Introduction
Esophageal cancer, a highly malignant cancer, is the
sixth leading cause of cancer deaths in the world and has a 5-year
survival rate of less than 25% (1,2).
Esophageal adenocarcinoma and esophageal squamous cell carcinoma
(ESCC) are the two main types of esophageal cancer. In Asia and
Northern Iran, ESCC is the most common histological subtype of
esophageal cancer and accounts for 90% of all esophageal cancer
patients (3). Radiotherapy plays a
crucial part in the management of patients with inoperable and
locally advanced ESCC. Unfortunately, radioresistance results in
local cancer recurrence and poor prognosis in ESCC patients
(4). Thus, the development of novel
radiosensitizing agents, which can enhance the response of cancer
cells to radiation and improve the survival of patients with
radioresistant ESCC is clinically warranted and significant.
Although several potential radiosensitizers of ESCC have been
investigated, a satisfactory agent has not yet been discovered.
The radiosensitivity of cancer cells depends on
multiple factors, including the regulation of the cell cycle,
apoptosis, and interference with DNA-repair pathways. Studies have
shown that cells are most radiosensitive in the G2/M phase and
least radiosensitive in the S phase. Several radiosensitizers were
found to possess the ability to regulate the cell cycle and result
in arrest at the G2/M stage (5,6). The
apoptotic pathway is involved in tumor cell survival after
radiation. Thus, radiosensitivity can also be increased by
promoting radiation-induced apoptosis of cancer cells (7). In addition, one of the most pivotal
mechanisms of radiation-induced cancer cell death is DNA damage,
especially DNA double-strand breaks (DSBs). Inhibition of the
repair of radiation-induced DSBs has been demonstrated to increase
the radiosensitivity of cancer cells (8,9).
Several traditional Chinese medicines with antitumor
properties and only few side effects have been studied. Sinomenine
is an alkaloid extracted from the traditional Chinese herb
Sinomenium acutum. Sinomenine has pharmacologically relevant
properties such as anti-arthritic (10), anti-inflammatory (11), analgesic (12), and immunosuppressive effects
(13). Sinomenine hydrochloride
(SH) has been effectively used to treat rheumatoid arthritis in
clinical practice (14). More
recently, several studies have demonstrated that SH has
antineoplastic effects against various types of cancer, including
lung cancer (15), hepatic cancer
(16), breast cancer (17), osteosarcoma (18), and colon cancer (19). The anticancer effects of SH include
anti-metastasis, anti-angiogenesis, anti-proliferation, and
apoptosis induction. Furthermore, it has been found that SH
inhibits the proliferation of ESCC cell line Eca109, promotes
apoptosis, and significantly increases chemosensitivity of cancer
cells to 5-fluorouracil (20).
However, to our knowledge, no study has yet investigated the
radiosensitizing effect of SH on ESCC. With this in mind, the
present study was undertaken with a view to determining the effects
of SH on the radiosensitivity of ESCC cells and clarifying the
molecular mechanisms underpinning these effects.
Materials and methods
Cell culture and chemicals
Eca109 and EC9706, two human ESCC cell lines, were
obtained from the Chinese Academy of Sciences Cell Bank (Shanghai,
China). Dulbecco modified Eagle medium (DMEM; Gibco, Carlsbad, CA,
USA) supplemented with 10% fetal bovine serum (Sijichun
Bioengineering Materials Inc., Zhejiang, China) was used as the
cell culture medium. For some studies, SH (Sigma-Aldrich Co., St.
Louis, MO, USA) was dissolved in DMEM to achieve a concentration of
10 mM. Cell cultures were housed at 37°C in a humidified incubator
with 5% CO2. The CCK-8 kits and cell cycle analysis kits
were acquired from Beyotime Institute of Biotechnology (Jiangsu,
China). Annexin V-7AAD apoptosis-detection kits were purchased from
BD Biosciences (Franklin Lakes, NJ, USA).
Cell proliferation assay
ESCC cell viability was assessed using CCK-8 assays.
Cells were placed in 96-well plates (3×103 cells/well)
and then exposed to 0, 0.04, 0.4, 1, 2.5, or 5 mM SH for 24, 48, or
72 h. In the combination treatment group, cells were pretreated
with SH for 24 h, and X-rays (8 Gy) for another 24 h. After the
treatments, 10 µl CCK-8 was added, and the incubation was continued
for another 4 h at 37°C. We then measured the absorbance of each
well at 450 nm by using spectrophotometry. The procedure was
performed in triplicate, and average values are reported.
Clonogenic assay
Cells were pretreated with or without SH for 48 h,
then seeded in 6-well plates and irradiated at 0, 2, 4, 6, or 8 Gy
(200, 200, 600, 1,000 or 3,000 cells per well) with 4-MV X-rays by
using a linear accelerator (Simens, Munich, Germany). After
incubation for 14 days at 37°C with 5% CO2, the colonies
formed were washed with phosphate-buffered saline (PBS), fixed with
methanol, and stained with Giemsa. Only colonies containing more
than 50 cells were counted. The experiment was performed in
triplicate. The survival curves were fitted using the single-hit
multi-target model in GraphPad Prism 5 (GraphPad Software Inc., La
Jolla, CA, USA). Then, the D0 (mean lethal dose), Dq
(quai-threshold), and SF2 (survival fraction at 2 Gy) were obtained
based on the clonogenic assay. The sensitizing enhancement ratio
(SER) was calculated as the ratio of D0-control cells to
D0-SH-treated cells. The interaction between SH and radiation was
examined using the combination index (CI) method of Chou and
Talalay (21) and CompuSyn software
(Biosoft, Cambridge, UK). CI<1 indicates synergic effect, CI=1
indicates additive effect, and CI>1 indicates antagonistic
effect.
Cell cycle analysis
ESCC cells were seeded in 6-well plates at a density
of 2×105 cells per well and divided into four groups:
Control group, SH group, radiation group, and SH + radiation group.
