Introduction
Renal cell carcinoma is the most common malignant
tumor of the kidney, accounting for almost 3% of all human
malignancies (1). Insensitivity to
radiotherapy and chemotherapy are a great obstacle for the
treatment of metastatic renal cell carcinoma.
Epithelial-mesenchymal transition (EMT), a hallmark of metastasis,
is a complicated process by which cells lose epithelial
characteristics and acquire a mesenchymal phenotype (2). In the process of EMT, cancer cells
escape from the primary site and invade to distant tissues through
blood and lymphatic vessels. In addition, an EMT phenotype is often
accompanied by the downregulation of epithelial markers E-cadherin
and zonuela occludens 1 (ZO-1), and upregulation of mesenchymal
markers such as N-cadherin, Twsit1 and Twist2, resulting in
enhanced motility (3,4). Furthermore, the expression of
E-cadherin or vimentin has been reported to be associated with
tumor progression and overall survival (OS) in a variety of
cancers, including lung cancer and nasopharyngeal carcinoma
(5,6). Additionally, Slug and Snail are found
to downregulate E-cadherin by binding to the promoter of E-cadherin
(7). In view of this, there is an
urgent need to identify potential therapeutic targets against renal
cell carcinoma.
High-mobility group AT-hook 2 (HMGA2), a member of
the high mobility group family, is a small non-histone
nuclear-binding protein, which contains three AT-hook structural
domains and an acid C-terminal tail. It has been reported that
HMGA2 participates in proliferation and differentiation during
embryonic development (8). In
addition, HMGA2 was found to be overexpressed in various types of
tumors, including ovarian cancer, hepatocellular carcinoma and
gliomas (9–11). Moreover, a high level of HMGA2 was
correlated with poor prognosis in breast cancer and lung cancer
patients (12,13). Although numerous studies have
validated that HMGA2 may play a crucial role in tumor progression,
only few studies have shown a correlation between HMGA2 and renal
cell carcinoma. In addition, the underlying role of HMGA2 in renal
cell carcinoma has not yet been elucidated.
In the present study, we focused on the role of
HMGA2 in cell migration, invasion and EMT of renal cell carcinoma
and revealed the possible mechanism by which HMGA2 regulates EMT
in vitro.
Materials and methods
Reagents
Rabbit monoclonal antibodies against HMGA2 (8179),
E-cadherin (E-Ca; 3195), N-cadherin (N-Ca; 13116), Twist1 (46702),
TGF-β (3709) phosphorylated-Smad2 (p-Smad2; 3108), Smad2 (5339),
Gli1 (3538), phosphorylated-β-catenin (p-β-catenin; 4176) and
β-actin (4970) were purchased from Cell Signaling Technology, Inc.
(Beverly, MA, USA). Rabbit polyclonal antibody against Twist2
(ab66031) was purchased from Abcam (Cambridge, UK). The appropriate
peroxidase-conjugated goat anti-rabbit IgG and goat anti-mouse IgG
secondary antibodies were purchased from Zhongshan Biotech
(Beijing, China). ACHN stably transfected with sh-HMGA2, 786-O
stably transfected with HMGA2 (OE-HMGA2), and their respective
corresponding empty vector control sublines (ACHN-scramble,
786-O-vector) were previously constructed.
Cell lines and culture
Human renal tubular epithelial HK2 cell line, and
five renal cell carcinoma cell lines 786-O, 769-P, OSRC-2, ACHN and
Caki-1 were purchased from the American Type Culture Collection
(ATCC, Manassas, VA, USA). All cells were maintained in RPMI-1640
medium and supplemented with 10% fetal bovine serum (Gibco, Grand
Island, NY, USA), 100 µg/ml streptomycin and 100 U/ml penicillin
(Invitrogen, Carlsbad, CA, USA) at 37°C in a humidified atmosphere
with 5% CO2.
Western blotting
For western blot analysis, cells were washed with
phosphate-buffered saline (PBS) after the specific treatment, and
proteins were extracted using a lysis buffer [10 mmol/l Tris-HCl
(pH 7.4), 150 mmol/l NaCl, 0.1% sodium dodecyl sulfate (SDS), 1
mmol/l ethylenediaminetetraacetic acid, 1 mmol/l ethylene glycol
tetraacetic acid, 0.3 mmol/l phenylmethylsulfonyl fluoride, 0.2
mmol/l sodium orthovanadate, 1% NP-40, 10 mg ml/1 leupeptin, and 10
mg/ml aprotinin]. Equal amounts of protein lysates (~40–60 µg) were
loaded onto a 10 or 15% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) gel and electrotransferred onto
polyvinylidene difluoride membranes (Millipore, Bedford, MA, USA)
for 2 h. The membranes were then blocked with 5% non-fat milk, and
incubated with primary antibodies (1:1,000) against E-cadherin
(E-Ca), N-cadherin (N-Ca), Twist1, Twist2, TGF-β, p-Smad2, Smad2,
Gli1, p-β-catenin and β-actin at 4°C overnight. The membranes were
subsequently washed with TBST (Tris-buffered saline with Tween)
buffer and incubated with corresponding secondary antibodies
(1:1,000) at room temperature (25°C) for 1 h. Ultimately, the bands
were visualized by an enhanced chemiluminescence kit (Bio-Rad,
Hercules, CA, USA) and analyzed using Image Lab 4.0 (Bio-Rad)
imaging software.
Wound healing assay
Cells were seeded onto 6-well plates. After the cell
density reached to complete cell culture medium, scratch wounds
were made across the monolayer with the tip of a 200-µl pipette.
Subsequently, the wounded cultures were visualized in serum-free
medium at 0 and 24 h, and images (×100) were then captured by
inverted microscopy (Olympus IX50; Olympus, Tokyo, Japan) to detect
the migratory ability. The experiments were performed in
triplicate.
