Introduction
Hepatocellular carcinoma (HCC) is the sixth most
prevalent cancer globally and the second leading cause of
cancer-related death (1). Due to
the important metabolic functions of the liver within the body,
malignancies in the liver typically lead to life-threatening
consequences (2–5). Early diagnosis is essential for the
successful treatment of liver cancer; however, there is typically a
lack of specific symptoms (2,6,7). Thus,
the identification of a novel biomarker of liver cancer may aid in
developing new methods for treating HCC (8).
RNAi is a gene-based technology that utilizes shRNA,
and is frequently used to suppress the expression of target genes
(9–11). RNAi and lentiviral technologies are
widely used in genetic research (12–14),
and RNAi technology could continue to provide novel methods for the
study of human gene function, early cancer diagnostics and targeted
therapy (15–17).
Ribosomes, as the sole organelles responsible for
protein synthesis, serve essential functions in all cells. The
assembly and activity of ribosomes are regulated by complex and
precise intracellular pathways. A study indicated that the
overexpression of oncogenes and downregulation of tumor suppressor
genes may lead to abnormal regulation of ribosome biogenesis and
the initiation of protein translation (18). In recent years, abundant research
has pointed out that dysfunction of ribosome biogenesis plays a
major role in the process of tumorigenesis (18–20).
Additionally, it has been shown that patients with quantitative and
qualitative changes in their ribosomes were more susceptible to
cancer (5). These data indicate
that the biogenesis of ribosomes may be involved in the development
and progression of cancers.
Regulator of ribosome synthesis 1 (RRS1), which
comprises 203 amino acids, was initially discovered in yeast
(21), and is a conserved nuclear
protein in eukaryotes (22). In a
murine model of Huntington's disease, high-level expression of RRS1
was identified in neurons (23).
Furthermore, immunohistochemical staining has shown that RRS1 is
not solely present in the nucleus, as in yeast, but is also present
in the endoplasmic reticulum (22,24),
and its expression may be induced by neuronal endoplasmic reticulum
stress (25,26). Additionally, secretion-deficient
yeast carrying a nonsense mutation in an amino codon in the
C-terminal of Rrs1 exhibited attenuated transcriptional suppression
of ribosomal (r)RNA and protein (21). Therefore, RRS1 is a key protein
connection between ribosome biogenesis and protein secretion
(27,28). It has been reported that RRS1 plays
an important role in the regulation of ribosome biogenesis,
particularly in the maturation of the ribosomal subunits 25S rRNA
and 60S rRNA (21,29,30).
Based on these critical functions of RRS1, we hypothesized that the
expression and activity of RRS1 may be abnormally regulated in
cancer cells, in which physiological function changes dramatically.
To the best of our knowledge, the expression of RRS1 in most cancer
types, including HCC, has not been reported. Therefore, the present
study investigated the expression of RRS1 in human HCC, as well as
the roles of RRS1 in the HCC cell line SMMC-7721 through the use of
lentivirus-mediated RNA interference (RNAi).
Materials and methods
Human tissue samples, cell lines,
reagents and antibodies
Human HCC tissues and corresponding paracancerous
tissues were obtained from 18 patients with HCC who first underwent
surgery at the Beijing 302 Hospital (Beijing, China) between
January 2013 and June 2013. All of the human tissue samples were
collected with the consent of patients or their legal
representatives and the approval of the local medical ethics
committee. All samples were immediately frozen in liquid nitrogen
and stored at −80°C to reduce RNA degradation. The human liver
cancer cell lines SMMC-7721, HepG2, Huh-7 and Hep3B, and the human
embryonic kidney cell line 293T, were purchased from the Type
Culture Collection of the Chinese Academy of Sciences (Shanghai,
China) and cultured in RPMI-1640 medium containing 10% fetal bovine
serum, 100 µg/ml streptomycin and 100 IU/ml penicillin at 37°C in
95% humidity and 5% CO2. Rabbit anti-RRS1 polyclonal
antibodies were purchased from Abcam (cat. no. AB188161, Shanghai,
China). Mouse anti-GAPDH monoclonal antibodies were obtained from
Santa Cruz Biotechnology, Inc. (cat. no. sc-32233, Dallas, TX,
USA). Anti-rabbit and anti-mouse IgG secondary antibodies were from
Santa Cruz Biotechnology, Inc. (cat. no. sc-2004). An APC Annexin V
apoptosis detection kit was from eBioscience (San Diego, CA, USA).
Propidium iodide (PI) was from Sigma-Aldrich (St. Louis, MO,
USA).
Lentivirus-mediated short hairpin
(sh)RNA delivery
An RRS1 shRNA sequence was inserted into a
pGCSIL-GFP lentivirus RNAi expression system (GeneChem Co., Ltd.,
Shanghai, China). The effective targeting sequence of RRS1 (GCT GCC
TTC ATT GAG TTT A) was selected by western blot analysis. A
non-silencing shRNA sequence (TTC TCC GAA CGT GTT CAC GT) was used
as a negative control, which does not target any gens in humans,
mice or rats as determined by screening with NCBI RefSeq. The shRNA
vectors were co-transfected with the lentiviral packaging plasmids
pHelper1.0 and pHelper2.0 into 293T cells using Lipofectamine 2000
(Invitrogen, Shanghai, China) to generate the respective
lentiviruses. Viral stocks were collected from the 293T cells 3
days after infection and were used to infect SMMC-7721 cells. The
SMMC-7721 cells were infected with the RRS1-siRNA-lentivirus
(Lv-shRRS1 group) or negative control lentivirus (Lv-shCon group)
according to the recommended multiplicity of infection (MOI). The
stem-loop-stem oligos (shRNAs) were synthesized, annealed, and
ligated into the pGCSIL-GFP lentivirus RNAi expression system. The
lentiviral-based shRNA-expressing vectors were confirmed by DNA
sequencing.
