Introduction
Colorectal cancer (CRC) is one of the most prevalent
tumors associated with cancer-related death worldwide (1). The development of drug resistance and
relapse with refractory disease, further limits the 5-year survival
of metastatic CRC patients to less than 10% (2). Aberrant alternate splicing commonly
observed in cancer cells, has been implicated in the transformation
of many types of cancers (3,4). For
instance, many genes involved in the proliferation and invasiveness
of cancer cells, are frequently alternatively spliced. These
specific splice variants, which are often upregulated in many
tumors, stimulate cell proliferation and migration and thus,
contribute to the transformed phenotype (5). However, the roles of different spliced
variants along with their splicing factors, involved in controlling
the alternate splicing have not been completely appreciated in
tumor development.
USP39 is one of the deubiquitinating enzymes (DUBs)
which do not have ubiquitin protease activity due to lack of three
important residues important for the protease activity (6). It encodes a conserved protein,
U4/U6·U5 tri-snRNP, which has 65% overall homology with yeast Sad1p
splice factor, and has been implicated in the assembly of mature
spliceosomes. In vitro studies have indicated that Usp39 is
essential for pre-mRNA splicing, but not for the stability of the
spliceosome complex after its formation (7). USP39 has also been shown to be
essential in the spindle assembly checkpoint and controls the
Aurora B mRNA levels in U2OS cells (6). In addition, USP39 mutation in
zebrafish induce G1/S arrest by rb1 splicing defect, and the e2f4
appears to be also the target of USP39. This mutation eventually
contributes to adenohypophyseal sensitivity to rb1 and e2f4, and
lead to pituitary tumorigenesis (8). Similarly, USP39 mutation in breast
cancer cells, also implicate it as an oncogenic factor, and its
downregulation induced apoptosis of the MCF-7 breast tumor cells
(9). A study (10) has further demonstrated that USP39
overexpression enhanced the proliferation of prostate cancer cells,
thereby suggesting that USP39 may also play a key role in prostate
cancer development. All these studies have emphasized that USP39
may be a potential molecular target of cancer.
In the present investigation, we first explored the
relationship between USP39 expression and colorectal cancer.
Initial results suggested that USP39 expression levels were higher
in tumor tissues than in adjacent normal tissues, and thus,
indicated that USP39 may act as a new target in colorectal cancer.
To further verify our hypothesis, we tested USP39 expression in
four different colorectal cancer cells, and later by knocking down
its expression by RNA interference (RNAi) technology in SW480 and
HT29 colorectal cancer cell lines, we investigated its role in
in vitro cell invasion and migration. Our results indicated
that USP39 inhibition suppressed the migration of colorectal cancer
cells through regulation of Wnt/β-catenin pathway.
Materials and methods
Cell culture
Human CRC cell lines, LoVo, Caco2, SW480 and HT29
were purchased from the Shanghai Institutes for Biological Sciences
(Shanghai, China). LoVo cells were cultured in F-12K medium, and
Caco2 cells in minimum essential nedium (MEM), SW480 were cultured
in L15 medium and HT29 cells in McCoys 5A medium. The culture
mediums of three cell lines were supplemented with 10% fetal bovine
serum (FBS), 100 U/ml of penicillin and 100 µg/ml of streptomycin.
The Caco2 cells, instead were cultured in medium with 20% FBS, in
addition to other ingredients. All cell types were incubated at
37°C with 5% CO2.
Immunostaining of tissue microarray
and clinical samples for USP39 expression
Colorectal cancer tissue arrays were purchased from
the National Engineering Center for Biochips (Shanghai, China). In
addition, the surgically excised CRC tissues and surrounding
non-tumor colon tissues were obtained from 9 CRC patients at the
Affiliated Drum Tower Hospital of Nanjing University Medical
School. This study was approved by the Ethics Committee of the
Affiliated Drum Tower Hospital of Nanjing University Medical School
and patient informed consent was obtained. The USP39 expression in
these tissues was evaluated by immunohistochemical staining with
USP39-specific antibody (1:500 dilution; Abcam, Cambridge, UK). The
expression was scored according to the staining intensity and the
percentage of cells stained. The final staining scores were
calculated by multiplying staining intensity with percentage of
stained cells.
MTT assay
Human CRC cells (1×105 in 0.2 ml/well) in
three replicates, were seeded in 96-well plate in the complete
medium and cultured at 37°C for 1–5 days. The 100 µl (5 mg/ml) of
MTT solution was added into each well and incubated at 37°C for 4
h. Next, the supernatant was removed and 150 µl of dimethyl
sulfoxide (DMSO) was added per well. The plate was oscillated for
30 min at room temperature, and later the absorbance at 490 nm was
measured. The values were calculated after background subtraction.
All the MTT experiments were repeated at least three times.
USP39 knockdown
The lenti-shRNA vector system to knock down USP39
was constructed, packed and purified by Shanghai GeneChem, Co.,
Ltd., (Shanghai, China) and was later manipulated according to the
protocol provided by the manufacturer.
