Introduction
Glioma is the most common intracranial malignant
tumor. Owing to the frequent infiltrating nature of glioma lesions,
radical surgery is not always feasible. Radiotherapy and
chemotherapy are conventional treatments after surgery. However,
even after standard treatment, the median survival time of patients
remains only 12–15 months after glioblastoma diagnosis (1,2). The
poor prognosis of glioblastoma patients has aroused increasing
interest in finding new approaches to glioma therapy. Agents
directed at molecular targets, such as abnormal signaling pathway
or tumor angiogenesis inhibitors, have gained wide attention
(3–5). Due to the hypervascular nature of
glioblastoma (6), the
identification of anti-angiogenesis-targeted chemotherapeutic
agents by exploring the regulatory factors of tumor-suppressor
genes is promising for the treatment of glioma.
Angiogenesis refers to the formation of new blood
vessels from original ones, which is essential for tumor blood
supply and is involved in various malignant biological behaviors of
tumors including growth, invasion and metastasis (7). Vascular endothelial cell
proliferation, migration and tube formation constitute the main
steps of the angiogenesis process (8). In the glioma microenviroment, vascular
endothelial cells respond to multiple proangiogenetic regulatory
factors such as vascular endothelial cell growth factor (VEGF),
transforming growth factor α (TGFα) and endothelin-1 directly
secreted by tumor cells (9,10). They are also altered in the
endogenous gene expession, such as Roundabout4 and miR-125b, which
indirectly affects tumor angiogenesis (11,12).
Long non-coding RNAs (lncRNAs) are defined as
non-coding RNAs between 200 nt and 100 kb in length. lncRNAs play
important roles in diverse biological processes, including
embryonic development, cell growth and tumorigenesis, by regulating
gene expression at the chromatin organization, transcriptional and
post-transcriptional levels (13).
Research has shown that abnormalities in lncRNAs are linked to
various tumor and non-tumor diseases. For example, expression of
lncRNA-HOTAIR was found to be upregulated in multiple malignant
tumors including colorectal cancer, hepatocellular carcinoma,
pancreatic and prostate cancer as well as glioma, and showed a
positive correlation with malignant biological behaviors of the
tumors (14–18). Recent studies also found lncRNAs to
be involved in the modulation of angiogenesis. Located in 7p13,
SNHG15 was found to be highly expressed in hepatocellular carcinoma
and gastric cancer (19,20). Nonetheless, the expression and
biological effect of SNHG15 in glioma vascular endothelial cells
remain unknown.
MicroRNAs (miRNAs) are a class of small non-coding
RNAs (18–25 nucleotides) which post-transcriptionally regulate gene
expression by negatively regulating the stability or translational
efficiency of their target mRNAs (21). Increasing evidence has shown that
abnormal expression of miRNAs is involved in tumor formation,
progression as well as angiogenesis (22,23).
miR-153, involved in the development of a number of tumors, was
found to be downregulated in breast, prostate and non-small cell
lung cancer as well as glioma (24–27).
However, the function of miR-153 in glioma vascular endothelial
cells is still unclear.
VEGFA is a member of the PDGF/VEGF growth factor
family which encodes a heparin-binding protein that plays
predominant roles in angiogenesis by inducing proliferation and
migration of vascular endothelial cells. It is upregulated in many
known tumors and its expression is correlated with tumor stage and
progression (28–30). Cdc42 is a small GTPase of the
Rho-subfamily which regulates cytoskeletal dynamics, cell shape,
and numerous other cellular processes including polarity,
migration, cell cycle progression and cell fate determination. Both
VEGFA and Cdc42 have been confirmed to be closely related to
angiogenesis.
In the present study, we aimed at investigating the
expression levels of SNHG15 and miR-153 in glioma microvascular
endothelial cells and the effect of SNHG15 on miR-153-induced
regulation of VEGF and Cdc42 as well as the underlying mechanism in
the process.
Materials and methods
Reagents and antibodies
Endothelial Basal Medium-2 (EBM-2) was purchased
from Lonza (Walkersville, MD, USA). Fetal bovine serum (FBS) ‘Gold’
and 10 mM HEPES were purchased from PAA Laboratories GmbH
(Pasching, Austria). Penicillin-streptomycin and chemically defined
lipid concentrate were purchased from Invitrogen (Life
Technologies, Carlsbad, CA, USA). Cultrex rat collagen I was
obtained from R&D Systems (Minneapolis, MN, USA), Human basic
fibroblast growth factor (bFGF), hydrocortisone and ascorbic acid
were obtained from Sigma-Aldrich (St. Louis, MO, USA). High-glucose
Dulbecco's modified Eagles medium (DMEM), DMEM-F12 mixed medium and
FBS were purchased from Gibco (Life Technologies). Cell Counting
Kit-8 (CCK-8) was purchased from Beyotime Institute of
Biotechnology (Jiangsu, China). VEGFA and Cdc42 antibodies were
purchased from the ProteinTech Group (Chicago, IL, USA). Antibodies
against GAPDH and the secondary antibodies were purchased from
Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Cell culture and treatment
Human cerebral microvascular endothelial cells
(hCMECs)/D3 were provided by Dr P.-O. Couraud (Institut Cochin,
Paris, France). The cells (passage 30–35) were cultured in culture
flasks coated with 150 µg/ml of cultrex rat collagen I. Primary
glioma cells were derived from intraoperative tissue samples from
patients who were diagnosed with primary glioblastoma. Primary
astrocyte cells derived from patients undergoing temporal lobe
resection for the surgical treatment of intractable epilepsy were
used as the negative control (NC). Detailed culture methodology was
carried out as previously described (31,32).
Immunohistochemistry against GFAP and β-tubulin III was used for
identification of glioma (or astrocyte) cells. Cells were stained
with anti-GFAP (1:100; Abcam, Cambridge, MA, USA) and
anti-β-tubulin III (1:100). Cells were also counterstained with
4,6-diamidino-2-phenylindole (DAPI) to identify all nuclei.
Glioma conditioned medium (GCM) was collected from
the indicated primary human glioma cells grown in 100-mm-diameter
Petri dishes. Primary astrocyte cell conditioned medium (ACM) was
used as the NC. Conditioned medium was prepared from the
glioblastoma (or astrocyte) cell culture grown to near confluency.
After being washed twice with serum-free medium, the cells were
incubated in serum-free EBM-2 medium in a humidified incubator at
37°C with 5% CO2 for 24 h. The supernatant was
harvested, centrifuged at 2,000 × g at 4°C for 10 min and
supplemented with 5% FBS ‘Gold’, 1% penicillin-streptomycin, 1%
chemically defined lipid concentrate, 1 ng/ml bFGF, 1.4 µM
hydrocortisone, 5 µg/ml ascorbic acid and 10 mM HEPES, EGF and
hydrocortisone prior to use. Primary astrocyte cell conditioned
medium was used as NC.
Reverse transcription and quantitative
real-time PCR (qRT-PCR)
Total RNA was extracted from cells using TRIzol
reagent (Life Technologies). RNA concentration and quality were
determined by the 260/280 nm ratio using a NanoDrop
spectrophotometer (ND-100). One-Step SYBR PrimeScript RT-PCR kit
(Perfect Real-Time) (Takara Bio, Inc., Otsu, Japan) was used for
qRT-PCR. The primers for SNHG15 were: 5′-GCATCTCTTCCCACTATCTGC-3′
and 5′-TGGTTTCATCTCCCAGCAC-3′; and for glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) 5′-GGTGAAGGTCGGAGTCAACG-3′ and
5′-CCATGTAGTTGAGGTCAATGAAG-3′. TaqMan MicroRNA Reverse
Transcription kit (Applied Biosystems, Foster City, CA, USA) was
used for miRNA reverse transcription and qRT-PCR was conducted
using TaqMan Universal Master Mix II with TaqMan microRNA assays of
miR-153 and U6, respectively. U6 and GAPDH were used as endogenous
controls for miRNA and gene expression detection. Expression levels
were normalized to endogenous controls, and the relative
quantification (2−ΔΔCt) method was used for fold change
calculation.
Cell transfections
The sh-SNHG15 plasmid and its respective
non-targeting sequence (NC); pre-miR-153 and anti-miR-153 plasmid,
and their respective non-targeting sequences were synthesized
(GenePharma, Shanghai, China). Cells were seeded into 24-well
plates (Corning Inc., Corning, NY, USA) until they achieved 50–70%
confluence, and then transfected using Opti-MEM I and Lipofectamine
2000 reagent (Life Technologies Corporation, Carlsbad, CA, USA).
After 6 h of transfection, the medium was replaced with EBM-2
medium with 10% FBS. The applicable stably transfected cells were
selected using G418 screening. The overexpression and the silencing
efficiency were analyzed using qRT-PCR. To determine the effect of
SNHG15 on glioma vascular endothelial cells (ECs), cells were
divided into 3 groups: control, SNHG15(−) NC (transfected with
sh-NC plasmid) and SNHG15(−) group (transfected with sh-SNHG15
plasmid). To determine the effect of miR-153 on glioma vascular
ECs, cells were divided into 5 groups: control, miR-153(+) NC
(transfected with pre-miR-153 NC plasmid), miR-153(+) (transfected
with pre-miR-153 plasmid), miR-153(−) NC (transfected with
anti-miR-153-NC plasmid) and miR-153(−) group (transfected with
anti-miR-153 plasmid). To determine whether SNHG15-mediated
regulation of miR-153 expression regulated the behavior of ECs,
cells were divided into 5 groups: control, SNHG15(−) NC +
miR-153(+) NC group (transfected with both sh-NC plasmid and
pre-miR-153-NC plasmid), SNHG15(−) + miR-153(+) group (transfected
with both sh-SNHG15 plasmid and pre-miR-153 plasmid), SNHG15(−) NC
+ miR-153(−) NC group (transfected with both sh-NC plasmid and
anti-miR-153-NC plasmid), SNHG15(−) + miR-153(−) group (transfected
with both sh-SNHG15 plasmid and anti-miR-153 plasmid).
