Introduction
Cholangiocarcinoma (CCA) is one of the highly
aggressive malignant tumors, arising from varying locations within
ductular epithelium of biliary tree (1,2). As
the second most common primary hepatobiliary malignancy, CCA has
very low postoperative 5-year survival rate, and treatment with
radiotherapy and chemotherapy also carries poor overall survival
rates (1,2). Disappointingly, epidemiologic report
showed that the overall incidence and mortality of CCA seem to be
increasing (3). Hence, defining the
molecular mechanisms of CCA is urgent and required for identifying
early diagnosis and effective chemotherapy markers for CCA
patients.
FBW7 (F-box and WD repeat domain-containing 7), also
known as CDC4, AGO and SEL10, is the substrate recognition
component of an evolutionary conserved SCF (complex of SKP1, CUL1
and F-box protein)-type E3 ubiquitin ligase (4,5). Human
FBW7 gene encodes three transcripts that are produced by
alternative splicing and each mRNA consists of an isoform-specific
first exon linked to ten shared exons, resulting in three protein
isoforms (FBW7α, β and γ) that only vary at the N-terminus
(6). Three FBW7 isoforms have
different subcellular location and tissue expression profile.
FBW7α, β and γ locate in nucleoplasm, cytoplasm, and nucleolus,
respectively (7). FBW7α mRNA is
expressed at much higher levels than either FBW7β or FBW7γ in most
human tissues and varieties of exponentially growing human cells
(6,8). Consistently, adult mouse tissues
express Fbw7α mRNA ubiquitously, while the FBW7β displays higher
levels in the brain and FBW7γ isoform exhibits an increased
expression in muscle tissue (9).
Grim et al utilized gene targeting to create
isoform-specific Fbw7-null mutations in human colon cancer cell
line HCT116 and found that FBW7α was the major isoform that
mediates the stability of cyclin E, c-Myc, and SREBP1 (sterol
regulatory element binding protein 1) (8). These studies suggest that FBW7α may
play more important roles in most physiological and pathological
process than other isoforms.
FBW7 targets multiple well-known oncoproteins
including c-Myc (10), cyclin E
(11), mTOR (12), Mcl-1 (13), Notch-1 (14,15)
and AIB1 (16) for
ubiquitination-mediated destruction, thus, it is a recognized tumor
suppressor (4,5). Consistent with the notion that FBW7 is
a tumor suppressor in various human malignancies, FBW7 mutations
are frequently identified in a variety of human malignancies. The
frequency of FBW7 mutations in all primary human cancers analyzed
was approximately 6% (87/1556). However, the mutation frequency in
cholangiocarcinomas reached up to 35% (7/20) and the report showed
that FBW7 hotspot mutants (4/7) in cholangiocarcinoma not only
reveal defects in localization and substrate binding but also can
abrogate wild-type FBW7 function through a dominant negative
mechanism (17). It has been
reported that the expression of FBW7 was downregulated in tumor
tissues compared with adjacent non-tumorous tissues in intrahepatic
(IHCC; 43 tumor specimens vs. 10 paired intrahepatic bile duct
tissues) and perihilar (PHCC; 64 tumor specimens vs. 10 paired
perihilar bile duct tissues) CCA (18). These results indicate that FBW7 is a
general tumor suppressor in various cancers and it may play more
important inhibitory roles in CCA progression. However, the
function of FBW7 in the proliferation of CCA remains unknown. In
the present study, we report that overexpression of FBW7α inhibited
CCA cell proliferatin in vitro and in vivo. Further
study indicated that FBW7α exerts antitumor activity in CCA at
least in part through downregulating c-Myc and cyclin E.
Materials and methods
Cell culture
CCA cell lines QBC-939 and MZ-cha1 were cultured in
RPMI-1640 (HyClone) supplemented with 10% fetal bovine serum
(Gibco) and 100 U/ml penicillin and 100 mg/ml streptomycin and were
maintained in the humidified incubator with 95% air and 5%
CO2 at 37°C. QBC-939 cells were obtained from Shuguang
Wang (The Third Military Medical University, China); Mz-Cha1 cells
were kindly provided by Dr Yabing Chen (University of Alabama at
Birmingham, Birmingham, AL, USA).
Small interfering RNA and cell
transfection
p21CIP/WAF1 siRNA
(CUUCGACUUUGUCACCGAGdTdT) (19),
FBW7α siRNA-1 (GGGCAACAACGACGCCGAAdTdT) (20), FBW7α siRNA-2:
(GUGAAGUUGUUGGAGUAGAdTdT) (21) and
nonspecific siRNA were purchased from Invitrogen. siRNAs were
transfected with X-tremeGENE transfection reagent to knock down
p21CIP/WAF1 or FBW7α expression in QBC-933 cells
following the manufacturer's instructions.
Generation of lentiviruses and
infection
The cDNA encoding FBW7α, c-Myc or cyclin E were
cloned in the lentiviral vector pLVCS2.0. Oligonucleotide encoding
shRNA targeting c-Myc or cyclin E or control oligonucleotide was
cloned in the lentiviral vector GV112. The generation of lentivirus
vectors was performed by co-transfecting pLV-CS2.0, GV112 or
pLV-CS2.0/GV112 carrying the expression cassette with helper
plasmids pVSV-G and pHR into 293T cells. The viral supernatant was
collected 48 h after transfection and CCA cells were infected with
viral supernatants containing 10 µg/ml Polybrene for 24 h and then
fresh medium was added to the infected cells. The shRNA target
sequences are listed as follows: the target sequence for c-myc
shRNA-1: CAAGGTA GTTATCCTTAAA; the target sequence for c-Myc
shRNA-2: GTTGAAACACAAACTTGAA; the target sequence for cyclin E
shRNA-1: ACATAGAGAACTGTGTCAA; the target sequence for cyclin E
shRNA-2: AATTCTTCTGGATTGG TTA; the target sequence for control
shRNA: TTCTCCGA ACGTGTCACGT.
Western blot analysis
Briefly, Equal amounts of protein lysates were
separated by SDS-PAGE and transferred onto PVDF membranes. After
incubation with the primary antibody, the membranes were extensive
washed by TBST buffer, then were incubated with horseradish
peroxidase conjugated secondary antibody and visualized by
chemiluminescence. Antibodies for FBW7 and c-Myc were purchased
from Abcam; Antibodies for p21CIP/WAF1, mTOR, AIB1 and
Notch1 were purchased from Cell Signaling Technology; Antibodies
for cyclin E and Mcl-1 were purchased from Santa Cruz
Biotechnology; Anti-β-actin antibody was purchased from Sigma.
