Introduction
Colorectal cancer (CRC) is one of the most
aggressive gastrointestinal malignancies and the third leading
cause of cancer deaths in the world (1,2).
However, in recent years, its incidence has been increasing in
economically transitioning countries due to physical inactivity and
consumption of calorie-dense food (3,4).
Although surgical resection has been considered to be the effective
treatment for early-stage patients, approximately 20–45% of
patients who underwent curative resection developed metastasis or
relapse (5–7). Tumor metastasis is a multistep process
in which cancer cells escape from the primary, penetrate
hematogenously and colonize at distant sites (8–10).
Thus, understanding the genetic basis and molecular mechanism of
initiation and progression of CRC will help to find novel
therapeutic targets for CRC patients.
Cortactin (CTTN), a v-Src substrate, was reported
overexpressed in a variety of cancers including lung, breast, and
prostate cancers (11). CTTN was
increased in head and neck carcinogenesis and correlated with poor
prognosis (12). CTTN promoted cell
motility and tumor metastasis by binding and activating the
actin-related protein 2/3 complex to regulate the formation of
invadopodia (13). In addition,
overexpression of cortactin has been shown to enhance cell adhesion
and form lamellipodium, which contributed to the lamellipodial
persistence and migration phenotypes (14). Conversely, CTTN silencing inhibited
cell motility in wound-healing, cell migration and Matrigel
invasion assays (15). CTTN has
been reported upregulated in colorectal cancer tissues and
overexpression of CTTN promoted cell migration and invasion by
interacting with zonula occludens-1 (ZO-1) (16). However, the clinical significance
and the underlying molecular mechanism of CTTN in the progression
of CRC remained to be determined.
In this study, we investigated CTTN expression in
CRC tissue and its relationship with the clinicopathological
factors and prognosis of the CRC patients. Furthermore, we explored
the potential role of CTTN in CRC cell mobility and the underlying
molecular mechanisms in vitro.
Materials and methods
Tissues specimens
A total of 146 paired formalin-fixed
paraffin-embedded CRC and adjacent normal tissue specimens were
obtained from stage I-III CRC patients who received curative
surgery in Ruijin hospital from May 2009 to July 2010. These
patients were diagnosed by pathological analysis. According to the
tumor-node-metastasis (TNM) staging system of the American Joint
Committee on Cancer (AJCC, 7th edition), 63 cases (43.2%) were in
disease stage I/II and 83 cases (56.8%) were in disease stage III.
Written informed consent was obtained from all patients. This study
was approved by the Committee of Medical Ethics at Shanghai Jiao
Tong University.
Immunohistochemistry
Immunohistochemistry of paraffin-embedded tissue
sections was performed as previously described (17). In brief, tissue sections from
paraffin blocks were deparaffinised and rehydrated. Endogenous
peroxidase activity was blocked with 3% H2O2
for 10 min. Antigen retrieval was performed in citrate buffer (10
mM, pH 6.0) for 15 min at 100°C. The sections were incubated with a
primary antibody (Anti-CTTN antibody, Sigma-Aldrich, HPA057242) at
4°C overnight in a moist chamber. Finally, the visualization signal
was developed with diaminobenzidine (Dako). Images of these slides
stained for CTTN were scanned at ×20 magnification and captured by
microscopy (DP70, Olympus, Tokyo, Japan). The final staining scores
were evaluated by two pathologists without prior knowledge of the
clinical data.
Cell culture
Human colorectal cancer cell lines (SW480 and SW620)
were obtained from Shanghai Cell Bank of the Chinese Academy of
Sciences (Shanghai, China) and maintained in RPMI-1640 (Hyclone,
Logan, UT, USA) supplemented with 10% fetal bovine serum
(Invitrogen, Grand Island, NY, USA), penicillin (100 U/ml), and
streptomycin (100 mg/ml) at 37°C in a humidified atmosphere of 5%
CO2.
Establishment of CTTN-knockdown and
CTTN-overexpressing cells
The plasmid and lentivirus particles for CTTN
overexpression or knockdown were designed and constructed by
Shanghai GenePharma Co., Ltd. (Shanghai, China). SW480 cells were
transduced with Lenti-GFP (vector) or Lenti-CTTN (CTTN) at 20 MOI
in the presence of the 8 µg/ml polybrene (Sigma). SW620
cells were transduced with Lenti-control (control) or Lenti-shCTTN
(shCTTN) at 20 MOI in the presence of the 8 µg/ml polybrene.
Cells were selected stable cell lines continuously in puromycin (2
µg/ml).
siRNA transfection
CTTN-overexpressing SW480 cells were transfected
with negative control (NC) or DOCK1 siRNA using Lipofectamine RNA
interference MAX Reagent (Life Technologies) according to the
manufacturer's instructions. The siRNAs targeting CTTN and negative
control were obtained from Shanghai GenePharma Co., Ltd.