Cell proliferation assay
ECs were seeded into 96-well plates at the density
of 2,000 cells/well, and cell viability rate was assayed using
CCK-8 (Beyotime Institute of Biotechnology) according to the
instructions provided by the manufacturer. CCK-8 (10 µl) was added
into each well and cells were incubated for another 2 h in a
humidified incubator. Optical density value was measured at 450 nm.
Five replicate wells were set up in each group and 3 independent
repeated experiments were performed.
Cell migration assay
EC migration in vitro was assayed using a
Transwell chamber (Costar, Corning, NY, USA) with a polycarbonate
membrane (6.5-mm in diameter, 8-µm pore size). ECs were trypsinized
and suspended into single cells with serum-free medium at the
density of 5×105 cells/ml. Cell suspension (100 µl) was
added to the upper chamber, and 600 µl of GCM supplemented with 10%
FBS was added to the lower chamber. Cells were incubated for 12 h
at 37°C. Non-migrating cells on the top surface of the membrane
were removed with cotton swabs. Cells that migrated to the lower
surface of the membrane were fixed with methanol and glacial acetic
acid in a 3:1 ratio, stained with 20% Giemsa solution for 30 min at
37°C and were washed twice with phosphate-buffered saline (PBS).
Then, stained cells were observed under an inverted microscope to
count the cell number within five randomly chosen fields at a
magnification of ×200 and the average number was calculated.
Tube formation assay
We used standard Matrigel assay to evaluate in
vitro angiogenesis activity by quantifying the tube formation,
as previously described (11).
Culture plates (48-well) were coated with 200 µl of Matrigel (BD
Biosciences, Bedford, MA, USA) per well, and then allowed to
polymerize for 30 min at 37°C. Confluent cells were suspended in
fresh GCM at the density of 1.5×105 cells/ml. Cell
suspension (100 µl) was seeded in wells of a 48-well culture plate
polymerized Matrigel and incubated at 37°C for 12 h. Each culture
was photographed at a magnification of ×100 using Olympus DP71
immunofluorescence microscopy (Olympus, Tokyo, Japan), and total
tubule length and number of tubule branches were measured using
ChemiImager 5500 V2.03 software (Alpha Innotech).
Western blot assay
Cells were lysed and the supernatant extracts were
quantified for protein using the BCA protein assay kit (Beyotime
Institute of Biotechnology). Total cell lysates containing 40 µg of
protein were fractionated using SDS-PAGE and transferred onto
polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA,
USA). After blocking with 5% non-fat dry milk in Tris-buffered
saline with Tween-20 (TBST) for 2 h, the membranes were incubated
with primary antibodies (anti-GAPDH antibody diluted at 1:1,000;
anti-VEGFA antibody diluted at 1:500; anti-Cdc42 antibody diluted
at 1:500) at 4°C overnight. After 3 washes with PBS-Tween (20 mM
Tris, 137 mM NaCl, 0.1% Tween-20, pH 7.6), the membranes were
incubated with the corresponding HRP-conjugated secondary antibody
(diluted at 1:5,000) at room temperature for 2 h. Protein bands
were visualized by ECL (Santa Cruz Biotechnology) and detected
using the ECL Detection System (Thermo Scientific, Waltham, MA,
USA). Then, the protein bands were scanned using ChemiImager 5500
V2.03 software, and integrated light density values (IDVs) were
calculated by Fluor Chen 2.0 software and normalized with those of
GAPDH.
Reporter vector construction and
luciferase assays
SNHG15 3-untranslated region (3′-UTR), VEGFA 3′-UTR
and Cdc42 3′-UTR sequences were amplified by PCR and cloned into a
pmirGlo Dual-luciferase MiRNA Target Expression Vector (Promega,
Madison, WI, USA) to construct 3′-UTR-luciferase reporter vector
(SNHG15-WT, VEGFA-WT and Cdc42-WT) (GenePharma). The sequence of
the putative binding site was replaced as indicated (SNHG15-Mut,
VEGFA-Mut and Cdc42-Mut) to mutate the putative binding site of
miR-153 in the 3′-UTR-containing vector. Glioma vascular ECs were
seeded in 96-well plates and the cells were co-transfected with
SNHG15-WT (or SNHG15-Mut), VEGFA-WT (or VEGFA-Mut) or Cdc42-WT (or
Cdc42-Mut) and miR-153 plasmids (or miR-153-NC) when they reached
50–70% confluence. The luciferase activities were measured at 48 h
after transfection using the Dual-Luciferase Reporter Assay kit
(Promega). The cells were divided in 5 groups, respectively:
control, SNHG15-WT + miR-153(+) NC, SNHG15-WT + miR-153(+),
SNHG15-Mut + miR-153(+) NC and SNHG15-Mut + miR-153(+) group;
control, VEGFA-WT + miR-153(+) NC, VEGFA-WT + miR-153(+), VEGFA-Mut
+ miR-153(+) NC and VEGFA-Mut + miR-153(+) group; control, Cdc42-WT
+ miR-153(+) NC, Cdc42-WT + miR-153(+), Cdc42-Mut + miR-153(+) NC
and Cdc42-Mut + miR-153(+) group.
RNA-binding protein
immunoprecipitation (RIP) assay
RNA-RIP was assayed using a Magna RNA-RIP kit
(Millipore) according to the manufacturers instructions. Briefly,
the glioma vascular ECs were lysed in complete RNA lysis buffer.
Whole cell lysate was incubated with RIP buffer containing magnetic
beads conjugated with human anti-Argonaute2 (Ago2) antibody, and NC
normal mouse IgG (both from Millipore). Samples were incubated with
proteinase K, and then immunoprecipitated RNA was isolated. The RNA
concentration was measured using a spectrophotometer (NanoDrop;
Thermo Scientific) and the RNA quality assessed using a bioanalyzer
(Agilent, Santa Clara, CA, USA). Furthermore, purified RNAs were
extracted and analyzed by qRT-PCR to demonstrate the presence of
the binding targets.
In vivo Matrigel plug assay
All experimental animal procedures were strictly
conducted in accordance with the Guide for the Care and Use of
Laboratory Animals, and approved by the Animal Care and Use
Committee of Shengjing Hospital. The male BALB/c athymic nude mice
(randomized to each group by 3 observers in a blinded manner,
n=7/group, 4–6 weeks old) were obtained from the Cancer Institute
of the Chinese Academy of Medical Sciences. Angiogenesis was also
measured by a Matrigel plug assay as previously described (33). Briefly, 3×106 GECs in 400
µl of solution containing 80% Matrigel were subcutaneously
injected. Plugs were harvested after 4 days, weighed, photographed
and dispersed in 400 µl of PBS (overnight incubation at 4°C) to
collect the hemoglobin. Hemoglobin content was measured using
Drabkin's solution (Sigma-Aldrich) according to the manufacturer's
recommendations.
Patients and specimens
The present study was approved by the Institutional
Review Board of Shengjing Hospital of China Medical University and
each patient signed a consent form to participate in the present
study. Human glioma specimens were obtained from primary patients
diagnosed with glioma who underwent gross total tumor resection at
the Department of Neurosurgery of Shengjing Hospital, China Medical
University, from April 2010 to January 2011. None of these patients
had undergone chemotherapy or radiotherapy before surgery, and
specimens were collected immediately after tumor excision during
surgery. All glioma cases were pathologically confirmed. Patient
survival information was acquired via out-patient review, telephone
and mail. Overall survival was calculated from the date of the
initial surgical operation to death from primary tumor-related
causes or up to December 2013, whichever came first. The follow-up
rate was 100% and median follow-up period was 39 months (range,
4–60 months).
Statistical analysis
Statistical analysis was performed using SPSS 18.0
statistical software. One-way ANOVA followed by Bonferroni post hoc
test was used to analyze the difference between two groups. Data
are expressed as mean ± standard deviation (SD). P<0.05 was
considered to indicate a statistically significant result.