Wound healing assay
CRC cells seeded in 6-well plates at a density of
105 cells/well, were incubated for 48 h. A 2-µl pipette
tip was used to scratch a linear wound in the cell monolayer and
cells were then allowed to grow in complete media. Images were
taken at 0 and 48 h after wounding, respectively.
Transwell migration
Cell migration assay was performed using Boyden
chambers (Transwell Costar; 6.5-mm diameter, 8-µm pore size)
according to the manufacturers instructions. Briefly,
3×104 cells were resuspended in 500 µl of serum-free
medium and seeded in the upper chamber. In parallel, 800 µl of the
complete medium containing 10% FBS was added into the lower
chamber. Cells were allowed to migrate for 48 h. Later, migrated
cells were stained with 1% crystal violet and the cell numbers were
counted under a microscope. Six fields of view from each chamber
were randomly selected for cell counting.
Quantitative real-time PCR
Total RNA was extracted from CRC cell lines using
TRIzol (Invitrogen) following the manufacturers instructions.
qRT-PCR was performed as previously descibed (11). Briefly, 1 µg of total RNA was
transcribed using random primers and PrimeScript reverse
transcriptase (Takara Bio, Dalian, China). Quantitative PCR
reaction for USP39 and GAPDH genes was carried out using SYBR-Green
qPCR kit (Takara) on a fluorescent temperature cycler (Mx3000P
real-time PCR system; Stratagene, La Jolla, CA, USA). The following
primers were used to detect the expression of USP39 (F,
5′-CCAGCGATGGCAACTAC-3′ and R, 5′-ACCACAACGGAAACACG-3′) and GAPDH
(F, 5′-TGACTTCAACAGCGACACCCA-3′ and R, 5′-CACCCTGTTGCTGTAGCCA
AA-3′). The PCR reaction was performed with the following
parameters; denaturation at 95°C for 5 min, followed by 45 cycles
of 95°C for 15 sec and 60°C for 1 min. Finally, using GAPDH as an
endogenous control, relative gene expression of USP39 was
determined by comparative ∆Ct method using the Stratagene analysis
software. The experiment was repeated at least three times.
Western blot analysis
CRC cells were lysed in RIPA lysis buffer (Tiangen
Biotech, Co., Ltd., Beijing, China) and extracted proteins (20 µg)
were separated on a 10% SDS-PAGE gel and then transferred to
