Introduction
Colorectal cancer (CRC) is one of the most commonly
diagnosed cancer and one of the leading causes of cancer mortality
worldwide (1). Metastasis accounts
for ~90% of patient deaths and is the most significant contributor
to cancer death (2,3). CRC is often diagnosed in its advanced
stage accompanied by metastasis, which is negatively correlated
with patient survival (4). Since
advanced metastatic CRC remains largely incurable, it is urgent to
discover novel biomarkers which can influence its progression and
contribute to diagnosis and treatment in early stage.
The tripartite motif (TRIM) proteins represent a
large protein family comprising >70 members (5), constituted with a common N-terminal
really interesting new gene (RING) finger domain, 1 or 2 B-box
motifs and coiled-coil sequences, and most of them functioned as E3
ubiquitin ligases (6). According to
previous studies, TRIM is implicated in many critical processes,
such as viral infections, developmental and neurodegenerative
disorders and cancer (5,7). In recent years, many scholars have
found that TRIM proteins act as oncogenes or tumor suppressors in
cancer growth. For example, TRIM13, TRIM19, TRIM24 and TRIM25 have
been linked to leukemia, breast and prostate cancer (8), respectively. TRIM59, through the
regulation of transcriptional factors or tumor suppressors, is
strongly associated with gastric tumor (9), NSCLC (10) and renal cell cancer (11), but the function of the TRIM59 in CRC
has not yet been reported.
In this study, we analyzed TRIM59 from independent
Gene Expression Omnibus (GEO) datasets (www.ncbi.nlm.nih.gov/gds/) to determine the
characteristic expression of TRIM59 in CRC. We found that TRIM59 is
upregulated in CRC and significantly shortens patient survival
through regulating cell proliferation, motility, cell cycle and
inhibiting apoptosis of CRC cells.
Materials and methods
Patients and immunohistochemistry
Institutional Review Board (IRB)/Ethics Committee
approval was obtained and the study was in adherence with the
Declaration of Helsinki. The CRC samples and paired non-neoplastic
tissues were obtained from Renmin Hospital of Wuhan University, and
the study was approved by the local ethics committee. Before use,
all cases were diagnosed by two pathologists without discrepancy.
Formalin-fixed, paraffin-embedded CRC and non-neoplastic epithelial
tissues were cut into 4 µm sections. The sections were
deparaffinized in xylene and rehydrated in a series of ethanol of
descending concentrations. For antigen unmasking, sections were
immersed in antigen-unmasking solution and boiled in a microwave
oven for 15 sec. Tissue sections were incubated with the TRIM59
primary antibody at room temperature for 60 min, followed by the
standard procedure for the S-P immunohistochemical kit (Fujian
Maixin Biological Technology, Co., Ltd., Fuzhou, China). Expression
level of TRIM59 was ascertained according to the evaluations of the
pathologists. The data of mRNA expression profile GSE14333
(12), GSE17536 (13), GSE4107 (14) and GSE32323 (15) were downloaded from the GEO
database.
Cell culture, small interfering RNA
and antibodies
HCT116, SW480, SW620, HT29, Caco2 and NCM460 cells
were maintained in PRMI-1640 medium (Invitrogen) supplemented with
10% fetal bovine serum (FBS), 100 U/ml penicillin and 100 mg/ml
streptomycin. Cells were cultured at 37°C in 5% CO2. Two
different small interfering RNA (siRNA) oligo duplexes were
produced by Invitrogen (Carlsbad, CA, USA). The target sequences
for the TRIM59 gene were: siRNA-1: GCCUCUCUAUCUGUUUACCAAAGUU;
siRNA-2: UCC UCGUGUACUGCCAUGCUCUCAU; The negative control from
Invitrogen was used as a control. The specific siRNA or negative
control was transfected into CRC cells with Lipofectamine 2000
transfection reagent (Invitrogen) according to the manufacturers
instructions. Primary antibody TRIM59 was purchased from Abcam
(ab69639) and anti-caspase-3 (25546–1-AP), PARP (13371–1-AP),
cyclin B1 (55004–1-AP), cyclin D1 (0186–1-Ig), CDC25C (16485–1-AP),
MMP2 (10373–2-AP) were purchased from Proteintech (Rosemont, IL,
USA).
Western blot analysis
CRC proteins were separated by SDS-PAGE. Proteins
were electro-blotted onto a polyvinylidene fluoride membrane and
were blocked for 30 min in TBST [0.05% Tween-20 in phosphate
buffered saline (PBS)] with 5% dried skimmed milk at room
temperature. Immunoblot analysis was performed using primary
antibodies overnight at 4°C. Membranes were then washed with TBST
and incubated with horseradish peroxidase (HRP)-labeled secondary
antibody in TBST for 45 min and then visualized using a
chemiluminescent ECL detection kit (Santa Cruz Biotechnology, Santa
Cruz, CA, USA). GAPDH was used as a control for protein
loading.
RNA isolation and real-time PCR
analysis
Total RNA was isolated from cell lines or tissues
with TRIzol reagents (Invitrogen) according to the manufacturers
instructions. Quatitative real-time PCR (qRT-PCR) was performed to
quantify mRNA expression with SYBR® Green PCR Master Mix
(Takara Bio, Shiga, Japan). GAPDH was used for TRIM59
normalization. Primers were as follow: TRIM59: forward,
5-TACGAGAGCAGCAGCTTGAA-3 and reverse, 5-ACGG GTTGAACCTCAGGAAG-3;
GAPDH: forward, 5-GTGGA CATCCGCAAAGAC-3 and reverse, 5-AAAGGGTGTAAC
GCAACTA-3.
Cell proliferation assay
Cells were plated in 96-well plates and examined at
12, 24, 48 and 72 h after plating (n=6). Cells were incubated with
the Cell Counting kit-8 (CCK-8; Dojindo Laboratories, Kumamoto,
Japan) for 1 h. Then the absorbance was measured with the
multifunctional microplate reader at 450 nm.
Cell cycle analysis
Cells were harvested and resuspended twice with PBS.