Results
SNHG15 is highly expressed in
glioma-induced hCMECs, VEGFA and Cdc42 are upregulated in
glioma-induced hCMECs while miR-153 is lowly expressed in
glioma-induced hCMECs
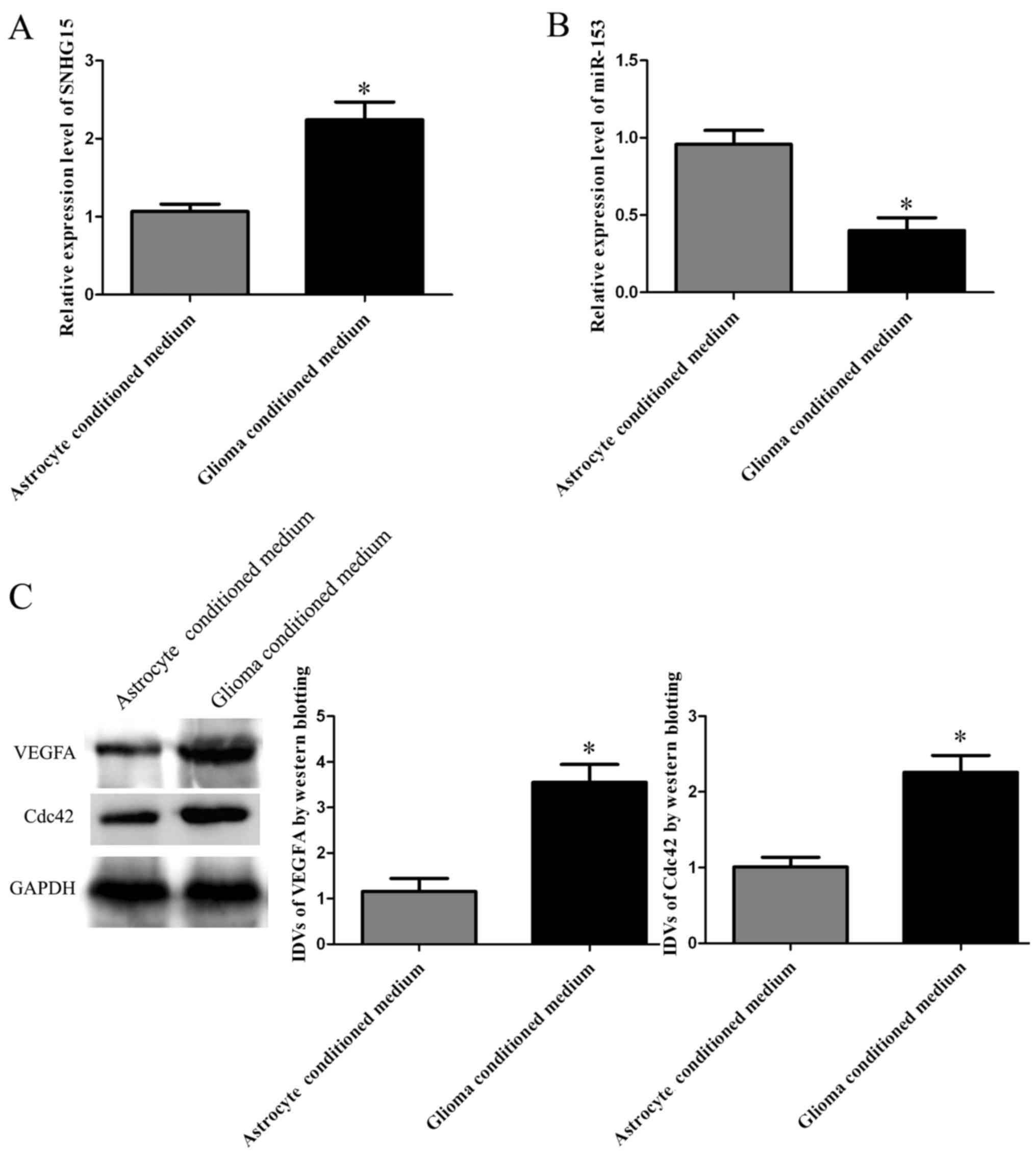
SNHG15 and miR-153 expression levels in hCMECs in
GCM and primary astrocyte cell conditioned medium were detected by
qRT-PCR. The SNHG15 expression level in glioma conditioned hCMECs
was increased compared with this level in the hCMECs in ACM
(P<0.05; Fig. 1A). miR-153
expression level in glioma conditioned hCMECs was decreased
compared with that in the hCMECs in ACM (P<0.05; Fig. 1B). Western blotting of VEGFA and
Cdc42 found increased expression of VEGFA and Cdc42 in the hCMECs
in GCM compared with these levels in the hCMECs in ACM (P<0.05;
Fig. 1C).
Knockdown of SNHG15 inhibits
proliferation, migration and tube formation of glioma vascular
endothelial cells
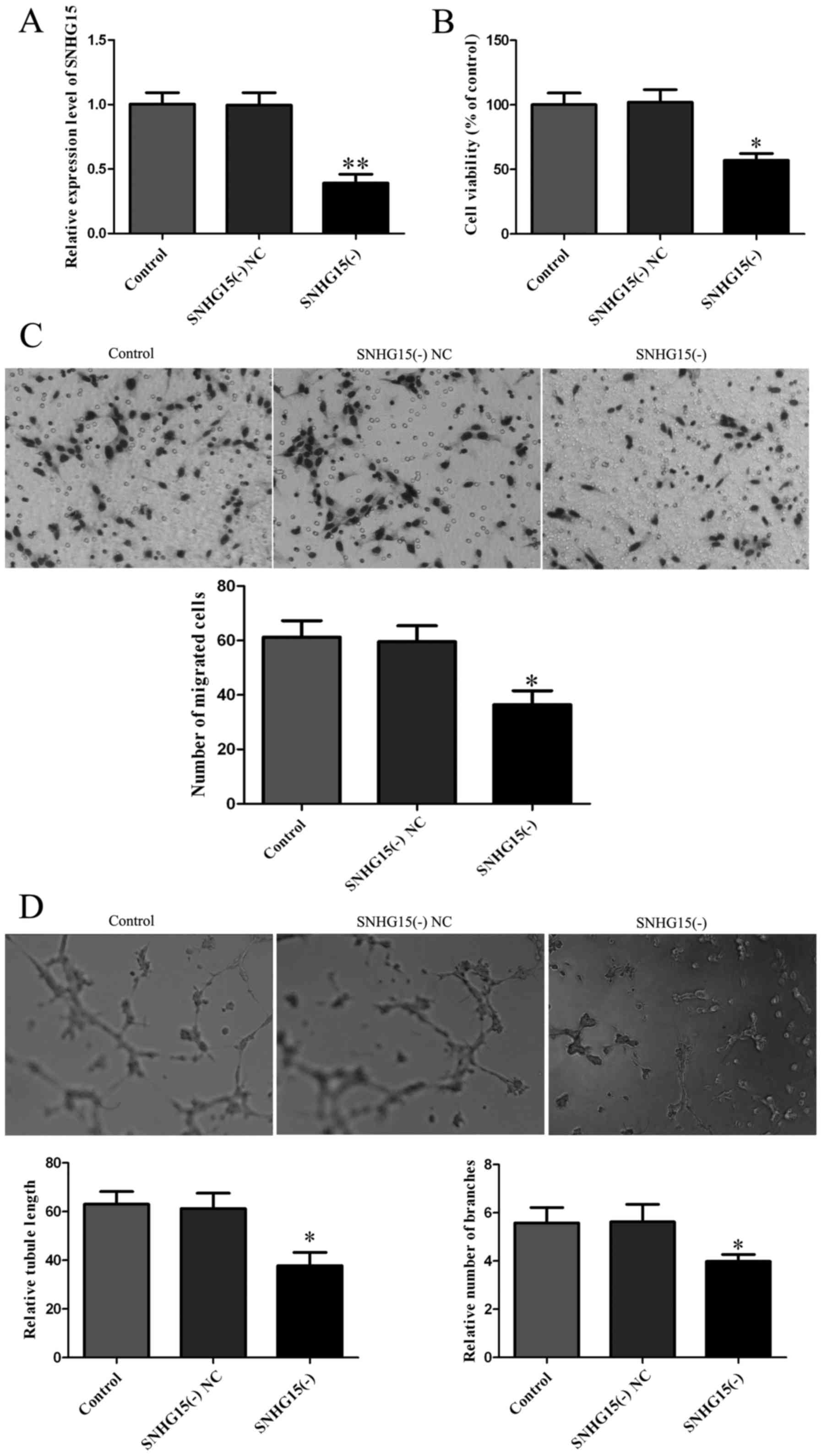
After investigation of the expression level of
SNHG15 in glioma vascular ECs, we further investigated the effect
of SNHG15 on proliferation, migration and tube formation of glioma
vascular ECs. The stable transfected EC line with knockdown of
SNHG15 was established. Expression of SNHG15 was 68.12% lower in
the SNHG15(−) group than that noted in the SNHG15(−) NC group
(P<0.01; Fig. 2A). Subsequently,
CCK-8 assay was performed to assess the effect of SNHG15 knockdown
on cell proliferation. The viability was lower in the SNHG15(−)
group than that noted in the SNHG15(−) NC group (P<0.05;
Fig. 2B). Cell migration assay
found that compared with the SNHG15(−) NC group, the SNHG15(−)
group showed a decrease in the migrated cell number (P<0.05;
Fig. 2C). Results of the tube
formation assay showed that relative tubule length and relative
number of branches were lower in the SNHG15(−) group in contrast to
these parameters in the SNHG15(−) NC group (P<0.05; Fig. 2D).
Knockdown of miR-153 promotes
proliferation, migration and tube formation of glioma vascular
endothelial cells
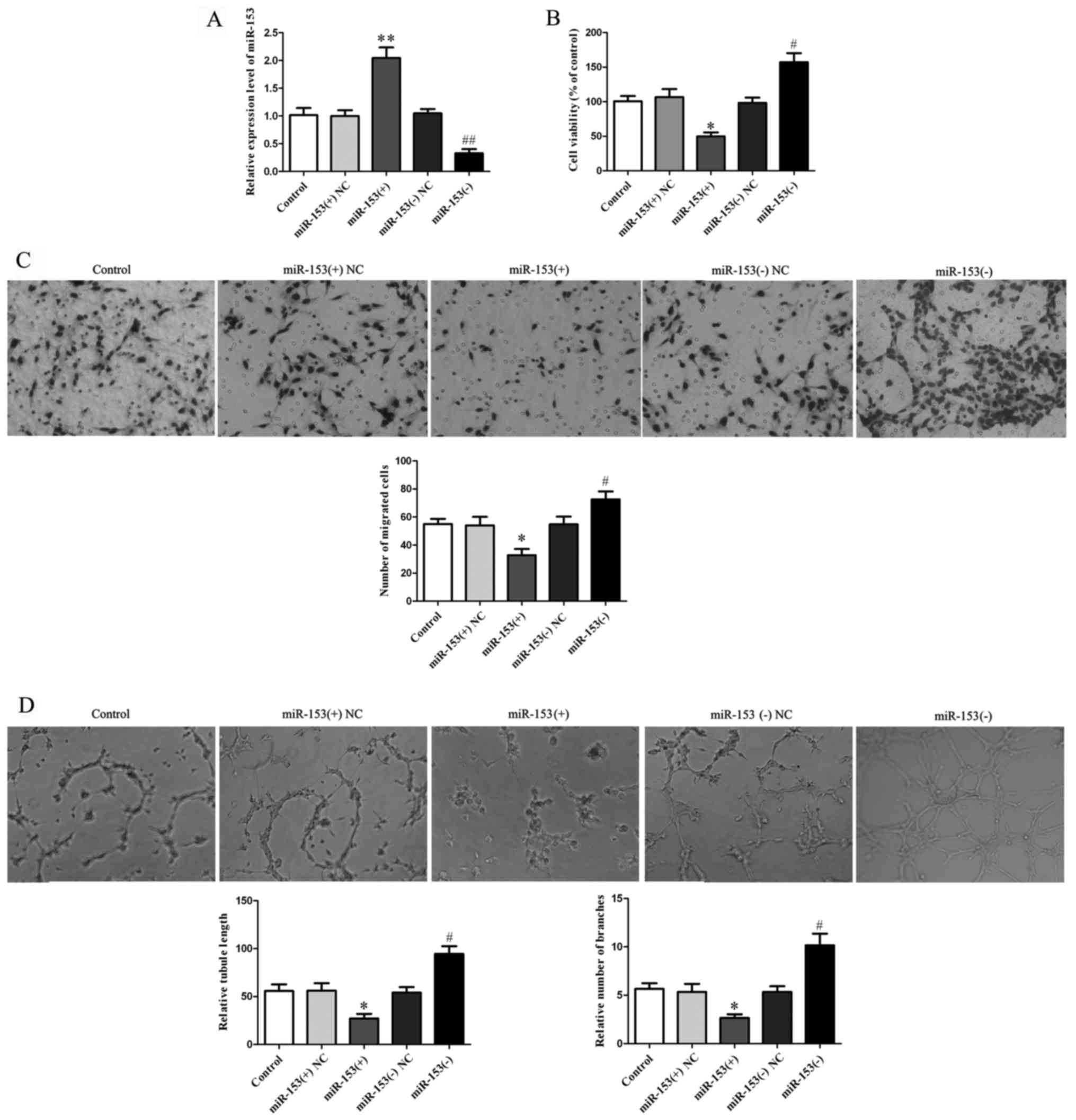
After investigation of miR-153 expression in glioma
vascular ECs, we further analyzed the effect on proliferation,
migration and tube formation of glioma vascular ECs. The expression
of miR-153 was 110.4% higher in the miR-153(+) group than that
noted in the miR-153(+) NC group and was 66.12% lower in the
miR-153(−) group than that in the miR-153(−) NC group (P<0.01;
Fig. 3A). CCK-8 assay found that
the viability was significantly lower in the miR-153(+) group
(P<0.01; Fig. 3B) and higher in
the miR-153(−) group (P<0.05; Fig.