Real-time RT-PCR
Briefly, total RNA was isolated with TRIzol reagent
(Invitrogen) according to the manufacturer's instructions. Reverse
transcription was performed using Revertra Ace qCR RT Master mix
(Toyobo) with random primer and olig dT primer. Real-time PCR
reactions were performed using FastStart Universal SYBR Green
Master (Roche). Relative quantification was achieved by
normalization to the amount of GAPDH. The primers used for
real-time PCR are listed as follow: FBW7α forward (22): GGAGATGG ACCAGGAGAGTG; FBW7α reverse:
GTTGGTGTTGC TGAACATGG; c-Myc forward: GCTGCTTAGACGCTG GATTT; c-Myc
reverse: CACCGAGTCGTAGTCGAGGT; cyclin E forward:
CTCCAGGAAGAGGAAGGCAA; cyclin E reverse: TCGATTTTGGCCATTTCTTCA;
p21CIP/WAF1 forward: CAGGGGAGCAGGCTGAAG;
p21CIP/WAF1 reverse: GGATTAGGGCTTCCTCTTGG; GAPDH
forward: CAC TCCTCCACCTTTGACGC; GAPDH reverse: TGCTGTAGC
CAAATTCGTTGT.
CHX (cycloheximide) treatment
CCA cells were treated with CHX (60 µg/ml) for
indicated times in the absence or presence of FBW7α overexpression.
Total protein was isolated at different times after CHX treatment,
and the levels of c-Myc and cyclin E protein were determined by
western blot analysis. The band density of c-Myc and cyclin E
protein was quantified by using Scion Imaging software and
normalized to β-actin levels.
MTT assay
CCA cell proliferation was analyzed by MTT assay. A
total of 3×103 QBC-939 or MZ-cha1 cells were seeded in
96-well plates and MTT was added to each well every 24 h. The
plates were incubated for 4 h before addition of solubilization
solution (10% SDS in 0.01 M HCl). The absorbance was measured at
560 nm using a microplate reader.
Focus formation assay
Five hundred cells were cultured in six-well plates
in RPMI-1640 with 10% FBS. Cells were grown for 2–3 weeks, then
colonies were stained with 0.05% crystal violet for 30 min and
counted.
Cell cycle analysis
For cell cycle analysis, 4×105 CCA cells
were synchronized by serum starvation for 24 h and induced to
re-enter the cell cycle by an exchange of 10% fetal bovine serum
for 9 h. Cells were harvested and fixed in 75% ethanol at 4°C
overnight. Cells were incubated with RNase A at 37°C for 30 min,
and then stained with propidium iodide (PI) at 37°C for 30 min.
Cell cycle was measured by flow cytometry.
Tumor xenograft experiments
After infection by lentivirus, QBC-939 cells were
selected with puromycin for 1 week, and then used to inject into
nude mice. The protocols for the in vivo studies were
approved by Institutional Animal Care and Use Committee of
Laboratory Animal Center of Xiamen University. Male nude mice
(4–6-week-old) were obtained from Laboratory Animal Center of
Xiamen University. Nude mice were injected subcutaneously in both
flanks with 4×106 QBC-939-FBW7α and control cells,
respectively. Ten days after cell injection, the volume of the
tumor was monitored and calculated following the formula: Volume =
length × Width2 ×0.52. After 28 days, tumors were
harvested and weighed, and then were dissected and fixed in 10%
formalin and embedded in paraffin for Ki67 staining. Remaining
tumors were homogenized in RIPA for western blot analysis.
Immunohistochemistry
Slides were soaked in preheated citrate buffer (pH
6.0) and heated in a microwave for 20 min to retrieve antigen.
After cooling, slides were washed with PBS three times, and then
incubated with Ki67 antibody (1:200; Invitrogen) overnight at 4°C.
On the second day, slides were washed with PBS three times, and
then incubated with a horseradish peroxidase-conjugated secondary
antibody for 1 h at room temperature. After washing, DAB reagent
was added to visualize the labeled protein.
Statistical analysis
All data are shown as the mean ± SD from the number
of replicates described in results. The statistically significant
effects between mean values (p<0.05) were assessed with the
two-tailed Student's t-test in SPSS.
Results
Overexpression of FBW7α inhibits CCA
cell proliferation
To investigate the role of FBW7α in CCA cell
proliferation, the FBW7α-encoding lentivirus was used to express
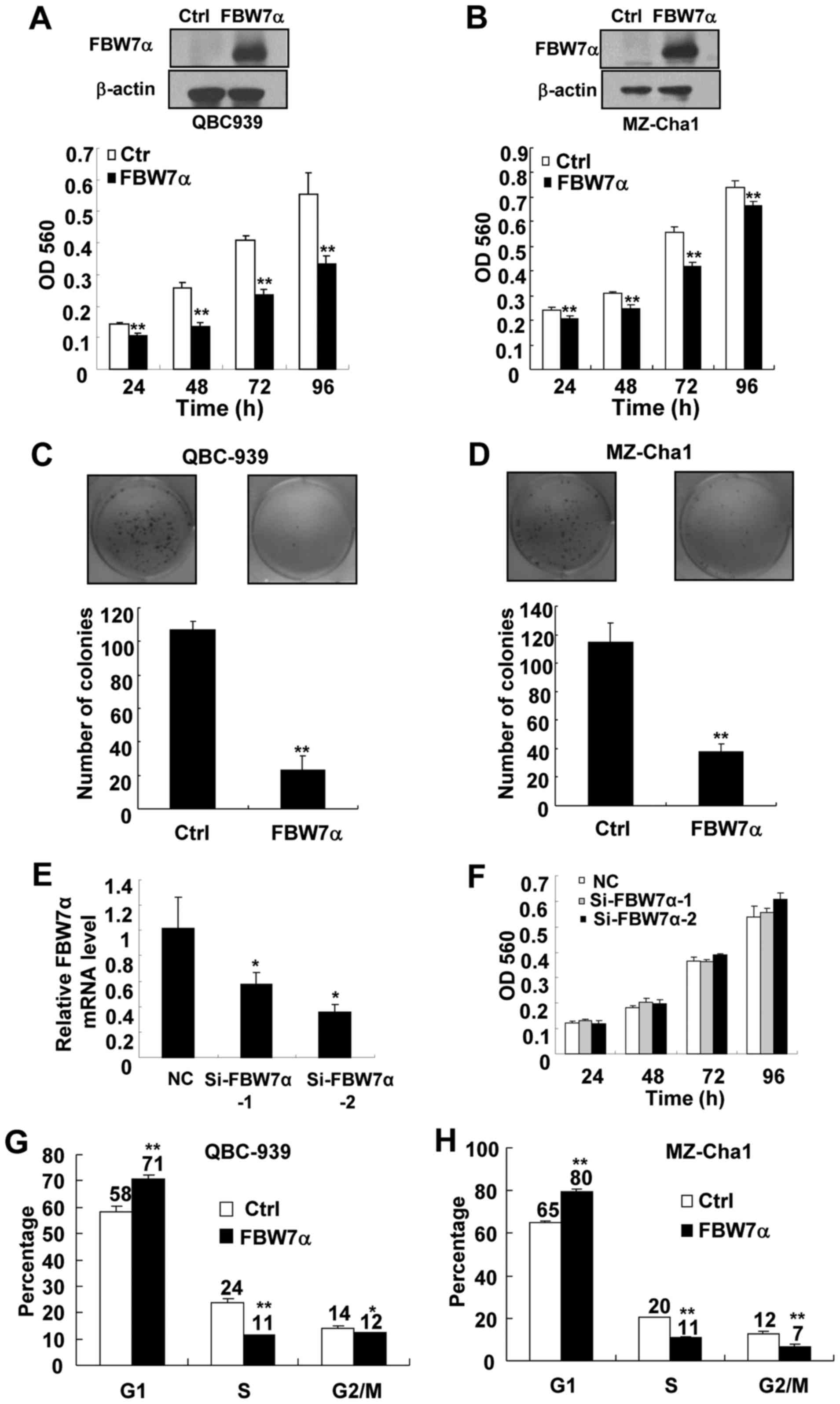
FBW7α in two CCA cell lines, QBC939 and MZ-Cha1, and then cell
proliferation was measured by MTT assay. As shown in Fig. 1A and B, FBW7α-encoding lenivirus,
but not control lentivirus, efficiently increased the levels of the
FBW7α protein in these cells. Significantly, upregulation of FBW7α
inhibited proliferation of QBC939 and MZ-Cha1 cells (Fig. 1A and B). Furthermore, upregulation
of FBW7α decreased focus formations in QBC939 and MZ-Cha1 cells
(Fig. 1C and D). Together, these
results indicate that FBW7α is important for CCA cell
proliferation.