Cell migration assay
The cell migration ability of renal cell carcinoma
cells following different treatments was assessed using Transwell
migration assay. Cells (ACHN, 2×104; 786-O,
1.5×104) in 200 µl serum-free medium were seeded into
the upper chambers, and 800 µl of 10% fetal calf serum-containing
medium was used as a chemoattractant in the lower chamber. After
incubation for 24 h, the cells which migrated onto the bottom of
the filter were then fixed with 4% paraformaldehyde, and stained
with 0.1% crystal violet (Beyotime, Shanghai, China). Cells were
then washed and photographed in five independent visual fields
using an inverted microscope (Olympus IX50; Olympus) at ×100
magnification.
Matrigel invasion assay
Cell invasive ability was detected by Matrigel
invasion assay using a Millicell chamber (Millipore, Billerica, MA,
USA). The Transwell chambers were precoated with 50 µl mixture
(Matrigel:serum-free medium 1:5) and the stable clone cell lines
(ACHN, 5×104; 786-O, 4×104) in 200 µl
serum-free medium were seeded onto the upper chamber for 24 h
according to the instructions of the Transwell migration assay. The
difference between two groups was analyzed by Sudent's t-test
(two-sided).
Quantitative real-time PCR assay
Total cellular RNA was extracted using TRIzol
reagent (Invitrogen). The RNA sample was reversely transcribed by
PrimerScript RT reagent kit (Takara, Dalian, China). Then the
relative levels of target gene messenger RNA (mRNA) were evaluated
by quantitative real-time PCR assay (qPCR) using FAST SYBR-Green
Master Mix with the gene-specific primers: E-cadherin (119 bp)
forward, 5′-CGAGAGCTACACGTTCACGG-3′; and reverse,
5′-GGGTGTCGAGGGAAAAATAGG-3′; N-cadherin (94 bp) forward,
5′-TCAGGCGTCTGTAGAGGCTT-3′ and reverse,
5′-ATGCACATCCTTCGATAAGACTG-3′; Twist1 (156 bp) forward,
5′-GTCCGCAGTCTTACGAGGAG-3′ and reverse,
5′-GCTTGAGGGTCTGAATCTTGCT-3′; Twist2 (200 bp) forward,
5′-GGAGTCCGCAGTCTTACGAG-3′; and reverse,
5′-TCTGGAGGACCTGGTAGAGG-3′; TGF-β (201 bp) forward,
5′-GGCCAGATCCTGTCCAAGC-3′ and reverse, 5′-GTGGGTTTCCACCATTAGCAC-3′;
Smad2 (182 bp) forward, 5′-CGTCCATCTTGCCATTCACG-3′; and reverse,
5′-CTCAAGCTCATCTAATCGTCCTG-3′; β-actin (250 bp) forward,
5′-CATGTACGTTGCTATCCAGGC-3′; and reverse
5′-CTCCTTAATGTCACGCACGAT-3′. The n-fold change in mRNA expression
was analyzed according to the method of 2−∆∆Ct.
Plasmid transfection
TGF-β cDNA was cloned into the pcDNA3.1 vector. The
plasmid was transfected into renal cell carcinoma ACHN cells with
sh-HMGA2 or 768-O cells with HMGA2 overexpression by Bo Kou using
X-treme GENE HP DNA transfection reagent (Roche, Germany) for 48 h
according to the manufacturer's instructions, and subsequently
prepared for the mechanistic research.
Statistical analysis
All experimental data were analyzed by GraphPad
Prism (San Diego, CA, USA) software and presented as means ± SD.
Differences between two groups were analyzed by Student's t-test
(two-sided). The correlations between HGMA2 and Twist1 or Twist2
were analyzed by Spearman's rank test. In addition, overall
survival (OS) was assessed by log-rank test, while a Kaplan-Meier
curve was generated. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of HMGA2 in human renal
epithelial cells and renal cell carcinoma cell lines
Firstly, we detected the expression of HMGA2 in
several renal cell carcinoma cell lines using western blotting. The
results demonstrated that HMGA2 was elevated in all renal cell
carcinoma cell lines, compared with that noted in the human renal
tubular epithelial HK2 cell line (Fig.
1A). Among these cell lines, HMGA2 was the most highly
expressed in the renal cell carcinoma ACHN cell line, whereas it
was lowly expressed in the 786-O cell line. In view of this, we
constructed stable clone cell lines with knockdown or
overexpression of HMGA2. In addition, we explored the HMGA2
expression level in ACHN cells with HMGA2 knockdown by qPCR and
western blotting, and validated a significant decrease in HMGA2
expression in the sh-HMGA2 ACHN cells compared with that in the
scramble or negative control (Fig. 1B
and C). Conversely, the expression of HMGA2 was significantly
increased in the 786-O cells overexpressing HMGA2 (OE-HMGA2) in
comparison with that noted in cells transfected with the vector or
negative control, as evidenced by qPCR and western blotting
(Fig. 1D and E).