In the SH group, cells were exposed to SH for 48 h, and in the
radiation group, cells were irradiated with 8 Gy X-rays for 24 h.
In the combination treatment group, cells were pretreated with SH
for 24 h and then irradiated at 8 Gy for another 24 h. After the
treatments, at least 1×106 cells were collected, fixed
with 70% ethanol (2 h, 4°C), and stained with propidium iodide and
RNase A (30 min, 37°C). We performed cell cycle analysis by using
flow cytometry (BD Biosciences).
Apoptosis assay
ESCC cells were divided into the following
experimental groups: SH group, radiation group, SH + radiation
group, and control group. ESCC cells (2×105 cells/well)
were seeded in 6-well plates and pretreated in the same way as they
were for the cell cycle analyses. Apoptosis was measured using flow
cytometry and the Annexin V-7AAD apoptosis-detection kit. At least
1×106 cells were incubated at 4°C with propidium iodide
and Annexin V-7AAD, and the percentage of apoptotic cells was
calculated using flow cytometry (BD Biosciences).
Western blot analysis
Cells were lysed with radioimmunoprecipitation assay
lysis and extraction buffers (Pioneer Technology, Xi'an, China),
separated using sodium dodecyl sulfate polyacrylamide gel
electrophoresis, and then transferred to polyvinylidene difluoride
membranes (Millipore, Billerica, MA, USA). The membranes were
incubated at 4°C with the following primary antibodies: Anti-Bax
(sc-20067, 1:1,000), anti-Bcl-2 (sc-509, 1:1,000), anti-Ku86
(sc-5280, 1:500), anti-Ku70 (sc-17789, 1:1,000), anti-Rad51
(sc-133089, 1:500), anti-cyclin B1 (sc-7393, 1:1,000), anti-CDK1
(sc-53219, 1:500), (all from Santa Cruz Biotechnology, Santa Cruz,
CA, USA), and anti-GAPDH (#5174, 1:3,000; Cell Signaling
Technology, Inc., Danvers, MA, USA). Then incubated with secondary
antibodies coupled with horseradish peroxidase at room temperature
for 1.5 h, anti-mouse (#4410, 1:5,000) or anti-rabbit (#4414,
1:5,000) antibodies (Cell Signaling Technology, Inc.). The
membranes were visualized using a chemiluminescence reagent
(Millipore) and the ChemiDoc System (Bio-Rad, Hercules, CA,
USA).
Xenograft tumor model
The animal experiments were approved by the
institutional animal Ethics Committee of the First Affiliated
Hospital of Xi'an Jiaotong University. The experimental protocol
complied with the animal ethics guidelines of the First Affiliated
Hospital of Xi'an Jiaotong University. Sixteen mice were housed in
sterile cages under standard condition (12-h light/dark cycles at
21±2°C) with ad libitum access to disinfected water and
food. An ESCC model was established in female BALB/c nude mice
(aged 4 weeks; Experimental Animal Center, Xi'an Jiaotong
University) by injecting 5×106 Eca109 cells
subcutaneously into the backs of the mice. When the tumor volume
was 100 mm3, the animals were randomly assigned to the
following groups (n=4 per group): radiation, SH, radiation + SH,
and control groups. SH was intraperitoneally injected at a dose of
75 mg/kg, once daily for 7 days. Tumors were treated with 4 Gy
X-rays for 3 consecutive days (total dose, 12 Gy), starting from
the second day of drug administration. The mice in the control
group were intraperitoneally inoculated with equal volumes of PBS.
Mouse body weight and tumor volume (length × width2 ×
0.5) were measured using calipers every 3 days for 30 days. All
mice were sacrificed using pentobarbital sodium at a dose of 100
mg/kg after 30 days, and the tumors were harvested.
Immunohistochemistry
Tumor tissue samples were fixed with 10% formalin,
paraffin embedded, and then stained with hematoxylin-eosin.
Immunohistochemical staining was performed according to the
standard protocol. Tumor-tissue sections were incubated overnight
at 4°C with primary antibodies against Ki-67 (sc-23900, 1:300;
Santa Cruz Biotechnology) and Bax (#5023, 1:300; Cell Signaling
Technology, Inc.), and then with anti-mouse or anti-rabbit
secondary antibodies for 1 h. Finally, images were captured using
microscopy, and five random fields were chosen in each specimen for
analysis.
Statistical analysis
The data were expressed as mean ± SEM. Statistical
analysis was performed using Graphpad Prism 5. Differences between
the control and treatment groups were tested using analysis of
variance (ANOVA) followed by Bonferroni's post-hoc test.
Differences were considered to be significant at P<0.05.
Results
SH inhibits ESCC cell growth and
enhances radiosensitivity of ESCC cells
To determine whether SH affected ESCC cell
proliferation, we treated ESCC cells with various concentration of
SH (0–5 mM) for 24–72 h. The CCK-8 assay was performed to estimate
cell viability. The results showed that SH significantly inhibited
ESCC cell viability in a time- and concentration-dependent manner
(P<0.05; Fig. 1A). In the case
of the 48 h treatment period, the half-maximal inhibitory
concentration (IC50) of SH for Eca109 and EC9706 cells
was 1.31 and 1.41 mM, respectively. We selected the 48 h
IC20 values (0.3 mM for Eca109 and 0.4 mM for EC9706) as
a appropriate concentration for the subsequent experiments. We then
evaluated the inhibitory effects of SH, radiation, and SH combined
with radiation on the proliferation of ESCC cells. The CCK-8 assay
showed that SH combined with radiation dramatically restrained ESCC
cell proliferation compared with SH or radiation group (P<0.05;
Fig. 1B).