After lentiviral infection, the SMMC-7721 cells were
observed under a fluorescence microscope (MicroPublisher 3.3RTV;
Olympus Corp., Tokyo, Japan) at 3 days post-infection. The
lentiviral transfection experiments were performed as in a previous
study (31).
Reverse transcription-quantitative PCR
(RT-qPCR) analysis
Total RNA was isolated from cell lines or frozen
tissues using TRIzol reagent (Invitrogen) and reverse transcribed
with a PrimeScript® RT reagent kit (Takara, Dalian,
China). cDNA was normalized with GAPDH. PCR was performed via a
two-step method using a SYBR® premix Ex Taq™ II kit
(Takara). The RT-qPCR comprised an initial denaturation at 95°C for
15 sec, then 45 cycles at 95°C for 5 sec and 60°C for 30 sec. The
melting curve was monitored with the following conditions: 60 sec
at 95°C, 60 sec at 55°C, followed by a temperature range from 55°C
to 95°C increased by 0.5°C every 4 sec. PCR reactions were carried
out in triplicate. GAPDH was used as internal control. The relative
amount of each cDNA was determined using the 2−∆∆Cq
method (32). The primer sequences
used for RRS1 and GAPDH were as follows: RRS1 forward,
CCGAAAAGGGGTTGAAACTTCC, and reverse, CCCTACCGGACACCAGAGTAA; and
GAPDH forward, TGACTTCAACAGCGACACCCA, and reverse,
CACCCTGTTGCTGTAGCCAAA. The PCR products of the RRS1 and GAPDH
primers were 153 and 121 bp in length, respectively.
Western blot analysis
Protein samples were prepared from SMMC-7721 cells 5
days after infection with RRS1 or control shRNA lentiviruses, and
were subjected to 10% SDS-PAGE (20 µg protein per lane), as
described by Laemmli (33). The
separated proteins were transferred onto polyvinylidene fluoride
(PVDF) membranes (EMD Millipore, Billerica, MA, USA) and probed
with rabbit anti-RRS1 (1:1,000) or mouse anti-GAPDH (1:5,000)
antibodies at room temperature for 2 h, followed by incubation with
the anti-rabbit and anti-mouse IgG secondary antibodies (1:5,000)
at room temperature for 2 h. GAPDH was used as an internal control.
After three washes with 10% bovine serum albumin in
phosphate-buffered saline (PBS), immunolabeled proteins on the PVDF
membranes were detected with an Enhanced Chemiluminescence kit
(Amersham, Uppsala, Sweden) and exposed to X-ray film. Bands on the
X-ray films were quantified with an ImageQuant densitometric
scanner (Molecular Devices LLC, Sunnyvale, CA, USA).
Cell proliferation assay
Five days after lentiviral infection, SMMC-7721
cells were trypsinized, resuspended, seeded into 96-well plates at
a concentration of 2,000 cells per well, and incubated at 37°C in
5% CO2. The number of viable cells was measured at daily
intervals (day 1, 2, 3, 4 and 5) using a Cellomics ArrayScan™ VT1
HCS automated reader (Cellomics, Inc., Pittsburgh, PA, USA).
Colony formation assay
SMMC-7721 cells were seeded into 6-well plates
(1,000 cell/well; three duplicate wells and cultured at 37°C in 5%
CO2. At 72 h after lentiviral infection, cells were
grown for 7 days with media replacement every 2 days to enable
colony formation. After 7 days, the cells were washed with PBS and
fixed with paraformaldehyde for 30 min, then washed with PBS and
stained with Giemsa for 20 min. The cells were subsequently washed
three times with ddH2O to obtain a clean background. The
number of colonies and the cell number in each colony were counted
and statistically analyzed according to the method by Zhang et
al (34).
Flow cytometric analysis
An apoptosis assay was performed with an APC Annexin
V staining kit. When cell confluence reached 85% (5–6 days after
infection), the cells were harvested by centrifugation at 10,000 ×
g for 5 min. The pellets were washed twice with cold PBS, fixed
with 70% chilled ethanol, centrifuged at 10,000 × g for 5 min to
remove the ethanol, and resuspended in PBS. The suspensions were
filtered through 400-mesh membranes and centrifuged at 10,000 × g
for 5 min. The cells were resuspended with 1X Annexin V staining
buffer and stained with Annexin V-APC at room temperature for 15
min in the dark. Approximately 1.0×105 fixed cells were
analyzed with a FACSCalibur flow cytometer (BD Biosciences,
Franklin Lakes, NJ, USA). The assay was carried out in
triplicate.
Cell cycle distribution was analyzed
with PI staining
In brief, 1.5×105 cells at 4 days
post-infection were seeded into 6-cm dishes and cultured for 40 h
at 37°C. Cells were harvested by centrifugation at 10,000 × g for 5
min, washed with PBS, and fixed with 70% cold ethanol for 1 h.
Cells were then collected by centrifugation at 10,000 × g for 5
min, resuspended in PBS containing 100 µg/ml of DNase-free RNase
and 40 µg/ml PI, and incubated for 1 h at 37°C. A total of
1.0×104 fixed cells were analyzed by flow cytometry
(FACSCalibur; BD Biosciences).
Statistical analysis
One-way ANOVA and a Student's t-test were used for
data analysis. Statistical analysis was performed using SPSS 22.0
software (IBM Corp., Armonk, NY, USA). All values were expressed as
the mean ± standard deviation (SD). P<0.05 was considered to
indicate a statistically significant difference.