3B), in contrast to the miR-153(+) NC and miR-153(−) NC group,
respectively. Cell migration assay results showed that the
miR-153(+) group had a decreased migratory cell number compared
with the miR-153(+) NC group while the miR-153(−) group had an
increased migratory cell number compared with that noted in the
miR-153(−) NC group (P<0.05; Fig.
3C). Results of the tube formation assay showed that the
miR-153(+) group had a decreased relative tubule length and
relative number of branches while the miR-153(−) group had enhanced
tube formation in vitro (P<0.05; Fig. 3D).
SNHG15 binds to and negatively
regulates miR-153
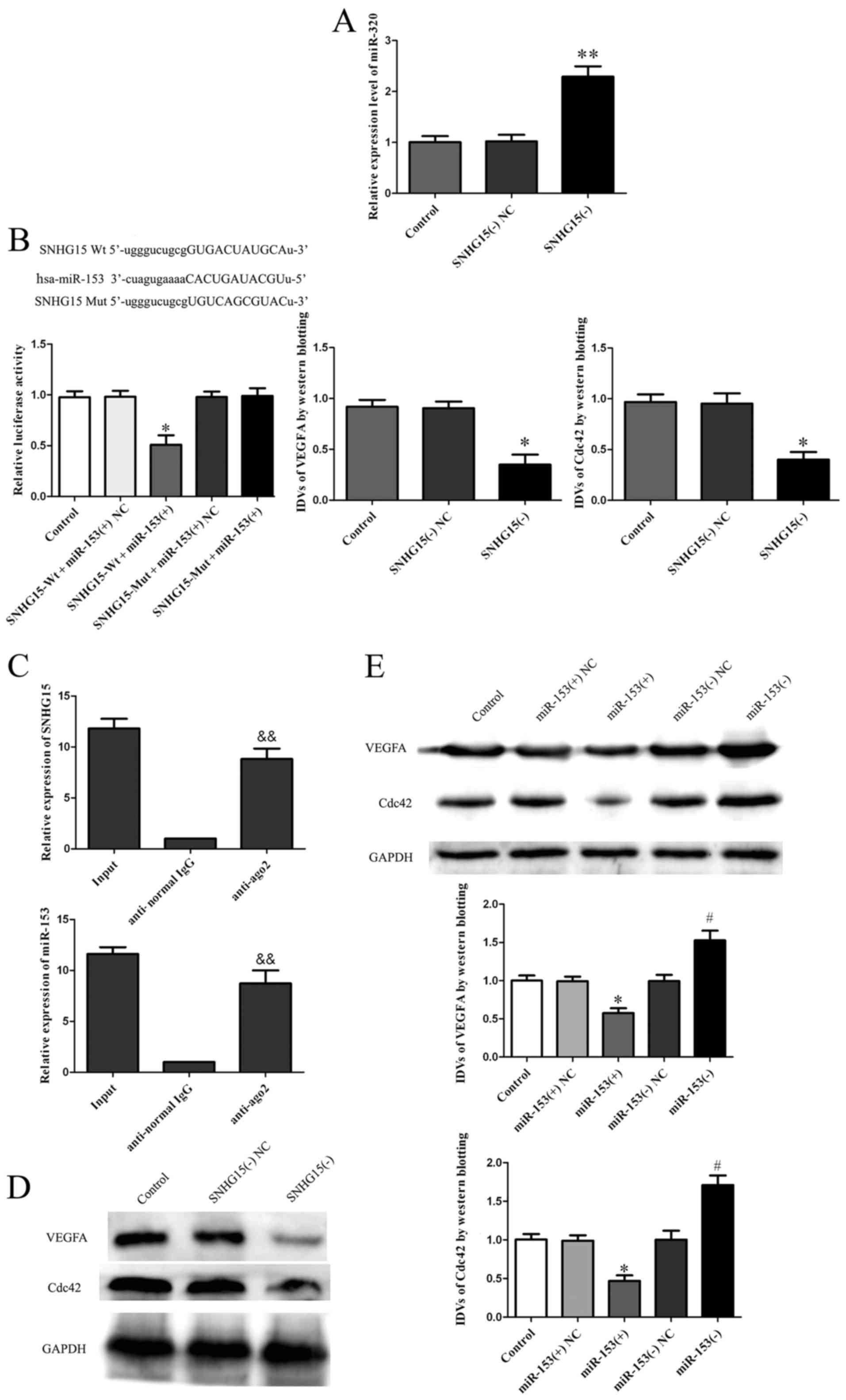
According to the bioinformatic database (RNAhybrid),
we predicted that SNHG15 may be associated with the miR-153 binding
sites. Furthermore, dual-luciferase gene reporter assay proved that
SNHG15 could bind to miR-153 at the predicted binding sites.
Quantitative RT-PCR results demonstrated that miR-153 expression
was upregulated in the SNHG15(−) group compared to the SNHG15(−) NC
group (P<0.01; Fig. 4A). Results
of dual-luciferase gene reporter assay showed that the luciferase
activity in the SNHG15-Wt + miR-153(+) group was lower than that in
the SNHG15-Wt + miR-153(+) NCgroup (P<0.05, Fig. 4B), indicating that SNHG15 bound to
miR-153 and negatively regulated its expression. To determine
whether SNHG15 is associated with the RNA-induced silencing complex
(RISC), we performed RNA-RIP assay. The RNA levels of SNHG15 and
miR-153 were higher in the anti-Ago2 group than levels in the
anti-normal IgG group (P<0.01; Fig.
4C).
Knockdown of SNHG15 downregulates the
expression of VEGFA and Cdc42 while knockdown of miR-153
upregulates the expression of VEGFA and Cdc42
Western blotting of VEGFA and Cdc42 indicated
decreased expression of VEGFA and Cdc42 in the SNHG15(−) group
compared with levels in the SNHG15(−) NC group (P<0.05; Fig. 4D). Furthermore, the expression of
VEGFA and Cdc42 was lower in the miR-153(+) group in contrast to
the miR-153(+) NC group and higher in the miR-153(−) group compared
with the miR-153(−) NC group (P<0.05; Fig. 4E).