Immobilon-P polyvinylidene difluoride membrane (PVDF) (Millipore,
Billerica, MA, USA). The expression of different proteins were
visualized by chemiluminescence using ECL-Advance Western blotting
detection kit (Millipore). The antibody dilutions used for USP39
(ab150393; Abcam) were 1:5,000; β-catenin (ab19452; Abcam) 1:1,000;
TCF4 (ab60727; Abcam) 1:1,000; MMP2 (#4022; Cell Signaling
Technology, Hitchin, UK) 1:2,000; MMP9 (#3852; Cell Signaling
Technology) 1:2,000; GAPDH (ab8227; Abcam) 1:7,500; and goat
anti-rabbit IgG H&R (Abcam) 1:6,000.
Statistical analyses
The data shown are presented as the mean ± standard
deviation (SD) of three independent experiments. All statistical
analyses were performed using SPSS 11.0 software (SPSS, Inc.,
Chicago, IL, USA) and P<0.05 was considered statistically
significant.
Results
USP39 expression is upregulated in CRC
tumor tissues and correlates with poor patient overall
survival
The USP39 expression was examined in human CRC
tissue array samples by immunohistochemical staining. The array
included 90 CRC patient tissue samples including cancer and
corresponding adjacent normal tissues. Among these, 48 tissues
samples were from male patients, while 42 were from female
patients, with a median age of 71 years (range, 24–90 years). As
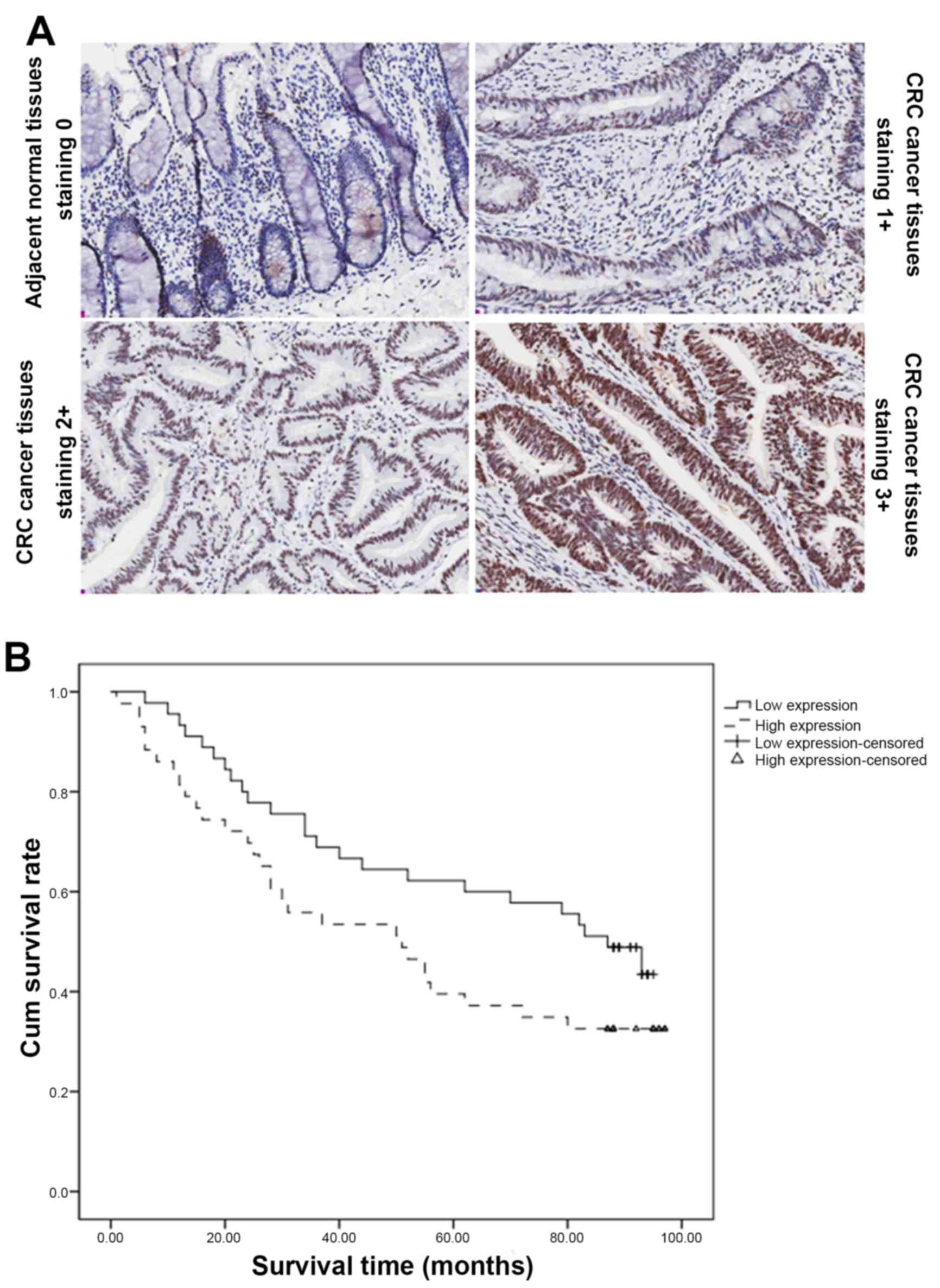
shown in Table I and Fig. 1A, USP39 had strong expression in CRC
tumor tissue samples, as 89 of these showed a score of ≥2. In
contrast, adjacent normal tissue samples displayed dramatically
reduced USP39 expression, compared with cancer tissues
(P<0.001). In addition, USP39 expression was not associated with
other factors. However, Kaplan-Meier survival curves and the
log-rank test survival analysis indicated that CRC patients with
high USP39 expression had significantly poor overall survival
(P<0.05), as shown in Fig. 1B.
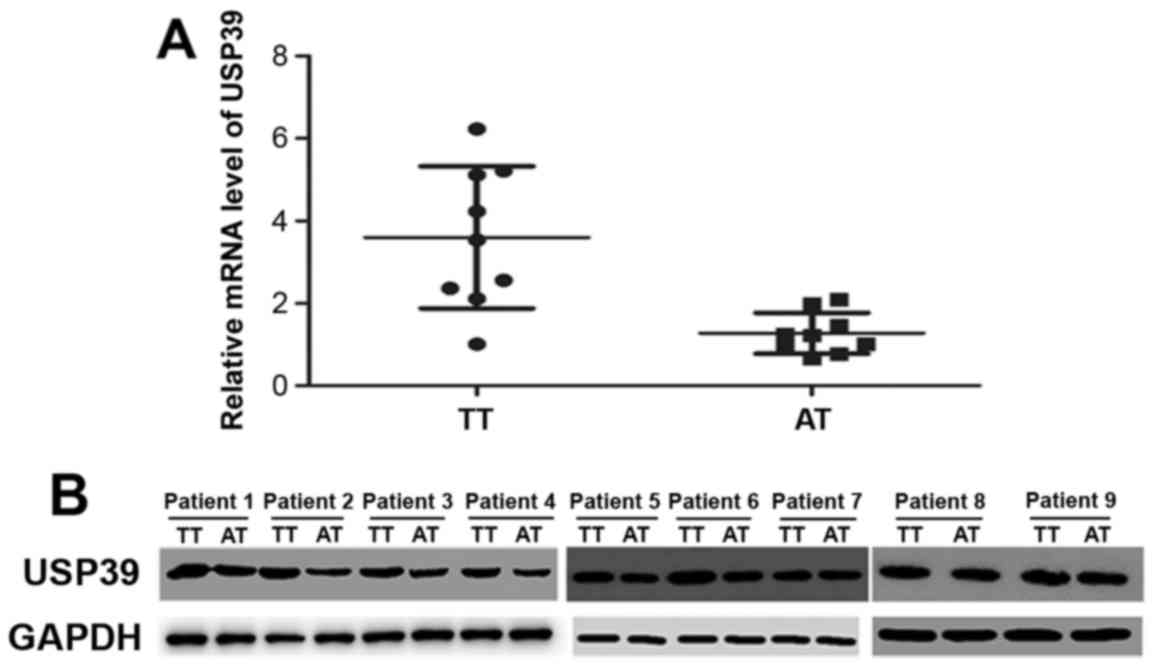
Additionally, the tissue sample analysis of 9 colorectal patients,
also revealed significantly higher mRNA and protein levels of USP39
in tumor tissues than corresponding adjacent normal tissues, as
shown in Fig. 2.
 | Table I.The expression of USP39 in human
colorectal cancer tissue array. |
Table I.