Results
RRS1 is expressed in human HCC tissues
and cell lines
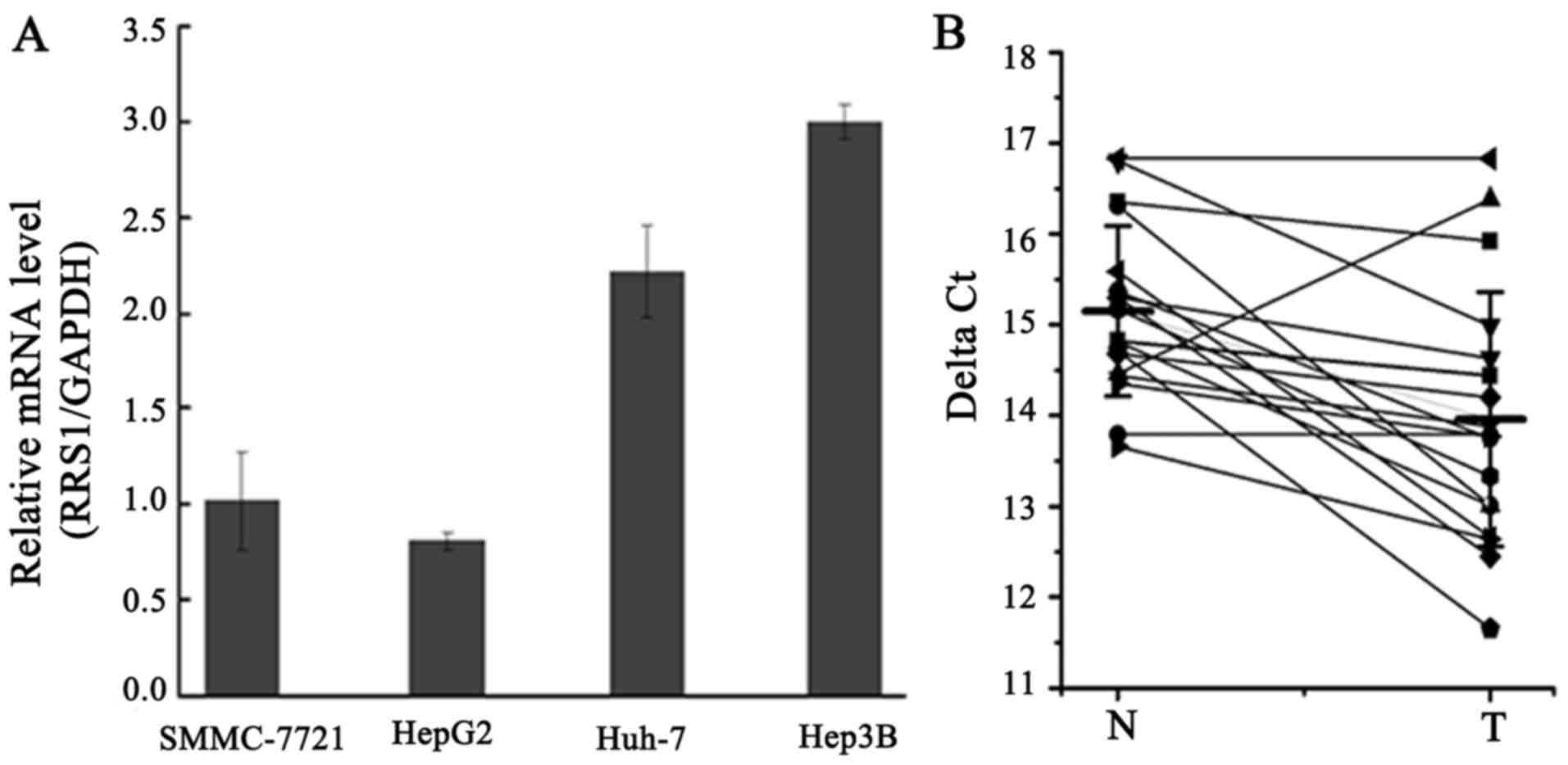
The expression of RRS1 mRNA was assessed in human
liver cancer cell lines SMMC-7721, HepG2, Huh-7, and Hep3B. PCR
analysis indicated that RRS1 mRNA was expressed in all four human
liver cancer cell lines (Fig. 1A).
Furthermore, the expression of RRS1 mRNA in 18 paired tissue
samples (HCC and paracancerous tissues) was measured by qPCR. As
shown in Fig. 1B, the level of RRS1
mRNA in HCC tissues was approximately 3–4-fold higher than that in
adjacent tissues. Thus, elevated expression of RRS1 may play an
important role in the pathogenesis of human HCC.
Lentiviral shRNA inhibits the mRNA and
protein expression of RRS1 in SMMC-7721 cells
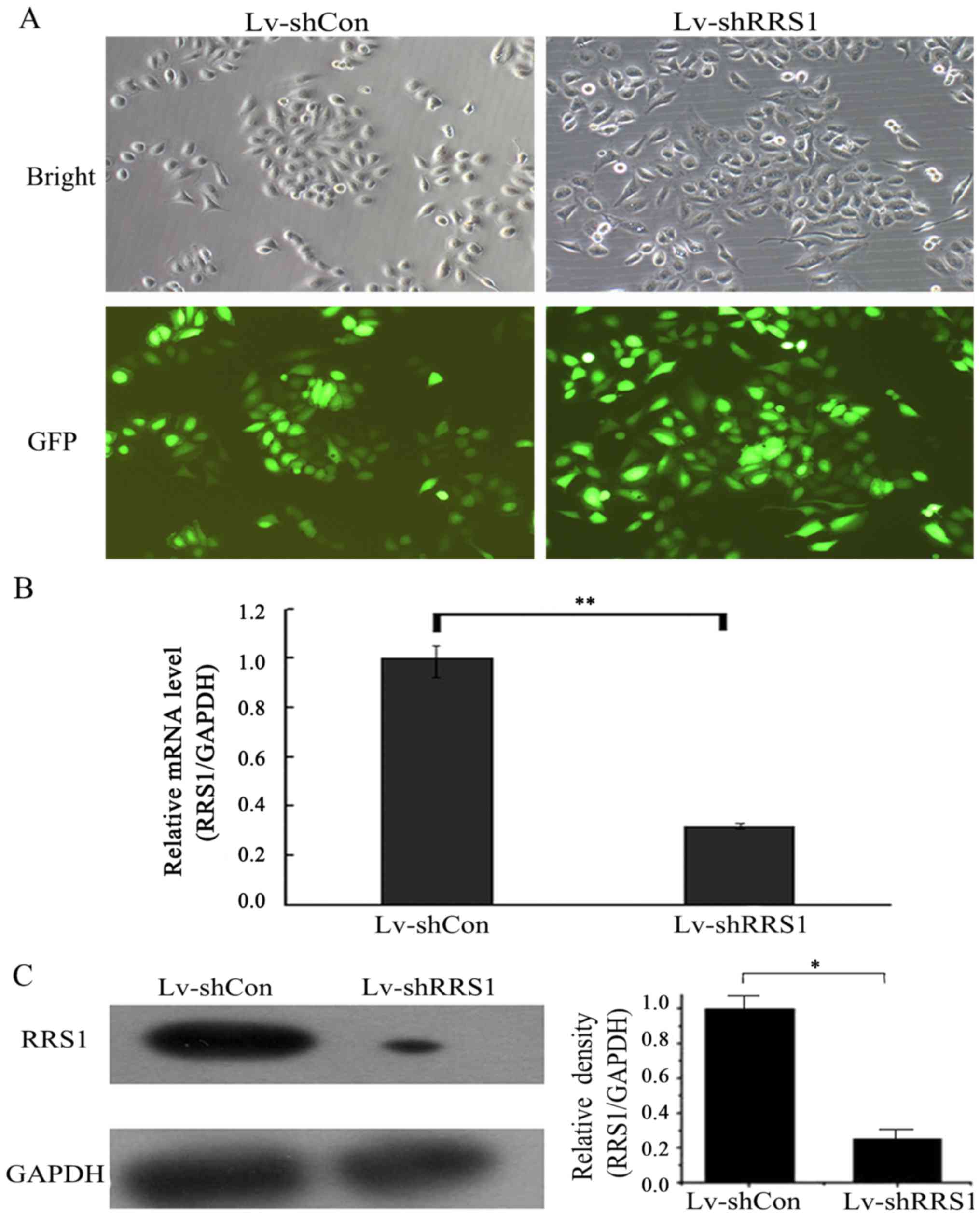
To investigate the role of RRS1 in HCC, the human
HCC cell line SMMC-7721 was selected for in vitro study. A
loss-of-function study was performed via lentivirus-mediated shRNA
knockdown of RRS1. A lentiviral vector system expressing shRNA
against RRS1 and GFP, as a reporter gene, was established. To