After centrifugation, cells were fixed in 70% ice-cold methanol for
at least 2 h at −20°C. Then the fixed cells were stained with
propidium iodide (PI) (50 µl/ml PI and 250 µl/ml RNase A) for 30
min. In each experiment, 2×105 cells were analyzed using
BD FACSAria (BD Biosciences, San Jose, CA, USA). Experiments were
performed in triplicate. ModFit 2.0 software was used to determine
the percentage of cells in G0/G1, S and G2/M phases.
Apoptosis analysis
Annexin V-FITC/PI staining was performed to
investigate whether TRIM59 regulates the apoptosis of CRC cells.
The cells of each group were stained with Annexin V-FITC/PI
apoptosis detection kit (Invitrogen), according to the
manufacturer's instructions. Experiments were performed in
triplicate. FlowJo 7.6.1 software was used to determine the
percentage of apoptotic cells.
In vitro migration and invasion
assays
In the scratch wound healing assay, cells were
cultured in serum-free medium for 24 h and wounded with pipette
tips. Then the medium was refreshed. The wound closing procedure
was observed 36 h later and images were taken. In vitro
tumor cell migration was measured using Transwell chambers
(Corning, Inc., Corning, NY, USA) according to the manufacturers
instruction. In brief, 2×105 cells with/without siRNA in
2% FBS RPMI-1640 medium were plated in the upper chamber and
incubated with RPMI-1640 medium containing 10% FBS in the bottom
chamber for 48 h. Subsequently, the number of cells on the bottom
surface, which migrated across the membranes, were counted after
staining by 0.1% crystal violet dye. Cell counting was carried out
in five separate areas under a microscope. The invasion assay was
the same except that Matrigel basement was contained in the BD
BioCoat Matrigel Transwell Chambers (BD Biosciences, Bedford, MA,
USA). The data are presented as mean ± standard deviation (SD).
Statistical analysis
Survival analyses were carried out using the
Kaplan-Meier method and the results were statistically analysed by
the log-rank test. The correlations between the gene expression and
the clinicopathological features were analyzed by the Chi-square
test. A significant difference was defined as P<0.05.
Statistical analyses were performed using the GraphPad Prism
(v.6.0).
Results
TRIM59 is upregulated in CRC on both
mRNA and protein levels
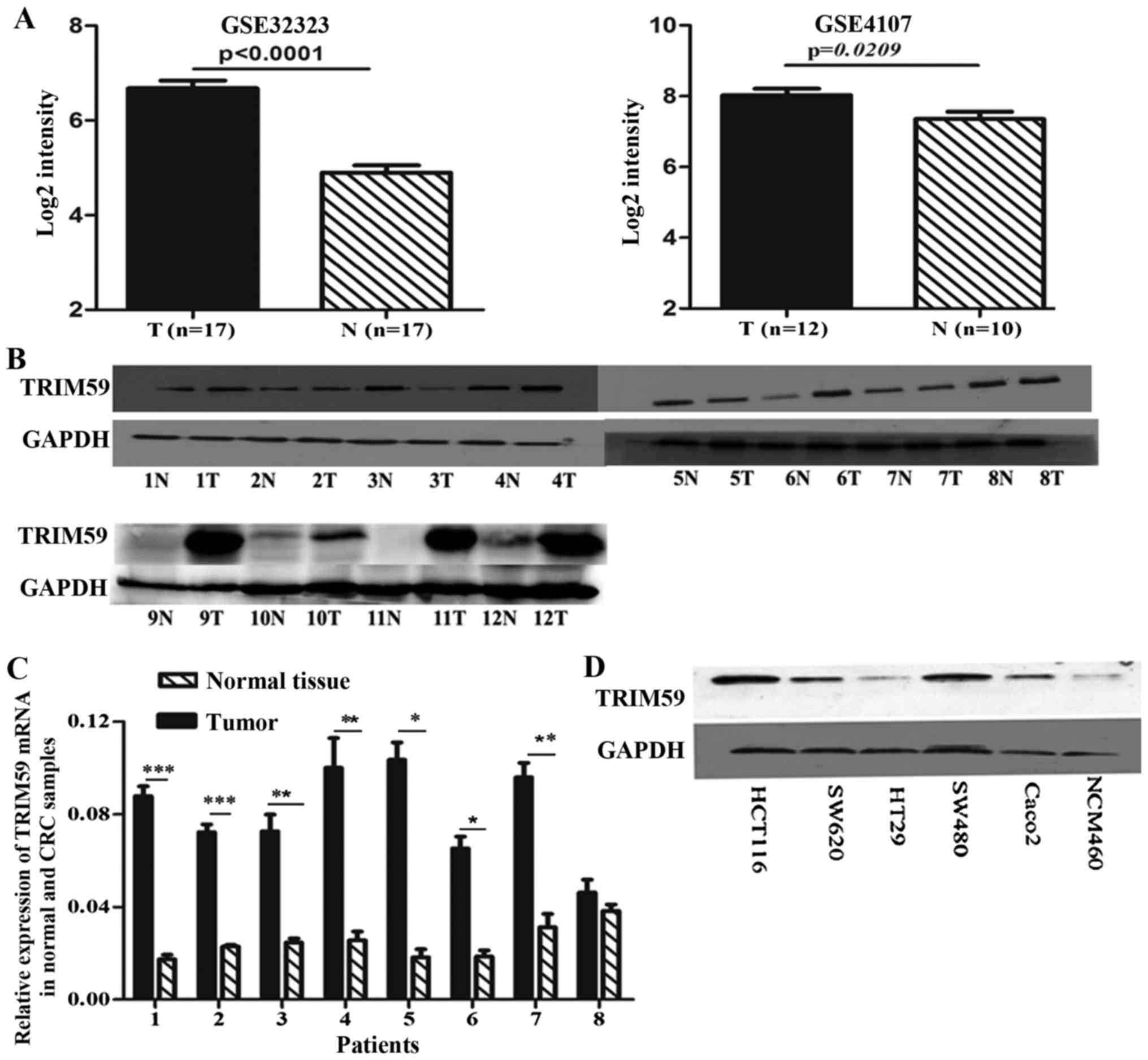
To determine the significance of TRIM59 in CRC, we
first analyzed the expression of TRIM59 at the mRNA level in the
cohorts of CRC patients available from two independent GEO datasets
GSE4107 and GSE32323. The two datasets involve paired CRC tumors
and adjacent non-cancerous tissue mRNA information. We chose all
the data of TRIM59 mRNA for statistical analyses. As shown in
Fig. 1A, TRIM59 mRNA levels
increased significantly in human colorectal tumor samples as
compared with the adjacent normal colorectal tissues.