We also observed that the decrease in growth of
FBW7α-overexpression MZ-Cha1 cells was less than
FBW7α-overexpression QBC-939 cells at 96 h (Fig. 1A and B), the reason may be that the
growth of MZ-Cha1 cells is quite nutrition-dependent in culture
medium and nutrition reduction in culture medium after 96-h cell
culture may result in a reduced growth of control cells.
Importantly, our focus forming assay demonstrated that upregulation
of FBW7α significantly decreased focus formations in MZ-Cha1 cells
as QBC-939 (Fig. 1C and D),
suggesting that FBW7α has the same tumor suppressor potential in
the MZ-Cha1 cells and QBC-939 cells.
To investigate the effect of knockdown of FBW7α on
CCA cell proliferation, RNA interference was used to knock down the
expression of FBW7α in QBC-939 cells and the cell proliferation was
measured by MTT assay. Since it is very difficult to detect
endogenous FBW7α protein expression, we use real-time PCR to detect
the level of FBW7α mRNA. As shown in Fig. 1E, FBW7α-specific siRNA, but not
control RNA, efficiently reduced the levels of endogenous FBW7α
mRNA. Although downregulation of FBW7α slightly increased the mean
of OD560 in MTT assay at 96 h, they were not statistically
significant (Fig. 1F). Since the
basal levels of FBW7α protein were too low in CCA cells, knockdown
of FBW7α could not cause significant changes in cell
proliferation.
As upregulation of FBW7α resulted in a decrease in
CCA cell proliferation, cell cycle analysis was performed to detect
whether overexpression of FBW7α induces cell arrest in a specific
phase of the cell cycle. The results showed that the percentage of
FBW7α-overexpression cells at G1 phase was significantly increased
as compared with control cells, and this was associated with a
concomitant decrease of cells at the S and G2/M of the cell cycle
(Fig. 1G and H). These results
indicate that upregulation of FBW7α induces CCA cell arrest in the
G1 phase of the cell cycle, which is at least in part responsible
for the suppression of cell proliferation by FBW7α.
FBW7α promotes the protein degradation
of c-Myc and cyclin E
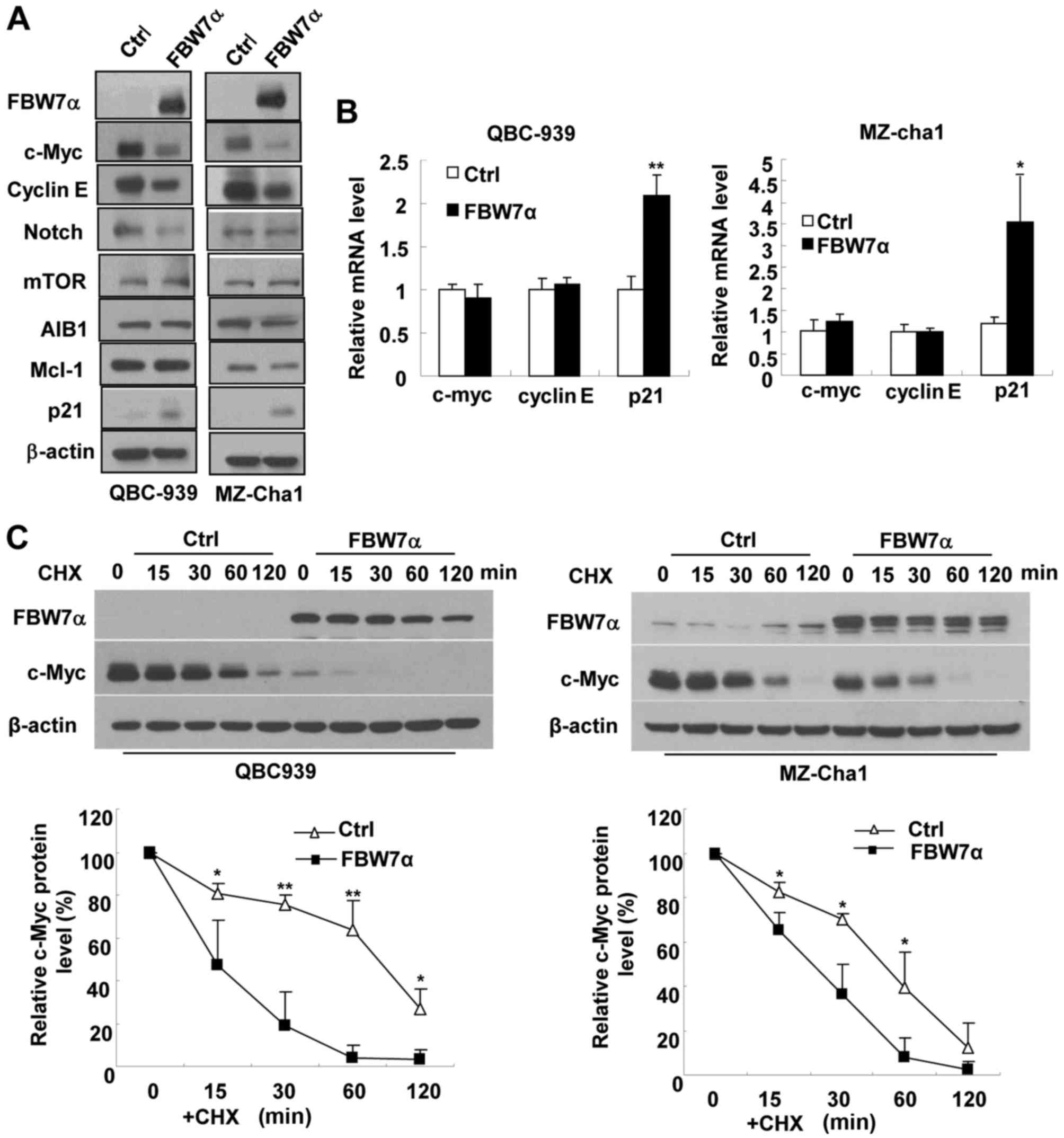
To explore the molecular mechanism underlying the
crucial role of FBW7α in control of CCA cell proliferation, we
examined the effect of FBW7α overexpression on the expression of
several cell proliferation/survival-related proteins which are
identified as the substrates of FBW7 (10–16).