determine the infection efficiency of lentivirus in SMMC-7721 cells
at 30% cell density, cells infected with Lv-shRRS1 or Lv-shCon
vector were observed under a fluorescence microscope at 3 days
post-infection. As shown in Fig.
2A, >90% of SMMC-7721 cells were found to express GFP, which
indicated a high infection efficiency of the lentivirus.
To verify the knockdown efficiency of Lv-shRRS1, the
mRNA and protein expression levels of RRS1 in SMMC-7721 cells were
detected by PCR and western blot analysis at 5 days post-infection.
As shown in Fig. 2B, the mRNA
expression levels of RRS1 in the Lv-shRRS1 infection group were
downregulated by 68.0% when compared with the Lv-shCon infection
group (P<0.01). As shown in Fig.
2C, the protein expression levels of RRS1 in the Lv-shRRS1
infection group were also significantly decreased (73.0% decrease)
when compared with the Lv-shCon infection group.
Knockdown of RRS1 significantly
inhibits SMMC-7721 cell proliferation
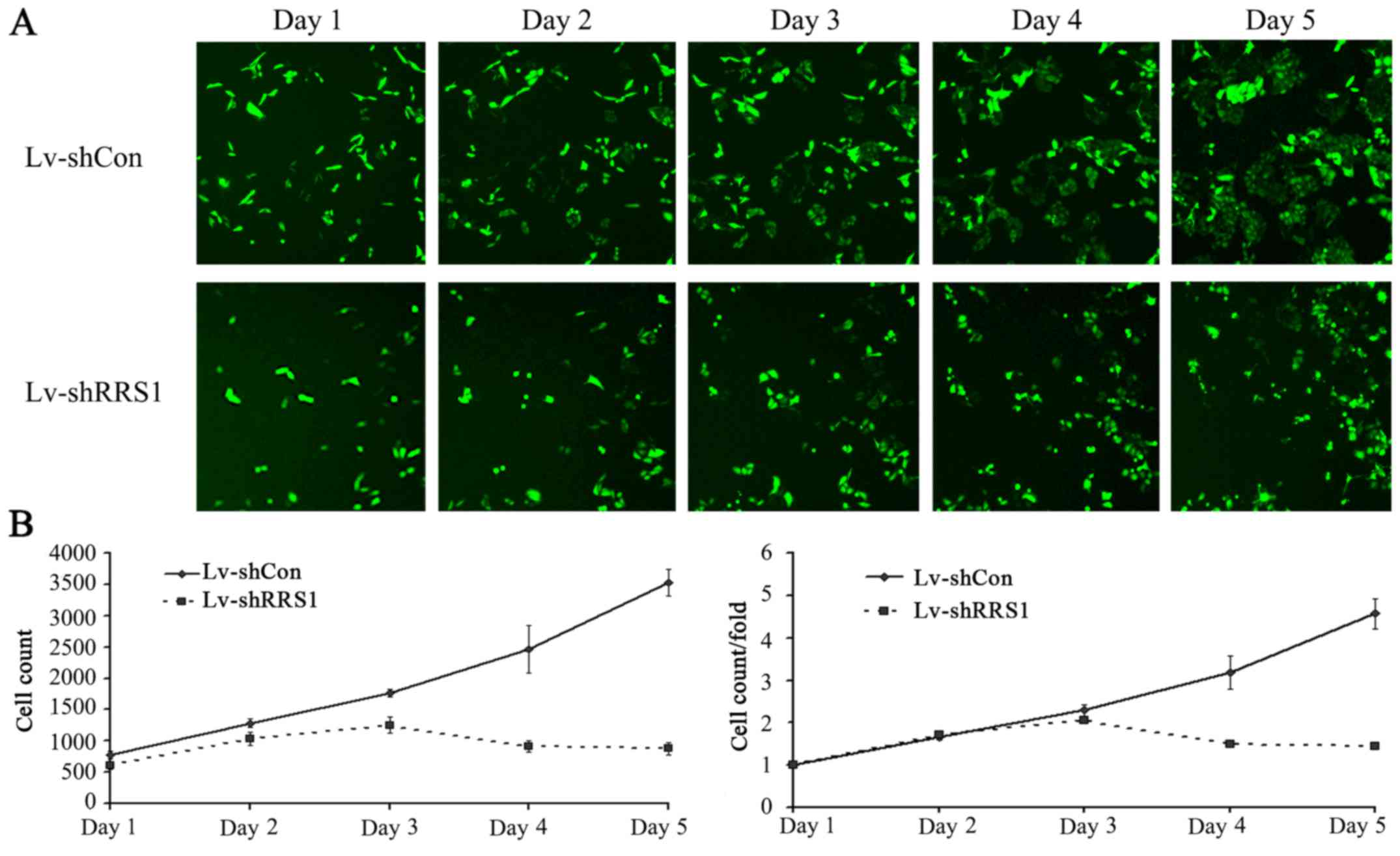
To investigate the effect of RRS1 knockdown on cell