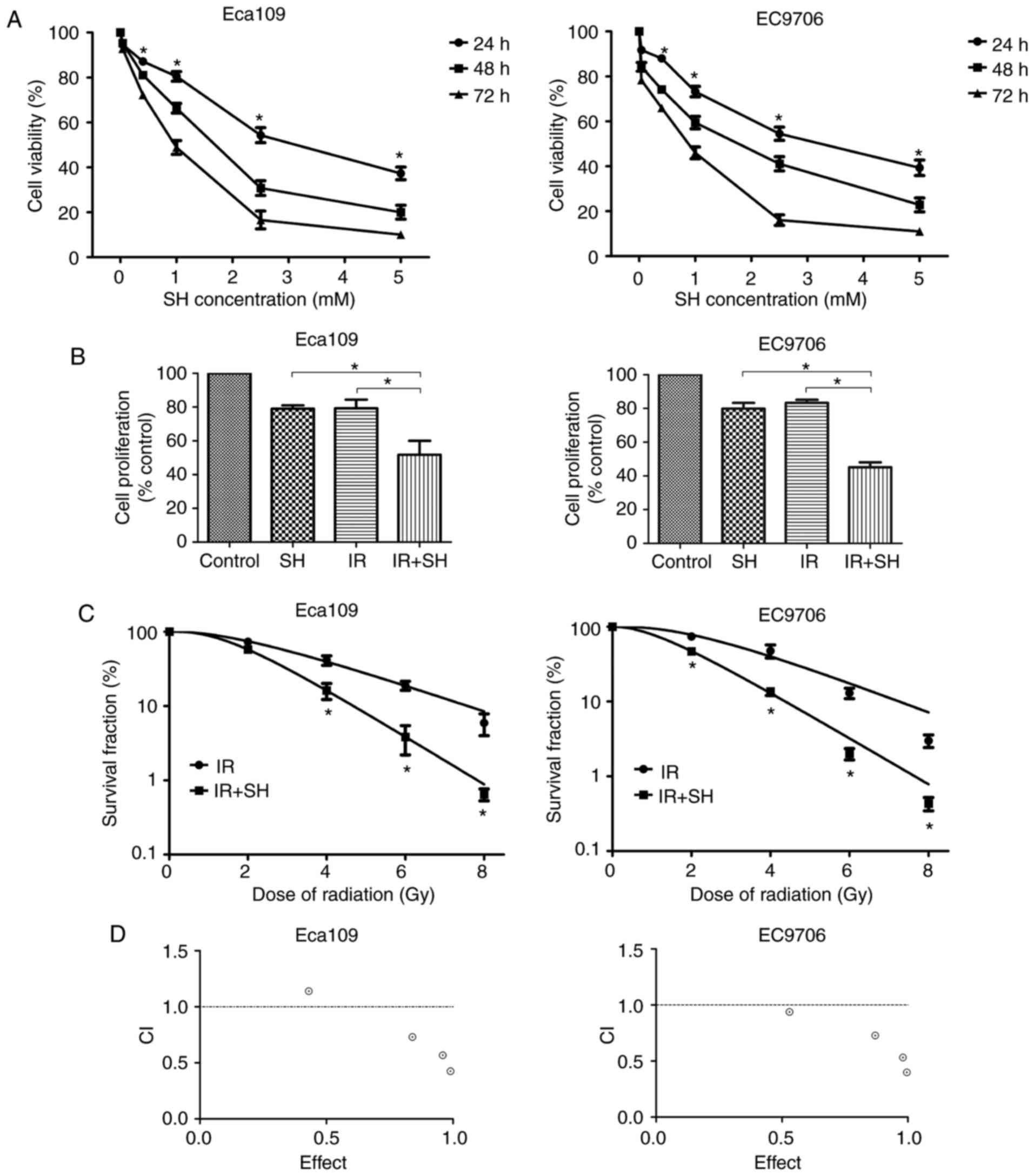 | Figure 1.SH enhances the radiosensitivity of
ESCC cells. (A) Eca109 and EC9706 cells were treated with SH (0,
0.04, 0.4, 1, 2.5, or 5 mM) for 24, 48, or 72 h, after which cell
viability was evaluated using the CCK-8 assay. (B) Cells were
pretreated with SH (0.3 mM for Eca109 and 0.4 mM for EC9706) and/or
exposed to 8 Gy X-rays, and then analyzed using the CCK-8 assay.
(C) Cells were pretreated with SH and exposed to 0, 2, 4, 6, or 8
Gy X-rays. After 14 days, colonies were stained and counted. The
survival curve was obtained using the multi-target model. (D) The
interaction between SH and radiation was examined using the
combination index (CI) method of Chou and Talalay and CompuSyn
software. CI=1, additive effect, CI<1, synergism, CI>1,
antagonism (*P<0.05). |
The radiosensitization effect of SH on ESCC cells
was assessed using the clonogenic assay. The results showed that SH
significantly improved the radiosensitivity of ESCC cells in
comparison with the control group (P<0.05; Fig. 1C). We calculated the radiation
parameters based on the results of the clonogenic survival assay.
The properties of a multi-target model in ESCC cells are detailed
in Table I. In the absence of SH,
the SF2 in Eca109 and EC9706 cells was 0.73 and 0.74, while after
treatment with SH, the SF2 decreased to 0.57 and 0.47,
respectively. The SER was 1.80 and 1.54 in Eca109 cells and EC9706
cells, respectively. CI values less than 1 indicated SH combined
with radiation resulted in synergic effect (Fig. 1D). These results indicate that SH
sensitized ESCC cells to radiotherapy.
 | Table I.The properties of a multi-target model
in ESCC cells as assessed through the clonogenic assay. |
Table I.