The expression of USP39 in human
colorectal cancer tissue array.
|
|
| USP39 expression |
|
|
|
|---|
|
|
|
|
|
|
|
|---|
| Characteristics | N | 0 | 1 | 2 | 3 | None | Mean rank | Z | P-value |
|---|
| Gender |
|
|
|
|
|
|
|
|
|
| Male | 48 | 0 | 10 | 21 | 16 | 1 | 32.47 | 0.938 | 0.348 |
| Female | 42 | 1 | 14 | 13 | 14 | 0 | 28.74 |
|
|
| Age (years) |
|
|
|
|
|
|
|
|
|
| ≤65 | 34 | 0 | 9 | 12 | 12 | 1 | 31.45 | 0.282 | 0.778 |
| >65 | 56 | 1 | 14 | 23 | 18 | 0 | 30.07 |
|
|
| Tumor size
(cm) |
|
|
|
|
|
|
|
|
|
| ≤5 | 45 | 1 | 12 | 16 | 16 | 0 | 31.13 | −0.005 | 0.996 |
| >5 | 45 | 0 | 12 | 18 | 14 | 1 | 30.27 |
|
|
|
Differentiation |
|
|
|
|
|
|
|
|
|
| I | 8 | 0 | 2 | 5 | 1 | 0 | 23.50 | −0.683 | 0.247 |
| II | 58 | 1 | 15 | 18 | 23 | 1 | 32.25 |
|
|
| III | 24 | 0 | 6 | 11 | 7 | 0 | 29.17 |
|
|
| TNM stage |
|
|
|
|
|
|
|
|
|
| I | 2 | 0 | 0 | 2 | 0 | 0 | 26.00 | −1.308 | 0.095 |
| II | 9 | 0 | 5 | 2 | 2 | 0 | 20.22 |
|
|
| III | 64 | 1 | 16 | 21 | 25 | 1 | 33.00 |
|
|
| IV | 15 | 0 | 3 | 9 | 3 | 0 | 28.00 |
|
|
| Location |
|
|
|
|
|
|
|
|
|
| Tumor tissue | 90 | 1 | 21 | 34 | 33 | 1 |
| 4.285 | 0.000 |
| Adjacent
tissue | 90 | 3 | 52 | 25 | 9 | 1 |
|
|
|
USP39 knockdown inhibits growth of
human CRC cells
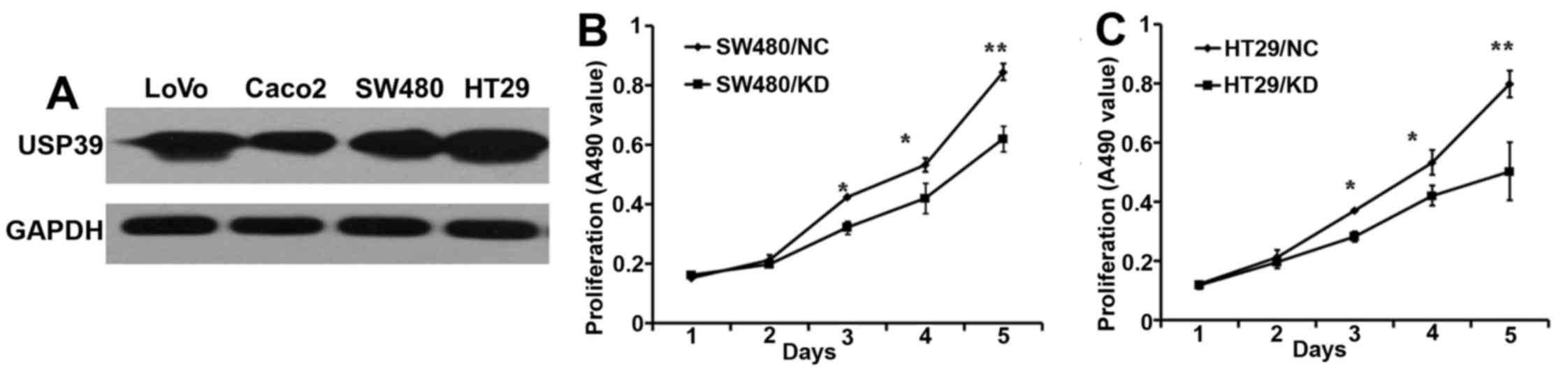
Next, we examined the USP39 protein expression in
four human CRC cell lines, LoVo, Caco2, SW480 and HT-29, using
western blot analysis, as shown in Fig.