proliferation, the two cell groups were seeded into 96-well plates
and subjected to daily cellomics analysis for 5 days (Fig. 3A). As shown in Fig. 3B, lentivirus-mediated shRNA
knockdown of RRS1 significantly inhibited the proliferation of
SMMC-7721 cells when compared with cells infected with control
lentivirus (P<0.05). This result indicated that RRS1 played an
important role in the proliferation of SMMC-7721 cells.
Knockdown of RRS1 significantly
inhibits the colony-forming ability of SMMC-7721 cells
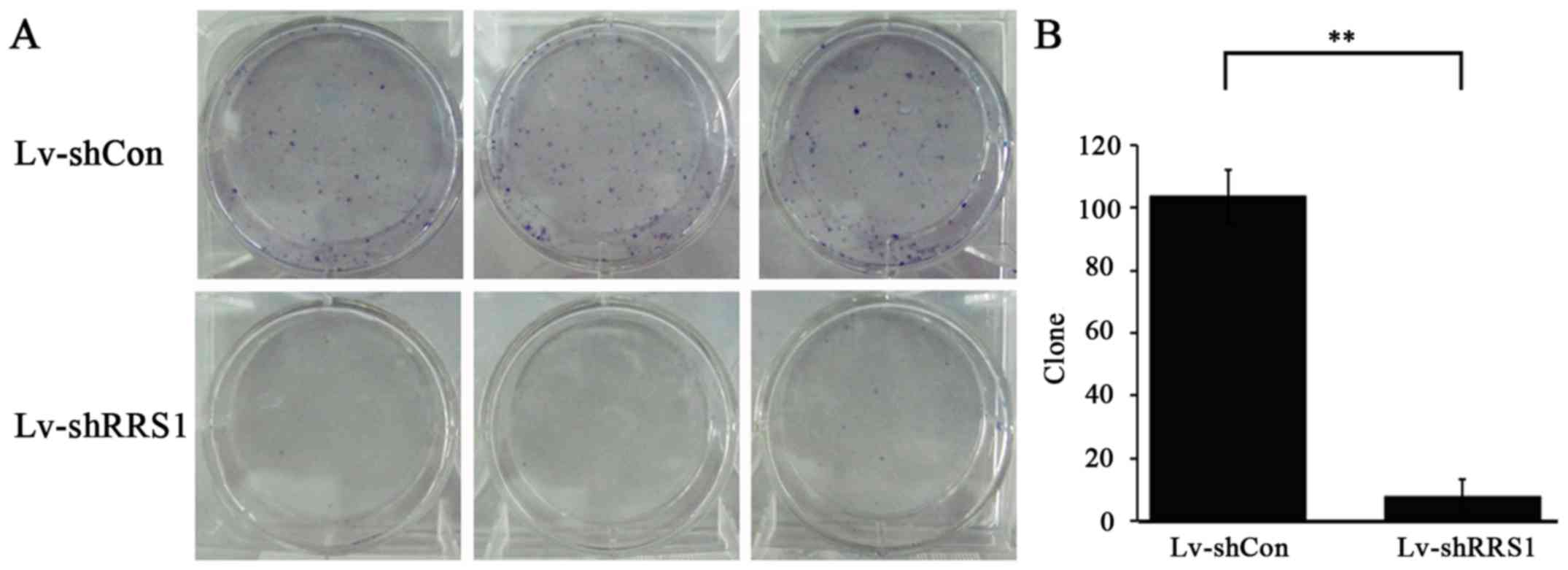
To study the long-term effect of RRS1 shRNA
lentivirus on cell growth, a colony forming assay was conducted in
SMMC-7721 cells. As shown in Fig.
4A, the number of colonies in the Lv-shRRS1 group was lower
than that in the Lv-shCon group. Specifically, the colony number
was 9±5 in the Lv-shRRS1 group compared with 104±9 in the Lv-shCon
group (P<0.01; Fig. 4B). These
results indicate that downregulation of RRS1 significantly
decreases the colony formation capacity of HCC cells in
vitro.