To verify the microarray analysis results, we
performed immunoblot and qRT-PCR experiments on human colorectal
adenocarcinoma specimens and their matched normal tissues. Seven of
12 tumor samples showed increased protein level of TRIM59 compared
with their respective paired normal tissues (Fig. 1B). In addition, qRT-PCR experiments
found significantly higher TRIM59 mRNA expression in the tumor
tissues (Fig. 1C). Expression of
TRIM59 was determined by western blot analysis in several CRC cell
lines, including HCT116, SW480, SW620, HT29, Caco2 and the normal
colon cell line NCM460. According to the tissue results,
significantly higher expression of TRIM59 was detected in most CRC
cells, compared to NCM460 cells (Fig.
1D). To further investigate the correlation of TRIM59 with CRC
progression, we also examined the expression levels of TRIM59 in 80
human CRC tissues (60 cases of colon cancer and 20 cases of rectal
carcinoma) and 16 normal colon tissues by immunohistochemical
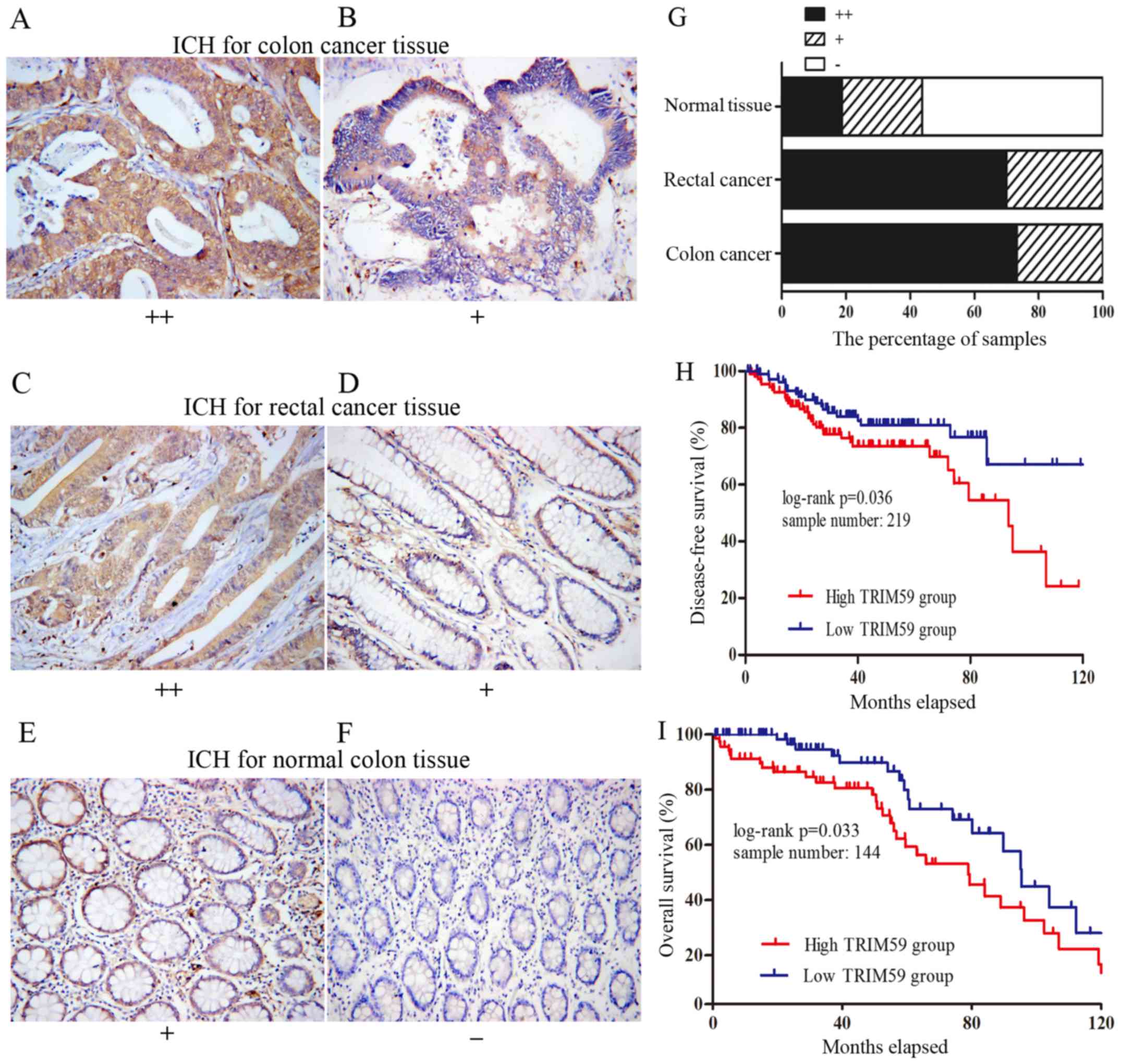
staining. TRIM59 was found mainly in the cytoplasm (Fig. 2A-E), which was consistent with
previous reports (9,16). The percentage of strongly positive
samples in cancer tissues (72.5%, 58/80) was significantly higher
than that in normal colon tissues (18.75%, 3/16) (P<0.0001,
Yates corrected χ2 analysis). Of the 60 colon cancer
samples, 44 (73.3%) were strongly positive and 16 (26.7%) were
weakly positive (Fig. 2A and B).
Whereas, 14 (70%) of the rectal cancer samples were strongly
positive and only 6 (30%) were weakly positive (Fig. 2C and D). In contrast, of the 16
normal tissue samples, 9 (56.25%) were negative, 4 (25%) were
weakly positive and only 3 (27.9%) were strongly positive (Fig. 2E and F). Distributions of TRIM59
staining grades are shown in Fig.
2G. Our results implied that TRIM59 expression was upregulated
at the protein level in CRC.