3A. Our results indicated that indeed all these cell lines have
elevated expression of USP39. Next, we knocked down the USP39
expression in these CRC cells and analyzed its effects on their
growth, using MTT assay. As shown in Fig. 3B and C, USP39 knockdown
significantly inhibited the growth of HT-29 and SW480 cells, as
compared to control knock down (NC). However, the LoVo and Caco2
cells, did not show any significant differences in their growth
(data not shown).
USP39 knockdown inhibits migration and
invasion of CRC cells
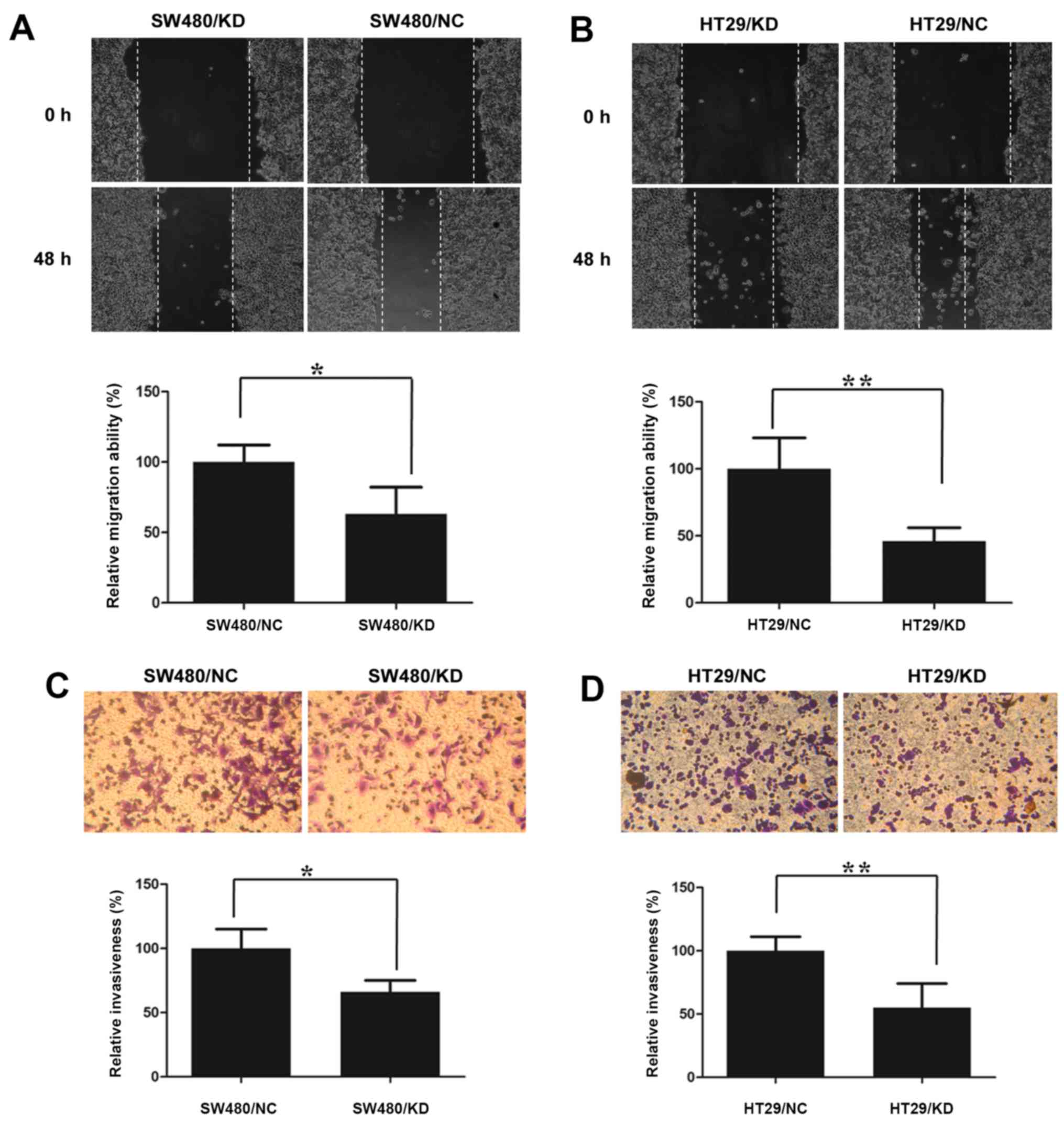
To assess the USP30 contribution to CRC cell (HT-29
and SW480 cells) migration and invasion, we performed the scratch
wound healing assay and Matrigel coated Transwell invasion assay.
As shown in Fig. 4A and B, the
scratch was about half closed after 48 h in control knockdown
cells, while the USP39 knocked down cells had very little effect on
the rate of wound closure. Similarly, USP39 knockdown also reduced
the cell invasion of both CRC cell types in comparison to the
control knockdown, as shown in Fig. 4C
and D.