As shown in Fig. 2A, overexpression
of FBW7α significantly reduced the protein levels of c-Myc and
cyclin E, but had no significant effect on the protein levels of
Notch, mTOR, AIB1 and Mcl-1 in either QBC939 or MZ-Cha1 cells.
Overexpression of FBW7α did not affect the mRNA levels of c-Myc and
cyclin E (Fig. 2B), indicating that
FBW7α decreases the expression of c-Myc and cyclin E at the
post-transcriptional level in CCA cells. It has been reported that
c-Myc can inhibit cell cycle inhibitor p21CIP/WAF1
expression at transcriptional level (23,24).
Our results showed that overexpression of FBW7α increased
p21CIP/WAF1 expression at both protein and mRNA levels
(Fig. 2A and 2B), and
downregulation of c-Myc increased p21CIP/WAF1 expression
at both protein and mRNA levels (Fig.
3A) in CCA cells, indicating that FBW7α increases cell cycle
inhibitor p21CIP/WAF1 expression at least in part
through downregulating c-Myc expression.
To determine whether FBW7α accelerates the
degradation of c-Myc and cyclin E proteins, we infected QBC939 and
MZ-Cha1 cells with control lentivirus or FBW7α-encoding lentivirus,
and then used cycloheximide (CHX) to block protein synthesis.
Overexpression of FBW7α significantly accelerated the degradation
of c-Myc and cyclin E proteins when CHX blocked protein synthesis
(Fig. 2C and D), but the effect of
FBW7α on the degradation of c-Myc and cyclin E proteins was
suppressed by the proteasome inhibitor MG132 (Fig. 2E). These results indicate that FBW7α
inhibits c-Myc and cyclin E protein expression dependent on
ubiquitin-proteasome degradation pathway in CCA cells. This notion
is consistent with previous reports in other cells (10,11).
Downregulation of either c-Myc or
cyclin E inhibits CCA cell proliferation
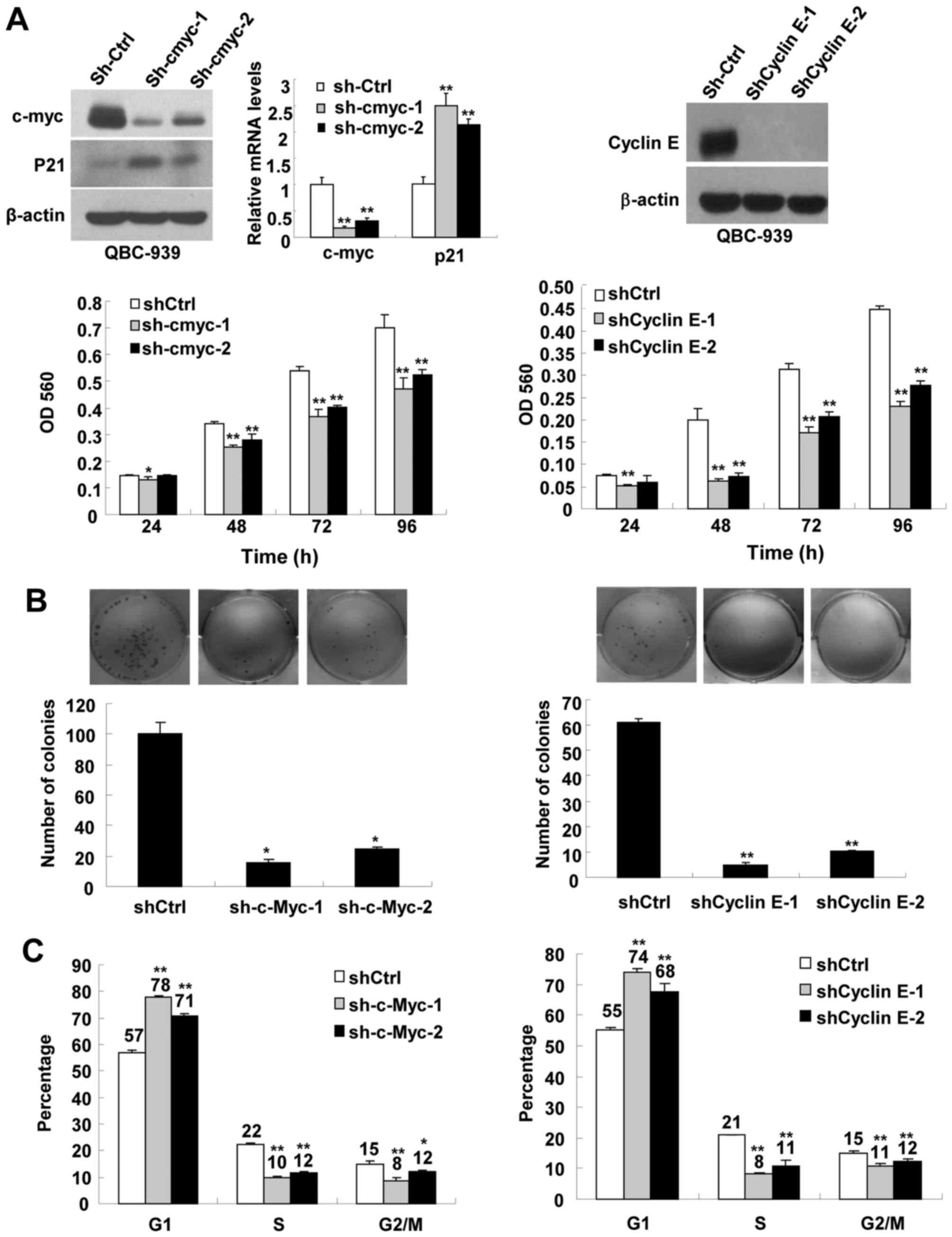
To determine whether FBW7α inhibits CCA
proliferation through promoting c-Myc or cyclin E protein
degradation, the effect of downregulation of c-Myc and cyclin E on
CCA proliferation were investigated. We respectively used two
different shRNA to knock down c-Myc and cyclin E in QBC-939 cells,
and then measured cell proliferation, focus formation and cell
cycle progression. As shown in Fig.