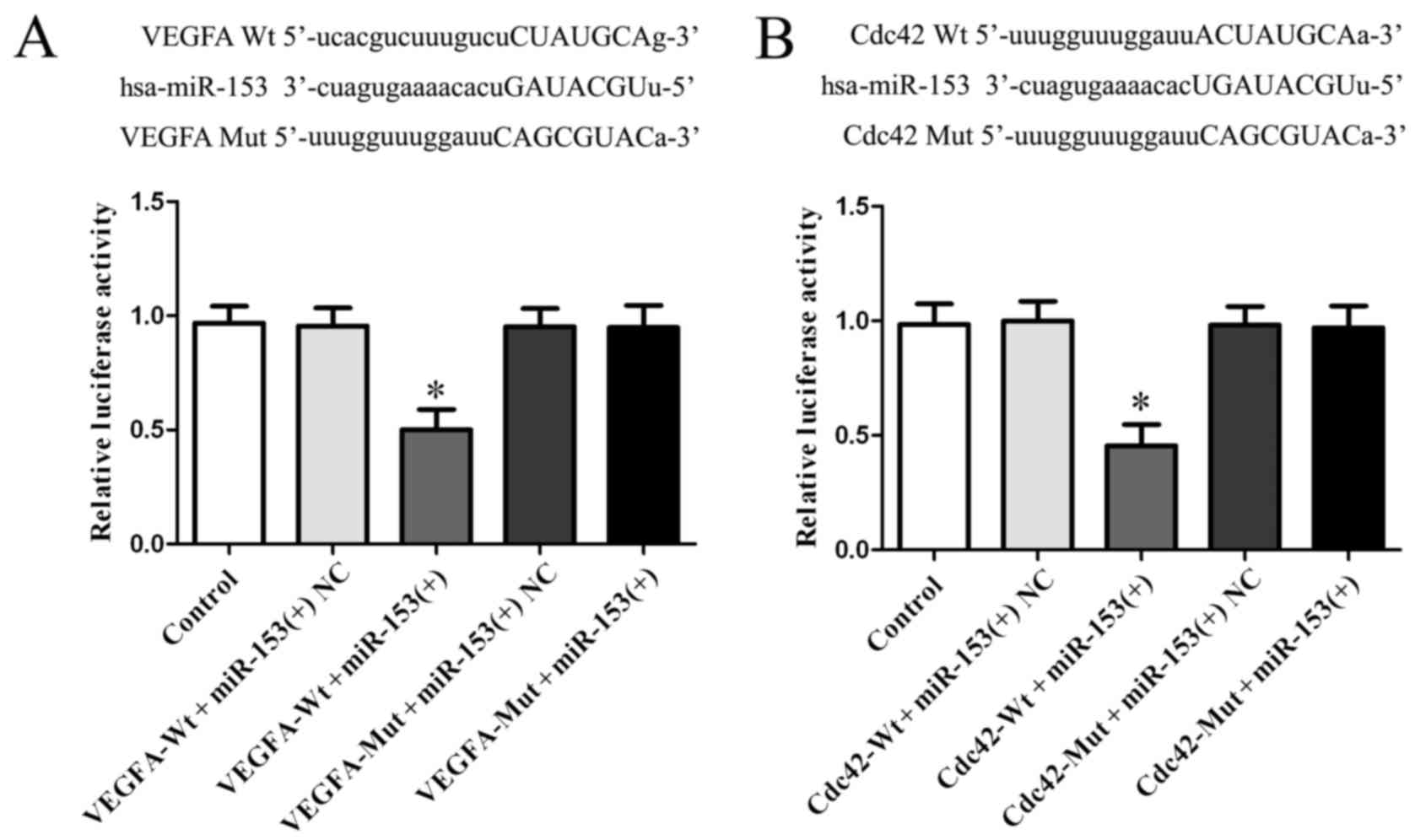
miR-153 binds to the 3′-UTR of VEGFA
and Cdc42
To investigate the targeted binding effect of
miR-153 to VEGFA and Cdc42, we searched the bioinformatic databases
(TargetScan, miRanda). Using TargetScan and miRanda we found
putative binding sites of miR-153 in the 3′-UTR of VEGFA and 3′-UTR
of Cdc42. To verify the combination between miR-153 and the 3′-UTR
of VEGFA and Cdc42, we performed dual-luciferase reporter assay.
Results showed that luciferase activity of VEGFA-Wt + miR-153(+)
and Cdc42-Wt + miR-153(+) group was lower than VEGFA-Wt +
miR-153(+) NC or Cdc42-Wt + miR-153(+) NC group (P<0.05;
Fig. 5A), while no significant
difference was found between VEGFA-Mut + miR-153(+) and VEGFA-Wt +
miR-153(+) NC group or between Cdc42-Mut + miR-153(+), and Cdc42-Wt
+ miR-153(+) NC group (P>0.05, Fig.
5A), which indicated that miR-153 bound with 3′-UTR of VEGFA
and Cdc42 by the putative binding sites.
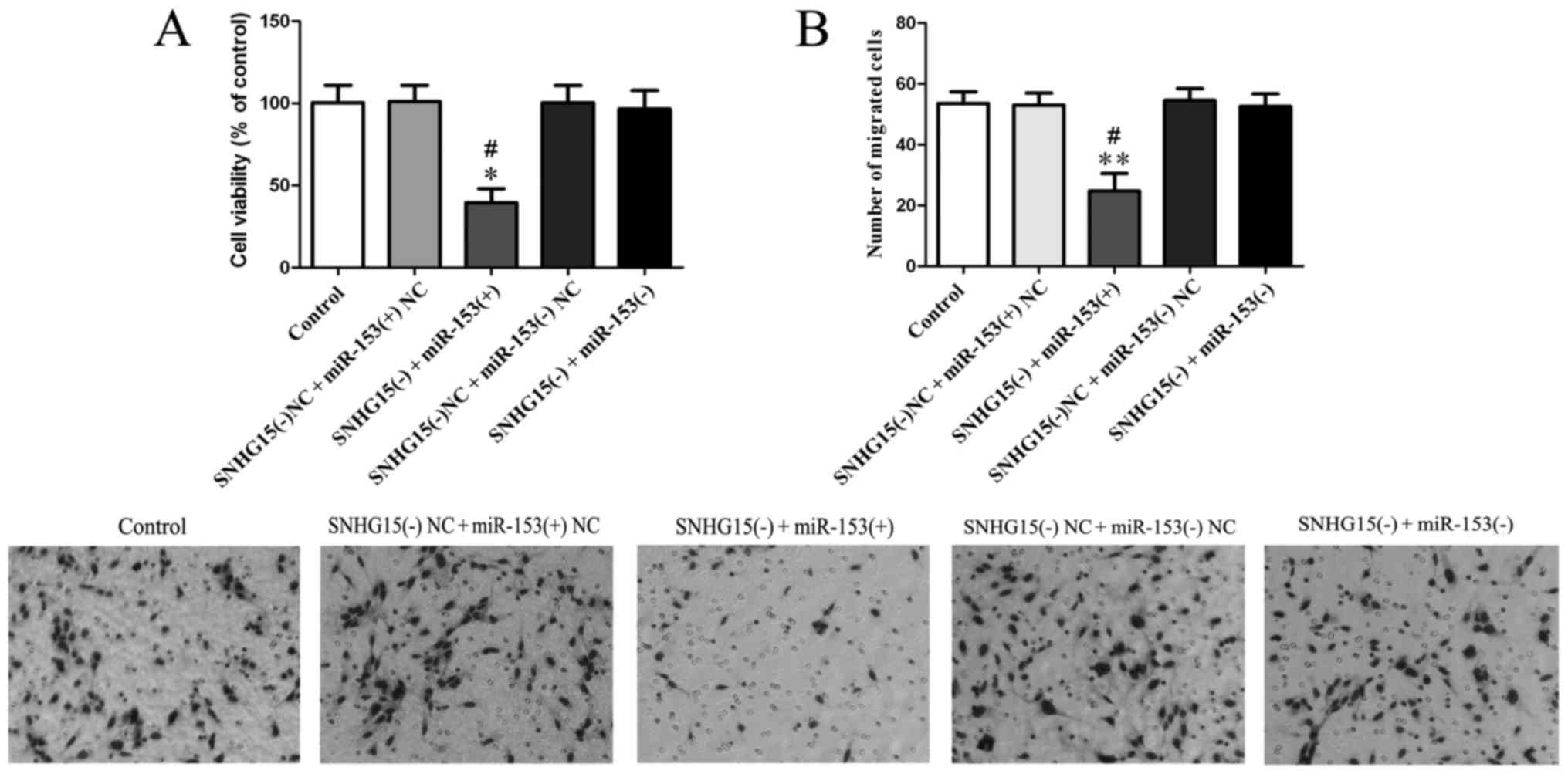
Knockdown of SNHG15 affects the
biological behaviors of glioma vascular endothelial cells and VEGFA
as well as Cdc42 expression by negative regulation of miR-153
To investigate whether SNHG15 affects the biological
behaviors of glioma vascular ECs by regulation of miR-153 and its
mechanism, we knocked down SNHG15 expression and also both knocked
down and overexpressed miR-153 simultaneously in the ECs. CCK-8
assay results showed that the viability of the SNHG15(−) +
miR-153(+) group was lower than both SNHG15(−) + miR-153(+) NC
(P<0.05; Fig. 6A) and SNHG15(−)
+ miR-153(−) groups (P<0.05; Fig.
6A).
The results of the cell migration assay showed that
the SNHG15(−) + miR-153(+) group had a decreased number of migrated
cells compared with the SNHG15(−) + miR-153(+) NC and SNHG15(−) +
miR-153(−) groups (P<0.05; Fig.
6B).
The tube formation assay found that the SNHG15(−) +
miR-153(+) group showed a decreased relative tubule length and a
decreased relative number of branches compared with the SNHG15(−) +
miR-153(+) NC and SNHG15(−) + miR-153(−) groups (P<0.05;
Fig. 6C).
Western blot analysis found that in the SNHG15(−) +
miR-153(+) group, VEGFA and Cdc42 expression was downregulated in
contrast to levels in the SNHG15(−) + miR-153(+) NC and SNHG15(−) +
miR-153(−) groups (P<0.05; Fig.
6D).