These data evidently demonstrated that TRIM59 was
upregulated in CRC in both mRNA and protein levels, implying its
importance in CRC pathogenesis.
The expression level of TRIM59 is
correlated with disease progression as well as shortened patient
survival
In order to confirm the correlations between the
TRIM59 expression level and the clinicopathological factors in CRC,
we downloaded clinical information for GSE14333 and analyzed it
statistically. The samples pooled in the dataset were then
classified into two groups according to the TRIM59 expression level
in tumor tissue and the χ2 test was applied. As shown
(Table I), higher TRIM59 expression
was closely associated with Dukes stage (P=0.004). The results
indicated that high expression of TRIM59 is related to rapid
carcinoma spread. Moreover, we downloaded clinical information and
performed Kaplan-Meier survival analysis. The two datasets
(GSE17536 and GSE14333) contain mRNA information of CRC and
disease-free survival (DFS) or overall survival (OS). We chose all
the samples that contained complete TRIM59 mRNA and survival
information for analysis. The results showed that CRC patients with
tumors displaying high TRIM59 expression level had significantly
shorter OS and DFS compared to those with tumors with low TRIM59
expression (Fig. 2H and I) (P=0.033
and P=0.036, respectively). The results strongly suggested that
TRIM59 may act as an oncogene in CRC and could represent a new
potential prognostic factor for CRC after curative colorectal
resection.
 | Table I.Correlations between the TRIM59
expression and the clinicopathological features of colorectal
carcinoma (GSE14333). |
Table I.
Correlations between the TRIM59
expression and the clinicopathological features of colorectal
carcinoma (GSE14333).
|
| TRIM59
expression |
|
|
|
|---|
|
|
|
|
|
|
|---|
| Characteristics | No. of patients | High | Low | Chi-square value | P-value |
|---|
| Age (years) |
|
|
| 0.258 | 0.612 |
|
>60 | 200 | 98 | 102 |
|
|
|
≤60 | 90 | 47 | 43 |
|
|
| Sex |
|
|
| 3.593 | 0.058 |
|
Male | 164 | 74 | 90 |
|
|
|
Female | 126 | 71 | 55 |
|
|
| Location |
|
|
| 1.466 | 0.48 |
|
L-colon | 138 | 64 | 74 |
|
|
|
R-colon | 111 | 60 | 51 |
|
|
|
Rectum | 39 | 19 | 20 |
|
|
| Dukes stage |
|
|
| 13.067 | 0.004 |
| A | 44 | 18 | 26 |
|
|
| B | 94 | 40 | 54 |
|
|
| C | 91 | 51 | 40 |
|
|
| D | 61 | 42 | 19 |
|
|
TRIM59 modulates CRC cell
proliferation
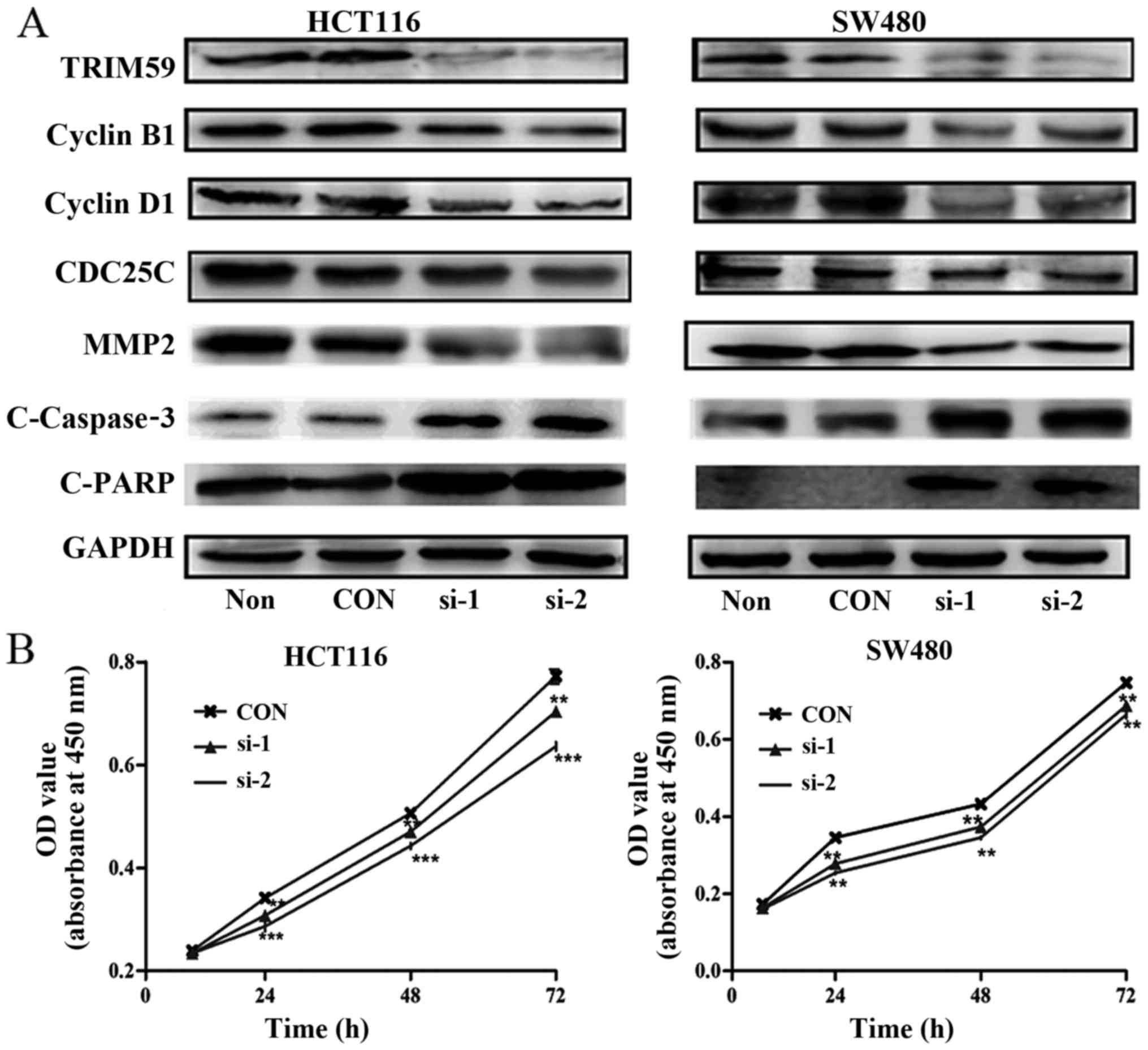
To validate the biological role of TRIM59 in the
proliferation of CRC cells, TRIM59 was depleted using three siRNAs
in HCT116 and SW480 cells, which exhibit a higher expression of
TRIM59. After transfecting the three siRNAs into CRC cells, the
protein level of TRIM59 was affirmed by western blot analysis. As
shown in the Fig. 3A, both siRNA-1
and siRNA-2 efficiently knocked down TRIM59 in the two cell lines
compared with that of control siRNA. Next, we examined the effects
of siRNA-induced knockdown of TRIM59 on the growth of CRC cell
lines. As shown in Fig. 3B, HCT116
and SW480 cells displayed a lower cell proliferation rate than
control cells, there was statistical significant difference
(P<0.05). These data suggested that TRIM59 is closely associated
with the proliferation of CRC cells.