3A, sh-c-Myc and shCyclin E efficiently knocked down the
expression of c-Myc and cyclin E, and downregulation of c-Myc and
cyclin E significantly inhibited CCA cells proliferation,
respectively. Downregulation of c-Myc and cyclin E also
significantly inhibited focus formation (Fig. 3B), and induced cell cycle arrest in
G1 phase (Fig. 3C). These results
indicate that downregulation of c-Myc and cyclin E indeed
contribute to the inhibitory effect of FBW7α on CCA
proliferation.
Restoration of the expression of
c-Myc, but not cyclin E, partially rescues proliferation of
FBW7α-overexpressing CCA cells
To further determine whether FBW7α inhibits CCA
proliferation through promoting c-Myc or cyclin E protein
degradation, FBW7α-overexpressing cells were infected with
c-Myc-encoding lentivirus and cyclin E-encoding lentivirus to
restore the expression of c-Myc and cyclin E, and then cell
proliferation was measured by MTT assay, respectively. Consistent
with the notion of Fig. 2 that
FBW7α could promote c-Myc and cyclin E protein degradation in CCA
cells, overexpression of FBW7α promoted degradation of exogenous
c-Myc (compare lanes 2 with lanes 5) or cyclin E (compare lanes 3
with lanes 6) protein (Fig. 4A).
Although, FBW7α could degrade exogenous c-Myc and cyclin E
proteins, as shown in Fig. 4B,
infection with c-Myc-encoding lentivirus and cyclin E-encoding
lentivirus efficiently restored the expression of c-Myc and cyclin
E in FBW7α-overexpression cells. Restoration of c-Myc expression,
but not cyclin E expression, suppressed FBW7α-medicated
upregulation of the cell cycle inhibitor p21CIP/WAF1 in
FBW7α-overexpression cells (Fig.
4B). Restoration of c-Myc expression partially rescued the
proliferation of FBW7α-overexpression cells as expected (Fig. 4C). Surprisingly, restoration of
cyclin E expression failed to rescue the proliferation of
FBW7α-overexpression cells (Fig.
4C). It may be due to relatively high levels of
p21CIP/WAF1 in FBW7α-overexpression cells (Fig. 4B), which counteracts the promoting
effect of cyclin E on cell proliferation. This notion was supported
by the observation that knockdown of p21CIP/WAF1
partially restored the proliferation of FBW7α-overexpression cells
(Fig. 4D and E).
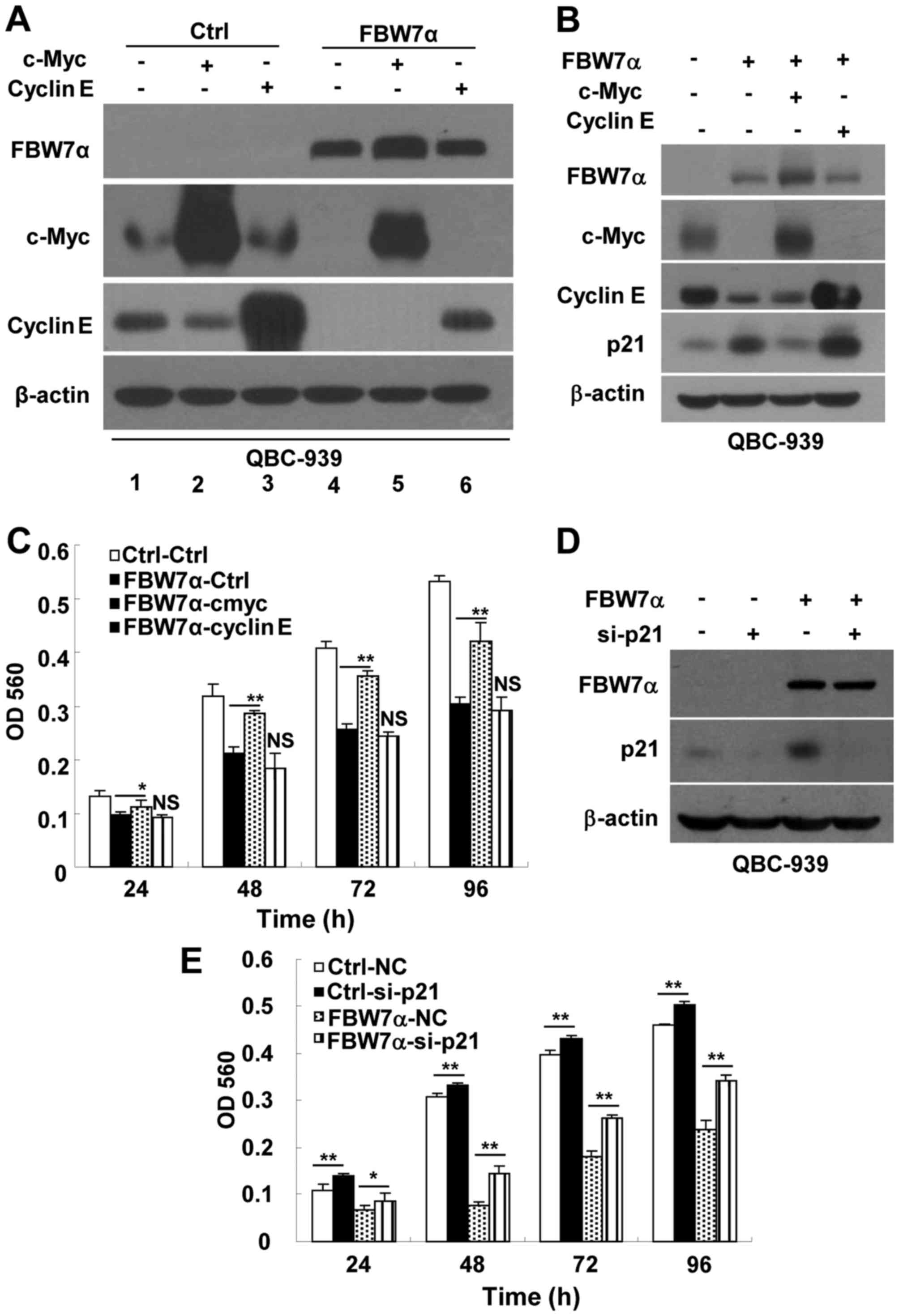 | Figure 4.Restoration of the expression of
c-Myc, but not cyclin E, partially rescues the proliferation of
FBW7α-overexpressing CCA cells. (A) QBC-939 cells were infected
with control or FBW7α-encoding lentivirus for 48 h, then control
cells and FBW7α-overexpressing cells were infected with control,