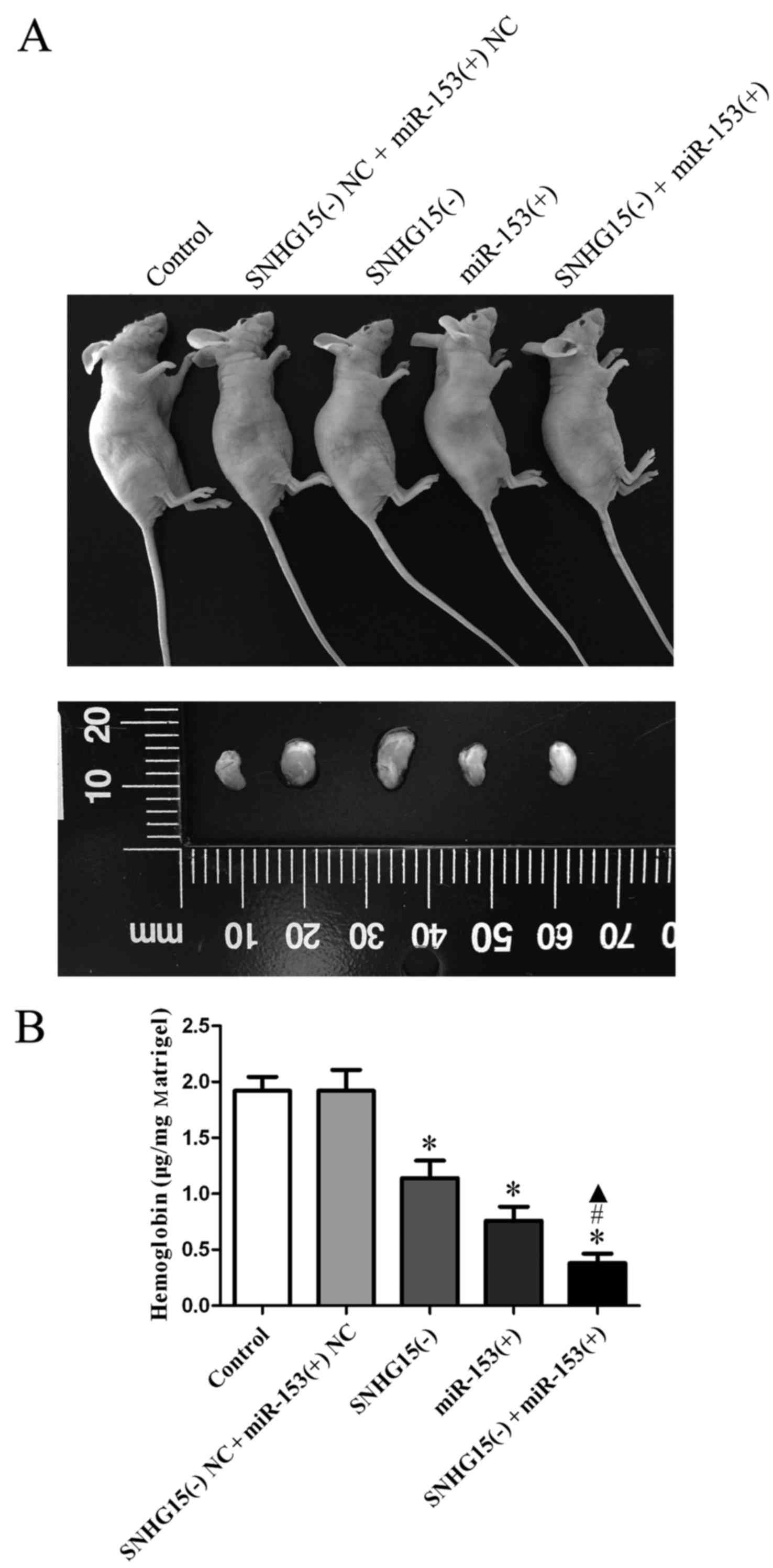
SNHG15 knockdown combined with miR-153
overexpression suppresses angiogenesis
Angiogenesis was also measured by a Matrigel plug
assay; the combinations of transfection were conducted prior to the
assessment of angiogenesis. The results revealed that the amount of
hemoglobin in the SNHG15(−), miR-153(+) and SNHG15(−) + miR-153(+)
groups was significantly decreased compared with that in the
SNHG15(−) NC + miR-153(+) NC group, and the amount of hemoglobin in
the SNHG15(−) + miR-153(+) group was found to be the least among
all the groups (P<0.05; Fig.
7B).
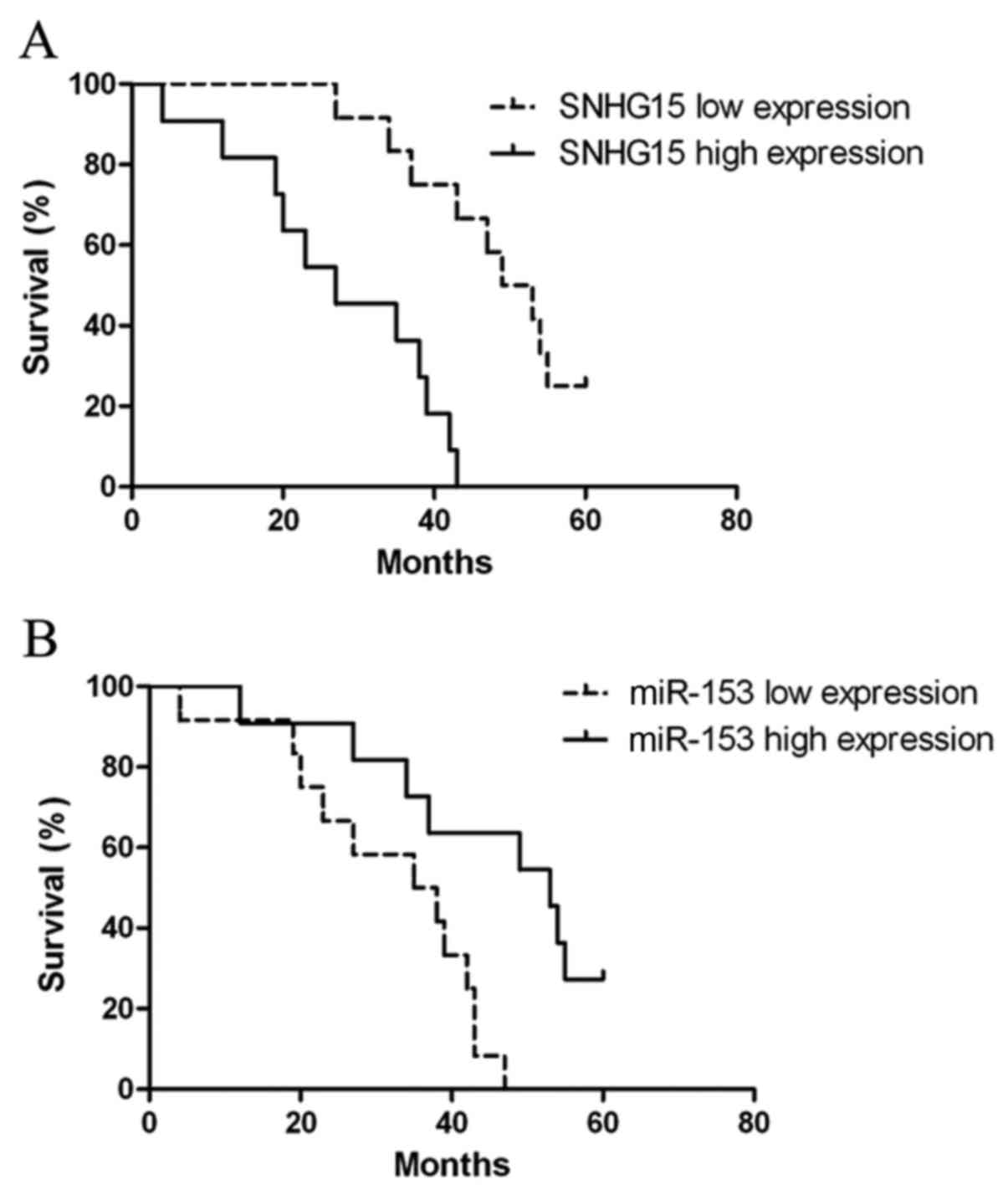
Association of SNHG15 and miR-153
expression with the survival of glioma patients
Kaplan-Meier analysis and log-rank test were
conducted to calculate the effects of SNHG15 and miR-153 on patient
survival. The overall survival rate was 17.39% (4 out of 23
patients). The median survival time of patients with low SNHG15
expression was 51 months, whereas the median survival time of those
with SNHG15 high expression was only 27 months. In log-rank test,
the results showed that patients with glioma of high SNHG15
expression tended to have poorer overall survival (log-rank test
P<0.001; Fig. 8A). The median
survival time of patients with low miR-153 expression was 36.5
months, whereas the median survival time of those with miR-153 high
expression was 53 months. In log-rank test, the results showed that
patients with glioma of high miR-153 expression tended to have
better overall survival (log-rank test P<0.01; Fig. 8B).
Discussion
In the present study, we found that SNHG15 was
highly expressed in glioma-induced endothelial cells while miR-153
was lowly expressed in glioma-induced endothelial cells. Knockdown
of SNHG15 or overexpression of miR-153 inhibited the proliferation,
migration and tube formation of glioma vascular endothelial cells
as well as decreased the expression of VEGFA and Cdc42. SNHG15 was
capable of binding to miR-153 and negatively regulating its
function. miR-153 targeted the 3′-UTR of VEGFA and Cdc42,
inhibiting their expression. SNHG15 affected proliferation,
migration and tube formation through negative regulation of
miR-153.
Tumor angiogenesis is a critical process during
cancer progression that modulates tumor growth and metastasis.
Solid tumors cannot grow beyond 2–3 mm in diameter without inducing
the formation of new blood vessels to support the energetic
requirements of tumor cells (7). As
for glioma, high microvessel density in glioma correlates with
tumor grade and is regarded as an independent marker of poor
patient prognosis (34), while
glioblastoma has been generally considered to be among the most
highlly vascularized tumor in humans (35). Vascular endothelial cells, as the
most important component of tumor vascularity, are the main target
through which glioma regulates angiogenesis by secretion of
multiple pro-angiogenic growth factors (9,10).
Further study showed that the glioma microenvironment also
triggered change in the expression of multiple genes in vascular
endothelial cells and whether this change takes part in glioma
angiogenesis remains controversal (36,37).
Recent studies have verified that lncRNAs not only
affect the biological behaviors of tumor cells directly, but also
modulate the function of tumor vascular endothelial cells. SNHG15,
firstly identified as one of the lncRNAs that respond to chemical
stressors [cisplatin, cycloheximide and mercury (II) oxide] in HeLa
Tet-off cells, was also found to be an oncogene (19,20,38).
Zhang et al showed that SNHG15 was highly expressed in
hepatocellular carcinoma and associated with poor overall survival
of patients (20). Furthermore,
Chen et al demonstrated that upregulated expression of
SNHG15 promoted cell proliferation and invasion in patients with
gastric cancer (19). To
investigate whether SNHG15 affects the function of glioma vascular
endothelial cells, we firstly detected the expression level of
SNHG15 in glioma conditioned endothelial cells and found that
SNHG15 was highly expressed in glioma vascular endothelial cells.