Cell adhesion assay
Approximately 2×105 cells were harvested
and resuspended in complete medium and then seeded into a 24-well
plate that was precoated with matrix proteins (20 µg/ml
fibronectin or 20 µg/ml laminin). After incubating for 1 h,
cells were washed twice with PBS to remove the non-adherent cells.
The bound cells were determined by WST-1 (Roche, Mannheim, Germany)
at a wavelength of 450 nm in a microplate reader. The change in
optical density was represented as fold of control.
Migration and invasion assay
Transwell migration assays were performed in 24-well
plates with 8 µm pore polycarbonate membrane (Corning,
Tewksbury, MA, USA). Cells (5×105) were starved for 12 h
and then added into the upper chambers of the wells in 100
µl serum-free medium. RPMI-1640 medium (600 µl) with
10% fetal bovine serum was added into the lower chamber. Cells were
incubated for 36 h at 37°C to allow cell invasion into the lower
chamber. The invaded cells were fixed and stained with 0.5% crystal
violet. Images were taken by microscopy (Olympus) at least in 8
representative fields. Invasion assay was performed with filters
pre-coated with Matrigel (BD Biosciences, Franklin Lakes, NJ, USA)
and the following protocols were the same as migration assay.
Western blot analysis
Cells were lysed with RIPA buffer (Cell Signaling
Technology, Danvers, MA, USA), and the protein concentration was
measured using the BCA assay (Thermo Fisher Scientific, Waltham,
MA, USA). Equal amounts of protein were subjected to 10% SDS-PAGE
for electrophoresis and transferred to a polyvinylidene difluoride
membrane (Millipore, Billerica, MA, USA). The membrane was blocked
with 5% fat-free milk at room temperate, then specific antibody
anti-CTTN antibody (Sigma-Aldrich), anti-DOCK1 antibody (Abcam,
Cambridge, MA, USA), anti-RAC1 antibody (Sigma-Aldrich), and
anti-GAPDH antibody (Cell Signaling Technology) were incubated
overnight at 4°C. Immunoreactive bands were visualized with