c-Myc-encoding or cyclin E-encoding lentivirus for 24 h,
respectively. Then these cells were seeded to perform western blot
analysis after 24 h. (B and C) After QBC-939 cells were infected
with control or FBW7α-encoding lentivirus for 48 h, the cells were
infected with control, c-Myc-encoding or cyclin E-encoding
lentivirus for 24 h. Then, QBC-939 cells were seeded to perform
western blot analysis (B) and MTT assay (C). (D and E) Knockdown of
p21CIP/WAF1 partially restored the proliferation of
FBW7α-overexpressing cells. After QBC-939 cells were infected with
control or FBW7α-encoding lentivirus for 48 h, these cells were
transfected with control siRNA or p21CIP/WAF1-specific
siRNA for 24 h. Then, QBC-939 cells were seeded to perform western
blot analysis (D) and MTT assay (E). Ctrl represents the infection
with pLV-CS 2.0-encoding lentivirus; FBW7α represents the infection
with pLV-CS2.0-FBW7α-encoding lentivirus; NC represents negtive
control for si-p21; si-p21 represents p21-specific siRNA. All data
are mean ± SD, n=3, *p <0.05, **p<0.01. |
Overexpression of FBW7α inhibits CCA
tumorigenesis in nude mice
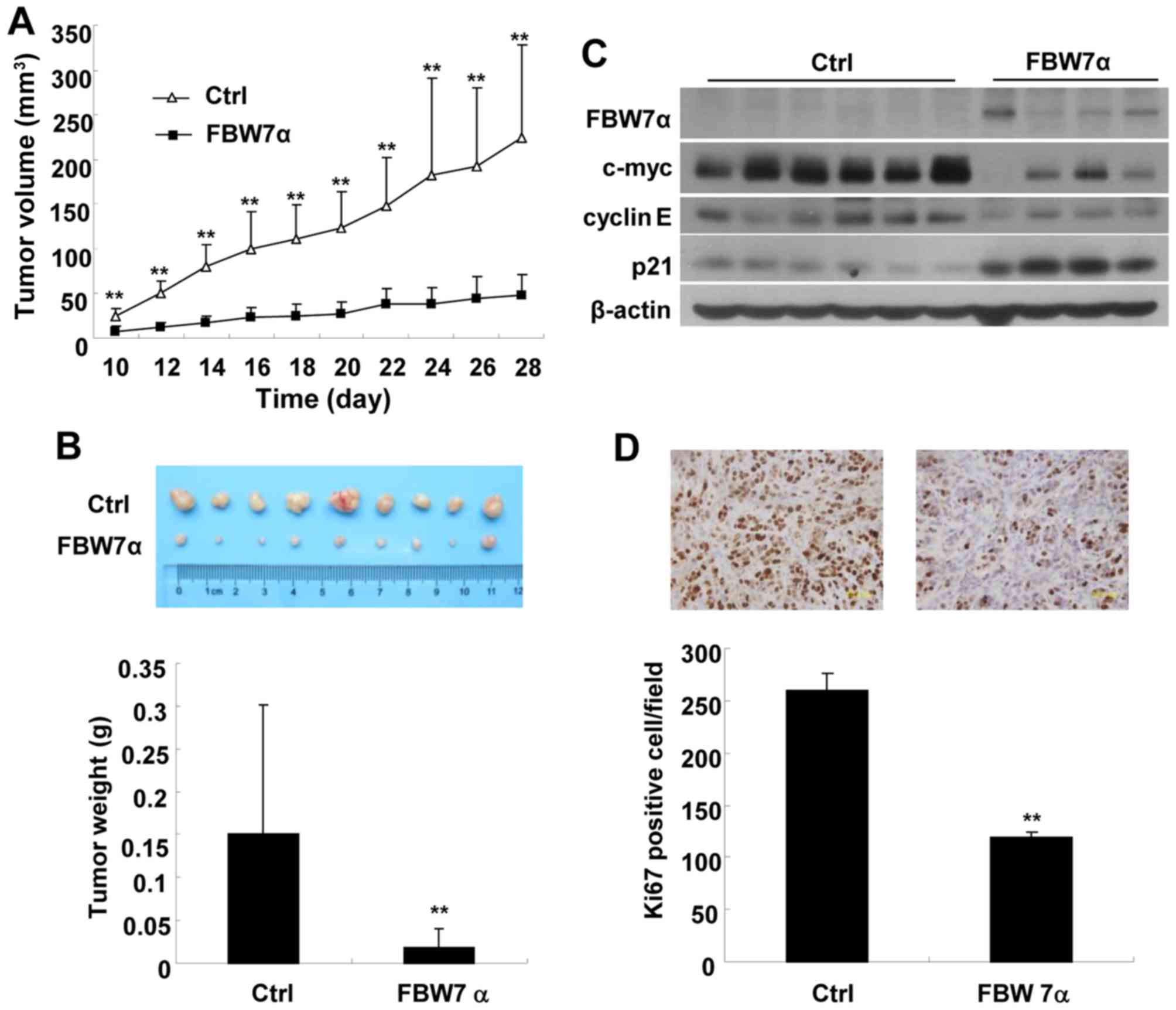
To invistigate the role of FBW7α in CCA progression
in vivo, we examined the effect of FBW7α overexpression on
the growth of CCA xenograft tumors in nude mice injected with
FBW7α-overexpression or control QBC939 cells. As shown in Fig. 5A, FBW7α-overexpression CCA tumors
grew much slower than control tumors. At the end of study (day 28),
tumor weight of FBW7α-overexpression group (0.0223±0.0185 g) was
only 15% of the control group (0.151±0.0728 g) (Fig. 5B). Western blot results showed that
the protein levels of c-Myc and cyclin E in FBW7α-overexpression
CCA tumors were much lower than that of control tumors, whereas
p21CIP/WAF1 expression was much higher (Fig. 5C). Furthermore, immunohistochemical
results showed that FBW7α-overexpression CCA tumors had much less
Ki67-positive cells (Ki-67 is known as a marker of cell
proliferation) than control tumors (Fig. 5D), indicating that overexpression of
FBW7α inhibits CCA tumor cell proliferation in vivo.
Collectively, these results demonstrate that FBW7α is able to
inhibit CCA tumor growth in nude mice.
Discussion
The mutation rates of FBW7 are relatively tumor
type-dependent. Cholangiocarcinoma and T-cell acute lymphocytic
leukemia harbor 35 and 31% frequency of mutations for FBW7,
respectively, while more than ten other types of tumors (such as
stomach, colon, breast, bladder, endometrium) only harbor 0–9%
frequency of mutations (17).