TRIM59 modulates cell migration and
invasion of CRC cells
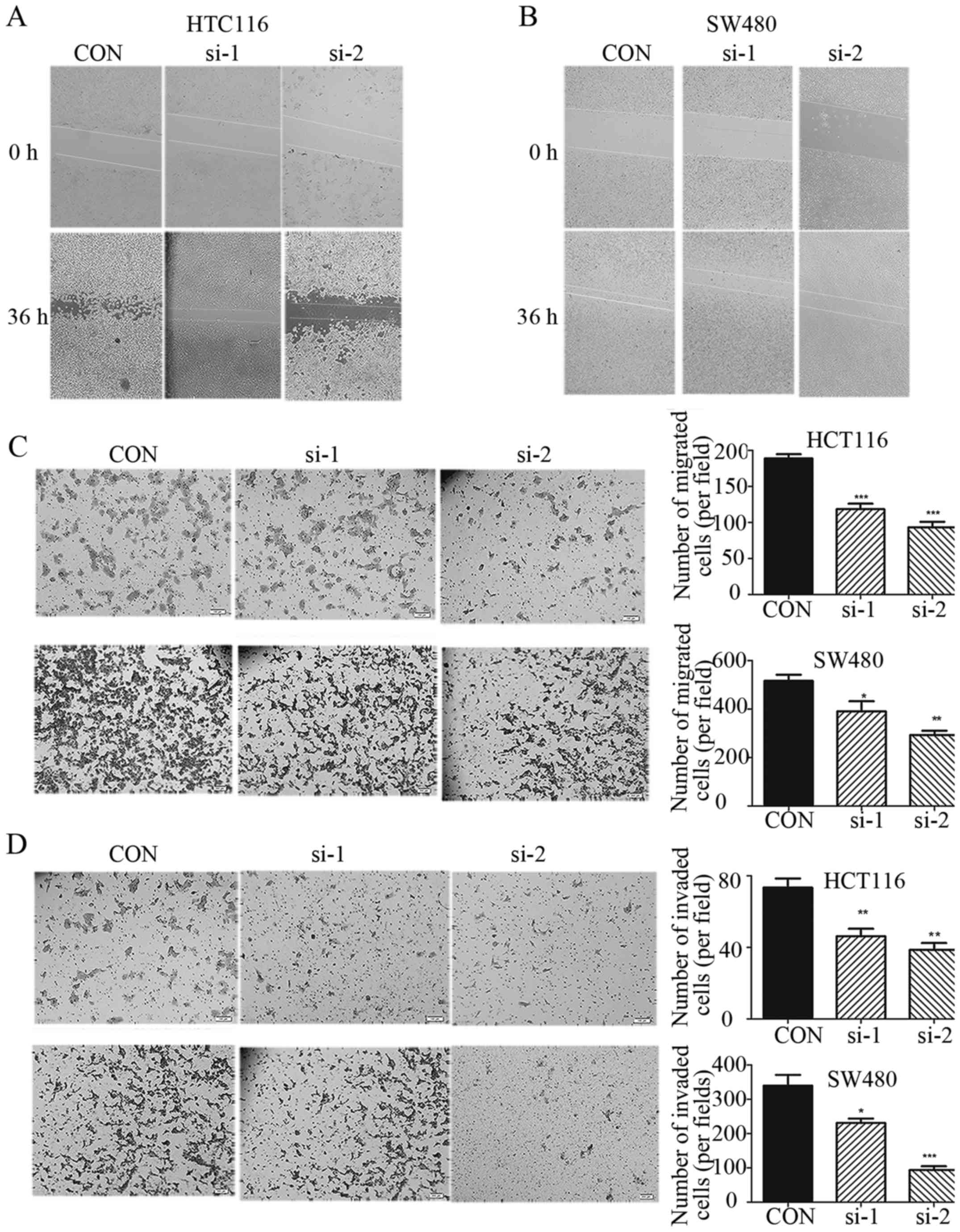
The role of TRIM59 in modulating CRC cell
proliferation is in agreement with previous reports in cervical
(17) and prostate cancer (16). In addition to cell proliferation,
the role of TRIM59 in cancer metastasis has not been well
characterized. We examined whether TRIM59 have an impact on cell
migration and invasion. As shown in Fig. 4A and B, we performed a wound healing
assay. The ‘wound’ was almost healed after 36 h in control cells.
However, the healing of the open area was markedly attenuated when
TRIM59 was knocked down. In order to further prove this effect, we
also performed a Transwell assay. Importantly, knockdown of TRIM59
suppressed the migration and invasion rates of HCT116 and SW480
cells (Fig. 4C and D). It is well
known that MMPs, especially MMP-2, is significantly involved in the
invasion and metastasis of human tumors (18). When TRIM59 is knocked down in HCT116
and SW480 cells, MMP2 is downregulated (Fig. 3A). TRIM59 modulates CRC cell
migration and invasion. This may be by regulating the expression of
MMP-2. These observations suggest that knockdown of TRIM59
significantly inhibits cell migration and invasion abilities in
HCT116 and SW480 cells and upregulation of TRIM59 may have
important consequences on the metastasis of CRC.