The properties of a multi-target model
in ESCC cells as assessed through the clonogenic assay.
| Cell line | D0 | Dq | SF2 | SER |
|---|
| Eca109 | 2.43 | 2.07 | 0.73 | 1.80 |
| Eca109+SH | 1.35 | 1.61 | 0.57 |
|
| EC9706 | 2.17 | 2.39 | 0.74 | 1.54 |
| EC9706+SH | 1.41 | 1.17 | 0.47 |
|
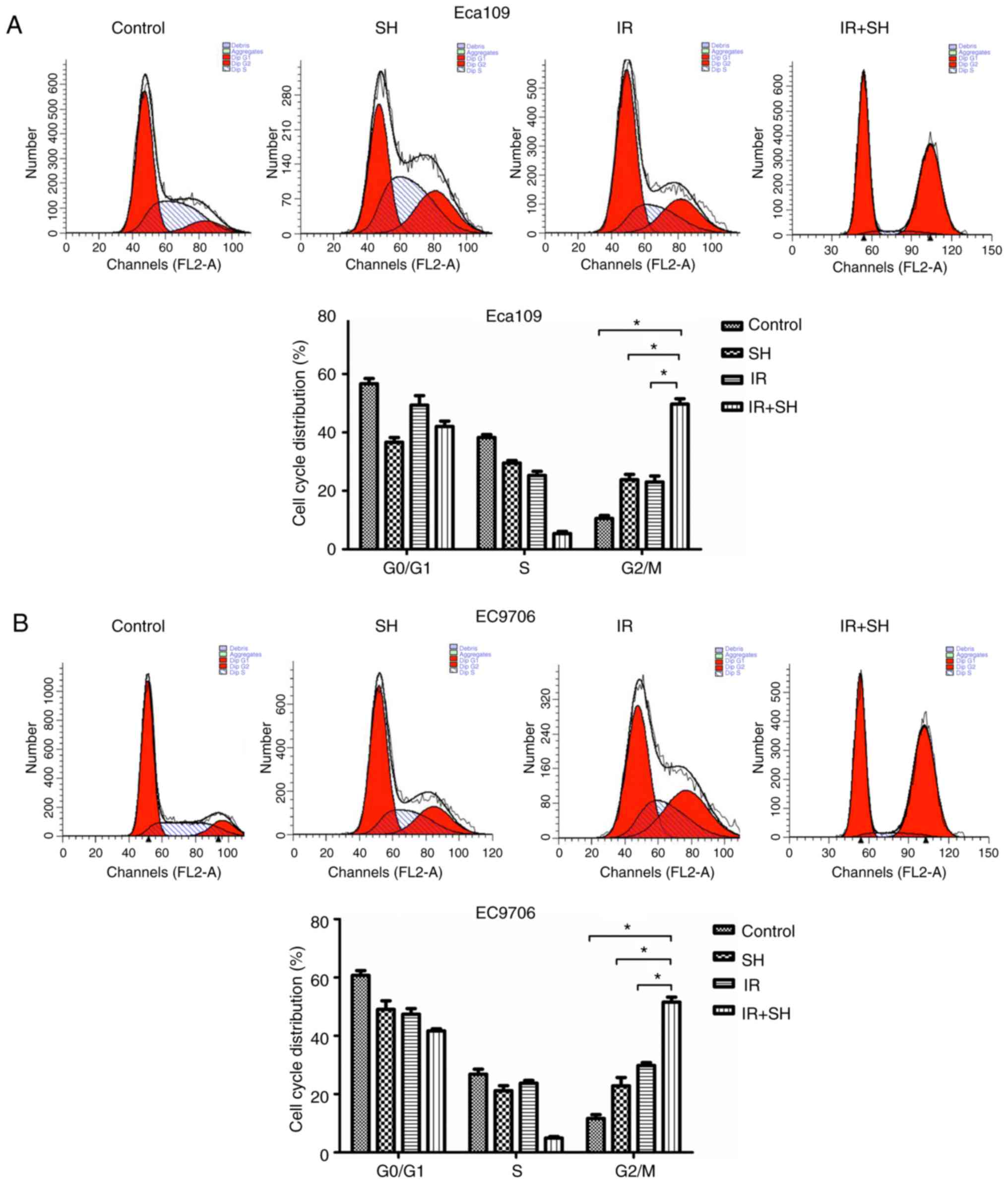
SH causes G2/M phase arrest in ESCC
cells
To observe the effect of SH on the cell cycle
distribution of ESCC cells, we used flow cytometry. Cell cycle
analysis demonstrated that SH combined with radiation arrested ESCC
cells in the G2/M phase (Fig. 2).
The number of cells in this phase was significantly higher in the
combination treatment group than in the SH, radiation, and control
groups (P<0.05). Thus, SH enhanced the radiosensitivity of ESCC
cells by increasing ratio of G2/M phase cells.
