Introduction
Head and neck squamous cell carcinoma (HNSCC) is the
sixth most common malignancy worldwide and the incidence rate in
developing countries is higher than the rate in developed countries
(1). The prognosis is poor and the
survival rate is barely over 50%. Oral leukoplakia (OL) is one of
the most common oral precancerous lesions. It was reported that the
percentage of malignant transformation rates of OL range from 17 to
24% of the patients with a median follow-up of >7 years
(2). There are no clinical
parameters that can predict the potential of malignant
transformation in patients with OL. The aim of the present study
was to investigate a potential role of SLPI in predicting the
development of oral squamous cell carcinoma (OSCC) in patients with
OL. Therefore, the development of effective diagnostic methods for
at-risk oral premalignant lesions (OPMLs) or early-stage OSCC is
essential in order to improve the survival rate of the patients as
well as alleviate patient pain.
By using in-depth mass spectrometry-based
quantitative shotgun proteomics study of non-invasively collected
oral brush biopsies, our previous study (3) indicated that SLPI abundance in OL
tissue samples is much lower than the expression of SLPI in human
normal oral epithelial tissues, with a more significant decrease in
OSCC tissues. Thus, it indicated that SLPI may be a promising,
mechanism-based oral cancer biomarker. In the present study, the
expression of SLPI in different histological stages of OL was
further investigated to determine whether SLPI is closely related
to carcinogenesis and may be used as an important reference for
grading, screening and treating dysplasia of oral mucosa. SLPI is a
11.7 kDa non-glycosylated serine protease inhibitor of a wide range
of serine proteases including neutrophil elastase, catharpin G,
chymotrypsin and trypsin. SLPI consists of 107 amino acids
possessing two homologous WFDC domains (4–7). It is
physiologically produced and secreted in the respiratory,
reproductive tract, oral, gastric and colonic mucosa, as well as
the lacrimal and salivary glands and other parts (4). According to previous research, the
expression of SLPI protein demonstrates diverse patterns in
different types of tumors. SLPI is highly upregulated in pancreatic
(8), gastric (9), lung (10,11),
endometrial (12) and ovarian
cancer (13). In contrast, it is
downregulated in human nasopharyngeal (14) and oral squamous cell carcinoma
(3,5,15).
Furthermore, the roles of SLPI in various types of tumors are not
entirely understood. It has been reported that SLPI exerts
pro-apoptotic and cell cycle-arrest effects in breast cancer
(16). On the contrary (17), SLPI, which provides protection
against paclitaxel-mediated cell injury, has pro-growth and
pro-survival effects in ovarian cancer. Similarly, it is considered
as a proliferation and survival factor for pancreatic cancer cells
(8). The role of SLPI in oral
cancer has not been entirely researched. However, Wen et al
(18) reported that SLPI protein
exerted an inhibitory effect on cell invasion and migration in oral
cancer cell lines. They provided evidence of a potential protective
role of SLPI in OSCC and suggested that SLPI may be used for a
possible stratification of oral cancer according to the risk of
occult metastasis. The aim of the present study was to further
investigate the potential predictive value of SLPI for the
identification of oral precancerous lesions with high risk of
progression to carcinoma and the biological effect in OL and OSCC,
especially in OPML. The effective risk prediction of the malignant
transformation of oral precancerous lesions is likely to
significantly improve the survival rate and to relieve patient
pain.
The regulatory mechanisms of SLPI on tumors are not
well known. One possible mechanism is through the NF-κB signaling
pathway. Several studies have reported that the SLPI protein
inhibits the activation of the NF-κB signaling pathway in
macrophages (19), alveolar
epithelial cells (20) and gingival
squamous cell carcinoma-derived Ca9-22 cells (21). It is known that the transcription
factor NF-κB is involved in the regulation of inflammatory
responses, cell growth, differentiation and apoptosis. Many of the
downstream target genes of the transcription factor NF-κB, such as
the baculoviral IAP repeat containing 2 (BIRC2), the baculoviral
IAP repeat containing 3 (BIRC3), the TRAF1 and TNF
receptor-associated factor 2 (TRAF2), are anti-apoptotic genes
(22). Furthemore, constitutive
activation of the NF-κB signaling pathway in HNSCC (23–25)
can promote the survival, growth, migration and secretion of
pro-inflammatory and pro-angiogenic cytokines.
In the present study, we aimed to investigate
whether the loss of SLPI protein was associated with oral cancer
progression using IHC. In addition, we explored the biological
effects of SLPI in OPML and OSCC cells and the possible regulatory
mechanism, especially focusing on changes in the NF-κB signaling
pathway.