We further studied the effect of SNHG15 on glioma vascular
endothelial cell function. Knockdown of SNHG15 inhibited
proliferation, migration and tube formation in vitro. In
recent years, accumulating evidence has demonstrated that lncRNAs
are involved in the regulation of various biological behaviors of
endothelial cells. Overexpression of lncRNA-NONHSAT073641 was shown
to increase sprout number and length as well as branch number and
tubule length in HUVECs, thus promoting endothelial angiogenic
function (39). Michalik et
al verified that lncRNA-MALAT1 inhibited human umbilical vein
endothelial cell migration, but promoted endothelial cell
proliferation and angiogenesis (40). He et al confirmed that
lncRNA-p21 induced vascular endothelial cell apoptosis (41). Similarly, in the present study,
SNHG15 promoted glioma vascular endothelial cell proliferation,
migration and tube formation in vitro. These findings
highlight the important role of lncRNAs in the regulation of
vascular endothelial cell function.
miRNAs are involved in various biological processes
of tumor and vascular endothelial cells. miR-153 was first
discovered as 1 of the 7 brain-specific miRNAs based on expression
analysis of 119 miRNAs in adult organs from the mouse and human
using Northern blot analysis (42).
In glioma, the study of Silber et al showed that the
expression level of miR-153 was significantly decreased in GBM
multiforme relative to non-neoplastic brain tissue and a recent
study by Cui et al further demonstrated the inhibitory
function of miR-153 on the growth of glioma cells. To study the
effect of miR-153 on glioma vascular endothelial cells, we detected
the expression level of miR-153 in glioma conditioned vascular
endothelial cells and found that miR-153 expression was
downregulated in glioma vascular endothelial cells. Further study
by overexpression of miR-153 showed suppressed proliferation,
migration and tube formation of the endothelial cells, which
suggested that the abnormal decrease in miR-153 expression possibly
contributed to the growth of glioma vascularity. Recently,
increasing attention has been paid to the influence of miRNAs on
endothelial cell function. miR-542-3p was found to inhibit
angiogenesis of vascular endothelial cells both in vivo and
in vitro (43). Moreover,
miR-18a inhibited proliferation and migration of choroid
endothelial cells (44). The
present study showed that overexpression of miR-153 inhibited
proliferation, migration and in vitro tube formation of
glioma vascular endothelial cells, suggesting that miR-153 could
become a potential target for glioma anti-angiogenesis therapy.
VEGFA has long been recognized as essential for
vasculogenesis, the differentiation of endothelial cell progenitors
and their assembly into a primary capillary plexus, and for
angiogenesis, or the sprouting of new capillaries from pre-existing
vessels (45). VEGF is also the
most important mediator of neovascularization associated with
diverse human diseases including cancer (46). Cui et al demonstrated that
the conditioned medium of esophageal squamous cell carcinoma
induced a 12-fold upregulation of VEGFA in HUVECs, which underlines
the role of endogenous VEGFA in vascular endothelial cells under
the tumor microenvironment (47).
Cdc42 has been proven to play an important role in
neovascularization and vascular biology. Despite its roles in lumen
formation, regulating endothelial cell cytoskeleton and restoring
endothelial cell barrier function, Cdc42 was also shown to improve
VEGF-driven angiogenesis (48–51).
Hoang et al showed that Cdc42 improved neovessel
architecture and lumen formation in vivo and coordinated
microtubules and actin filaments and improved capillary
morphogenesis in vitro (48). The results of the present study
showed that both VEGFA and Cdc42 expression was upregulated in
glioma vascular endothelial cells while knockdown of SNHG15 and
overexpression of miR-153 downregulated the expression of VEGFA and
Cdc42, which lead to inhibition of proliferation, migration and
tube formation of endothelial cells.
miRNAs modulate target gene expression by binding to
the 3′-UTR of their target genes. Jin et al demonstrated
that miR-214 regulates endothelial progenitor cell function via
targeting the 3′-UTR of VEGF. Tang et al demonstrated that
overexpression of miR-185 inhibited glioma cell invasion by
targeting the 3′-UTR of Cdc42. Using several bioinformatic
databases (TargetScan, miRanda), we identified the 3′-UTR of VEGFA
and Cdc42 as binding sites for miR-153. We further confirmed the
combination between miR-153 and 3′-UTR of VEGFA as well as Cdc42 by
dual-luciferase reporter assay.
lncRNAs interact with miRNAs, as lncRNAs function as
endogenous sponge or bait, thus negatively regulating expression
and activity of miRNAs (52). In
the present study, we identified the effects of SNHG15 and miR-153
on endothelial cell function. The present study also showed that
knockdown of SNHG15 in vascular endothelial cells led to a
significant increase in miR-153 expression, indicating that miR-153
was involved in the effect of SNHG15 on endothelial cells. We found
a binding site between SNHG15 and miR-153 according to a
bioinformatic database (RNAhybrid). We further confirmed that
SNHG15 was capable of binding to miR-153 by dual-luciferase
reporter assay, which indicated that SNHG15 may affect the function
of endothelial cells by regulating miR-153. The results of the RIP
assay also support the involvement of RNA-induced silencing complex
in this reciprocal repression process, which indicated that SNHG15
may affect the function of endothelial cells by regulating miR-153.
To investigate whether SNHG15 affected the biological behaviors of
endothelial cells through regulation of miR-153, we transfected the
cells with knockdown of SNHG15 and both overexpression or knockdown
of miR-153 simultaneously. The results revealed that knockdown of
SNHG15 along with overexpression of miR-153 suppressed, to the
greatest extent, endothelial cell proliferation, migration, tube
formation and decreased the expression of VEGFA and Cdc42, while
knockdown of miR-153 reversed the suppressing effect of the
knockdown of SNHG15 on endothelial cell function and expression of
VEGFA and Cdc42. In addition, the in vivo studies also
supported the above findings. The results above suggest that SNHG15
affected glioma vascular endothelial cells through negative
regulation of miR-153. Interaction with miRNAs is a common way of
lncRNA function in vascular endothelial cells. Yan et al
showed that lncRNA-MIAT affected proliferation, migration and
angiogenesis of vascular endothelial cells by binding with
miR-150-5p (41).
To elucidate the effect of SNHG15 and miR-153 on
glioma prognosis, we investigated the association of SNHG15 and
miR-153 expression with the survival of glioma patients. Our
results demonstrated that SNHG15 is a potential prognostic factor
for glioma patients, and has a negative impact on overall survival
of primary glioma patients while miR-153 showed a contrasting
result.
In summary, this is the first study to demonstrate
that SNHG15 is highly expressed in glioma-induced endothelial cells
while miR-153 is lowly expressed in glioma-induced endothelial
cells. SNHG15 negatively regulates the expression of miR-153 and
miR-153 negatively regulates the expression of VEGFA and Cdc42,
which in turn affects glioma vascular endothelial cell
proliferation, migration and tube formation in vitro.
Therefore, we suggest that SNHG15 and miR-153 are novel targets for
glioma anti-angiogenesis therapy.
Acknowledgements
The present study was supported by grants from the
Natural Science Foundation of China (nos. 81573010 and 81672511),
the Liaoning Science and Technology Plan Project (no. 2015225007),
the Shenyang Science and Technology Plan Projects (nos.
F15-199-1-30 and F15-199-1-57) and the Outstanding Scientific Fund
of Shengjing Hospital (no. 201304).
References
|
1
|
Weller M, Van den Bent M, Hopkins K, Tonn
JC, Stupp R, Falini A, Cohen-Jonathan-Moyal E, Frappaz D,
Henriksson R, Balana C, et al European Association for
Neuro-Oncology (EANO) Task Force on Malignant Glioma, : EANO
guideline for the diagnosis and treatment of anaplastic gliomas and
glioblastoma. Lancet Oncol. 15:e395–e403. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dimberg A: The glioblastoma vasculature as
a target for cancer therapy. Biochem Soc Trans. 42:1647–1652. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Polivka J Jr, Polivka J, Rohan V, Topolcan
O and Ferda J: New molecularly targeted therapies for glioblastoma
multiforme. Anticancer Res. 32:2935–2946. 2012.PubMed/NCBI
|
|
4
|
Patel M, Vogelbaum MA, Barnett GH, Jalali
R and Ahluwalia MS: Molecular targeted therapy in recurrent
glioblastoma: Current challenges and future directions. Expert Opin
Investig Drugs. 21:1247–1266. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang W, Barth RF, Huo T, Nakkula RJ,
Weldon M, Gupta N, Agius L and Grecula JC: Radiation therapy
combined with intracerebral administration of carboplatin for the
treatment of brain tumors. Radiat Oncol. 9:252014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Brem S, Cotran R and Folkman J: Tumor
angiogenesis: A quantitative method for histologic grading. J Natl
Cancer Inst. 48:347–356. 1972.PubMed/NCBI
|
|
7
|
Lopes-Bastos BM, Jiang WG and Cai J:
Tumour-endothelial cell communications: Important and indispensable
mediators of tumour angiogenesis. Anticancer Res. 36:1119–1126.
2016.PubMed/NCBI
|
|
8
|
Zecchin A, Borgers G and Carmeliet P:
Endothelial cells and cancer cells: Metabolic partners in crime?
Curr Opin Hematol. 22:234–242. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dunn IF, Heese O and Black PM: Growth
factors in glioma angiogenesis: FGFs, PDGF, EGF, and TGFs. J
Neurooncol. 50:121–137. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Egidy G, Eberl LP, Valdenaire O, Irmler M,
Majdi R, Diserens AC, Fontana A, Janzer RC, Pinet F and
Juillerat-Jeanneret L: The endothelin system in human glioblastoma.
