013010) and the Certificate of China Postdoctoral Science
Foundation Grant (2015M571818).
References
|
1
|
Malek SN: The biology and clinical
significance of acquired genomic copy number aberrations and
recurrent gene mutations in chronic lymphocytic leukemia. Oncogene.
32:2805–2817. 2013. View Article : Google Scholar :
|
|
2
|
Zhu DX, Zhu W, Fang C, Fan L, Zou ZJ, Wang
YH, Liu P, Hong M, Miao KR, Liu P, et al: miR-181a/b significantly
enhances drug sensitivity in chronic lymphocytic leukemia cells via
targeting multiple anti-apoptosis genes. Carcinogenesis.
33:1294–1301. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wójtowicz M and Wołowiec W: Dysregulation
of apoptosis and proliferation in CLL cells. Chronic Lymphocytic
Leukemia. Intech; Rijeka: pp. 37–62. 2012, View Article : Google Scholar
|
|
4
|
Bianchi S, Dighiero G and Pritsch O:
Selected topics in chronic lymphocytic leukemia pathogenesis.
Chronic Lymphocytic Leukemia. Intech; Rijeka: pp. 3–18. 2012,
http://cdn.intechopen.com/pdfs-wm/27981.pdf.
|
|
5
|
Rozovski U, Hazan-Halevy I, Keating MJ and
Estrov Z: Personalized medicine in CLL: Current status and future
perspectives. Cancer Lett. 352:4–14. 2014. View Article : Google Scholar
|
|
6
|
Antosz H, Paterski A, Marzec-Kotarska B,
Sajewicz J and Dmoszyńska A: Alterations in TP53, cyclin D2, c-Myc,
p21WAF1/CIP1 and p27KIP1 expression associated with
progression in B-CLL. Folia Histochem Cytobiol. 48:534–541. 2010.
View Article : Google Scholar
|
|
7
|
Marschitz I, Tinhofer I, Hittmair A, Egle
A, Kos M and Greil R: Analysis of Bcl-2 protein expression in
chronic lymphocytic leukemia. A comparison of three
semiquantitation techniques. Am J Clin Pathol. 113:219–229. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Podhorecka M, Halicka D, Klimek P, Kowal
M, Chocholska S and Dmoszynska A: Resveratrol increases rate of
apoptosis caused by purine analogues in malignant lymphocytes of
chronic lymphocytic leukemia. Ann Hematol. 90:173–183. 2011.
View Article : Google Scholar :
|
|
9
|
Salakou S, Kardamakis D, Tsamandas AC,
Zolota V, Apostolakis E, Tzelepi V, Papathanasopoulos P, Bonikos
DS, Papapetropoulos T, Petsas T, et al: Increased Bax/Bcl-2 ratio
up-regulates caspase-3 and increases apoptosis in the thymus of
patients with myasthenia gravis. In Vivo. 21:123–132.
2007.PubMed/NCBI
|
|
10
|
Fu NY, Sukumaran SK and Yu VC: Inhibition
of ubiquitin-mediated degradation of MOAP-1 by apoptotic stimuli
promotes Bax function in mitochondria. Proc Natl Acad Sci USA.
104:10051–10056. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Woyach JA and Johnson AJ: Targeted
therapies in CLL: Mechanisms of resistance and strategies for
management. Blood. 126:471–477. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang Y, Wu JZ, Li JY and Xu W: Know the
enemy as well as the weapons in hand: The aberrant death pathways
and therapeutic agents in chronic lymphocytic leukemia. Am J Cancer
Res. 5:2361–2375. 2015.PubMed/NCBI
|
|
13
|
Wang YC and Huang TL: Screening of
anti-Helicobacter pylori herbs deriving from Taiwanese folk
medicinal plants. FEMS Immunol Med Microbiol. 43:295–300. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang YC and Huang TL: Anti-Helicobacter
pylori activity of Plumbago zeylanica L. FEMS Immunol Med
Microbiol. 43:407–412. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Eldhose B, Gunawan M, Rahman M, Latha MS
and Notario V: Plumbagin reduces human colon cancer cell survival
by inducing cell cycle arrest and mitochondria-mediated apoptosis.