enhanced chemiluminescence (Thermo Fisher Scientific) using LAS
4000 imaging system (Fijifilm, Tokyo, Japan).
Statistical analysis
Data are presented as the mean ± standard deviation
of the mean. The χ2 test and the Fisher's test were used
to test correlations between CTTN expression and
clinicopathological parameters. Kaplan-Meier analysis with log-rank
test was performed to assess patient survival. Data were analyzed
using software SPSS 17.0 (SPSS Inc., Chicago, IL, USA) and a value
of P<0.05 was considered to indicate a statistically significant
difference.
Results
CTTN is upregulated in colorectal cancer
tissues
Previous studies have shown that CTTN was increased
in many types of human cancers, including colorectal cancer
(11). However, the expression of
CTTN in Chinese colorectal cancer was unknown. IHC was used to
detect the expression of CTTN in 146 paired CRC and adjacent normal
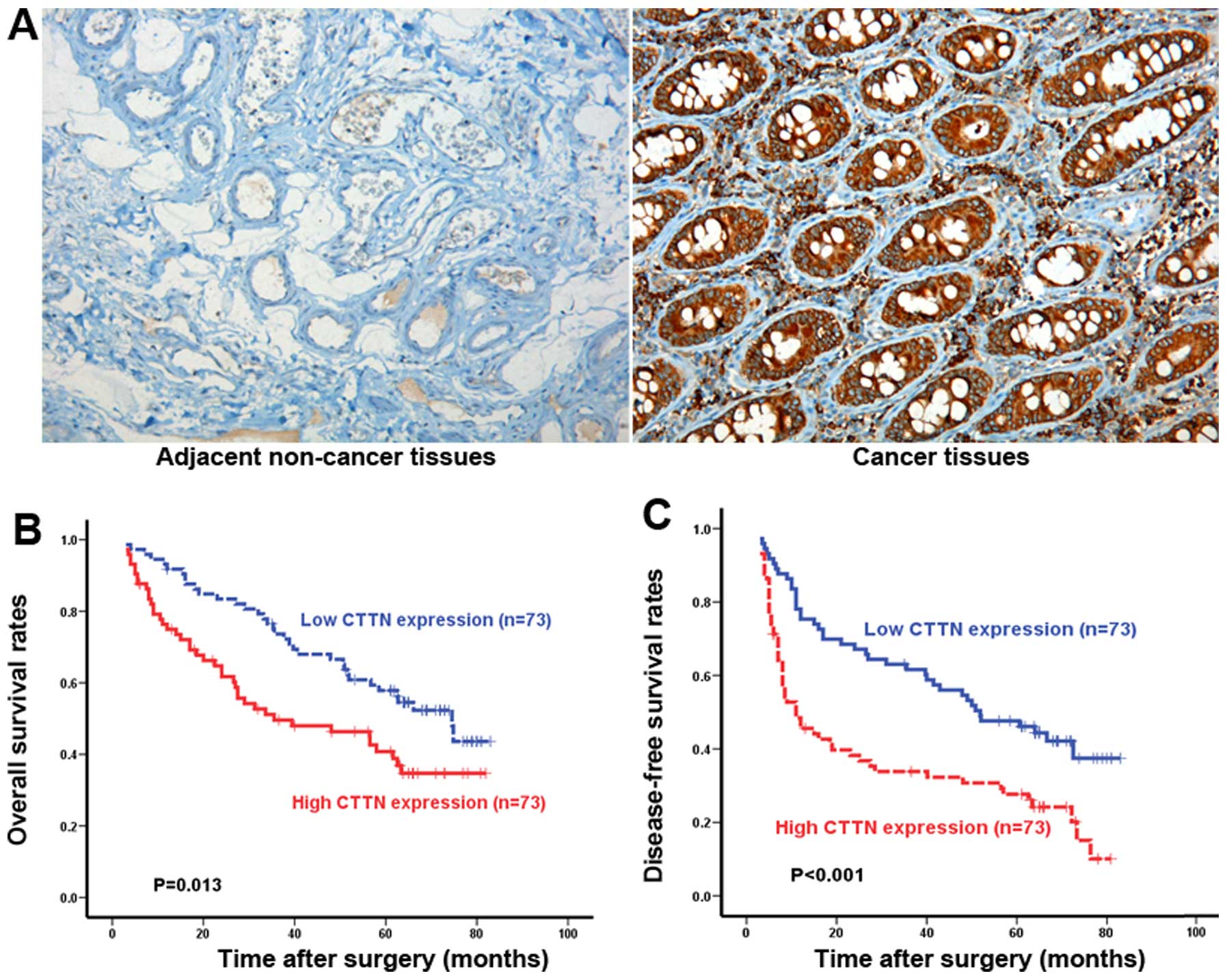
tissues. In 94 cases (64%), CTTN is expressed in the tumor tissues,
mainly localized in the cell cytoplasm (Fig. 1A). In contrast, no or weak staining
is seen in the cytoplasm of adjacent normal colorectal tissues
(Fig. 1A). The relationship between
CTTN expression and clinicopathological features is displayed in
(Table I). We found CTTN expression
was significantly correlated with depth of invasion (P<0.001),
lymph node involvement (P<0.001), distant metastasis
(P<0.031), lymph node metastasis (P<0.005), and TNM stage
(P=0.012). On the contrary, no significant differences between CTTN
expression and gender, age, tumor size, tumor differentiation, or
preoperative CEA level were observed. To further determine if the
expression of CTTN was associated with colorectal cancer patient
survival, Kaplan-Meier curve analysis is performed to assess
prognosis. As shown in Fig. 1B,
patients with high (n=73) CTTN expression had a significantly short
overall survival (OS) than those with low (n=73) CTTN expression