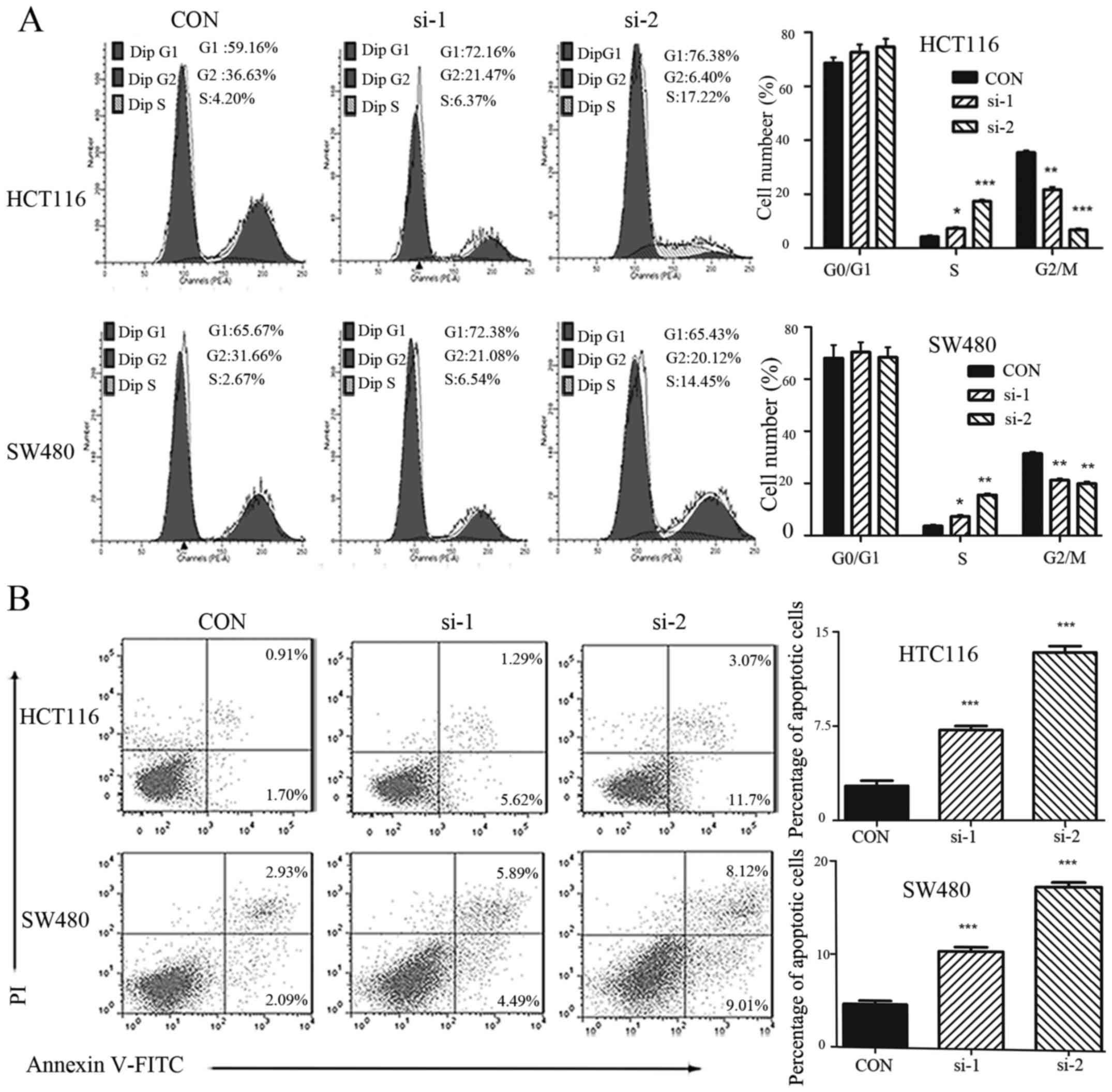
Knockdown of TRIM59 arrests cell cycle
in S phase
Cell proliferation depends largely on cell cycle
progression. Hence, the effect of TRIM59 knockdown on cell cycle
progression was also assessed using flow cytometry. After treatment
with si-TRIM59 or control siRNA for 48 h, cells were collected and
stained by PI. TRIM59 knockdown led to a significant accumulation
of cells at the S-phase with cell proportion in G2/M phase largely
decrease in HCT116 and SW480 cells (Fig. 5A). These data suggest that knockdown
of TRIM59 interrupts cell cycle progression leading to the
accumulation of cells in S phase. We proceeded to detect the
expression levels of cyclins, which are key cell cycle regulators
(19). As shown in Fig. 3A, the cyclin family proteins cyclin
D1, CDC25C and cyclin B1 were downregulated following TRIM59
knockdown, while the level of the internal control GAPDH remained
stable. The regulation of cyclins by TRIM59 might underlie the
TRIM59-mediated cell cycle arrest in CRC cells.
Downregulation of TRIM59 causes
apoptosis in CRC cells
Inhibiting apoptosis is crucial for sustaining
proliferation. To further explore how TRIM59 regulates the
proliferation of CRC cells, we investigated the contribution of
apoptosis to growth inhibition in SW480 and HCT116 cells with or
without TRIM59 knockdown. As expected, the apoptotic indexes of
TRIM59-silenced cells and control cells were 7.19% (si-1), 13.37%
(si-2) and 2.76% (control), (HCT116, P<0.05), 10.39% (si-1),
17.25% (si-2) and 4.66% (control) (SW480, P<0.05) (Fig. 5B), suggesting that silencing TRIM59
significantly promoted cell apoptosis.
Cell apoptosis is closely related to the regulation
of apoptosis-related genes including Bcl-2 and caspase gene
families (20). We measured
apoptotic markers and the cleavage of caspase-3 and PARP
(indicating a pro-apoptotic phenotype). These were higher when
TRIM59 was depleted (Fig. 3A).
These results revealed that TRIM59 may exert an
inhibitory effect on the caspase-dependent apoptosis pathway.