Knockdown of RRS1 significantly
increases apoptosis of SMMC-7721 cells
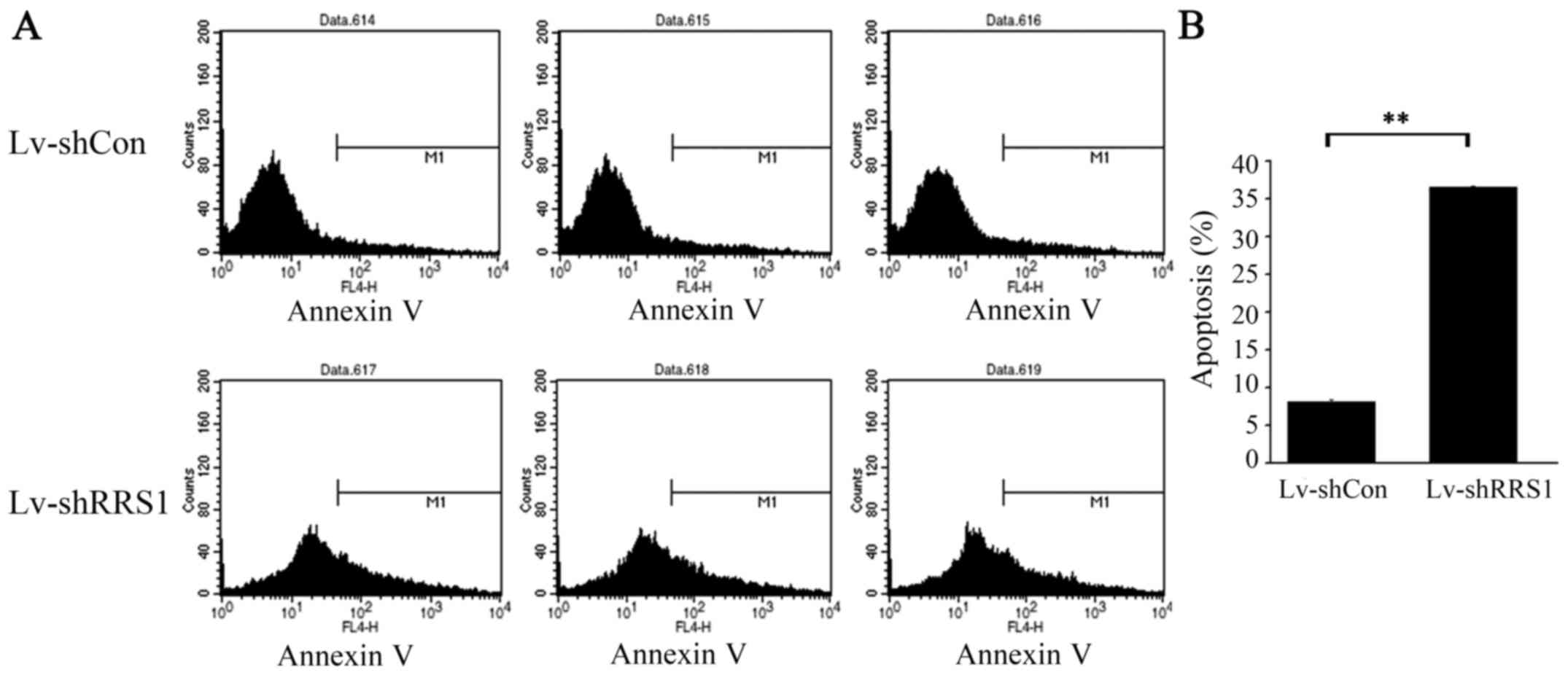
To determine the effect of RRS1 knockdown in HCC
cells, Annexin V staining and flow cytometric analysis were
performed on SMMC-7721 cells 5–6 days after lentiviral infection,
when cell confluence reached 85%. As shown in Fig. 5, the apoptotic rate of SMMC-7721
cells in the Lv-shRRS1 group was significantly higher than that in
the Lv-shCon group (36.52±0.19% vs. 8.15±0.30%; P<0.001).
Effects of RRS1 knockdown on the cell
cycle distribution of SMMC-7721 cells
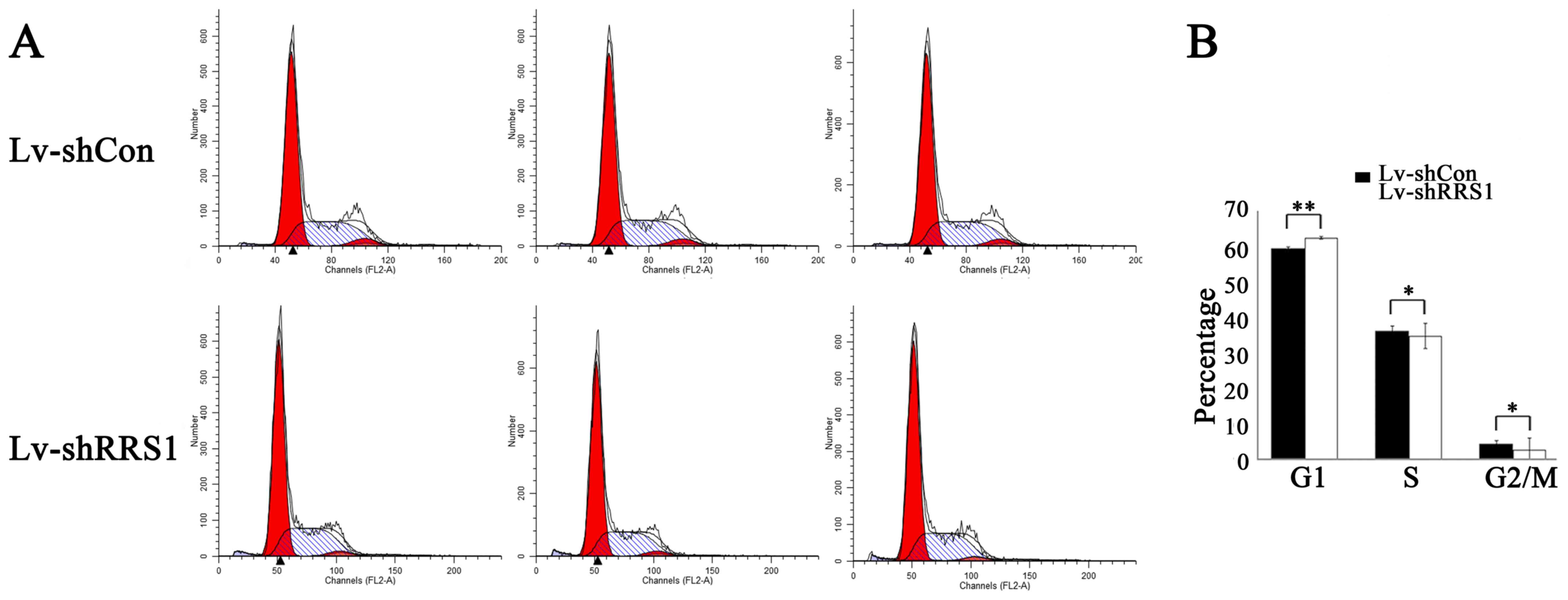
To elucidate the impact of RRS1 knockdown on the
cell cycle progression of HCC cells, SMMC-7721 cells were subjected
to a PI staining flow cytometry assay 6 days after lentiviral
infection. The Lv-shRRS1 infection group exhibited an increased
proportion of cells in G1 phase (Lv-shRRS1 62.49±0.59% vs. Lv-shCon
59.41±0.41%, P=0.003) and reduced proportions of cells in S phase
(Lv-shRRS1 34.77±0.55% vs. Lv-shCon 36.21±0.51%, P=0.03) and G2/M
phase (Lv-shRRS1 2.74±0.50% vs. Lv-shCon 4.39±0.12%, P=0.024) when
compared with the Lv-shCon group (Fig.