(P=0.013). There is also a statistically significant difference in
disease-free survival (DFS) between patients whose tumors had high
or low CTTN expression (P<0.001) (Fig. 1C). These data strongly indicated
that CTTN expression play an important role in the progression of
colorectal cancer.
 | Table IAssociations between CTTN expression
and multiple clinicopathological factors in CRC patients. |
Table I
Associations between CTTN expression
and multiple clinicopathological factors in CRC patients.
| Variables | CTTN expression level
| P-value |
|---|
Low
N=73 | High
N=73 |
|---|
| Gender | | | |
| Female | 25 | 27 | NS |
| Male | 48 | 46 | |
| Age, years | | | |
| ≤60 | 40 | 31 | NS |
| >60 | 33 | 42 | |
| Tumor size, cm | | | |
| ≤5 | 38 | 42 | NS |
| >5 | 35 | 31 | |
| Tumor
differentiation | | | |
| Well | 19 | 21 | NS |
| Moderate | 44 | 36 | |
| Poor | 10 | 16 | |
| Preoperative CEA
(ng/ml) | | | |
| ≤5 | 49 | 44 | NS |
| >5 | 24 | 29 | |
| Depth of
invasion | | | |
| Muscular layer | 40 | 27 | 0.031 |
| Serosa layer | 33 | 46 | |
| Lymph node
metastasis | | | |
| No | 48 | 27 | 0.0005 |
| Yes | 25 | 46 | |
| TNM stage | | | |
| I–II stage | 39 | 24 | 0.012 |
| III stage | 34 | 49 | |
Knockdown of CTTN inhibits cellular
matrix adhesion, migration and invasion in vitro
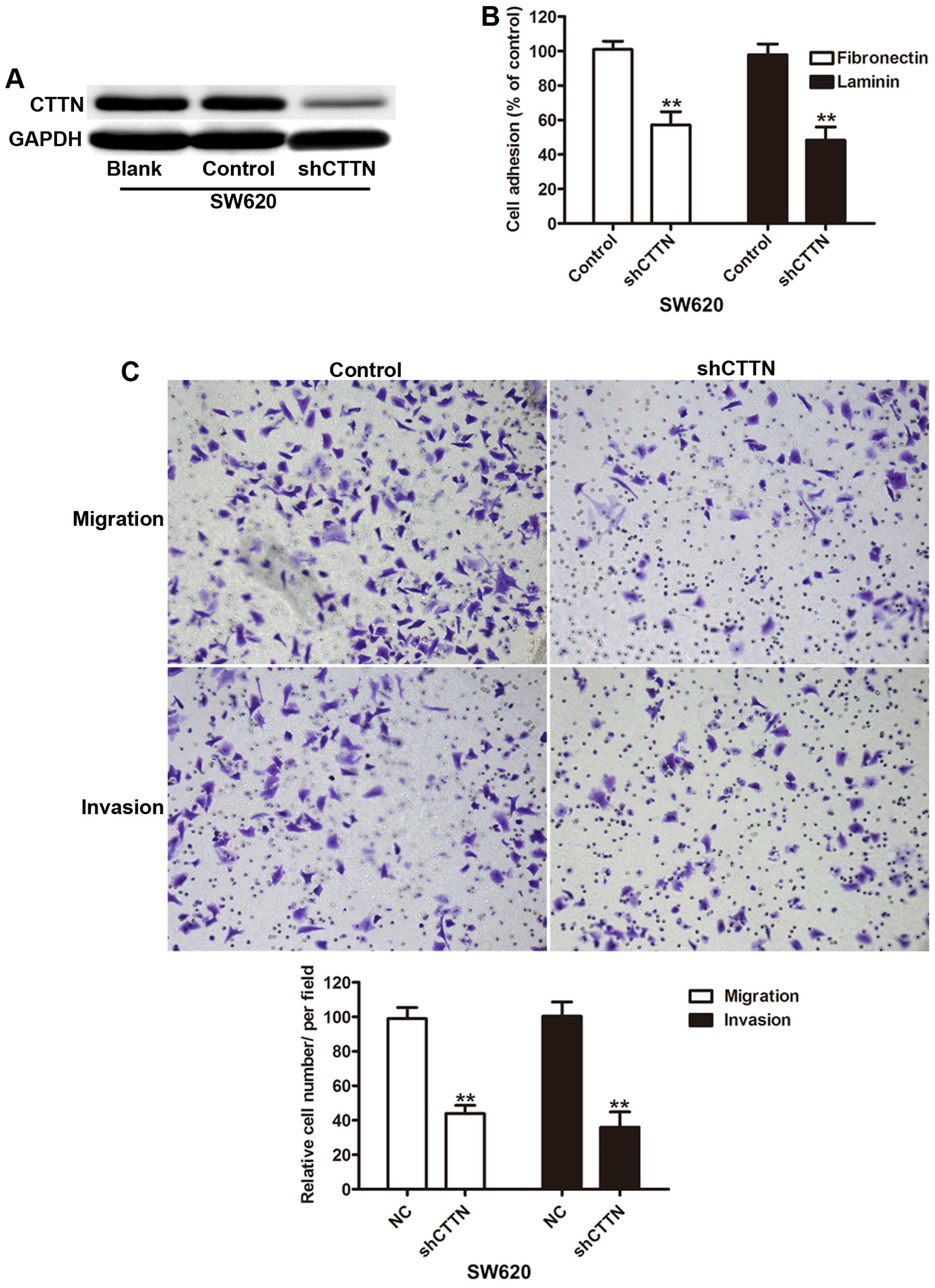
Since CTTN is increased in colorectal cancer
patients, we knocked down the CTTN expression in SW620 cells (high
invasion capability) to investigate the biological behavior.
Compared with the negative control (control) and blank, the levels
of CTTN protein was significantly decreased in SW620 cells
transfected with shCTTN (Fig. 2A).
To investigate the effect of CTTN on the migration and invasion of
SW620 cells, cell adhesion was first assessed and the results
indicated knockdown of CTTN caused lower adhesion capability
(Fig. 2B). Furthermore, Transwell
analysis showed that knockdown of CTTN decreases the migration and
invasion of SW620 cells compared to control (Fig. 2C).