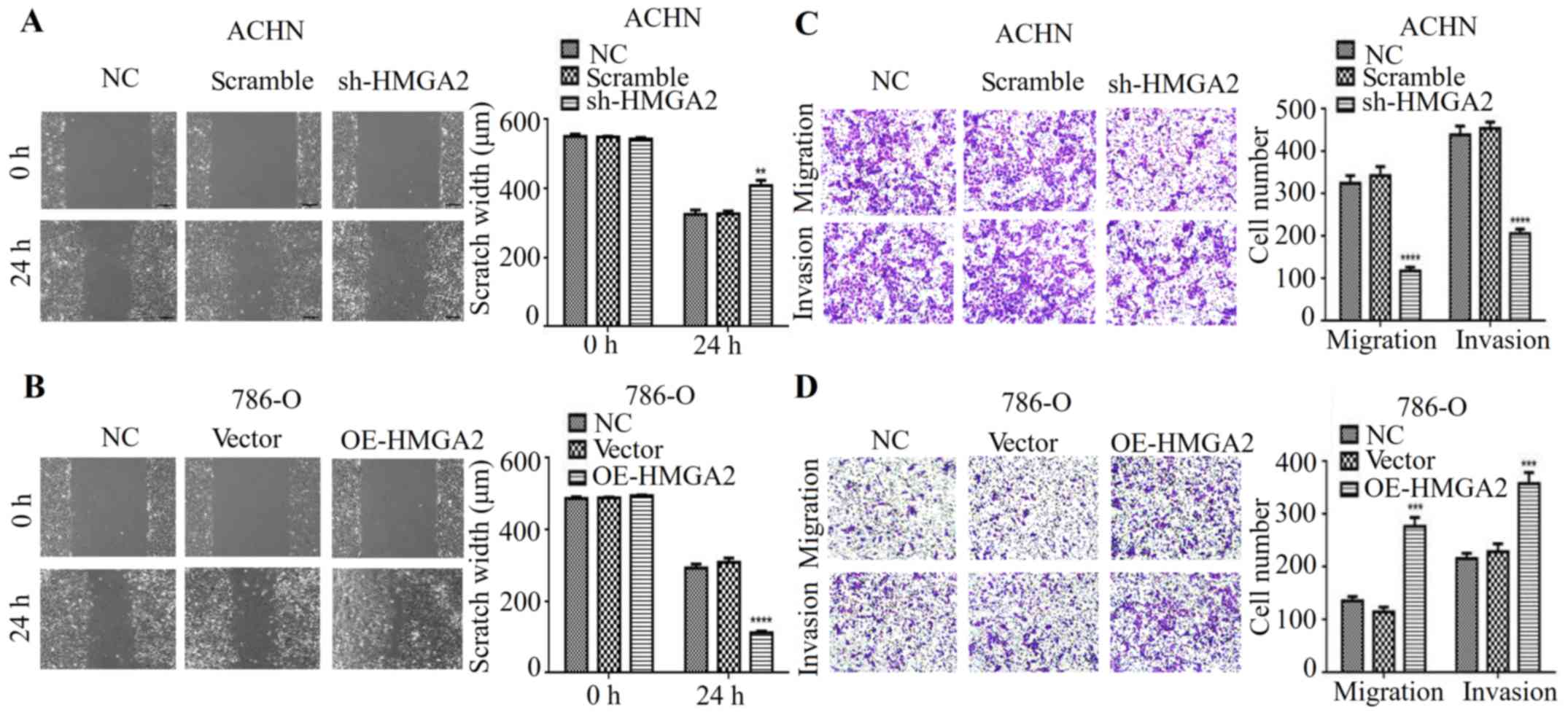
HMGA2 regulates the metastatic
phenotype of renal cell carcinoma
Studies showed that HMGA2 is associated with cancer
metastasis in a variety of cancers (14,15).
Thus, we hypothesized that HMGA2 is implicated in the migration and
invasion of human renal cell carcinoma. To verify this hypothesis,
wound healing and Transwell migration assays with statistical
quantification were used. As expected, our results indicated that
HMGA2 knockdown had a much wider scratch width and restrained cell
migration when compared to the negative control or scramble in ACHN
cell line (Fig. 2A and C). In
contrast, HMGA2 overexpression promoted cell migration of renal
cell carcinoma (Fig. 2B and D).
Next, we determined whether HMGA2 affects invasive ability in
vitro. As shown in Fig. 2C, the
silencing of HMGA2 expression decreased the number of invaded
cells, while overexpressing HMGA2 increased the invasive property
of 786-O cell line compared with negative control (Fig. 2D). These data indicated that HMGA2
knockdown inhibited the metastatic phenotype of human renal cell
carcinoma cells in vitro.
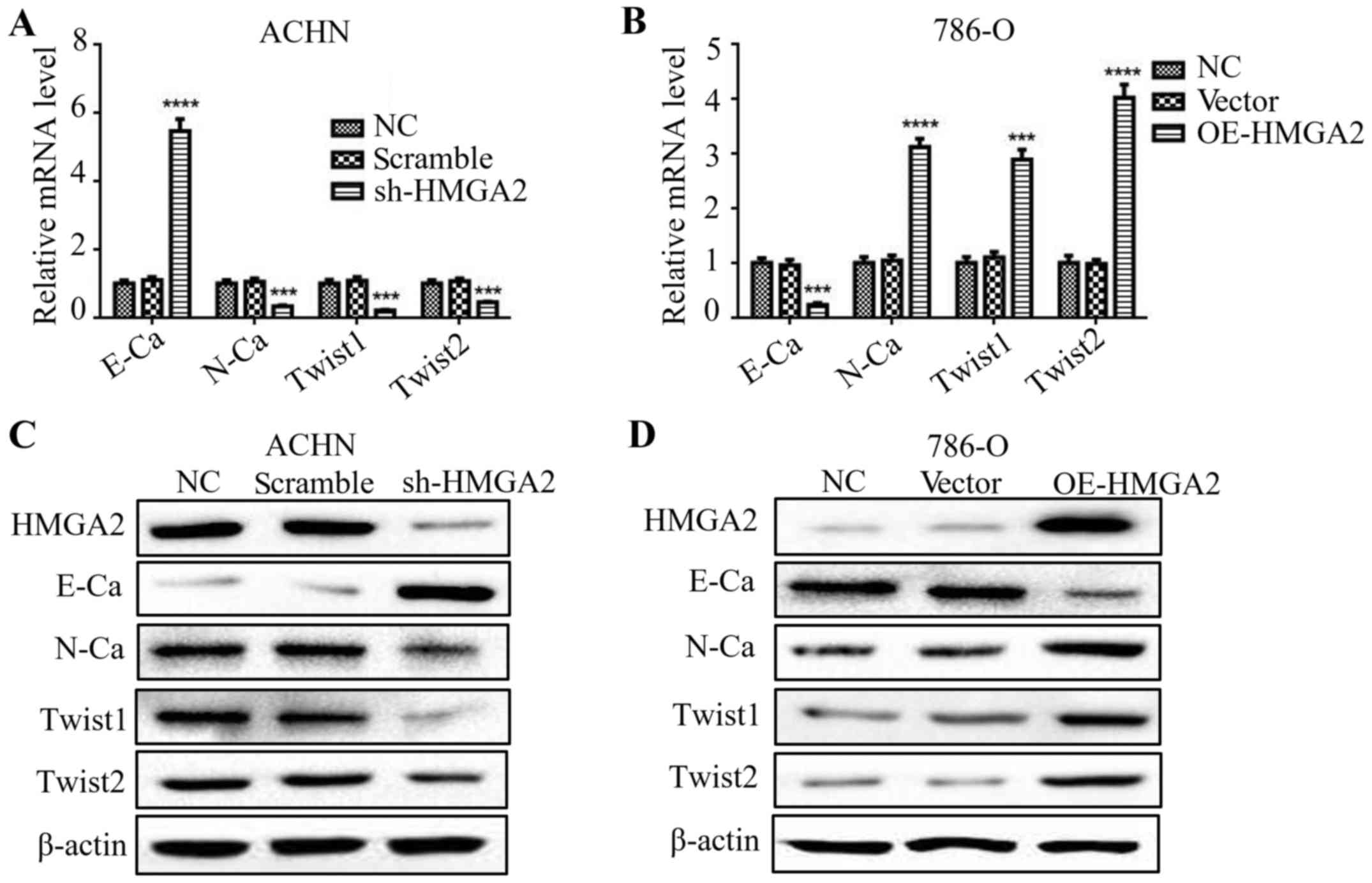
HMGA2 regulates the EMT of renal cell
carcinoma