Discussion
The present study indicates that TRIM59 promotes CRC
progression, which is similar to previous studies in other cancer
types such as NSCLC (10), gastric
cancer (9,21), renal cell carcinoma (11) and osteosarcoma (22). Significantly shortened survival is
seen in patients with high TRIM59 expression compared with those
with low TRIM59 expression. Our data showed knockdown of TRIM59
inhibited the cell cycle and promoted apoptosis in CRC cells in
vitro, which may result in slower cancer cell proliferation.
However, the underlying mechanism still remains unclear, and here
several possible explanations are proposed.
Firstly, a hallmark of cancer cells is sustaining
proliferative signaling, this can be achieved by inactivation of
tumor suppressor genes. p53, a widely known tumor suppressor
(23). At present, whether p53 is
regulated by TRIM59 remain controversial. Some authors hold that
p53 is negatively regulated by TRIM59 via ubiquinitation, by which
TRIM59 promotes tumor growth, cell proliferation and migration
(9). Hence, the SV40 Tag/p53/pRB
routes can be considered one mechanism for TRIM59 functioning in
the process of tumorigenesis. However, others believed that TRIM59
may promote cancer cell growth through other pathways but not the
p53 signaling pathway (10). In
this study, HCT116 is a p53 wild-type; in contrast, SW480 is mutant
p53 cell line. When knocking down TRIM59, the two different cells
have similar response, inferring that possibly TRIM59 is not
mediated through p53, so further research is needed. In addition,
the TRIM family is an evolutionarily conserved gene family
implicated in a number of critical processes including
transcriptional regulation (24,25).
The domain of the really interesting new gene (RING) is frequently
involved in proteolysis, acting as E3 ubiquitin ligase and the
ubiquitin-proteasome system in the regulation of numerous cellular
processes including cell cycle regulatory proteins, transcription
factors and signal transducers (26). Valiyeva et al (27) recently identified TRIM59 as an early
signal transducer in two (SV40Tag and Ras) oncogene pathways in
murine prostate cancer models. High expression level of TRIM59 may
activate the transcription of some crucial downstream oncogenes,
which, in turn promotes the progression of cancers. Furthermore, it
is reported that TRIM59 is involved in NF-κB and IRF3/IRF7 mediated
signaling pathways (28), both of
which are dysregulated in nearly all tumors and considered to be
driver genes of cancer. We hypothesize that high expression of
TRIM59 will regulate NF-κB and IRF3/IRF7 pathways in CRC, which
remains to be validated. Lastly, c-Myc overexpression and DNA
promoter hypermethylation repress the expression of TRIM59 in
cancers (29). These reports
suggest several possible mechanisms by which TRIM59 may regulate
cell cycle progression in colorectal cancer.
In the present study, we demonstrated that TRIM59
modulates not only proliferation, but also motility, migration and
invasion. In CRC, metastasis accounts for ~90% of patient deaths,
representing the most lethal event during the course of the disease
(30). Metastasis is directly
linked to patient survival, and critically limits successful
therapy (31,32). Metastasis formation is a major
hurdle in CRC therapy. Therefore, to identify patients at high risk
for metastasis formation, early diagnosis and molecular
characterization of the primary tumor is crucial to define
prognostic and therapeutic targets (33). Our results indicated that TRIM59 is
an important target for blocking metastasis in CRC. We propose that
quantification of TRIM59 in colon biopsies could be used in
combination with pathologic examination to predict biological
behavior of the CRC. The molecular pathologic diagnosis will be
helpful in personalized treatment optimization.
In summary, the present study has shown the
biological and clinical significance of TRIM59 in CRC. This study
demonstrated that TRIM59 expression is significantly upregulated in
CRC tissues and associated with tumor Dukes stage. Knockdown of
TRIM59 with specific siRNA inhibited cancer cell proliferation and
led to cell cycle arrest in the S-phase. Regulation of cell cycle
progression and promotion of apoptosis may underlie TRIM59-mediated
CRC development. Our evidence provides novel clues that may aid CRC
diagnosis and treatment in the future. However, further in depth
exploration of the molecular mechanisms of TRIM59 in promoting
proliferation and metastasis will be needed.
Acknowledgements
The authors would like to thank the anonymous
reviewers for their helpful comments. The authors also would like
to thank Dr Katelyn ONeill from the University of Nebraska Medical
Center for improving the language of this manuscript.
References
|
1
|
Terzic J, Grivennikov S, Karin E and Karin
M: Inflammation and colon cancer. Gastroenterology.
138:2101–2114.e5. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stein U and Schlag PM: Clinical,
biological, and molecular aspects of metastasis in colorectal
cancer. Recent Results Cancer Res. 176:61–80. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li W, Cai S, Wang L, Yang C, Zhou B and
Wang H: HINT2 downregulation promotes colorectal carcinoma
migration and metastasis. Oncotarget. 8:13521–13531.
2017.PubMed/NCBI
|
|
4
|
Mathot L, Kundu S, Ljungström V, Svedlund
J, Moens L, Adlerteg T, Falk-Sörqvist E, Rendo V, Bellomo C,
Mayrhofer M, et al: Somatic ephrin receptor mutations are
associated with metastasis in primary colorectal cancer. Cancer
Res. 77:1730–1740. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Elabd S, Meroni G and Blattner C: TRIMming
p53s anticancer activity. Oncogene. 35:5577–5584. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ozato K, Shin DM, Chang TH and Morse HC
III: TRIM family proteins and their emerging roles in innate
immunity. Nat Rev Immunol. 8:849–860. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tomar D and Singh R: TRIM family proteins:
Emerging class of RING E3 ligases as regulator of NF-κB pathway.
Biol Cell. 107:22–40. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hatakeyama S: TRIM proteins and cancer.
Nat Rev Cancer. 11:792–804. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou Z, Ji Z, Wang Y, Li J, Cao H, Zhu HH
and Gao WQ: TRIM59 is up-regulated in gastric tumors, promoting
ubiquitination and degradation of p53. Gastroenterology.