Immunohistochemistry staining showed that the expression of FBW7
was downregulated in tumor tissues compared with adjacent
non-tumorous tissues in intrahepatic (IHCC) and perihilar (PHCC)
CCA (18). The relatively high
frequency of impaired FBW7 function in CCA implicate that FBW7 may
play a specific role in CCA progression. In the present study, we
defined the role of FBW7 in CCA cell proliferation. Our results
showed that overexpression of FBW7α significantly inhibited CCA
cell proliferation and CCA xenograft tumor growth, suggesting that
FBW7α plays a tumor suppressor role in CCA progression.
Several oncogenic proteins, such as c-Myc, c-Jun,
cyclin E, mTOR, Notch-1, Notch-3, and AIB1, have been identified as
the substrates of FBW7 (4,5). However, FBW7 targets different
substrates for degradation in a cell type-specific manner and the
role of FBW7 in cell proliferation may depend on cell and signaling
context. For example, conditional deletion of FBW7 in mouse
keratinocytes significantly increased the protein levels of Notch1
and c-Myc, but not c-Jun, Notch3 and cyclin E (25). FBW7 deletion in mouse thymocytes
significantly increased the expression of Notch1, Notch3 and c-Myc,
but not cyclin E (26). Although
conditional deletion of FBW7 in hematopoietic stem cells,
keratinocytes or T cells promoted cell proliferation (25–27),
FBW7 deletion inhibited cell proliferation in mouse embryonic
fibroblasts (28,29). In the present study, we found that
the decrease in the amounts of c-Myc and cyclin E (rather than
mTOR, Notch, AIB1 and Mcl-1) in FBW7α-overexpression CCA cells were
the most pronounced and reproducible in different CCA cell lines,
suggesting that downregulation of c-Myc and cyclin E may mediate
the inhibitory effect of FBW7α in CCA cell proliferation. This
notion is supported by the observation that knockdown of c-Myc or
cyclin E significantly inhibited CCA cell proliferation.
Noteworthy, while restoration of c-Myc expression in
FBW7α-overexpressing CCA cells partially rescued the cell
proliferation, restoration of cyclin E expression in
FBW7α-overexpressing CCA cells failed to rescue the cell
proliferation. Restoration of c-Myc expression in
FBW7α-overexpression CCA cells could significantly downregulate
p21CIP/WAF1 expression, but restoration of cyclin E
expression in FBW7α-overexpression CCA cells could not downregulate
p21CIP/WAF1 expression. These results suggest that high
level of p21CIP/WAF1 could abolish the promotive effect
of cyclin E on proliferation in cyclin E/FBW7α-overexpression CCA
cells. Downregulation of p21CIP/WAF1 in
FBW7α-overexpressing CCA cells could partially rescue the cell
proliferation.
Our study demonstrated that FBW7α plays an essential
inhibitory role in CCA progression, suggesting that targeting FBW7α
pathway becomes an attracting strategy in CCA treatment. Our
results showed that restoration of the expression of c-Myc
partially rescues proliferation of FBW7α-overexpressing CCA cells,
suggesting that downregulation of c-Myc is partially responsible
for FBW7α-induced inhibition of CCA cell proliferation. Consistent
with our results, cre-mediated ablation of c-Myc (but not
inactivation of notch signaling) prevented the stimulatory effect
of FBW7 ablation on keratinocyte or T cell proliferation in mice
(25,26), suggesting that FBW7-c-Myc axis is
important for controlling cell proliferation. The expression of
FBW7α substrate c-Myc was induced in different animal models of CCA
and promoted CCA progression (30–32),
suggesting that c-Myc plays an important role in CCA
progression.
It has been reported that FBW7 hotspot mutants in
human CCA not only reveal defects in localization and substrate
binding, but also can abrogate wild-type FBW7 function through a
dominant negative mechanism (17).
Since mutated endogenous FBW7 proteins are not only functionally
inactivated, but also impair wild-type FBW7 function, it is not a
good strategy to target FBW7 directly for CCA treatment. Given that
our results demonstrate that downregulation of c-Myc expression
plays an important role in inhibition of proliferation of
FBW7α-overexpression CCA cells, therefore, targeting FBW7α
substrate c-Myc with a pharmacological c-Myc inhibitor may be a
viable strategy for CCA treatment. It has been reported that a
small molecular c-Myc inhibitor 10058-F4 could induce
p21CIP/WAF1 expression, inhibit tumor cell proliferation
and enhance chemosensitivity of human hepatocellular carcinoma
cells and acute myeloid leukemia (33,34).
Therefore, c-Myc inhibitor may represent a promising therapeutic
approach for CCA with FBW7 mutated or downregulated.
Acknowledgements
This work was supported by grants from the Natural
Science Foundation of China (no. 81272246 to W.L.), the Xiamen
Science and Technology Plan (no. 3502Z20154037 to M.L.) and Natural
Science Foundation of Fujian Province (no. 2015J01502 to Z.Z.).
References
|
1
|
Seeree P, Pearngam P, Kumkate S and
Janvilisri T: An omics perspective on molecular biomarkers for
diagnosis, prognosis, and therapeutics of cholangiocarcinoma. Int J
Genomics. 2015:1795282015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Razumilava N and Gores GJ:
Cholangiocarcinoma. Lancet. 383:2168–2179. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bergquist A and von Seth E: Epidemiology
of cholangiocarcinoma. Best Pract Res Clin Gastroenterol.
29:221–232. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Welcker M and Clurman BE: FBW7 ubiquitin
ligase: A tumour suppressor at the crossroads of cell division,
growth and differentiation. Nat Rev Cancer. 8:83–93. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang L, Ye X, Liu Y, Wei W and Wang Z:
Aberrant regulation of FBW7 in cancer. Oncotarget. 5:2000–2015.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Spruck CH, Strohmaier H, Sangfelt O,
Müller HM, Hubalek M, Müller-Holzner E, Marth C, Widschwendter M
and Reed SI: hCDC4 gene mutations in endometrial cancer. Cancer
Res. 62:4535–4539. 2002.PubMed/NCBI
|
|
7
|
Welcker M, Orian A, Grim JE, Eisenman RN
and Clurman BE: A nucleolar isoform of the Fbw7 ubiquitin ligase
regulates c-Myc and cell size. Curr Biol. 14:1852–1857. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Grim JE, Gustafson MP, Hirata RK, Hagar
AC, Swanger J, Welcker M, Hwang HC, Ericsson J, Russell DW and
Clurman BE: Isoform- and cell cycle-dependent substrate degradation
by the Fbw7 ubiquitin ligase. J Cell Biol. 181:913–920. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Matsumoto A, Onoyama I and Nakayama KI:
Expression of mouse Fbxw7 isoforms is regulated in a cell cycle- or
p53-dependent manner. Biochem Biophys Res Commun. 350:114–119.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yada M, Hatakeyama S, Kamura T, Nishiyama
M, Tsunematsu R, Imaki H, Ishida N, Okumura F, Nakayama K and
Nakayama KI: Phosphorylation-dependent degradation of c-Myc is
mediated by the F-box protein Fbw7. EMBO J. 23:2116–2125. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Koepp DM, Schaefer LK, Ye X, Keyomarsi K,
Chu C, Harper JW and Elledge SJ: Phosphorylation-dependent
ubiquitination of cyclin E by the SCFFbw7 ubiquitin ligase.