Several studies have stated that HMGA2 is involved
in the process of EMT (16,17). To validate the function of HMGA2 in
the EMT process of renal cell carcinoma cells, we evaluated the
levels of both epithelial and mesenchymal markers by qPCR and
western blotting. The deficiency in HMGA2 presented an elevated
level of E-cadherin, while a reduction in N-cadherin, Twist1 and
Twist2 (Fig. 3A and C). Conversely,
786-O cells with HMGA2 overexpression exhibited a significant
decline in E-cadherin level, while an increase in N-cadherin,
Twist1 and Twist2. (Fig. 3B and D).
These results indicated that knockdown of HMGA2 reversed the EMT of
human renal cell carcinoma.
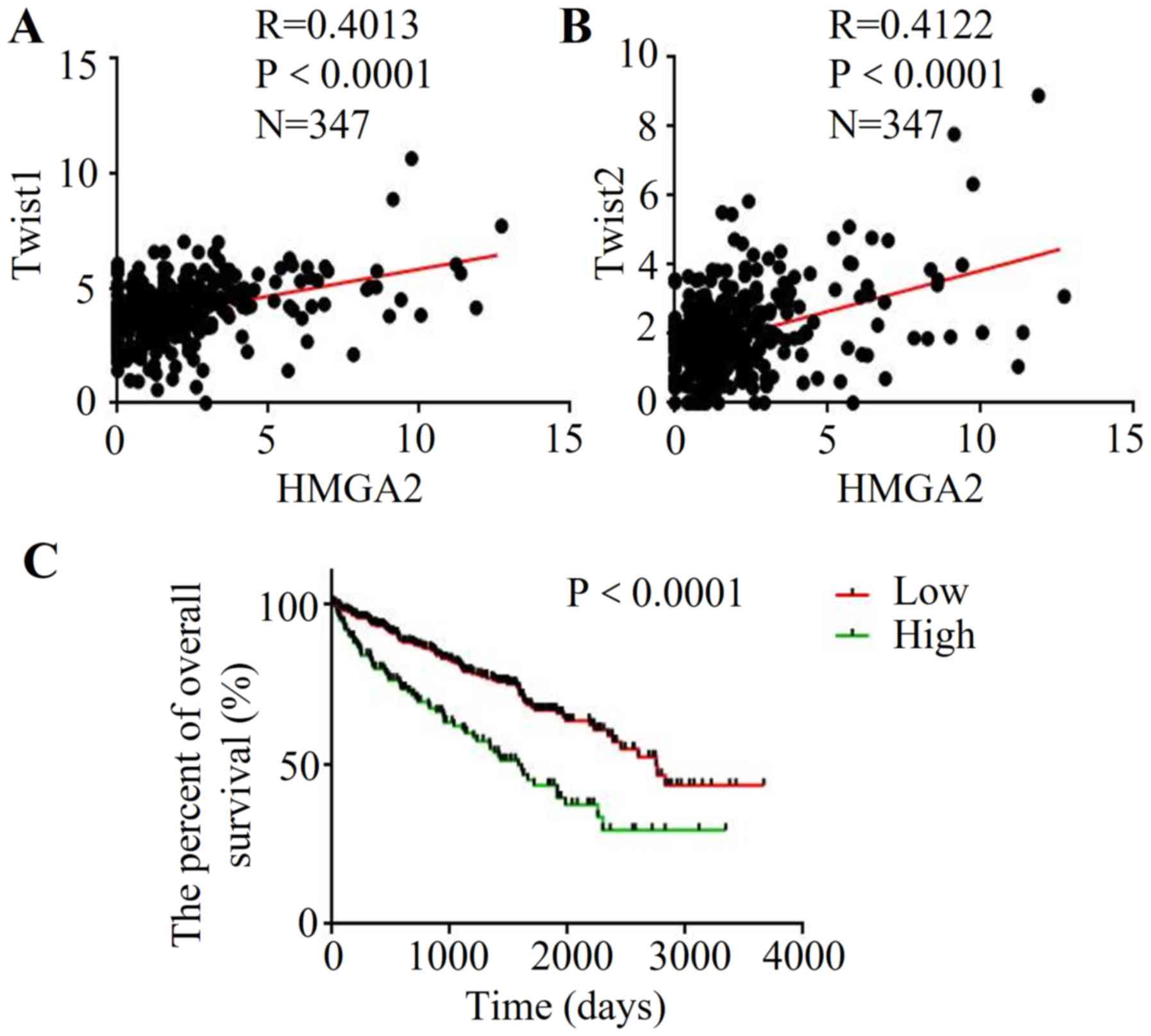
Correlation between HMGA2 and EMT
markers based on Cancer Browser database
To further explore the association between HMGA2 and
EMT in renal cell carcinoma, the clinical data and gene expression
from database Cancer Browser
(TCGA_KIRC_exp_HiSeqV2-2015-02-24) were extracted. The results
demonstrated that high expression of HMGA2 was correlated with
increased Twist1 expression (R=0.0.4013, P<0.0001) (Fig. 4A). Meanwhile, HMGA2 expression was
positively correlated with the Twist2 level in renal cell carcinoma
(R=0.0.4122, P<0.0001) (Fig.