CTTN promotes cellular matrix adhesion,
migration and invasion in vitro
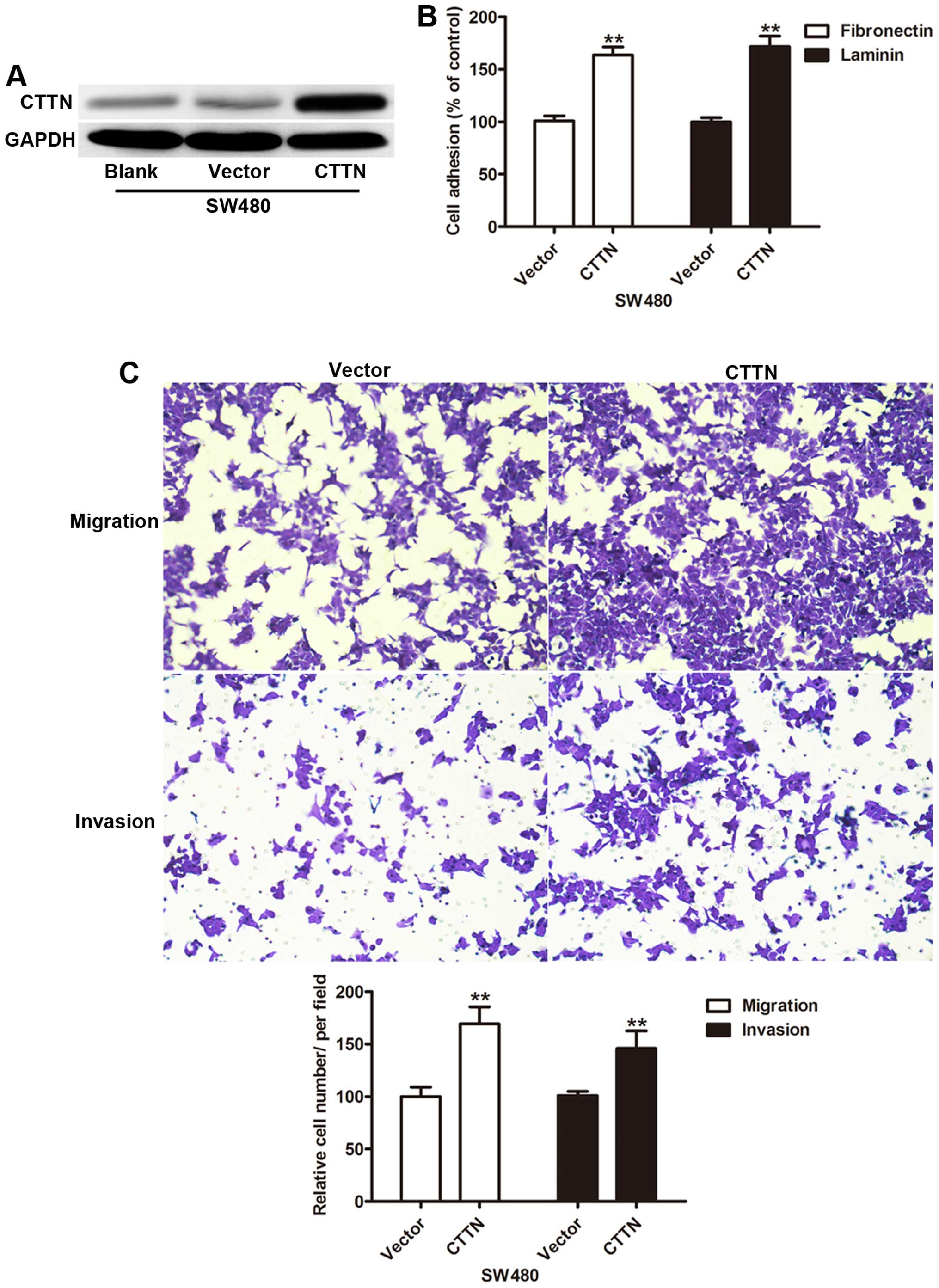
Next we explored whether CTTN expression impacts the
invasion capability, SW480 cells (low invasion capability) were
transfected with CTTN. Western blot results showed CTTN expression
was significantly increased in the CTTN group compared with the
control vector and blank groups (Fig.
3A). Gain of function of CTTN enhanced the adhesion capability
of SW480 cells transfected with CTTN (Fig. 3B). The cell invasion assay revealed
a significant increase in the average number of cells penetrating
the Transwell membrane and Matrigel in the CTTN group (Fig. 3C). These findings suggested that
CTTN played an important role in cell invasion.
CTTN mediates DOCK1 expression in
colorectal cancer cells
To further determine the mechanisms that CTTN
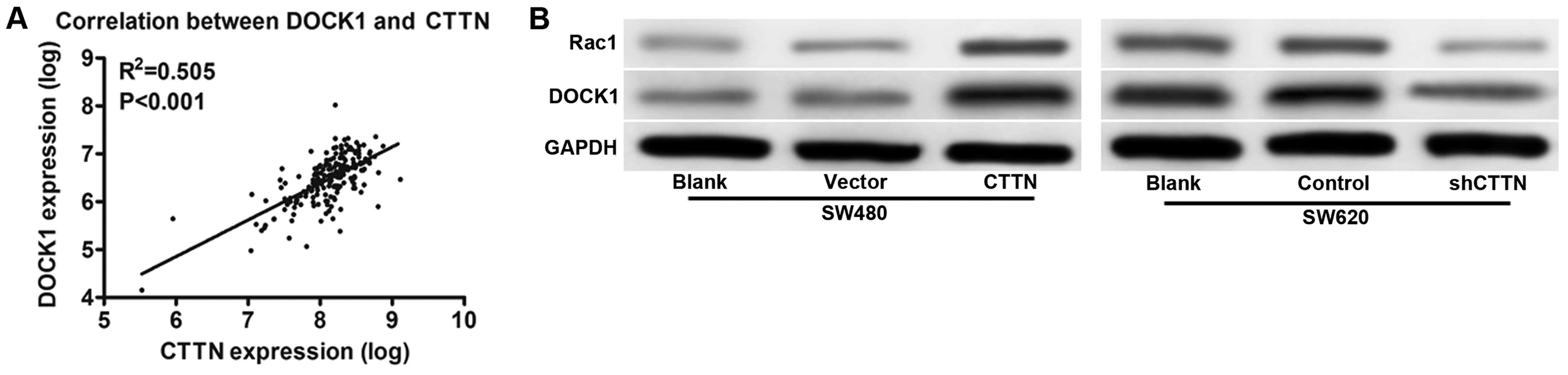
promote CRC cell invasion, we performed bioinformatic analysis to
predict protein interactions. We focused on DOCK1, which has been
documented as an oncogene in human cancers (Fig. 4A) (18,19).