Lab Invest. 80:1681–1689. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cai H, Xue Y, Li Z, Hu Y, Wang Z, Liu W,
Li Z and Liu Y: Roundabout4 suppresses glioma-induced endothelial
cell proliferation, migration and tube formation in vitro by
inhibiting VEGR2-mediated PI3K/AKT and FAK signaling pathways. Cell
Physiol Biochem. 35:1689–1705. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Smits M, Wurdinger T, van het Hof B,
Drexhage JA, Geerts D, Wesseling P, Noske DP, Vandertop WP, de
Vries HE and Reijerkerk A: Myc-associated zinc finger protein (MAZ)
is regulated by miR-125b and mediates VEGF-induced angiogenesis in
glioblastoma. FASEB J. 26:2639–2647. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Angrand PO, Vennin C, Le Bourhis X and
Adriaenssens E: The role of long non-coding RNAs in genome
formatting and expression. Front Genet. 6:1652015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Svoboda M, Slyskova J, Schneiderova M,
Makovicky P, Bielik L, Levy M, Lipska L, Hemmelova B, Kala Z,
Protivankova M, et al: HOTAIR long non-coding RNA is a negative
prognostic factor not only in primary tumors, but also in the blood
of colorectal cancer patients. Carcinogenesis. 35:1510–1515. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ding C, Cheng S, Yang Z, Lv Z, Xiao H, Du
C, Peng C, Xie H, Zhou L, Wu J, et al: Long non-coding RNA HOTAIR
promotes cell migration and invasion via down-regulation of RNA
binding motif protein 38 in hepatocellular carcinoma cells. Int J
Mol Sci. 15:4060–4076. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim K, Jutooru I, Chadalapaka G, Johnson
G, Frank J, Burghardt R, Kim S and Safe S: HOTAIR is a negative
prognostic factor and exhibits pro-oncogenic activity in pancreatic
cancer. Oncogene. 32:1616–1625. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Okugawa Y, Toiyama Y, Hur K, Toden S,
Saigusa S, Tanaka K, Inoue Y, Mohri Y, Kusunoki M, Boland CR, et
al: Metastasis-associated long non-coding RNA drives gastric cancer
development and promotes peritoneal metastasis. Carcinogenesis.
35:2731–2739. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ke J, Yao YL, Zheng J, Wang P, Liu YH, Ma
J, Li Z, Liu XB, Li ZQ, Wang ZH, et al: Knockdown of long
non-coding RNA HOTAIR inhibits malignant biological behaviors of
human glioma cells via modulation of miR-326. Oncotarget.
6:21934–21949. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen SX, Yin JF, Lin BC, Su HF, Zheng Z,
Xie CY and Fei ZH: Upregulated expression of long noncoding RNA
SNHG15 promotes cell proliferation and invasion through regulates
MMP2/MMP9 in patients with GC. Tumour Biol. 37:6801–6812. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang JH, Wei HW and Yang HG: Long
noncoding RNA SNHG15, a potential prognostic biomarker for
hepatocellular carcinoma. Eur Rev Med Pharmacol Sci. 20:1720–1724.
2016.PubMed/NCBI
|
|
21
|
Zheng Y, Zhu C, Ma L, Shao P, Qin C, Li P,
Cao Q, Ju X, Cheng G, Zhu Q, et al: miRNA-154-5p inhibits
proliferation, migration and invasion by targeting E2F5 in prostate
cancer cell lines. Urol Int. 98:102–110. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xue Z, Yilan D, Ping J and Fei M:
Bioinformatic analysis of cancer-related microRNAs and their target
genes. Yi Chuan. 37:855–864. 2015.PubMed/NCBI
|
|
23
|
Roy S and Sen CK: miRNA in wound
inflammation and angiogenesis. Microcirculation. 19:224–232. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu X, Li L, Li Y and Liu Z: MiR-153
promotes breast cancer cell apoptosis by targeting HECTD3. Am J
Cancer Res. 6:1563–1571. 2016.PubMed/NCBI
|
|
25
|
Sun Y, Jia X, Hou L and Liu X: Screening
of differently expressed miRNA and mRNA in prostate cancer by
integrated analysis of transcription data. Urology.
94:313.e1–313.e6. 2016. View Article : Google Scholar
|
|
26
|
Chen WJ, Zhang EN, Zhong ZK, Jiang MZ,
Yang XF, Zhou DM and Wang XW: MicroRNA-153 expression and prognosis
in non-small cell lung cancer. Int J Clin Exp Pathol. 8:8671–8675.
2015.PubMed/NCBI
|
|
27
|
Ghasemi A, Fallah S and Ansari M: miR-153
as a tumor suppressor in glioblastoma multiforme is downregulated
by DNA methylation. Clin Lab. 62:573–580. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang L, Hu S, Tan J, Zhang X, Yuan W, Wang
Q, Xu L, Liu J, Liu Z, Jia Y, et al: Pregnancy-specific
glycoprotein 9 (PSG9), a driver for colorectal cancer, enhances
angiogenesis via activation of SMAD4. Oncotarget. 7:61562–61574.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu J, Yuan P, Mao Q, Lu P, Xie T, Yang H
and Wang C: miR-613 inhibits proliferation and invasion of breast
cancer cell via VEGFA. Biochem Biophys Res Commun. 478:274–278.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cai H, Liu X, Zheng J, Xue Y, Ma J, Li Z,
Xi Z, Li Z, Bao M and Liu Y: Long non-coding RNA taurine
upregulated 1 enhances tumor-induced angiogenesis through
inhibiting microRNA-299 in human glioblastoma. Oncogene.
36:318–331. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Garzon-Muvdi T, Schiapparelli P, ap Rhys
C, Guerrero-Cazares H, Smith C, Kim DH, Kone L, Farber H, Lee DY,
An SS, et al: Regulation of brain tumor dispersal by NKCC1 through
a novel role in focal adhesion regulation. PLoS Biol.
10:e10013202012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Senger DL, Tudan C, Guiot MC, Mazzoni IE,
Molenkamp G, LeBlanc R, Antel J, Olivier A, Snipes GJ and Kaplan
DR: Suppression of Rac activity induces apoptosis of human glioma
cells but not normal human astrocytes. Cancer Res. 62:2131–2140.
2002.PubMed/NCBI
|
|
33
|
Jia P, Cai H, Liu X, Chen J, Ma J, Wang P,
Liu Y, Zheng J and Xue Y: Long non-coding RNA H19 regulates glioma
angiogenesis and the biological behavior of glioma-associated
endothelial cells by inhibiting microRNA-29a. Cancer Lett.
381:359–369. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Leon SP, Folkerth RD and Black PM:
Microvessel density is a prognostic indicator for patients with
astroglial brain tumors. Cancer. 77:362–372. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Plate KH and Risau W: Angiogenesis in
malignant gliomas. Glia. 15:339–347. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pen A, Moreno MJ, Martin J and
Stanimirovic DB: Molecular markers of extracellular matrix
remodeling in glioblastoma vessels: Microarray study of
laser-captured glioblastoma vessels. Glia. 55:559–572. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mustafa DA, Dekker LJ, Stingl C, Kremer A,
Stoop M, Smitt PA Sillevis, Kros JM, Luider TM, et al: A proteome
comparison between physiological angiogenesis and angiogenesis in
glioblastoma. Mol Cell Proteomics. 11:M111.0084662012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tani H and Torimura M: Identification of
short-lived long non-coding RNAs as surrogate indicators for
chemical stress response. Biochem Biophys Res Commun. 439:547–551.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Josipovic I, Fork C, Preussner J, Prior
KK, Iloska D, Vasconez AE, Labocha S, Angioni C, Thomas D,
Ferreirós N, et al: PAFAH1B1 and the lncRNA NONHSAT073641 maintain
an angiogenic phenotype in human endothelial cells. Acta Physiol.
218:13–27. 2016.
|
|
40
|
Michalik KM, You X, Manavski Y,
Doddaballapur A, Zörnig M, Braun T, John D, Ponomareva Y, Chen W,
Uchida S, et al: Long noncoding RNA MALAT1 regulates endothelial
cell function and vessel growth. Circ Res. 114:1389–1397. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yan B, Yao J, Liu JY, Li XM, Wang XQ, Li
YJ, Tao ZF, Song YC, Chen Q and Jiang Q: lncRNA-MIAT regulates
microvascular dysfunction by functioning as a competing endogenous
RNA. Circ Res. 116:1143–1156. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sempere LF, Freemantle S, Pitha-Rowe I,
Moss E, Dmitrovsky E and Ambros V: Expression profiling of
mammalian microRNAs uncovers a subset of brain-expressed microRNAs
with possible roles in murine and human neuronal differentiation.
Genome Biol. 5:R132004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
He T, Qi F, Jia L, Wang S, Wang C, Song N,
Fu Y, Li L and Luo Y: Tumor cell-secreted angiogenin induces
angiogenic activity of endothelial cells by suppressing miR-542-3p.
Cancer Lett. 368:115–125. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Miao YS, Zhao YY, Zhao LN, Wang P, Liu YH,
Ma J and Xue YX: MiR-18a increased the permeability of BTB via
RUNX1 mediated down-regulation of ZO-1, occludin and claudin-5.
Cell Signal. 27:156–167. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Carmeliet P: Mechanisms of angiogenesis
and arteriogenesis. Nat Med. 6:389–395. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ferrara N: VEGF: An update on biological
and therapeutic aspects. Curr Opin Biotechnol. 11:617–624. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jin G, Yang Y, Liu H, Liu K, Zhao J, Chen
X, Zhang X, Zhang Y, Lu J and Dong Z: Genome-wide analysis of the
effect of esophageal squamous cell carcinoma on human umbilical
vein endothelial cells. Oncol Rep. 36:155–164. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hoang MV, Nagy JA and Senger DR:
Cdc42-mediated inhibition of GSK-3β improves angio-architecture and
lumen formation during VEGF-driven pathological angiogenesis.
Microvasc Res. 81:34–43. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tzima E, Kiosses WB, del Pozo MA and
Schwartz MA: Localized cdc42 activation, detected using a novel
assay, mediates microtubule organizing center positioning in
endothelial cells in response to fluid shear stress. J Biol Chem.
278:31020–31023. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kouklis P, Konstantoulaki M, Vogel S,
Broman M and Malik AB: Cdc42 regulates the restoration of
endothelial barrier function. Circ Res. 94:159–166. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Koh W, Mahan RD and Davis GE: Cdc42- and
Rac1-mediated endothelial lumen formation requires Pak2, Pak4 and
Par3, and PKC-dependent signaling. J Cell Sci. 121:989–1001. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Cesana M, Cacchiarelli D, Legnini I,
Santini T, Sthandier O, Chinappi M, Tramontano A and Bozzoni I: A
long noncoding RNA controls muscle differentiation by functioning
as a competing endogenous RNA. Cell. 147:358–369. 2011. View Article : Google Scholar : PubMed/NCBI
|