Int J Oncol. 45:1913–1920. 2014.PubMed/NCBI
|
|
16
|
Pekarsky Y, Zanesi N and Croce CM:
Molecular basis of CLL. Semin Cancer Biol. 20:370–376. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hanada M, Delia D, Aiello A, Stadtmauer E
and Reed JC: bcl-2 gene hypomethylation and high-level expression
in B-cell chronic lymphocytic leukemia. Blood. 82:1820–1828.
1993.PubMed/NCBI
|
|
18
|
Liu FT, Giustiniani J, Farren T, Jia L,
Bensussan A, Gribben JG and Agrawal SG: CD160 signaling mediates
PI3K-dependent survival and growth signals in chronic lymphocytic
leukemia. Blood. 115:3079–3088. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Furman RR, Asgary Z, Mascarenhas JO, Liou
HC and Schattner EJ: Modulation of NF-kappa B activity and
apoptosis in chronic lymphocytic leukemia B cells. J Immunol.
164:2200–2206. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yan CH, Li F and Ma YC: Plumbagin shows
anticancer activity in human osteosarcoma (MG-63) cells via the
inhibition of S-Phase checkpoints and down-regulation of c-myc. Int
J Clin Exp Med. 8:14432–14439. 2015.PubMed/NCBI
|
|
21
|
Qiu JX, He YQ, Wang Y, Xu RL, Qin Y, Shen
X, Zhou SF and Mao ZF: Plumbagin induces the apoptosis of human
tongue carcinoma cells through the mitochondria-mediated pathway.
Med Sci Monit Basic Res. 19:228–236. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu X, Cai W, Niu M, Chong Y, Liu H, Hu W,
Wang D, Gao S, Shi Q, Hu J, et al: Plumbagin induces growth
inhibition of human glioma cells by downregulating the expression
and activity of FOXM1. J Neurooncol. 121:469–477. 2015. View Article : Google Scholar
|
|
23
|
Wang J, Guo W, Zhou H, Luo N, Nie C, Zhao
X, Yuan Z, Liu X and Wei Y: Mitochondrial p53 phosphorylation
induces Bak-mediated and caspase-independent cell death.
Oncotarget. 6:17192–17205. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pan ST, Qin Y, Zhou ZW, He ZX, Zhang X,
Yang T, Yang YX, Wang D, Qiu JX and Zhou SF: Plumbagin induces
G2/M arrest, apoptosis, and autophagy via p38 MAPK- and
PI3K/Akt/mTOR-mediated pathways in human tongue squamous cell
carcinoma cells. Drug Des Devel Ther. 9:1601–1626. 2015.
|
|
25
|
Hafeez BB, Zhong W, Fischer JW, Mustafa A,
Shi X, Meske L, Hong H, Cai W, Havighurst T, Kim K, et al:
Plumbagin, a medicinal plant (Plumbago zeylanica)-derived
1,4-naphthoquinone, inhibits growth and metastasis of human
prostate cancer PC-3M-luciferase cells in an orthotopic xenograft
mouse model. Mol Oncol. 7:428–439. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dyer MJ, Zani VJ, Lu WZ, O'Byrne A, Mould
S, Chapman R, Heward JM, Kayano H, Jadayel D, Matutes E, et al:
BCL2 translocations in leukemias of mature B cells. Blood.
83:3682–3688. 1994.PubMed/NCBI
|
|
27
|
Vrhovac R, Delmer A, Tang R, Marie JP,
Zittoun R and Ajchenbaum-Cymbalista F: Prognostic significance of
the cell cycle inhibitor p27Kip1 in chronic B-cell
lymphocytic leukemia. Blood. 91:4694–4700. 1998.PubMed/NCBI
|