As showed in Fig. 4B, DOCK1 was
significantly increased in CTTN overexpressing cells as compared
with control vector and consistently decreased DOCK1 expression in
SW620 cells by knockdown of CTTN. In addition, we also observed
Rac1, a downstream molecule, had a similar expression (Fig. 4B). These results indicated that the
expression level of DOCK1 was upregulated by CTTN in CRC cell lines
and prompted us to investigate the role of DOCK1 in CTTN-mediated
cell invasion.
DOCK1 is required for CTTN-induced cell
migration and invasion
To further investigate the role of DOCK1 in
CTTN-mediated cell mobility, we knocked down DOCK1 in SW480 cells
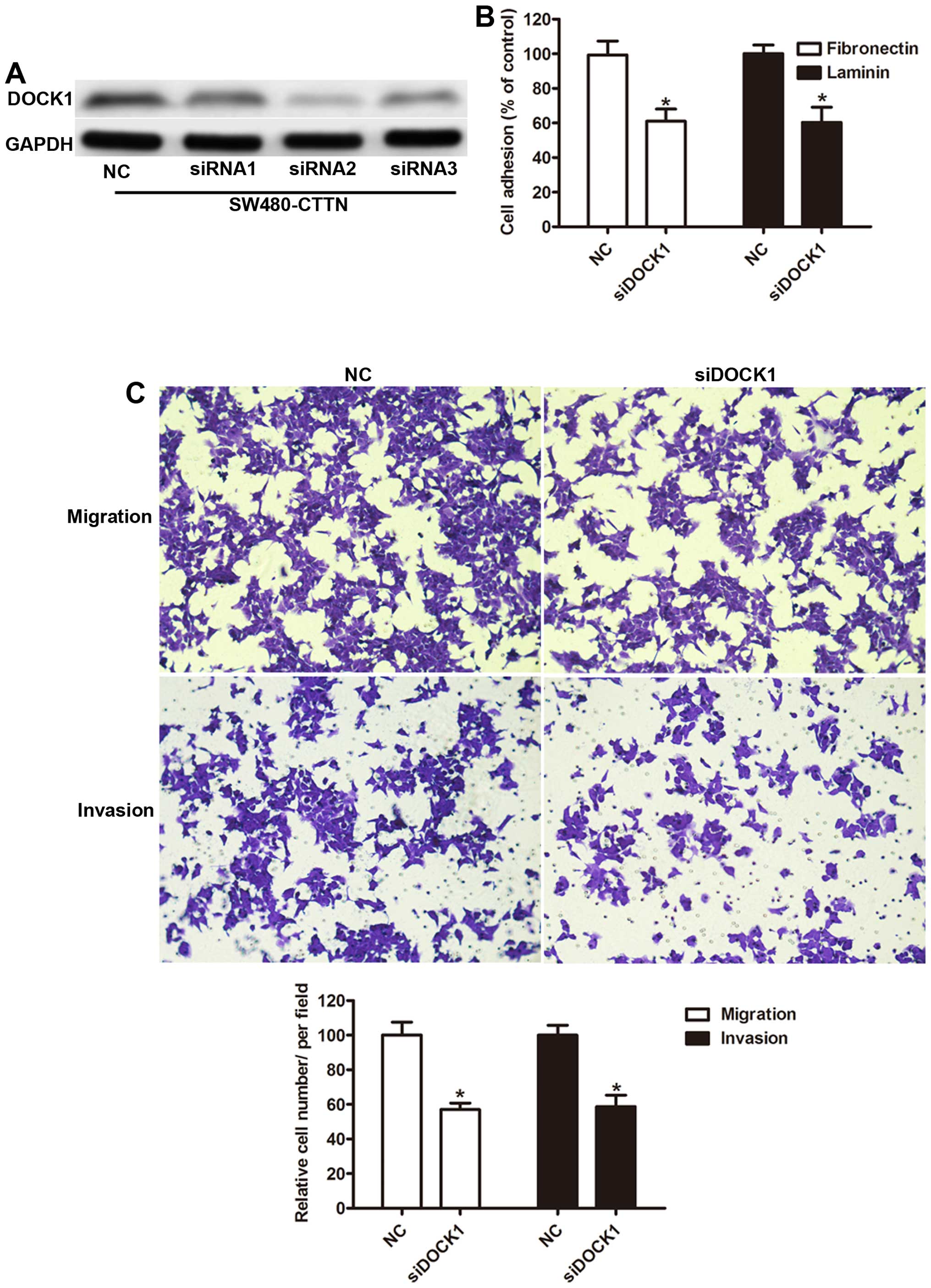
with stable overexpression of CTTN. As shown in Fig. 5A, compared to negative control (NC),
transfection with the siRNA1, siRNA2, and siRNA3 results in
inhibition of DOCK1 expression. siRNA2 had the most effective
inhibition and was used in the remainder of the study. Then we
measured the adhesion capability of SW480 cells and found DOCK1
silencing abrogated the effects of CTTN on adhesion capability
(Fig. 5B). Furthermore, inhibition
of DOCK1 also reduced the mean number of cells penetrating the
Transwell membrane and Matrigel (Fig.