Materials and methods
Patients and specimens
Sixty-seven formalin-fixed paraffin-embedded (FFPE)
specimens were obtained from the Department of Oral Histology and
Pathology, of the Shanghai Ninth People's Hospital Affiliated to
the Shanghai Jiao Tong University School of Medicine (Shanghai,
China) between May 2016 and December 2016. All patients provided
written informed consent. The specimens consisted of: 6 OL tissues
without dysplasia, 14 OL tissues with mild dysplasia, 10 OL tissues
with mild to moderate dysplasia, 7 OL tissues with moderate
dysplasia, 13 OL tissues with moderate to severe dysplasia and 17
OSCC specimens. The diagnosis of the samples was confirmed by oral
pathologists of the Shanghai Ninth People's Hospital according to
the World Health Organization (WHO) classification system in 2016.
The study was approved by the Institutional Ethics Committee of the
Shanghai Ninth People's Hospital, Affiliated to the Shanghai Jiao
Tong University School of Medicine.
Hematoxylin and eosin (H&E)
staining
H&E staining was conducted according to routine
protocol (26). After
deparaffinization and rehydration, 5-µm tissue sections were
stained with hematoxylin solution for 5 min, followed by 5
immersions in 1% acid ethanol (1% HCl in 70% ethanol), and then
rinsed in distilled water. Subsequently the sections were stained
with eosin solution for 3 min and then dehydrated with graded
alcohol and cleared in xylene.
IHC staining
The FFPE specimens were cut in 5-µm tissue sections
and then used for IHC as previously described (27,28).
The slides were incubated overnight at 4°C with primary antibody
diluted in primary antibody diluent (Beyotime Institute of
Biotechnology, Beijing, China). The primary antibody used was mouse
monoclonal anti-SLPI antibody (dilution 1:100; cat. no. ab17157;
Abcam, Cambridge, MA, USA). The negative staining control was
phosphate-buffered saline (PBS). For the antigen retrieval,
sections were pre-treated by boiling in 0.01 M sodium citrate
(Solarbio Science & Technology, Beijing, China) for 20 min in a
steaming pressure cooker. Endogenous peroxidase activity was then
blocked by 0.3% (v/v) H2O2 for 20 min. The
sections were incubated for 30 min at room temperature with
anti-mouse secondary antibody (cat. no. GK500705; Gene Tech
Biotechnology, Shanghai, China), stained with 3,39-diaminobenzidine
(DAB) (Gene Tech Biotechnology), and then counterstained with
haematoxylin for nuclear staining. The slides were observed under a
microscope with ×200 and ×400 magnification.
IHC scoring
IHC scoring was evaluated independently by two
authors as previously described (27). The results were assessed based on
staining intensity and staining area. The intensity score was the
average of different field scores as follows: 0, negative; 1, weak
staining; 2, distinctly enhanced staining; and 3, strong staining.
The staining area score was determined by the percentage of
positive cells (stained with DAB in the cytomembrane, cytoplasm and
cell nucleus) in the slide. Five random fields of one sample
(counting >500 cells/field) were analyzed and categorized as
follows: 0, 0% positive; 1, 1–25% positive; 2, 26–50% positive; 3,
51–75% positive and 4, 76–100% positive. For the statistical
analysis, the expression index (EI) for each specimen was
calculated by the following equation: staining intensity score ×
staining area score; the values given ranged from 0 to 12.
Cell lines
The human HNSCC cell lines WSU-HN4, HN6 and HN30
(kindly provided by the University of Maryland Dental School,
Baltimore, MD, USA) were cultured in Dulbecco's modified Eagle's
medium (DMEM; Basalmedia, Shanghai, China) supplemented with 10%
heat-inactivated FBS (Gibco; Thermo Fisher Scientific, Carlsbad,
CA, USA), penicillin (100 U/ml) and streptomycin (100 µg/ml) at
37°C in a humidified 5% CO2 atmosphere. SCC-9 and SCC-25
cells (kindly provided by the Shanghai Key Laboratory of
Stomatology, Shanghai, China) were cultured in DMEM/F12 medium
(Basalmedia) supplemented with 10% heat-inactivated FBS, penicillin
(100 U/ml) and streptomycin (100 µg/ml). Leuk1 cells (kindly
provided by Professor Li Mao of the University of Maryland Dental
School) were cultured in keratinocyte serum-free medium (KSF;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) with 0.2
ng/ml recombinant epidermal growth factor (rEGF). Leuk-1 cell line
is spontaneously derived from an OL lesion that was adjacent to
OSCC. It exhibits an immortalized but non-tumorigenic
phenotype.