6). These results indicated that knockdown of RRS1 could induce
G1 phase arrest in HCC cells.
Discussion
The incidence and mortality rates of HCC have
increased over recent years. An estimated 782,500 new liver cancer
cases and 745,500 liver cancer-related deaths occurred worldwide
during 2012, with China alone accounting for approximately 50% of
the total cases and deaths (2).
RRS1 is a conserved eukaryotic nuclear protein. RRS1
can be regulated to control the speed of ribosome biogenesis
according to the energy state of the cell, thereby maintaining
cellular homeostasis. Additionally, RRS1 is an important protein in
signal transduction pathways related to protein secretion and
ribosome synthesis. It has been reported that the RRS1 gene may be
involved in the pathogenesis of Huntington's disease, although the
specific underlying mechanism is unclear (26). A study by Gambe et al
(24) showed that RRS1 was involved
in chromosome aggregation. Notably, their experiments identified
abnormalities in the chromosome alignment and spindle organization
of RRS1-depleted cells, which resulted in mitotic delay.
Furthermore, it has been suggested that RRS1 may play a role in the
development of cervical cancer through regulation of the cell
cycle. However, the expression and function of RRS1 in other
tumors, including HCC, have not been reported.
In the present study, we successfully constructed an
shRNA lentiviral vector that targeted RRS1 mRNA, which may provide
an experimental basis for further research. To evaluate the
function of RRS1 in HCC cells, we efficiently silenced the RRS1
gene, and found that the proportion of SMMC-7721 cells in G1 phase
was increased, while the proportions of cells in S and G2/M phases
were decreased, relative to the control group. Further assays
verified that silencing of the RRS1 gene decreased cell
proliferation and colony formation ability, and promoted apoptosis.
These results suggest that RRS1 may act as an oncogenic factor in
liver cancer. However, the specific signaling pathway underlying
the involvement of RRS1 in the biological behavior of HCC cells
remains unclear, and further study is ongoing.
In conclusion, this is the first study to
demonstrate that the downregulation of RRS1 in SMMC-7721 cells by
RNAi could decrease cell proliferation, inhibit colony formation
ability and induce cell apoptosis. Therefore, RRS1 may be involved
in the regulation of hepatocellular proliferative activity, cell
cycling and apoptotic ability. These findings may aid in developing
gene targeting therapies for patients with liver cancer, and
provide an experimental basis for further research into the
functions of RRS1 in the progression, invasion, metastasis and
tumor recurrence of HCC.
Glossary
Abbreviations
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
RSS1
|
regulator of ribosome synthesis 1
|
|
RNAi
|
RNA interference
|
|
shRNA
|
short hairpin RNA
|
References
|
1
|
World Health Organization, . World Cancer
Report 2014Stewart BW and Wild CP: World Health Organization;
International Agency for Research on Cancer. WHO Press, Geneva:
2015, View Article : Google Scholar
|
|
2
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zuo TT, Zheng RS, Zhang SW, Zeng HM and
Chen WQ: Incidence and mortality of liver cancer in China in 2011.
Chin J Cancer. 34:508–513. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wei KR, Yu X, Zheng RS, Peng XB, Zhang SW,
Ji MF, Liang ZH, Ou ZX and Chen WQ: Incidence and mortality of
liver cancer in China, 2010. Chin J Cancer. 33:388–394.
2014.PubMed/NCBI
|
|
5
|
Petrick JL, Braunlin M, Laversanne M,
Valery PC, Bray F and McGlynn KA: International trends in liver
cancer incidence, overall and by histologic subtype, 1978–2007. Int
J Cancer. 139:1534–1545. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fitzmorris P, Shoreibah M, Anand BS and
Singal AK: Management of hepatocellular carcinoma. J Cancer Res
Clin Oncol. 141:861–876. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nguyen MH and Keeffe EB: Screening for
hepatocellular carcinoma. J Clin Gastroenterol. 35 Suppl 2:S86–S91.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shimizu D, Inokawa Y, Sonohara F, Inaoka K
and Nomoto S: Search for useful biomarkers in hepatocellular
carcinoma, tumor factors and background liver factors (Review).