5C), suggesting that DOCK1 plays a crucial role in cell
mobility of CRC cells and is involved in CTTN-mediated tumor
metastasis.
Discussion
Growing evidence shows a direct link between
disorder gene expression and cancer invasion and metastasis
(20). Aberrant activation of
cortactin (CTTN) has been implicated in tumorigenesis and
metastasis of numerous malignant tumors including esophageal
squamous cell carcinoma, breast, and colon cancer (16,21–23).
However, the prognostic significance of CTTN in CRC has not yet
been reported. Our results showed that the expression of CTTN was
increased in CRC tissues. Pathologic investigation of CRC patient
tissues suggested a significant correlation between CTTN expression
depth of invasion, lymph node metastasis, and Tumor-Node-Metastasis
stage. Kaplan-Meier analysis showed that patients with high CTTN
expression had a shorter OS and DFS. In vitro experiments,
CTTN silencing inhibited the invasive capability of CRC cells,
whereas overexpression of CTTN promote CRC invasion. These data
imply that CTTN might be involved in CRC initiation and progression
by acting as an oncogene.
Aberrant expression of CTTN was associated with
worse prognosis in esophageal cancer and laryngeal premalignancy
(12,15). Our immunohistochemical analysis
showed an increase of CTTN protein levels in CRC tumors compared
with the adjacent non-tumor tissues, which agreed with a previous
study (16). In addition, we found
that high CTTN expression had poor prognosis, which further
reinforced that CTTN played a significant role in the progress of
CRC. In order to understand the connection between CTTN and CRC
cell invasion in more detail, it was necessary to identify genes by
which DJ-1 regulated invasion. DOCK1, a guanine nucleotide exchange
factor, has been reported to promote cell proliferation, motility
and survival by activating Rac1 (18,24).
DOCK1 also interacted with ELMO1 and Gαi2 regulated the actin
cytoskeleton enhanced tumor metastasis in breast cancer (25). In the present study, our data
suggested that invasion by CTTN CRC cells is dependent of the
activation of DOCK1-Rac1, which was confirmed by inhibition of
DOCK1 abolishing the effects of CTTN on migration and invasion.
However, the expression and roles of DOCK1 require further study in
CRC.
In summary, our data indicated that the level of
CTTN expression correlates with clinicopathological features and
patient survival in CRC and contributed to invasion and metastasis
of CRC in vitro. Furthermore, our findings emphasize the
potential role of DOCK1 in CTTN-mediated cell migration and
invasion. Our findings may shed new light on the prognostic markers
for CRC and novel therapeutic targets for CRC invasion
intervention.
Acknowledgments
This study was supported by grants from the Nature
Science Foundation of China (no. 81272751).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu Z, Zhang Y, Franzin L, Cormier JN,
Chan W, Xu H and Du XL: Trends and variations in breast and
colorectal cancer incidence from 1995 to 2011: A comparative study
between Texas Cancer Registry and National Cancer Institute's
Surveillance, Epidemiology and End Results data. Int J Oncol.
46:1819–1826. 2015.PubMed/NCBI
|
|
3
|
Jemal A, Center MM, DeSantis C and Ward
EM: Global patterns of cancer incidence and mortality rates and
trends. Cancer Epidemiol Biomarkers Prev. 19:1893–1907. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ramasamy TS, Ayob AZ, Myint HH,
Thiagarajah S and Amini F: Targeting colorectal cancer stem cells
using curcumin and curcumin analogues: Insights into the mechanism
of the therapeutic efficacy. Cancer Cell Int. 15:962015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kang H, O'Connell JB, Maggard MA, Sack J
and Ko CY: A 10-year outcomes evaluation of mucinous and
signet-ring cell carcinoma of the colon and rectum. Dis Colon
Rectum. 48:1161–1168. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cunningham D, Atkin W, Lenz HJ, Lynch HT,
Minsky B, Nordlinger B and Starling N: Colorectal cancer. Lancet.
375:1030–1047. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ni B, Yu X, Guo X, Fan X, Yang Z, Wu P,
Yuan Z, Deng Y, Wang J, Chen D, et al: Increased urothelial cancer
associated 1 is associated with tumor proliferation and metastasis
and predicts poor prognosis in colorectal cancer. Int J Oncol.