147:1043–1054. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhan W, Han T, Zhang C, Xie C, Gan M, Deng
K, Fu M and Wang JB: TRIM59 promotes the proliferation and
migration of non-small cell lung cancer cells by upregulating cell
cycle related proteins. PLoS One. 10:e01425962015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Khatamianfar V, Valiyeva F, Rennie PS, Lu
WY, Yang BB, Bauman GS, Moussa M and Xuan JW: TRIM59, a novel
multiple cancer biomarker for immunohistochemical detection of
tumorigenesis. BMJ Open. 2:e0014102012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jorissen RN, Gibbs P, Christie M, Prakash
S, Lipton L, Desai J, Kerr D, Aaltonen LA, Arango D, Kruhøffer M,
et al: Metastasis-associated gene expression changes predict poor
outcomes in patients with Dukes stage B and C colorectal cancer.
Clin Cancer Res. 15:7642–7651. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Freeman TJ, Smith JJ, Chen X, Washington
MK, Roland JT, Means AL, Eschrich SA, Yeatman TJ, Deane NG and
Beauchamp RD: Smad4-mediated signaling inhibits intestinal
neoplasia by inhibiting expression of β-catenin. Gastroenterology.
142:562–571.e2. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hong Y, Ho KS, Eu KW and Cheah PY: A
susceptibility gene set for early onset colorectal cancer that
integrates diverse signaling pathways: Implication for
tumorigenesis. Clin Cancer Res. 13:1107–1114. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Khamas A, Ishikawa T, Shimokawa K, Mogushi
K, Iida S, Ishiguro M, Mizushima H, Tanaka H, Uetake H and Sugihara
K: Screening for epigenetically masked genes in colorectal cancer
Using 5-Aza-2-deoxycytidine, microarray and gene expression
profile. Cancer Genomics Proteomics. 9:67–75. 2012.PubMed/NCBI
|
|
16
|
Lin WY, Wang H, Song X, Zhang SX, Zhou PS,
Sun JM and Li JS: Knockdown of tripartite motif 59 (TRIM59)
inhibits tumor growth in prostate cancer. Eur Rev Med Pharmacol
Sci. 20:4864–4873. 2016.PubMed/NCBI
|
|
17
|
Aierken G, Seyiti A, Alifu M and Kuerban
G: Knockdown of tripartrtite-59 (TRIM59) inhibits cellular
proliferation and migration in human cervical cancer cells. Oncol
Res. 25:381–388. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xing P, Liao Z, Ren Z, Zhao J, Song F,
Wang G, Chen K and Yang J: Roles of low-density lipoprotein
receptor-related protein 1 in tumors. Chin J Cancer. 35:62016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Koosha S, Alshawsh MA, Looi CY, Seyedan A
and Mohamed Z: An association map on the effect of flavonoids on
the signaling pathways in colorectal cancer. Int J Med Sci.
13:374–385. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shu YJ, Weng H, Ye YY, Hu YP, Bao RF, Cao
Y, Wang XA, Zhang F, Xiang SS, Li HF, et al: SPOCK1 as a potential
cancer prognostic marker promotes the proliferation and metastasis
of gallbladder cancer cells by activating the PI3K/AKT pathway. Mol
Cancer. 14:122015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Luo D, Wang Y, Huan X, Huang C, Yang C,
Fan H, Xu Z and Yang L: Identification of a synonymous variant in
TRIM59 gene for gastric cancer risk in a Chinese population.
Oncotarget. 8:11507–11516. 2016.
|
|
22
|
Liang J, Xing D, Li Z, Shen J, Zhao H and
Li S: TRIM59 is upregulated and promotes cell proliferation and
migration in human osteosarcoma. Mol Med Rep. 13:5200–5206. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Charni M, Aloni-Grinstein R, Molchadsky A
and Rotter V: p53 on the crossroad between regeneration and cancer.
Cell Death Differ. 24:8–14. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gack MU, Shin YC, Joo CH, Urano T, Liang
C, Sun L, Takeuchi O, Akira S, Chen Z, Inoue S, et al: TRIM25
RING-finger E3 ubiquitin ligase is essential for RIG-I-mediated
antiviral activity. Nature. 446:916–920. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lerner M, Corcoran M, Cepeda D, Nielsen
ML, Zubarev R, Pontén F, Uhlén M, Hober S, Grandér D and Sangfelt
O: The RBCC gene RFP2 (Leu5) encodes a novel transmembrane E3
ubiquitin ligase involved in ERAD. Mol Biol Cell. 18:1670–1682.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Deshaies RJ and Joazeiro CA: RING domain
E3 ubiquitin ligases. Annu Rev Biochem. 78:399–434. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Valiyeva F, Jiang F, Elmaadawi A, Moussa
M, Yee SP, Raptis L, Izawa JI, Yang BB, Greenberg NM, Wang F, et
al: Characterization of the oncogenic activity of the novel TRIM59
gene in mouse cancer models. Mol Cancer Ther. 10:1229–1240. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kondo T, Watanabe M and Hatakeyama S:
TRIM59 interacts with ECSIT and negatively regulates NF-κB and
IRF-3/7-mediated signal pathways. Biochem Biophys Res Commun.
422:501–507. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Licchesi JD, Van Neste L, Tiwari VK, Cope
L, Lin X, Baylin SB and Herman JG: Transcriptional regulation of
Wnt inhibitory factor-1 by Miz-1/c-Myc. Oncogene. 29:5923–5934.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dahlmann M, Kobelt D, Walther W, Mudduluru
G and Stein U: S100A4 in cancer metastasis: Wnt signaling-driven
interventions for metastasis restriction. Cancers (Basel).
8:E592016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fodde R, Smits R and Clevers H: APC,
signal transduction and genetic instability in colorectal cancer.
Nat Rev Cancer. 1:55–67. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Stein U and Schlag PM: Clinical,
biological, and molecular aspects of metastasis in colorectal
cancer. Recent Results Cancer Res. 176:61–80. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dahlmann M, Okhrimenko A, Marcinkowski P,
Osterland M, Herrmann P, Smith J, Heizmann CW, Schlag PM and Stein
U: RAGE mediates S100A4-induced cell motility via MAPK/ERK and
hypoxia signaling and is a prognostic biomarker for human
colorectal cancer metastasis. Oncotarget. 5:3220–3233. 2014.
View Article : Google Scholar : PubMed/NCBI
|