USP39 knockdown in migration and
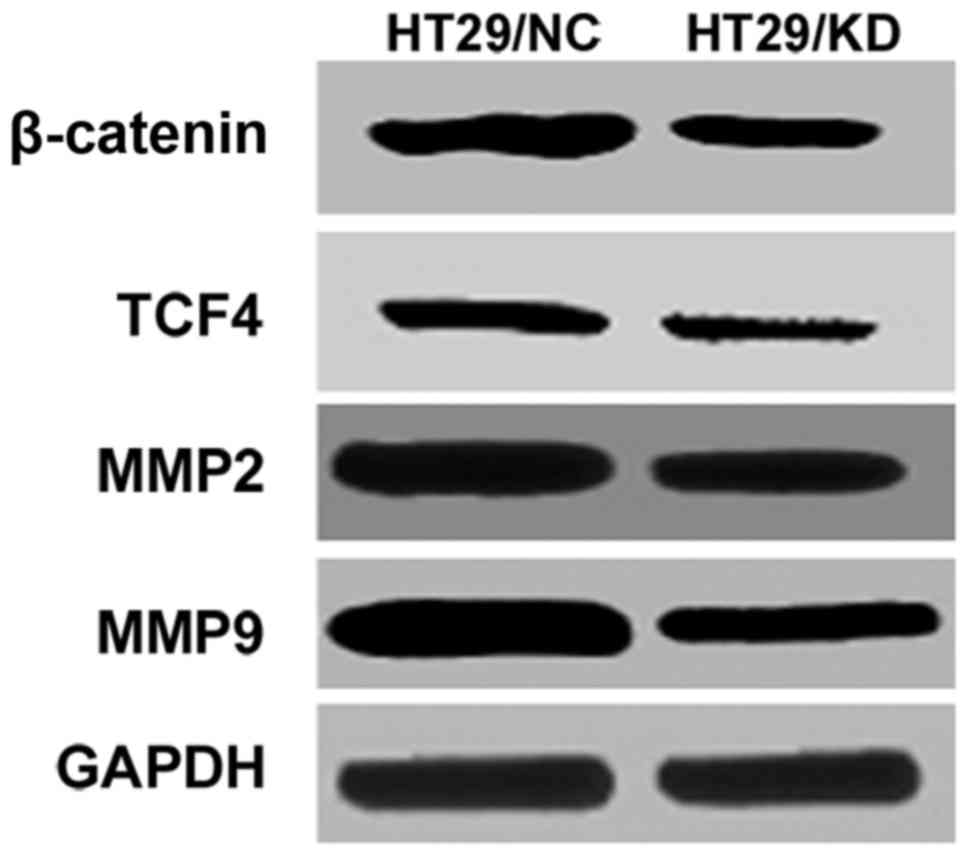
invasion involved the Wnt signaling pathway
Wnt signaling pathway and its modulators play a
significant oncogenic role in CRC development and progression
(12–14), we tested the effect of USP39
knockdown on proteins related to Wnt/β-catenin pathway and its
downstream factor matrix metalloproteinases (MMPs) (15,16).
Our results showed that USP39 knockdown significantly reduced the
expression of β-catenin, TCF4, MMP2 and MMP9 proteins, as shown in
Fig. 5.
Discussion
In our previous study, we have shown that USP39
promotes hepatocellular carcinoma (11), however, herein we first demonstrated
that USP39 expression in colorectal cancer tissues was higher as
compared with corresponding adjacent normal tissues. In addition,
we also reported that USP39 was highly expressed in multiple CRC
cell lines, and series of in vitro functional experiments
revealed that it promotes the growth, migration and invasion of CRC
cell lines. Knockdown of USP39 inhibited not only the growth of
HT29 and SW480 cells, but also their migration and invasion
ability. Further mechanistic insight into its function at molecular
level indicated that USP39 might be mediating these effects through
regulation of some of the proteins involved in Wnt/β-catenin
pathway, as Wnt/β-catenin pathway has been shown to influence the
growth and metastasis of CRC.
Wnt signaling is a crucial mechanism involved in the
regulation of cell proliferation, differentiation and morphogenesis
(17). Numerous studies have
suggested that the abnormal activation of Wnt/β-catenin signaling
pathway have a significant role in human tumorigenesis, especially
colorectal cancer (18). β-catenin
which is the key molecule in this pathway, is a multifunctional
protein (19), and acts as a key
transcriptional factor in the Wnt pathway. It widely participates
in the regulation of cell proliferation and migration (20,21).
In the present study, we observed a significant effect of USP39
knockdown on β-catenin levels, consistent with reduced growth and
migration.
MMP2 and MMP9 are proteins expressed on many tumor
cells, and act as potential indicators of invasion or metastasis in
colorectal tumors (22–24). These MMPs are regulated by
Wnt/β-catenin signaling pathway (25,26).