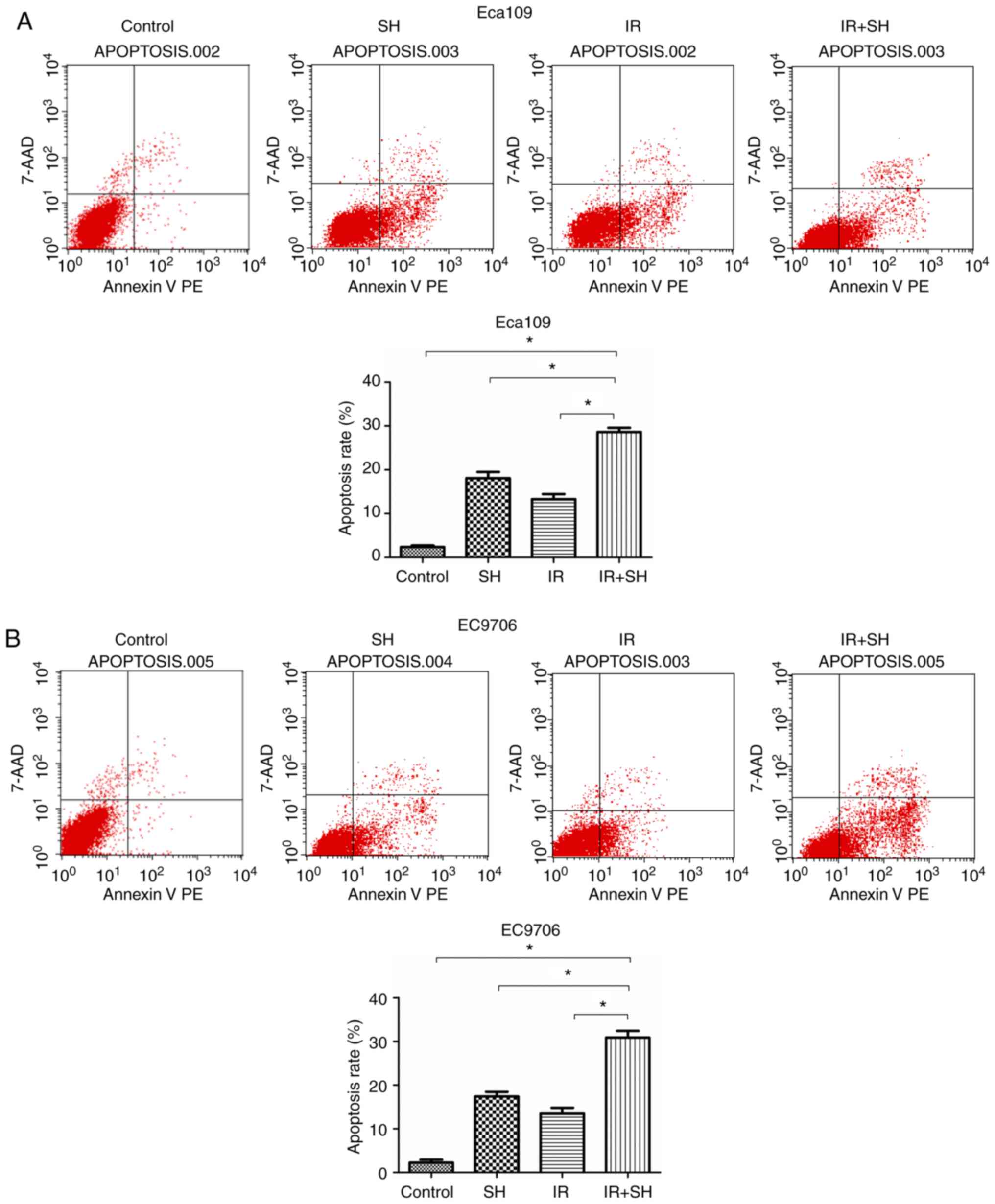
SH induces apoptosis of ESCC
cells
We performed Annexin V-PE/7AAD flow cytometry to
measure the effect of SH on apoptosis of ESCC cells. We found that
SH and radiation in combination increased the apoptosis ratio in
ESCC cells as compared with the control, SH and radiation group
(P<0.05; Fig. 3). Based on this
finding, SH increased radiosensitivity by promoting
radiation-induced apoptosis of ESCC cells.
SH upregulates Bax and downregulates
Bcl-2, cyclin B1, CDK1, Ku-70, Ku-86, and Rad51 in ESCC cells
To probe the molecular mechanism underpinning the
effects of SH on the radiosensitivity of ESCC cells, we examined
the expression of G2/M phase-associated proteins, apoptotic
proteins and DSB-repair proteins using western blot analysis. Our
findings showed that the combination of SH and 8 Gy irradiation
could upregulate the expression of Bax compared with the SH group
and radiation group; conversely, the expression of Bcl-2, cyclin
B1, CDK1, Ku-70, Ku-86, and Rad51 was downregulated in the
combination-treatment group compared with other three groups
(Fig. 4).
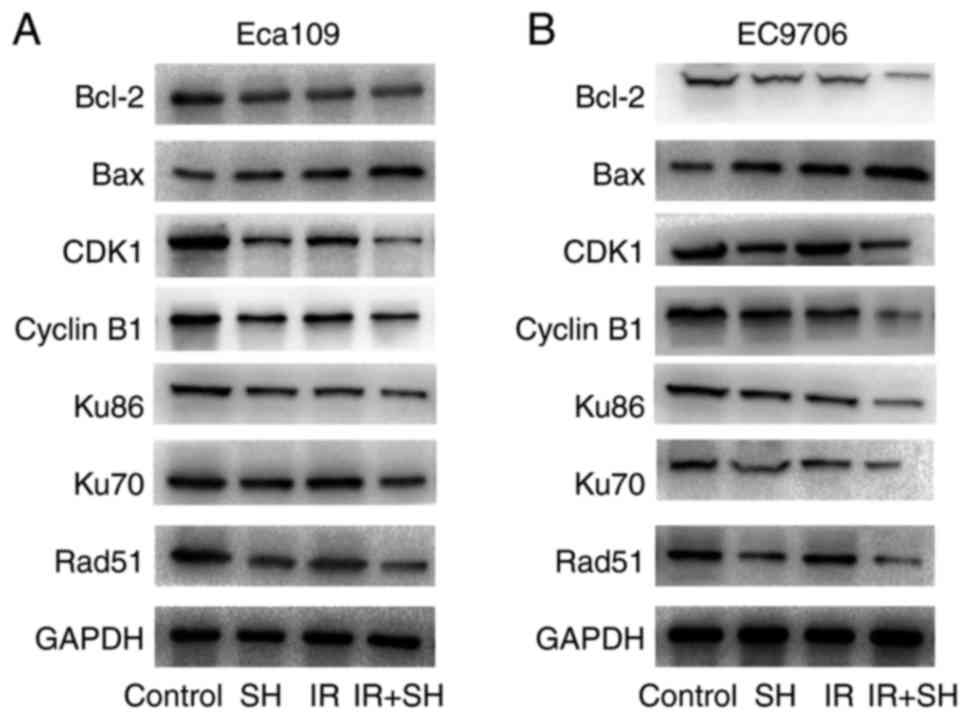 | Figure 4.SH regulates the expression of Bax,
Bcl-2, cyclin B1, CDK1, Ku-70, Ku-86, and Rad51 in ESCC cells. SH
markedly promoted Bax expression and inhibited Ku-70, Ku-86, Bcl-2,
cyclin B1, CDK1, and Rad51 expression. GAPDH was the internal
control. (A) Eca109, (B) EC9706. |
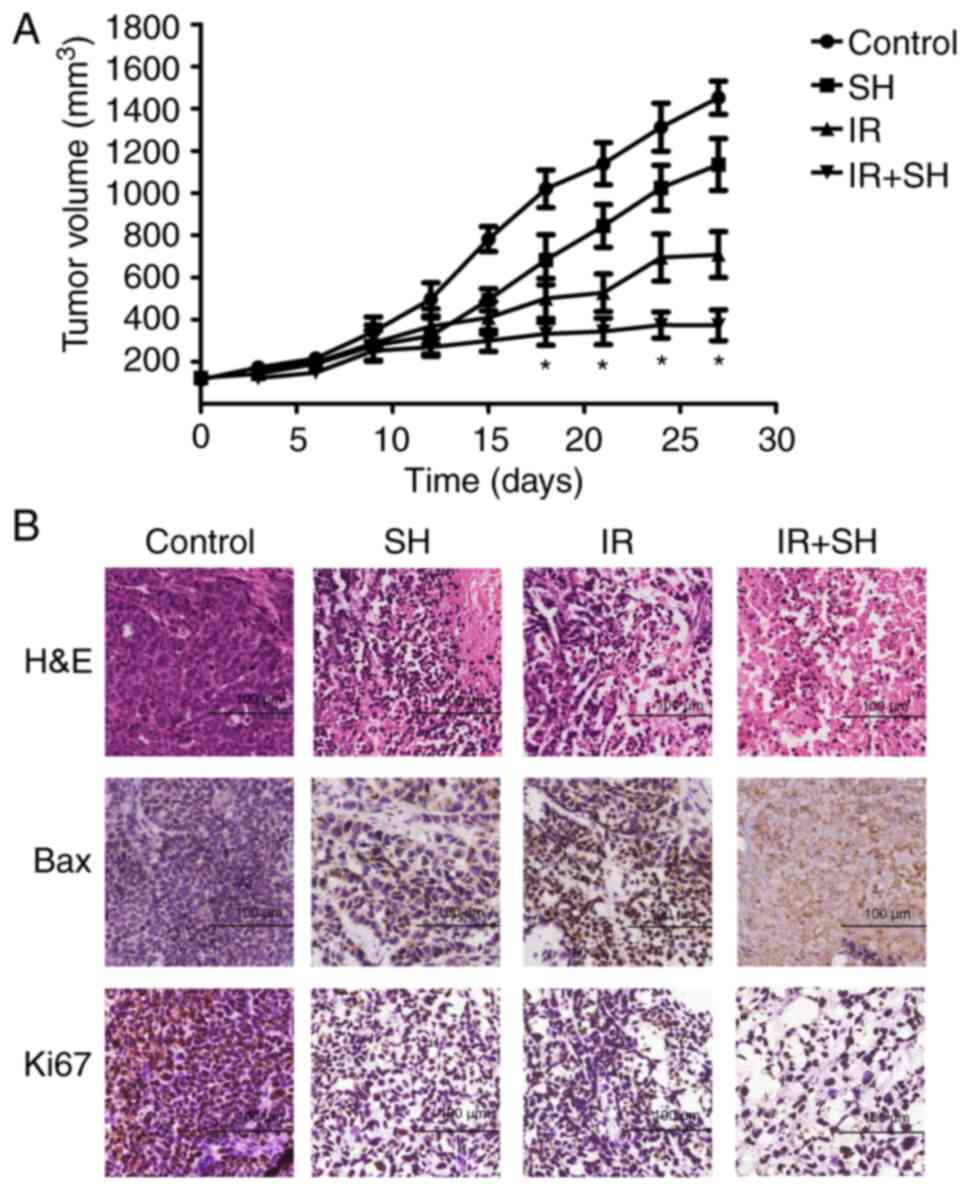
Combined treatment with radiation and
SH delays ESCC tumor growth in vivo
To determine whether SH affected the
radiosensitivity of ESCC cells in vivo, an ESCC model was
established by injecting Eca109 cells into nude mice. We found that
SH could considerably inhibit tumor growth. The tumor volume was
markedly smaller in the radiation + SH group than in the other
three groups (P<0.05; Fig. 5A).
Furthermore, immunohistochemical analysis showed that in the