Science. 294:173–177. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mao JH, Kim IJ, Wu D, Climent J, Kang HC,
DelRosario R and Balmain A: FBXW7 targets mTOR for degradation and
cooperates with PTEN in tumor suppression. Science. 321:1499–1502.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Inuzuka H, Shaik S, Onoyama I, Gao D,
Tseng A, Maser RS, Zhai B, Wan L, Gutierrez A, Lau AW, et al:
SCF(FBW7) regulates cellular apoptosis by targeting MCL1 for
ubiquitylation and destruction. Nature. 471:104–109. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tsunematsu R, Nakayama K, Oike Y,
Nishiyama M, Ishida N, Hatakeyama S, Bessho Y, Kageyama R, Suda T
and Nakayama KI: Mouse Fbw7/Sel-10/Cdc4 is required for notch
degradation during vascular development. J Biol Chem.
279:9417–9423. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu G, Lyapina S, Das I, Li J, Gurney M,
Pauley A, Chui I, Deshaies RJ and Kitajewski J: SEL-10 is an
inhibitor of notch signaling that targets notch for
ubiquitin-mediated protein degradation. Mol Cell Biol.
21:7403–7415. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu RC, Feng Q, Lonard DM and O'Malley BW:
SRC-3 coactivator functional lifetime is regulated by a
phospho-dependent ubiquitin time clock. Cell. 129:1125–1140. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Akhoondi S, Sun D, von der Lehr N,
Apostolidou S, Klotz K, Maljukova A, Cepeda D, Fiegl H, Dafou D,
Marth C, et al: FBXW7/hCDC4 is a general tumor suppressor in human
cancer. Cancer Res. 67:9006–9012. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang H, Lu X, Liu Z, Chen L, Xu Y, Wang Y,
Wei G and Chen Y: FBXW7 suppresses epithelial-mesenchymal
transition, stemness and metastatic potential of cholangiocarcinoma
cells. Oncotarget. 6:6310–6325. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Barsotti AM and Prives C:
Pro-proliferative FoxM1 is a target of p53-mediated repression.
Oncogene. 28:4295–4305. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang C, Li S, Wang M, Chang AK, Liu Y,
Zhao F, Xiao L, Han L, Wang D, Li S, et al: PTEN suppresses the
oncogenic function of AIB1 through decreasing its protein stability
via mechanism involving Fbw7 alpha. Mol Cancer. 12:212013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
van Drogen F, Sangfelt O, Malyukova A,
Matskova L, Yeh E, Means AR and Reed SI: Ubiquitylation of cyclin E
requires the sequential function of SCF complexes containing
distinct hCdc4 isoforms. Mol Cell. 23:37–48. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang R, Wang Y, Liu N, Ren C, Jiang C,
Zhang K, Yu S, Chen Y, Tang H, Deng Q, et al: FBW7 regulates
endothelial functions by targeting KLF2 for ubiquitination and
degradation. Cell Res. 23:803–819. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gartel AL and Radhakrishnan SK: Lost in
transcription: p21 repression, mechanisms, and consequences. Cancer
Res. 65:3980–3985. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bretones G, Delgado MD and León J: Myc and
cell cycle control. Biochim Biophys Acta. 1849:506–516. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ishikawa Y, Hosogane M, Okuyama R, Aoyama
S, Onoyama I, Nakayama KI and Nakayama K: Opposing functions of
Fbxw7 in keratinocyte growth, differentiation and skin
tumorigenesis mediated through negative regulation of c-Myc and
Notch. Oncogene. 32:1921–1932. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Onoyama I, Tsunematsu R, Matsumoto A,
Kimura T, de A, lborán IM, Nakayama K and Nakayama KI: Conditional
inactivation of Fbxw7 impairs cell-cycle exit during T cell
differentiation and results in lymphomatogenesis. J Exp Med.
204:2875–2888. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Matsuoka S, Oike Y, Onoyama I, Iwama A,
Arai F, Takubo K, Mashimo Y, Oguro H, Nitta E, Ito K, et al: Fbxw7
acts as a critical fail-safe against premature loss of
hematopoietic stem cells and development of T-ALL. Genes Dev.
22:986–991. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ishikawa Y, Onoyama I, Nakayama KI and
Nakayama K: Notch-dependent cell cycle arrest and apoptosis in
mouse embryonic fibroblasts lacking Fbxw7. Oncogene. 27:6164–6174.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Masuda K, Ishikawa Y, Onoyama I, Unno M,
de A, lborán IM, Nakayama KI and Nakayama K: Complex regulation of
cell-cycle inhibitors by Fbxw7 in mouse embryonic fibroblasts.
Oncogene. 29:1798–1809. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang H, Li TW, Ko KS, Xia M and Lu SC:
Switch from Mnt-Max to Myc-Max induces p53 and cyclin D1 expression
and apoptosis during cholestasis in mouse and human hepatocytes.
Hepatology. 49:860–870. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Prakobwong S, Khoontawad J, Yongvanit P,
Pairojkul C, Hiraku Y, Sithithaworn P, Pinlaor P, Aggarwal BB and
Pinlaor S: Curcumin decreases cholangiocarcinogenesis in hamsters
by suppressing inflammation-mediated molecular events related to
multistep carcinogenesis. Int J Cancer. 129:88–100. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang H, Li TW, Peng J, Tang X, Ko KS, Xia
M and Aller MA: A mouse model of cholestasis-associated
cholangiocarcinoma and transcription factors involved in
progression. Gastroenterology. 141:378–388, 388.e1-4. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lin CP, Liu JD, Chow JM, Liu CR and Liu
HE: Small-molecule c-Myc inhibitor, 10058-F4, inhibits
proliferation, downregulates human telomerase reverse transcriptase
and enhances chemosensitivity in human hepatocellular carcinoma
cells. Anticancer Drugs. 18:161–170. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang MJ, Cheng YC, Liu CR, Lin S and Liu
HE: A small-molecule c-Myc inhibitor, 10058-F4, induces cell-cycle
arrest, apoptosis, and myeloid differentiation of human acute
myeloid leukemia. Exp Hematol. 34:1480–1489. 2006. View Article : Google Scholar : PubMed/NCBI
|