Real-time polymerase chain reaction
(real-time PCR) and analysis
Total RNA was extracted with TRIzol reagent (Takara
Bio, Inc., Shiga, Japan) and 1 µg total RNA was reverse transcribed
into complementary DNA (cDNA) in a total volume of 20 µl using the
PrimerScript RT reagent kit (Takara Bio). Subsequently, 100 ng of
each cDNA was subjected to PCR amplification using the SYBR Premix
Ex Taq reagent kit (Takara Bio) by ABI 7300 real-time PCR system
(Applied Biosystems, Foster City, CA, USA). The primer sequences
were synthesized as displayed in Table
I. An initial denaturation step was performed for 5 min at 96°C
and 35 cycles were performed with the following PCR program:
denaturing at 96°C for 15 sec, annealing at 56°C for 30 sec and
elongating at 72°C for 50 sec. The program was completed with a
final extension step at 72°C for 5 min. The results were calculated
by the 2−ΔΔCt equation.
 | Table I.The primer sequences used in the
present study. |
Table I.
The primer sequences used in the
present study.
| Name | Forward primer | Reverse primer |
|---|
| SLPI |
5′-GCATCAAATGCCTGGATCCT-3′ |
5′-GCATCAAACATTGGCCATAAGTC-3′ |
| XIAP |
5′-AATAGTGCCACGCAGTCTACA-3′ |
5′-CAGATGGCCTGTCTAAGGCAA-3′ |
| BIRC2 |
5′-AGCACGATCTTGTCAGATTGG-3′ |
5′-GGCGGGGAAAGTTGAATATGTA-3′ |
| BIRC3 |
5′-AAGCTACCTCTCAGCCTACTTT-3′ |
5′-CCACTGTTTTCTGTACCCGGA-3′ |
| TRAF1 |
5′-TCCTGTGGAAGATCACCAATGT-3′ |
5′-GCAGGCACAACTTGTAGCC-3′ |
| TRAF2 |
5′-GCTCATGCTGACCGAATGTC-3′ |
5′-GCCGTCACAAGTTAAGGGGAA-3′ |
| BCL2L1 |
5′-GACTGAATCGGAGATGGAGACC-3′ |
5′-GCAGTTCAAACTCGTCGCCT-3′ |
| BCL2 |
5′-GAGGATTGTGGCCTTCTTTG-3′ |
5′-GCCGGTTCAGGTACTCAGTC-3′ |
| GAPDH |
5′-CGGGAAACTGTGGCGTGAT-3′ |
5′-GTCGCTGTTGAAGTCAGAGGAG-3′ |
Western blotting experiments
Sixty micrograms of total protein from the cell
lysates of the cell lines were separated by 12% SDS-PAGE. The
proteins were then transferred to a 0.22-µm PVDF membrane (EMD
Millipore, Billerica, MA, USA) and incubated with a goat polyclonal
anti-SLPI antibody (dilution 0.1 µg/ml; cat. no. AF1274; R&D
Systems, Minneapolis, MN, USA). The loading control was the mouse
monoclonal GAPDH antibody (dilution 1:5,000; cat. no. D190090;
Sangon Biotech Co., Ltd., Shanghai, China). The membrane was
labeled with HRP-labeled rabbit anti-goat monoclonal IgG (dilution
1:10,000; cat. no. WB0178; Biotechwell, Shanghai, China) or
HRP-labeled goat anti-mouse monoclonal IgG (dilution 1:10,000; cat.
no. WB0176; Biotechwell) and visualized with an ECL detection
system. The quantity of the total protein was assessed using the
BCA assay (Sangon Biotech).
Intake of exogenous SLPI
To ensure that exogenous SLPI (Sino Biological,
Beijing, China) was absorbed in human oral epithelial cells, the
Leica SP8 confocal laser scanning microscope (Leica Microsystems,
Wetzlar, Germany) was used to observe the localization of SLPI in
WSU-HN4 cells by immunofluorescent assay. SLPI protein (40 µg/ml)
or equal-volume ddH2O was added to the cell culture for
1 h. Images were captured using Zen-Software (Carl Zeiss AG,
Oberckochen, Germany).
Cell proliferation assay
As previously described (29), Leuk1 and WSU-HN4 cells were plated
in 96-well plates at concentrations of 5×103 and
8×103 cells/well, respectively. After the Leuk1 cells
had achieved adherence they were incubated with 20, 40 or 60 µg/ml
exogenous SLPI or ddH2O. In addition, the WSU-HN4 cells
were mixed with 10, 20 or 40 µg/ml exogenous SLPI or
ddH2O. In the beginning of the experiment, Leuk1 cells
were incubated with 20, 40 or 60 µg/ml exogenous SLPI or
equal-volume ddH2O. After 3 repetitions, the Leuk1 cells
were almost dead at the concentration of 60 µg/ml. Based on these
results, we reduced the concentration of SLPI in WSU-HN4 cell line.