47:1329–1338. 2015.PubMed/NCBI
|
|
8
|
Gupta GP and Massagué J: Cancer
metastasis: Building a framework. Cell. 127:679–695. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tsai JH and Yang J: Epithelial-mesenchymal
plasticity in carcinoma metastasis. Genes Dev. 27:2192–2206. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liao MY, Kuo MY, Lu TY, Wang YP and Wu HC:
Generation of an anti-EpCAM antibody and epigenetic regulation of
EpCAM in colorectal cancer. Int J Oncol. 46:1788–1800.
2015.PubMed/NCBI
|
|
11
|
Weaver AM: Cortactin in tumor
invasiveness. Cancer Lett. 265:157–166. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rodrigo JP, Álvarez-Alija G, Menéndez ST,
Mancebo G, Allonca E, García-Carracedo D, Fresno MF, Suárez C and
García-Pedrero JM: Cortactin and focal adhesion kinase as
predictors of cancer risk in patients with laryngeal premalignancy.
Cancer Prev Res (Phila). 4:1333–1341. 2011. View Article : Google Scholar
|
|
13
|
Weed SA, Karginov AV, Schafer DA, Weaver
AM, Kinley AW, Cooper JA and Parsons JT: Cortactin localization to
sites of actin assembly in lamellipodia requires interactions with
F-actin and the Arp2/3 complex. J Cell Biol. 151:29–40. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bryce NS, Clark ES, Leysath JL, Currie JD,
Webb DJ and Weaver AM: Cortactin promotes cell motility by
enhancing lamellipodial persistence. Curr Biol. 15:1276–1285. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hill A, McFarlane S, Mulligan K, Gillespie
H, Draffin JE, Trimble A, Ouhtit A, Johnston PG, Harkin DP,
McCormick D, et al: Cortactin underpins CD44-promoted invasion and
adhesion of breast cancer cells to bone marrow endothelial cells.
Oncogene. 25:6079–6091. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hirakawa H, Shibata K and Nakayama T:
Localization of cortactin is associated with colorectal cancer
development. Int J Oncol. 35:1271–1276. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chi Y, Huang S, Liu M, Guo L, Shen X and
Wu J: Cyclin D3 predicts disease-free survival in breast cancer.
Cancer Cell Int. 15:892015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Feng H, Hu B, Liu KW, Li Y, Lu X, Cheng T,
Yiin JJ, Lu S, Keezer S, Fenton T, et al: Activation of Rac1 by
Src-dependent phosphorylation of Dock180(Y1811) mediates
PDGFRα-stimulated glioma tumorigenesis in mice and humans. J Clin
Invest. 121:4670–4684. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li H, Yang L, Fu H, Yan J, Wang Y, Guo H,
Hao X, Xu X, Jin T and Zhang N: Association between Gαi2 and
ELMO1/Dock180 connects chemokine signalling with Rac activation and
metastasis. Nat Commun. 4:17062013. View Article : Google Scholar
|
|
20
|
Jing F, Kim HJ, Kim CH, Kim YJ, Lee JH and
Kim HR: Colon cancer stem cell markers CD44 and CD133 in patients
with colorectal cancer and synchronous hepatic metastases. Int J
Oncol. 46:1582–1588. 2015.PubMed/NCBI
|
|
21
|
Luo ML, Shen XM, Zhang Y, Wei F, Xu X, Cai
Y, Zhang X, Sun YT, Zhan QM, Wu M, et al: Amplification and
overexpression of CTTN (EMS1) contribute to the metastasis of
esophageal squamous cell carcinoma by promoting cell migration and
anoikis resistance. Cancer Res. 66:11690–11699. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee MS, Kim S, Kim BG, Won C, Nam SH, Kang
S, Kim HJ, Kang M, Ryu J, Song HE, et al: Snail1 induced in breast
cancer cells in 3D collagen I gel environment suppresses cortactin
and impairs effective invadopodia formation. Biochim Biophys Acta.
1843:2037–2054. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kozyreva VK, McLaughlin SL, Livengood RH,
Calkins RA, Kelley LC, Rajulapati A, Ice RJ, Smolkin MB, Weed SA
and Pugacheva EN: NEDD9 regulates actin dynamics through cortactin
deacetylation in an AURKA/HDAC6-dependent manner. Mol Cancer Res.
12:681–693. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Côté JF and Vuori K: GEF what? Dock180 and
related proteins help Rac to polarize cells in new ways. Trends
Cell Biol. 17:383–393. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Laurin M, Huber J, Pelletier A, Houalla T,
Park M, Fukui Y, Haibe-Kains B, Muller WJ and Côté JF: Rac-specific
guanine nucleotide exchange factor DOCK1 is a critical regulator of
HER2-mediated breast cancer metastasis. Proc Natl Acad Sci USA.
110:7434–7439. 2013. View Article : Google Scholar : PubMed/NCBI
|