4B). Then, we used Kaplan-Meier analysis to evaluate the
prognostic value of HMGA2 in renal cell carcinoma. Importantly, the
low HMGA2 patient group had a better OS than that of the
high-expression group (Fig. 4C),
indicating that high HMGA2 may be a poor prognostic predictor of
renal cell carcinoma.
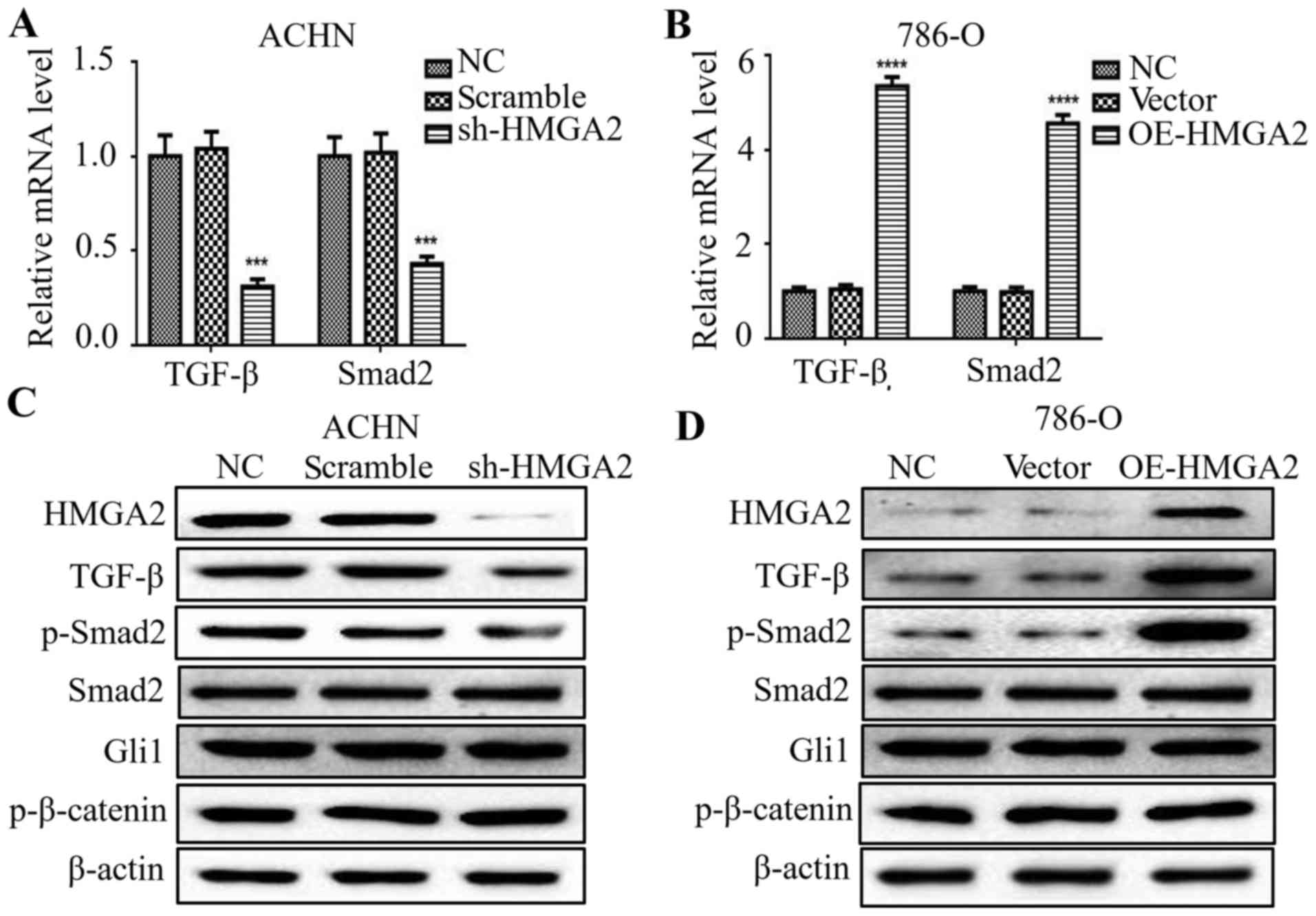
Silencing of HMGA2 decreases TGF-β and
Smad2 expression in renal cell carcinoma
Previous studies have reported that the EMT process
is governed by various regulatory networks, such as TGF-β, Wnt and
Hedgehog signaling. To clarify the correlation among HMGA2 and
several signals in renal cell carcinoma, we firstly evaluated that
the change in TGF-β-, Wnt- and Hedgehog-related markers in
HMGA2-knockdown ACHN cells or HMGA2-overexpressing 786-O cells. The
mRNA levels of TGF-β and Smad2 were downregulated by silencing of
HMGA2 (Fig. 5A), and upregulated in
the 786-O cells with HMGA2 overexpression (Fig. 5B). To further examine the protein
levels of the above markers, we found a marked decrease of TGF-β
and phosphorylated-Smad2 in the HMGA2-depleted ACHN cells, and a
marked increase in TGF-β and phosphorylated-Smad2 in the
HMGA2-overexpressing 786-O cells (Fig.