Activation of cytokine signaling and cell recruitment in the areas
of silica dust deposition can lead to excessive disruption of
extracellular matrix, which is associated with the secretion of
matrix metalloproteinases (MMPs). In the development of pulmonary
fibrosis, MMPs specifically cleave important constituents of
extracellular matrix (ECM) such as gelatin and collagen, and
therefore damage the structure of the alveolar wall (22). It has been reported that β-catenin
mediated expression of matrix metalloproteinases (MMPs) enhance
epithelial-mesenchymal transition (27). The key feature of tumor invasion and
metastasis is degrading extracellular matrix, and MMPs take part in
its degradation, and hence is related closely with the tumor
occurrence and development (28).
Thus, decreased expression of MMP2 and MMP9 due to USP39 knockdown,
coincide with role of USP39 in promoting growth and metastasis in
CRC tumor cells.
The treatment of colorectal cancer affecting
millions of people worldwide each year, is confronted by the
challenges of metastasis, relapse and drug resistance (29). Molecular targeted therapies have
become an alternative therapeutic approach, and are being used as a
combination therapy with standard chemotherapy agents. For example,
the recombinant monoclonal antibody bevacizumab against VEGF-A, and
the chimeric monoclonal antibody cetuximab against EGFR have been
recently approved for clinical use in combination with 5-FU and
irinotecan, respectively, to treat metastatic colorectal cancer
(30,31). Thus, identifying novel molecular
targets would provide additional options for effective treatment,
and the present study suggests that USP39 can be a tentative target
for CRC treatment.
In summary, the present study identified that USP39
expression is not only upregulated in CRC patients, but also
correlates with their overall survival. In addition, it is involved
in regulation of CRC cells growth, migration and invasion. In terms
of mechanistic understanding, USP39 effects are mediated through
Wnt/β-catenin pathway, as its inhibition significantly reduced the
expression of many proteins of this pathway. Thus, we conclude that
USP39 promotes growth and metastasis of colorectal cancer mainly
through the Wnt/β-catenin pathway.
Acknowledgements
The present study was supported by the grants from
the National Natural Science Foundation of China (nos. 31300103,
31171306, 31371373 and 31530046); the Clinical Medical Center for
Hepatobiliary Disease of Jiangsu Province (no. ZX201105); the
Clinical Medical Center for Digestive Disease of Jiangsu Province
(no. BL2012001); the Natural Science Foundation of Jiangsu Province
(BK20151395); the National Basic Research Program of China (973
program, 2012CB524900); and the Open Fund of State Key Laboratory
of Natural Medicines (no. SKLNMKF201606).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Goldberg RM, Rothenberg ML, Van Cutsem E,
Benson AB III, Blanke CD, Diasio RB, Grothey A, Lenz HJ, Meropol
NJ, Ramanathan RK, et al: The continuum of care: A paradigm for the
management of metastatic colorectal cancer. Oncologist. 12:38–50.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang J and Manley JL: Misregulation of
pre-mRNA alternative splicing in cancer. Cancer Discov.
3:1228–1237. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ladomery M: Aberrant alternative splicing
is another hallmark of cancer. Int J Cell Biol. 2013:4637862013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhou X, Li X, Cheng Y, Wu W, Xie Z, Xi Q,
Han J, Wu G, Fang J and Feng Y: BCLAF1 and its splicing regulator
SRSF10 regulate the tumorigenic potential of colon cancer cells.
Nat Commun. 5:45812014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Reyes-Turcu FE, Ventii KH and Wilkinson
KD: Regulation and cellular roles of ubiquitin-specific
deubiquitinating enzymes. Annu Rev Biochem. 78:363–397. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
van Leuken RJ, Luna-Vargas MP, Sixma TK,
Wolthuis RM and Medema RH: Usp39 is essential for mitotic spindle
checkpoint integrity and controls mRNA-levels of aurora B. Cell
Cycle. 7:2710–2719. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ríos Y, Melmed S, Lin S and Liu NA:
Zebrafish usp39 mutation leads to rb1 mRNA splicing defect and
pituitary lineage expansion. PLoS Genet. 7:e10012712011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang H, Ji X, Liu X, Yao R, Chi J, Liu S,
Wang Y, Cao W and Zhou Q: Lentivirus-mediated inhibition of USP39
suppresses the growth of breast cancer cells in vitro. Oncol
Rep. 30:2871–2877. 2013.PubMed/NCBI
|
|
10
|
Huang Y, Pan XW, Li L, Chen L, Liu X, Lu
JL, Zhu XM, Huang H, Yang QW, Ye JQ, et al: Overexpression of USP39
predicts poor prognosis and promotes tumorigenesis of prostate
cancer via promoting EGFR mRNA maturation and transcription
elongation. Oncotarget. 7:22016–22030. 2016.PubMed/NCBI
|
|
11
|
Yuan X, Sun X, Shi X, Jiang C, Yu D, Zhang
W, Guan W, Zhou J, Wu Y, Qiu Y, et al: USP39 promotes the growth of
human hepatocellular carcinoma in vitro and in vivo.