Oncol Rep. 37:2527–2542. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pecot CV, Calin GA, Coleman RL,
Lopez-Berestein G and Sood AK: RNA interference in the clinic:
challenges and future directions. Nat Rev Cancer. 11:59–67. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sioud M: RNA interference: Mechanisms,
technical challenges, and therapeutic opportunities. Methods Mol
Biol. 1218:1–15. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Holoch D and Moazed D: RNA-mediated
epigenetic regulation of gene expression. Nat Rev Genet. 16:71–84.
2015. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu H, Liang S, Yang X, Ji Z, Zhao W, Ye X
and Rui J: RNAi-mediated RPL34 knockdown suppresses the growth of
human gastric cancer cells. Oncol Rep. 34:2267–2272. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu K, Li X, Cao Y, Ge Y, Wang J and Shi
B: MiR-132 inhibits cell proliferation, invasion and migration of
hepatocellular carcinoma by targeting PIK3R3. Int J Oncol.
47:1585–1593. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Q, Hu H, Shi X and Tang W: Knockdown
of S100P by lentiviral-mediated RNAi promotes apoptosis and
suppresses the colony-formation ability of gastric cancer cells.
Oncol Rep. 31:2344–2350. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Du P, Ye L, Yang Y and Jiang WG: Candidate
of metastasis 1 regulates in vitro growth and invasion of
bladder cancer cells. Int J Oncol. 42:1249–1256. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Roop RP and Ma CX: Endocrine resistance in
breast cancer: Molecular pathways and rational development of
targeted therapies. Future Oncol. 8:273–292. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li J and Huang L: Targeted delivery of
RNAi therapeutics for cancer therapy. Nanomedicine (Lond).
5:1483–1486. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Montanaro L, Treré D and Derenzini M:
Changes in ribosome biogenesis may induce cancer by down-regulating
the cell tumor suppressor potential. Biochim Biophys Acta.
1825:101–110. 2012.PubMed/NCBI
|
|
19
|
Donati G, Montanaro L and Derenzini M:
Ribosome biogenesis and control of cell proliferation: p53 is not
alone. Cancer Res. 72:1602–1607. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Montanaro L, Treré D and Derenzini M:
Nucleolus, ribosomes, and cancer. Am J Pathol. 173:301–310. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tsuno A, Miyoshi K, Tsujii R, Miyakawa T
and Mizuta K: RRS1, a conserved essential gene, encodes a novel
regulatory protein required for ribosome biogenesis in
Saccharomyces cerevisiae. Mol Cell Biol. 20:2066–2074. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang J, Harnpicharnchai P, Jakovljevic J,
Tang L, Guo Y, Oeffinger M, Rout MP, Hiley SL, Hughes T and
Woolford JL Jr: Assembly factors Rpf2 and Rrs1 recruit 5S rRNA and
ribosomal proteins rpL5 and rpL11 into nascent ribosomes. Genes
Dev. 21:2580–2592. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fossale E, Wheeler VC, Vrbanac V, Lebel
LA, Teed A, Mysore JS, Gusella JF, MacDonald ME and Persichetti F:
Identification of a presymptomatic molecular phenotype in Hdh CAG
knock-in mice. Hum Mol Genet. 11:2233–2241. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gambe AE, Matsunaga S, Takata H,
Ono-Maniwa R, Baba A, Uchiyama S and Fukui K: A nucleolar protein
RRS1 contributes to chromosome congression. FEBS Lett.
583:1951–1956. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wan K, Kawara H, Yamamoto T, Kume K,
Yabuki Y, Goshima T, Kitamura K, Ueno M, Kanai M, Hirata D, et al:
The essential function of Rrs1 in ribosome biogenesis is conserved
in budding and fission yeasts. Yeast. 32:607–614. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Carnemolla A, Fossale E, Agostoni E,
Michelazzi S, Calligaris R, De Maso L, Del Sal G, MacDonald ME and
Persichetti F: Rrs1 is involved in endoplasmic reticulum stress
response in Huntington disease. J Biol Chem. 284:18167–18173. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Miyoshi K, Tsujii R, Yoshida H, Maki Y,
Wada A, Matsui Y, Toh-E A and Mizuta K: Normal assembly of 60 S
ribosomal subunits is required for the signaling in response to a
secretory defect in Saccharomyces cerevisiae. J Biol Chem.
277:18334–18339. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Morita D, Miyoshi K, Matsui Y, Toh-E A,
Shinkawa H, Miyakawa T and Mizuta K: Rpf2p, an evolutionarily
conserved protein, interacts with ribosomal protein L11 and is
essential for the processing of 27 SB Pre-rRNA to 25 S rRNA and the
60 S ribosomal subunit assembly in Saccharomyces cerevisiae.
J Biol Chem. 277:28780–28786. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nariai M, Tanaka T, Okada T, Shirai C,
Horigome C and Mizuta K: Synergistic defect in 60S ribosomal
subunit assembly caused by a mutation of Rrs1p, a ribosomal protein
L11-binding protein, and 3′-extension of 5S rRNA in
Saccharomyces cerevisiae. Nucleic Acids Res. 33:4553–4562.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Miyoshi K, Shirai C, Horigome C, Takenami
K, Kawasaki J and Mizuta K: Rrs1p, a ribosomal protein L11-binding
protein, is required for nuclear export of the 60S pre-ribosomal
subunit in Saccharomyces cerevisiae. FEBS Lett. 565:106–110.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lois C, Hong EJ, Pease S, Brown EJ and
Baltimore D: Germline transmission and tissue-specific expression
of transgenes delivered by lentiviral vectors. Science.
295:868–872. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Laemmli UK: Cleavage of structural
proteins during the assembly of the head of bacteriophage T4.
Nature. 227:680–685. 1970. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang C, Liu K, Li T, Fang J, Ding Y, Sun
L, Tu T, Jiang X, Du S, Hu J, et al: miR-21: A gene of dual
regulation in breast cancer. Int J Oncol. 48:161–172. 2016.
View Article : Google Scholar : PubMed/NCBI
|