5C and D). Meanwhile, the protein level of total Smad2, Gli1
and p-β-catenin had no significant change following HMGA2 knockdown
or overexpression. These findings suggest that HMGA2 regulated
TGF-β/Smad2 signaling in renal cell carcinoma.
HMGA2 participates in the EMT process
of renal cell carcinoma by regulating the TGF-β/Smad2 signaling
pathway
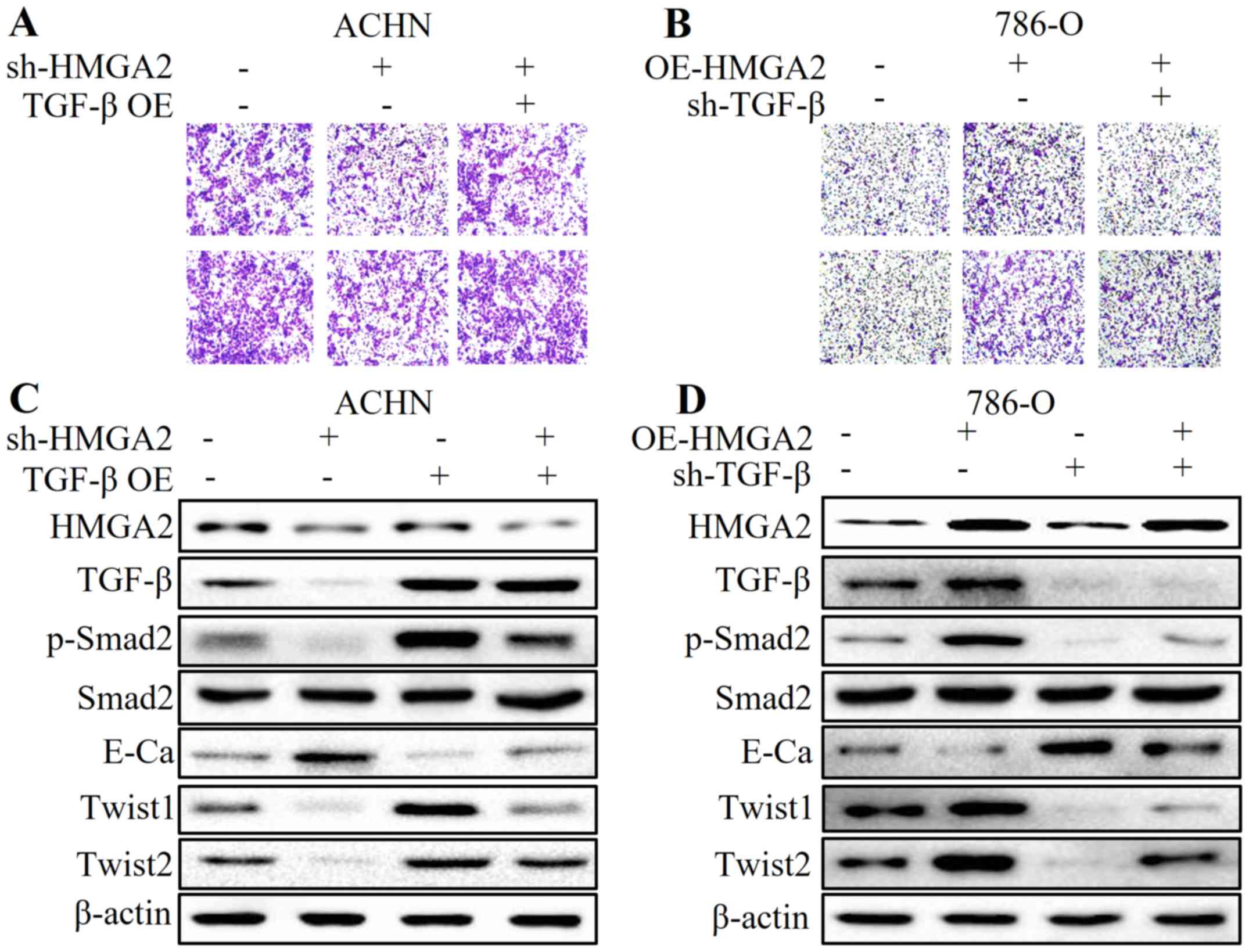
To further elucidate the role of TGF-β in
HMGA2-regulated EMT in renal cell carcinoma, we applied plasmid
transfection to overexpress TGF-β in HMGA2-deficient ACHN cells,
and to knock down TGF-β in HMGA2-overexpressing 786-O cells. We
found that overexpression of TGF-β partially reversed the
anti-metastatic effect and MET by HMGA2 loss (Fig. 6A and C). Conversely, the
pro-metastatic phenotype and high expression of TGF-β- and
EMT-related markers induced by HMGA2 overexpression were abolished
by TGF-β deficiency (Fig. 6B and
D). These results strongly suggest that the TGF-β/Smad2
signaling pathway is involved in the HMGA2-mediated EMT of renal
cell carcinoma.