Oncol Rep. 34:823–832. 2015.PubMed/NCBI
|
|
12
|
Lou YF, Zou ZZ, Chen PJ, Huang GB, Li B,
Zheng DQ, Yu XR and Luo XY: Combination of gefitinib and DNA
methylation inhibitor decitabine exerts synergistic anti-cancer
activity in colon cancer cells. PLoS One. 9:e977192014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang J, Tsoi H, Li X, Wang H, Gao J, Wang
K, Go MY, Ng SC, Chan FK, Sung JJ, et al: Carbonic anhydrase IV
inhibits colon cancer development by inhibiting the Wnt signalling
pathway through targeting the WTAP-WT1-TBL1 axis. Gut. 13:e78–e79.
2015.
|
|
14
|
Park J and Jeong S: Wnt activated
β-catenin and YAP proteins enhance the expression of non-coding RNA
component of RNase MRP in colon cancer cells. Oncotarget.
6:34658–34668. 2015.PubMed/NCBI
|
|
15
|
Jung O, Lee J, Lee YJ, Yun JM, Son YJ, Cho
JY, Ryou C and Lee SY: Timosaponin AIII inhibits migration and
invasion of A549 human non-small-cell lung cancer cells via
attenuations of MMP-2 and MMP-9 by inhibitions of ERK1/2, Src/FAK
and β-catenin signaling pathways. Bioorg Med Chem Lett.
26:3963–3967. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shalaby MA, Nounou HA, Ms AOA, Azzam N and
Saeed HM: Associations between single nucleotide polymorphisms of
COX-2 and MMP-2 genes and colorectal cancer susceptibility in the
Saudi population. Asian Pac J Cancer Prev. 15:4989–4994. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ao N and Liu Y, Bian X, Feng H and Liu Y:
Ubiquitin-specific peptidase 22 inhibits colon cancer cell invasion
by suppressing the signal transducer and activator of transcription
3/matrix metalloproteinase 9 pathway. Mol Med Rep. 12:2107–2113.
2015.PubMed/NCBI
|
|
18
|
Moon RT, Brown JD and Torres M: WNTs
modulate cell fate and behavior during vertebrate development.
Trends Genet. 13:157–162. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu M, Wang S, Song YU, Yao J, Huang K and
Zhu X: Apigenin suppresses colorectal cancer cell proliferation,
migration and invasion via inhibition of the Wnt/β-catenin
signaling pathway. Oncol Lett. 11:3075–3080. 2016.PubMed/NCBI
|
|
20
|
Li X, Zhang X, Liu X, Tan Z, Yang C, Ding
X, Hu X, Zhou J, Xiang S, Zhou C, et al: Caudatin induces cell
apoptosis in gastric cancer cells through modulation of
Wnt/β-catenin signaling. Oncol Rep. 30:677–684. 2013.PubMed/NCBI
|
|
21
|
Fu Y, Zheng S, An N, Athanasopoulos T,
Popplewell L, Liang A, Li K, Hu C and Zhu Y: β-catenin as a
potential key target for tumor suppression. Int J Cancer.
129:1541–1551. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim W, Kim M and Jho EH: Wnt/β-catenin
signalling: From plasma membrane to nucleus. Biochem J. 450:9–21.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tutton MG, George ML, Eccles SA, Burton S,
Swift RI and Abulafi AM: Use of plasma MMP-2 and MMP-9 levels as a
surrogate for tumour expression in colorectal cancer patients. Int
J Cancer. 107:541–550. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee MA, Park JH, Rhyu SY, Oh ST, Kang WK
and Kim HN: Wnt3a expression is associated with MMP-9 expression in
primary tumor and metastatic site in recurrent or stage IV
colorectal cancer. BMC Cancer. 14:125–131. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu B, Crampton SP and Hughes CC: Wnt
signaling induces matrix metalloproteinase expression and regulates
T cell transmigration. Immunity. 26:227–239. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zuo F, Kaminski N, Eugui E, Allard J,
Yakhini Z, Ben-Dor A, Lollini L, Morris D, Kim Y, DeLustro B, et
al: Gene expression analysis reveals matrilysin as a key regulator
of pulmonary fibrosis in mice and humans. Proc Natl Acad Sci USA.
99:6292–6297. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Orlichenko LS and Radisky DC: Matrix
metalloproteinases stimulate epithelial-mesenchymal transition
during tumor development. Clin Exp Metastasis. 25:593–600. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fagan-Solis KD, Schneider SS, Pentecost
BT, Bentley BA, Otis CN, Gierthy JF and Arcaro KF: The RhoA pathway
mediates MMP-2 and MMP-9-independent invasive behavior in a
triple-negative breast cancer cell line. J Cell Biochem.
114:1385–1394. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gill S, Blackstock AW and Goldberg RM:
Colorectal cancer. Mayo Clin Proc. 82:114–129. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cheng YD, Yang H, Chen GQ and Zhang ZC:
Molecularly targeted drugs for metastatic colorectal cancer. Drug
Des Devel Ther. 7:1315–1322. 2013.PubMed/NCBI
|
|
31
|
Peng X, Luo Z, Kang Q, Deng D, Wang Q,
Peng H, Wang S and Wei Z: FOXQ1 mediates the crosstalk between
TGF-β and Wnt signaling pathways in the progression of colorectal
cancer. Cancer Biol Ther. 16:1099–1109. 2015. View Article : Google Scholar : PubMed/NCBI
|