combined treatment group, the expression of Bax was significantly
upregulated while that of Ki-67 was downregulated as compared with
the expressions in the other three groups (Fig. 5B).
Discussion
Radiotherapy is an effective therapeutic strategy
for inoperable and locally advanced ESCC, although it is sometimes
limited by the development of radioresistance (4). Thus, a radiosensitizer with a high
benefit-to-risk ratio is required. SH has been reported to exhibit
antitumor action by regulating cell proliferation, inhibiting
metastasis, and inducing apoptosis (15,22).
However, to date, whether or not SH affects the radiosensitivity of
ESCC cells is unknown. Our study demonstrated that SH can increase
the radiosensitivity of ESCC cells both in vitro and in
vivo. We found that SH impeded ESCC cell proliferation in a
time- and concentration-dependent manner and decreased the fraction
of cells surviving after irradiation. Furthermore,
radiosensitization of SH was related to G2/M phase arrest through
the downregulation of cyclin B1 and CDK1, apoptosis via the
regulation of Bcl-2 and Bax expression, and downregulation of Ku86,
Ku70, and Rad51 expression, which resulted in the inhibition of
DNA-damage repair.
The sensitivity of cells to radiation is related to
the cell cycle phase. Cells in the specific cell cycle phases
exhibit different degrees of radiosensitivity. In general, cells
are most sensitive to irradiation in the G2 and M phases and are
most radioresistant in the S phase. Cyclins and cyclin-dependent
kinases (CDKs) are directly involved in the progression of the cell
cycle (23). The cyclin B1/CDK1
complex plays an important role in G2/M phase transition (24). Numerous studies have demonstrated
that many radiosensitizing agents, such as astaxanthin, sunitinib,
and oxamate increase the cells in the G2/M cell cycle via
inhibiting the expression of cyclin B1 and CDK1, which enhances the
radiosensitivity of many malignant tumors (7,25,26).