Discussion
Accumulating evidence indicates that HMGA2 is highly
expressed in solid tumors and is regulated by complicated
regulatory systems (18–20). Studies have shown that in renal cell
carcinoma, HMGA2 expression is significantly higher than that in
benign and normal renal tissues (21). Moreover, there is a positive
correlation between HMGA2 and clinical staging and lymph node
metastasis. Also, another study reported that the expression of
HMGA2 was significantly associated with tumor size and Fuhrman
grade in patients with clear cell renal cell carcinoma (ccRCC)
(22). In the present study, we
firstly confirmed that HMGA2 was highly expressed in five renal
cell carcinoma cell lines compared with that in the normal renal
tubular epithelial HK2 cell line. Recent studies have shown that
HMGA2 may play an essential role in cancer proliferation, migration
and metastasis (23–25). It was reported that deficiency of
HMGA2 reduced the metastatic potential of breast cancer cells
(26). Additionally, HMGA2 seems to
have the potential of enhancing self-renewal capacity in cancer
stem cells (27). The present study
demonstrated that HMGA2 knockdown inhibited cell migration and
invasion, while overexpression of HMGA2 facilitated the metastatic
phenotype in renal cell carcinoma cells in vitro, as
evidenced by wound healing and Transwell assays. These results
emphasize the critical role of HGMA2 in renal cell carcinoma
metastasis.
EMT is a complex process during which tumor cells
gain more aggressive and metastatic ability (28). Previous studies have reported that
silencing of HMGA2 may attenuate migration and invasion, and
reverse epithelial-mesenchymal transition (EMT) in nasopharyngeal
cancer cells (29). Furthermore,
HMGA2 was found to promote EMT by inducing Slug expression in colon
cancer (30). Our findings revealed
that the E-cadherin level was upregulated, while N-cadherin, Twist1
and Twist2 were downregulated in HMGA2-depleted ACHN cells. In
contrast, overexpression of HMGA2 in 786-O cells enhanced EMT.
Subsequently, the analysis of database Cancer Browser
further validated the positive correlation between HGMA2 and Twist1
or Twist2 in renal cell carcinoma. In addition, the Kaplan-Meier
analysis indicated that low HMGA2 expression was closely associated
with an increased overall survival (OS) in renal cell carcinoma,
which was in accordance with a previous study (22).
Multiple signaling pathways are found to participate
in the EMT process. Sonic hedgehog-Gli1 signals are considered to
be critical for EMT in ovarian cancer (31). In addition, the Wnt/β-catenin
signaling pathway is reported to promote EMT in oral squamous
carcinoma stem cells (32). Studies
have shown that TGF-β induces PLOD2 expression to promote EMT in
cervical cancer (33). In the
present study, there was no significant change in Gli1 and
p-β-catenin by HMGA2 depletion or overexpression. In addition, our
results showed that silencing of HMGA2 downregulated the mRNA and
protein levels of TGF-β and Smad2, while HMGA2 overexpression had
the opposite effect. In addition, TGF-β overexpression by transient
transfection partially abolished the anti-metastatic phenotype and
mesenchymal-epithelial transition (MET) by HMGA2 loss, whereas
TGF-β deficiency reversed the pro-metastatic effect and high
expression of TGF-β- and EMT-related markers by overexpression of
HMGA2. These results confirmed the vital role of the TGF-β
signaling pathway in HMGA2-mediated EMT.
In conclusion, our results revealed, for the first
time, that HMGA2 facilitated a metastatic phenotype and the EMT
process in renal cell carcinoma in vitro through a
TGF-β-dependent pathway. In addition, our findings strongly suggest
that HGMA2 may serve as a potential therapeutic target and
prognostic biomarker against renal cell carcinoma in the future,
although further investigation is needed.
Acknowledgements
This study was partially supported by the National
Natural Science Foundation of China (no. 81602562) and the
International Science and Technology Cooperative Project of Shaanxi
Province (no. 2017KW-063).
References
|
1
|
Fayaz MS, Al-Qaderi AE and El-Sherify MS:
Metastatic renal cell carcinoma with undetectable renal mass
presenting as lymphadenopathy. CEN Case Rep. 6:36–38. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu Y, Zeng S, Jiang X, Lai D and Su Z:
SOX4 induces tumor invasion by targeting EMT-related pathway in
prostate cancer. Tumour Biol. 39:10104283176945392017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chang JW, Gwak SY, Shim GA, Liu L, Lim YC,
Kim JM, Jung MG and Koo BS: EZH2 is associated with poor prognosis
in head-and-neck squamous cell carcinoma via regulating the
epithelial-to-mesenchymal transition and chemosensitivity. Oral
Oncol. 52:66–74. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xia P and Xu XY: Epithelial-mesenchymal
transition and gastric cancer stem cell. Tumour Biol.
39:10104283176983732017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kong FF, Qu ZQ, Yuan HH, Wang JY, Zhao M,
Guo YH, Shi J, Gong XD, Zhu YL, Liu F, et al: Overexpression of
FOXM1 is associated with EMT and is a predictor of poor prognosis
in non-small cell lung cancer. Oncol Rep. 31:2660–2668. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Luo W, Fang W, Li S and Yao K: Aberrant
expression of nuclear vimentin and related epithelial-mesenchymal
transition markers in nasopharyngeal carcinoma. Int J Cancer.
131:1863–1873. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Naber HP, Drabsch Y, Snaar-Jagalska BE,
ten Dijke P and van Laar T: Snail and Slug, key regulators of
TGF-β-induced EMT, are sufficient for the induction of single-cell
invasion. Biochem Biophys Res Commun. 435:58–63. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kumar MS, Armenteros-Monterroso E, East P,
Chakravorty P, Matthews N, Winslow MM and Downward J: HMGA2
functions as a competing endogenous RNA to promote lung cancer
progression. Nature. 505:212–217. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu J and Wei JJ: HMGA2 and high-grade
serous ovarian carcinoma. J Mol Med (Berl). 91:1155–1165. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Luo Y, Li W and Liao H: HMGA2 induces
epithelial-to-mesenchymal transition in human hepatocellular
carcinoma cells. Oncol Lett. 5:1353–1356. 2013.PubMed/NCBI
|
|
11
|
Liu B, Pang B, Hou X, Fan H, Liang N,
Zheng S, Feng B, Liu W, Guo H, Xu S, et al: Expression of
high-mobility group AT-hook protein 2 and its prognostic
significance in malignant gliomas. Hum Pathol. 45:1752–1758. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Langelotz C, Schmid P, Jakob C, Heider U,
Wernecke KD, Possinger K and Sezer O: Expression of
high-mobility-group-protein HMGI-C mRNA in the peripheral blood is
an independent poor prognostic indicator for survival in metastatic
breast cancer. Br J Cancer. 88:1406–1410. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang X, Liu X, Li AY, Chen L, Lai L, Lin
HH, Hu S, Yao L, Peng J, Loera S, et al: Overexpression of HMGA2
promotes metastasis and impacts survival of colorectal cancers.
Clin Cancer Res. 17:2570–2580. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xi YN, Xin XY and Ye HM: Effects of HMGA2
on malignant degree, invasion, metastasis, proliferation and
cellular morphology of ovarian cancer cells. Asian Pac J Trop Med.
7:289–292. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zou Q, Wu H, Fu F, Yi W, Pei L and Zhou M:
RKIP suppresses the proliferation and metastasis of breast cancer
cell lines through up-regulation of miR-185 targeting HMGA2. Arch
Biochem Biophys. 610:25–32. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao XP, Zhang H, Jiao JY, Tang DX, Wu YL
and Pan CB: Overexpression of HMGA2 promotes tongue cancer
metastasis through EMT pathway. J Transl Med. 14:262016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cai J, Shen G, Liu S and Meng Q:
Downregulation of HMGA2 inhibits cellular proliferation and
invasion, improves cellular apoptosis in prostate cancer. Tumour
Biol. 37:699–707. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sun J, Sun B, Zhu D, Zhao X, Zhang Y, Dong
X, Che N, Li J, Liu F, Zhao N, et al: HMGA2 regulates CD44
expression to promote gastric cancer cell motility and sphere
formation. Am J Cancer Res. 7:260–274. 2017.PubMed/NCBI
|
|
19
|
Dong J, Wang R, Ren G, Li X, Wang J, Sun
Y, Liang J, Nie Y, Wu K, Feng B, et al: HMGA2-FOXL2 axis regulates
metastases and epithelial-to-mesenchymal transition of
chemoresistant gastric cancer. Clin Cancer Res. 23:3461–3473. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang H, Jiang Z, Chen H, Wu X, Xiang J and
Peng J: MicroRNA-495 inhibits gastric cancer cell migration and
invasion possibly via targeting high mobility group AT-hook 2
(HMGA2). Med Sci Monit. 23:640–648. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Y, Fu QZ, Pu L, Meng QG, Liu XF, Dong
SF, Yang JX and Lv GY: HMGA2 expression in renal carcinoma and its
clinical significance. J Med Biochem. 34:338–343. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Na N, Si T, Huang Z, Miao B, Hong L, Li H
and Qiu J and Qiu J: High expression of HMGA2 predicts poor
survival in patients with clear cell renal cell carcinoma. Onco
Targets Ther. 9:7199–7205. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shi Z, Wu D, Tang R, Li X, Chen R, Xue S,
Zhang C and Sun X: Silencing of HMGA2 promotes apoptosis and
inhibits migration and invasion of prostate cancer cells. J Biosci.
41:229–236. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun M, Song CX, Huang H, Frankenberger CA,
Sankarasharma D, Gomes S, Chen P, Chen J, Chada KK, He C, et al:
HMGA2/TET1/HOXA9 signaling pathway regulates breast cancer growth
and metastasis. Proc Natl Acad Sci USA. 110:pp. 9920–9925. 2013;
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Natarajan S, Hombach-Klonisch S, Dröge P
and Klonisch T: HMGA2 inhibits apoptosis through interaction with
ATR-CHK1 signaling complex in human cancer cells. Neoplasia.
15:263–280. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang E, Cisowski J, Nguyen N, O'Callaghan
K, Xu J, Agarwal A, Kuliopulos A and Covic L: Dysregulated protease
activated receptor 1 (PAR1) promotes metastatic phenotype in breast
cancer through HMGA2. Oncogene. 35:1529–1540. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kaur H, Ali SZ, Huey L, Hütt-Cabezas M,
Taylor I, Mao XG, Weingart M, Chu Q, Rodriguez FJ, Eberhart CG, et
al: The transcriptional modulator HMGA2 promotes stemness and
tumorigenicity in glioblastoma. Cancer Lett. 377:55–64. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Micalizzi DS, Haber DA and Maheswaran S:
Cancer metastasis through the prism of epithelial-to-mesenchymal
transition in circulating tumor cells. Mol Oncol. 11:770–780. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xia YY, Yin L, Jiang N, Guo WJ, Tian H,
Jiang XS, Wu J, Chen M, Wu JZ and He X: Downregulating HMGA2
attenuates epithelial-mesenchymal transition-induced invasion and
migration in nasopharyngeal cancer cells. Biochem Biophys Res
Commun. 463:357–363. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li Y, Zhao Z, Xu C, Zhou Z, Zhu Z and You
T: HMGA2 induces transcription factor Slug expression to promote
epithelial-to-mesenchymal transition and contributes to colon
cancer progression. Cancer Lett. 355:130–140. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ke Z, Caiping S, Qing Z and Xiaojing W:
Sonic hedgehog-Gli1 signals promote epithelial-mesenchymal
transition in ovarian cancer by mediating PI3K/AKT pathway. Med
Oncol. 32:3682015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qiao B, He BX, Cai JH, Tao Q and King-Yin
Lam A: MicroRNA-27a-3p modulates the Wnt/β-catenin signaling
pathway to promote epithelial-mesenchymal transition in oral
squamous carcinoma stem cells by targeting SFRP1. Sci Rep.
7:446882017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu F, Zhang J, Hu G, Liu L and Liang W:
Hypoxia and TGF-β1 induced PLOD2 expression improve the migration
and invasion of cervical cancer cells by promoting
epithelial-to-mesenchymal transition (EMT) and focal adhesion
formation. Cancer Cell Int. 17:542017. View Article : Google Scholar : PubMed/NCBI
|