Our study showed that radiation combined with SH could increase the
number of Eca109 and EC9706 cells at G2/M phase, suggesting that SH
arrested ESCC cells at G2/M phase and enhanced the lethal effects
of irradiation on ESCC cells. The results of western blot showed
that SH combined radiation significantly decreased the expression
of cyclin B1 and CDK1, which arrested ESCC cells at G2/M phase.
Apoptosis is considered to be the primary process of
cell death following radiotherapy (27). It is activated through different
pathways, including both intrinsic (mitochondrial death pathway)
and extrinsic pathways (death receptor pathway). The former pathway
is regulated by the balance of anti-apoptotic and apoptogenic
proteins, such as Bcl-2 and Bax, respectively (28). There is increasing evidence that the
cell-apoptosis rate usually depends on the ratio of anti- and
pro-apoptotic members, especially the ratio of Bcl-2 to Bax
(29). SH has been demonstrated to
promote apoptosis in various cancer cells (16,30).
Other anticancer agents, such as oroxylin A and astaxanthin, have
been shown to improve the sensitivity of radiation in cancer cells
by promoting Bax expression and inhibiting Bcl2 expression
(6,25). These findings suggest that
aberrations in Bcl2 and Bax expression may determine the cell fate
and the anticancer effect of radiation therapy. In this study, ESCC
cells were subjected to SH and radiation, both separately and in
combination. Our results demonstrated that radiation and SH could
synergistically increase the fraction of apoptotic cells. To
investigate the mechanisms underpinning this synergistic effect, we
performed western blot analysis. The results demonstrated that the
radiation + SH group had increased Bax expression compared with
other groups, in both Eca109 and EC9706 cells, which promoted cell
apoptosis. In addition, Bcl-2 expression was considerably inhibited
in the combination group. Furthermore, Bax expression in tumor
tissues was upregulated in the combination group, as revealed by
immunohistochemistry. Therefore, it seems that Bcl2 and Bax act as
downstream signals mediating the effects of SH combined with
radiation to induce apoptosis in ESCC cells.
The repair of DNA damage, especially double-strand
breaks (DSBs), is a vital factor determining radiosensitivity.
Non-homologous end joining (NHEJ) and homologous recombination (HR)
are two major pathways of DNA-DSBs repair. RAD51, Ku86, and Ku70
are critical downstream factors of the NHEJ and HR pathways. Thus,
the HR and NHEJ pathways are potential targets for radiosensitizing
agents. Studies have revealed that cancer cells can be rendered
more radiosensitive by inhibiting DNA-repair pathways. For example,
the inhibition of RAD51 enhanced the radiosensitivity of non-small
cell lung cancer cells, and the siRNA-mediated downregulation of
Ku80 expression can sensitize osteosarcoma cells to radiation
probably via telomere length shortening (31,32).
In this study, western blot analysis indicated that SH sensitizes
ESCC cells to radiation and impairs DNA repair via the
downregulation of Ku-70, Ku-86, and Rad51 expression. Therefore, SH
increased ESCC cell radiosensitivity by inhibiting DSB-repair
pathways.
In addition, SH combined with radiation considerably
inhibited the growth of tumor xenografts in vivo. Our data
showed that the observed suppression in tumor growth was related
with the inhibition of cell proliferation and the enhancement of
apoptosis. Immunohistochemical analysis showed that in the combined
treatment group, the expression of Bax was significantly higher
while that of Ki-67 was lower than the expressions in the other
three groups.
In conclusion, our study is the first to demonstrate
the effect of SH on the radiosensitivity of ESCC cells. Our
findings provide evidence that SH is a promising radiosensitizer
for improving the therapeutic efficacy of radiation in ESCC.
Acknowledgements
We are grateful to Medical Radiation Technologists
of Radiation Oncology, the First Affiliated Hospital of Xi'an
Jiaotong University for their significant contributions to this
study.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pennathur A, Gibson MK, Jobe BA and
Luketich JD: Oesophageal carcinoma. Lancet. 381:400–412. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Enzinger PC and Mayer RJ: Esophageal
cancer. N Engl J Med. 349:2241–2252. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu YM, Liu YK, Wang LW, Huang YC, Huang
PI, Tsai TH and Chen YJ: The medicinal fungus Antrodia cinnamomea
regulates DNA repair and enhances the radiosensitivity of human
esophageal cancer cells. Onco Targets Ther. 9:6651–6661. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tan C, Qian X, Ge Y, Yang B, Wang F, Guan
Z and Cai J: Oroxylin a could be a promising radiosensitizer for
esophageal squamous cell carcinoma by inducing G2/M arrest and
activating apoptosis. Pathol Oncol Res. 23:323–328. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhai X, Yang Y, Wan J, Zhu R and Wu Y:
Inhibition of LDH-A by oxamate induces G2/M arrest, apoptosis and
increases radiosensitivity in nasopharyngeal carcinoma cells. Oncol
Rep. 30:2983–2991. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kulkarni R, Thomas RA and Tucker JD:
Expression of DNA repair and apoptosis genes in mitochondrial
mutant and normal cells following exposure to ionizing radiation.
Environ Mol Mutagen. 52:229–237. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Choudhury A, Zhao H, Jalali F, Al Rashid
S, Ran J, Supiot S, Kiltie AE and Bristow RG: Targeting homologous
recombination using imatinib results in enhanced tumor cell
chemosensitivity and radiosensitivity. Mol Cancer Ther. 8:203–213.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Y, Fang Y, Huang W, Zhou X, Wang M,
Zhong B and Peng D: Effect of sinomenine on cytokine expression of
macrophages and synoviocytes in adjuvant arthritis rats. J
Ethnopharmacol. 98:37–43. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qian L, Xu Z, Zhang W, Wilson B, Hong JS
and Flood PM: Sinomenine, a natural dextrorotatory morphinan
analog, is anti-inflammatory and neuroprotective through inhibition
of microglial NADPH oxidase. J Neuroinflammation. 4:232007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kok TW, Yue PY, Mak NK, Fan TP, Liu L and
Wong RN: The anti-angiogenic effect of sinomenine. Angiogenesis.
8:3–12. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cheng Y, Zhang J, Hou W, Wang D, Li F,
Zhang Y and Yuan F: Immunoregulatory effects of sinomenine on the
T-bet/GATA-3 ratio and Th1/Th2 cytokine balance in the treatment of
mesangial proliferative nephritis. Int Immunopharmacol. 9:894–899.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yamasaki H: Pharmacology of sinomenine, an
anti-rheumatic alkaloid from sinomenium acutum. Acta Med Okayama.
30:1–20. 1976.PubMed/NCBI
|
|
15
|
Jiang T, Zhou L, Zhang W, Qu D, Xu X, Yang
Y and Li S: Effects of sinomenine on proliferation and apoptosis in
human lung cancer cell line NCI-H460 in vitro. Mol Med Rep.
3:51–56. 2010.PubMed/NCBI
|
|
16
|
Lu XL, Zeng J, Chen YL, He PM, Wen MX, Ren
MD, Hu YN, Lu GF and He S: Sinomenine hydrochloride inhibits human
hepatocellular carcinoma cell growth in vitro and in vivo:
Involvement of cell cycle arrest and apoptosis induction. Int J
Oncol. 42:229–238. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li X, Li P, Liu C, Ren Y, Tang X, Wang K
and He J: Sinomenine hydrochloride inhibits breast cancer
metastasis by attenuating inflammation-related
epithelial-mesenchymal transition and cancer stemness. Oncotarget.
8:13560–13574. 2017.PubMed/NCBI
|
|
18
|
Xie T, Ren HY, Lin HQ, Mao JP, Zhu T, Wang
SD and Ye ZM: Sinomenine prevents metastasis of human osteosarcoma
cells via S phase arrest and suppression of tumor-related
neovascularization and osteolysis through the CXCR4-STAT3 pathway.
Int J Oncol. 48:2098–2112. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang H, Yin P, Shi Z, Ma Y, Zhao C, Zheng
J and Chen T: Sinomenine, a COX-2 inhibitor, induces cell cycle
arrest and inhibits growth of human colon carcinoma cells in vitro
and in vivo. Oncol Lett. 11:411–418. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liao F, Yang Z, Lu X, Guo X and Dong W:
Sinomenine sensitizes gastric cancer cells to 5-fluorouracil in
vitro and in vivo. Oncol Lett. 6:1604–1610. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chou TC and Talalay P: Quantitative
analysis of dose-effect relationships: The combined effects of
multiple drugs or enzyme inhibitors. Adv Enzyme Regul. 22:27–55.
1984. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang S, Gao Y, Hou W, Liu R, Qi X, Xu X,
Li J, Bao Y, Zheng H and Hua B: Sinomenine inhibits A549 human lung
cancer cell invasion by mediating the STAT3 signaling pathway.
Oncol Lett. 12:1380–1386. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pawlik TM and Keyomarsi K: Role of cell
cycle in mediating sensitivity to radiotherapy. Int J Radiat Oncol
Biol Phys. 59:928–942. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lindqvist A, van Zon W, Karlsson Rosenthal
C and Wolthuis RM: Cyclin B1-Cdk1 activation continues after
centrosome separation to control mitotic progression. PLoS Biol.
5:e1232007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qian X, Tan C, Yang B, Wang F, Ge Y, Guan
Z and Cai J: Astaxanthin increases radiosensitivity in esophageal
squamous cell carcinoma through inducing apoptosis and G2/M arrest.
Dis Esophagus. 30:1–7. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ding YQ, Zhu HC, Chen XC, Sun XC, Yang X,
Qin Q, Zhang H, Yang Y, Yang YH, Gao L, et al: Sunitinib modulates
the radiosensitivity of esophageal squamous cell carcinoma cells in
vitro. Dis Esophagus. 29:1144–1151. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Maier P, Hartmann L, Wenz F and Herskind
C: Cellular pathways in response to ionizing radiation and their
targetability for tumor radiosensitization. Int J Mol Sci. 17(pii):
E1022016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cory S, Huang DC and Adams JM: The Bcl-2
family: Roles in cell survival and oncogenesis. Oncogene.
22:8590–8607. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Naseri MH, Mahdavi M, Davoodi J, Tackallou
SH, Goudarzvand M and Neishabouri SH: Up regulation of Bax and down
regulation of Bcl2 during 3-NC mediated apoptosis in human cancer
cells. Cancer Cell Int. 15:552015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li X, Wang K, Ren Y, Zhang L, Tang XJ,
Zhang HM, Zhao CQ, Liu PJ, Zhang JM and He JJ: MAPK signaling
mediates sinomenine hydrochloride-induced human breast cancer cell
death via both reactive oxygen species-dependent and -independent
pathways: An in vitro and in vivo study. Cell Death Dis.
5:e13562014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhong X, Luo G, Zhou X, Luo W, Wu X, Zhong
R, Wang Y, Xu F and Wang J: Rad51 in regulating the
radiosensitivity of non-small cell lung cancer with different
epidermal growth factor receptor mutation status. Thorac Cancer.
7:50–60. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hu L, Wu QQ, Wang WB, Jiang HG, Yang L,
Liu Y, Yu HJ, Xie CH, Zhou YF and Zhou FX: Suppression of Ku80
correlates with radiosensitivity and telomere shortening in the
U2OS telomerase-negative osteosarcoma cell line. Asian Pac J Cancer
Prev. 14:795–799. 2013. View Article : Google Scholar : PubMed/NCBI
|