Cell Counting Kit (CCK)-8 solution (10 µl; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) was added to each well at 0,
24, 48 and 72 h and incubated for 1 h. The number of viable cells
in each well were assessed by reading the optical density (OD) of
absorbance at 490 nm.
Apoptosis assay
Leuk1 and WSU-HN4 cells, which were separately
incubated with 40 µg/ml exogenous SLPI or equal-volume
ddH2O, were harvested at 12 h. At the concentration of
40 µg/ml, the outcomes of the CCK-8 assay were statistically
significant in Leuk1 and WSU-HN4 cells. Therefore, the
concentration of 40 µg/ml was chosen in the assay. According to the
manufacturer's protocols, both the adherent cells and the floating
cells were harvested, washed twice with cold PBS and resuspended in
1Χ binding buffer which was obtained from the FITC Annexin V
Apoptosis Detection kit (BD Biosciences, Bedford, MA, USA) at a
concentration of 1×106 cells/ml. Then, 5 µl FITC Annexin
V and 5 µl propidium iodide (PI) were added to cell suspension. The
cells were incubated for 15 min at 25°C in the dark, resuspended in
400 µl of 1X binding buffer and then quantified immediately by BD
LSR II flow cytometry (BD Biosciences).
Caspase-3/-7 activity assay
The Caspase-Glo® 3/7 assay (Promega,
Madison, WI, USA) is a homogeneous, luminescent assay that assesses
the activities of caspase-3 and −7. 4,000 cells/well of Leuk1 or
WSU-HN4 were seeded in 384-well plates for 24 h. Then different
concentrations (0, 10, 20 and 40 µg/ml) of SLPI protein were added
to the corresponding wells and incubated for 2, 4, 6, 8, 12 and 24
h. After incubation, 25 µl Caspase-Glo 3/7 reagent was added to
each well. The plate was gently mixed using a plate shaker at
300–500 rpm for 30 sec and the contents were incubated at room
temperature for 1 h. The luminescence of each sample was assessed
in a plate-reading luminometer (SpectraMax i3; Molecular Devices,
Sunnyvale, CA, USA).
NF-κB luciferase reporter gene
assay
The activation of NF-κB was determined by a reporter
gene assay. The cells were plated in the 24-well plate to a
confluency of 70–80%. After adherence, 600 ng of NF-κB luciferase
reporter gene constructs (Beyotime Institute of Biotechnology) was
co-transfected with 200 ng of Renilla luciferase (Beyotime
Institute of Biotechnology) which was used to normalize data for
transfection efficiency. After 24 h of transfection, the cells were
treated with recombinant human SLPI at 0, 20 or 30 µg/ml. In order
to ensure the green fluorescent protein expression of NF-κB
luciferase reporter gene, the cells were then cultivated for 12 h.
The cell lysates were analyzed via dual luciferase reporter gene
assay kit (Beyotime Institute of Biotechnology) on Modulus™ (Turner
Biosystems, Sunnyvale, CA, USA).
mRNA expression levels of various
anti-apoptotic proteins
The NF-κB signaling pathway regulates different
anti-apoptotic proteins, such as XIAP, BIRC2, BIRC3, TRAF1, TRAF2,
BCL2L1 and BCL2. When the cells achieved 80–90% confluency in
24-well plate, recombinant human SLPI was added to the
corresponding well at the concentration of 0, 10, 20 or 40 µg/ml
for 4 h. Using TRIzol-based RNA extraction method, the mRNA
expression levels of various anti-apoptotic proteins which were
subjected to the transcription factor NF-κB were assessed by
real-time PCR.
Statistical analysis
The relationship between the EI of SLPI and the
degree of OL was analyzed using Spearman's rank correlation
analysis. The correlation coefficient (r) was calculated to
determine the correlation degree between the EI of SLPI and the
degree of OL. Comparison of the expression of SLPI with clinical
parameters was evaluated using the χ2/Fisher's exact
test. Other outcomes were determined using Student's t test or
one-way ANOVA. Calculations were carried out by SPSS version 16.0
(SPSS, Inc., Chicago, IL, USA). A two-sided P-value of <0.05 was
considered to indicate statistically significant differences.
Results
Negative correlation between the SLPI
protein expression and the degree of OL
Firebrowse (http://firebrowse.org/), which is a website with data
from The Cancer Genome Atlas (TCGA) and demonstrates the different
expression of certain proteins in various cancer types through the
form of images, was used to analyze the different expression of
SLPI in 28 cancer types in the present study (Fig. 1A). It demonstrated that the
expression of SLPI had diverse patterns in different types of
tumors. As displayed, compared with matched normal tissues, the
expression of SLPI in OSCC decreased. To investigate the expression
of SLPI at different histological stages of OL, we used the method
of IHC. There was a negative association between the expression
level of SLPI and the pathological differentiation of oral tissues
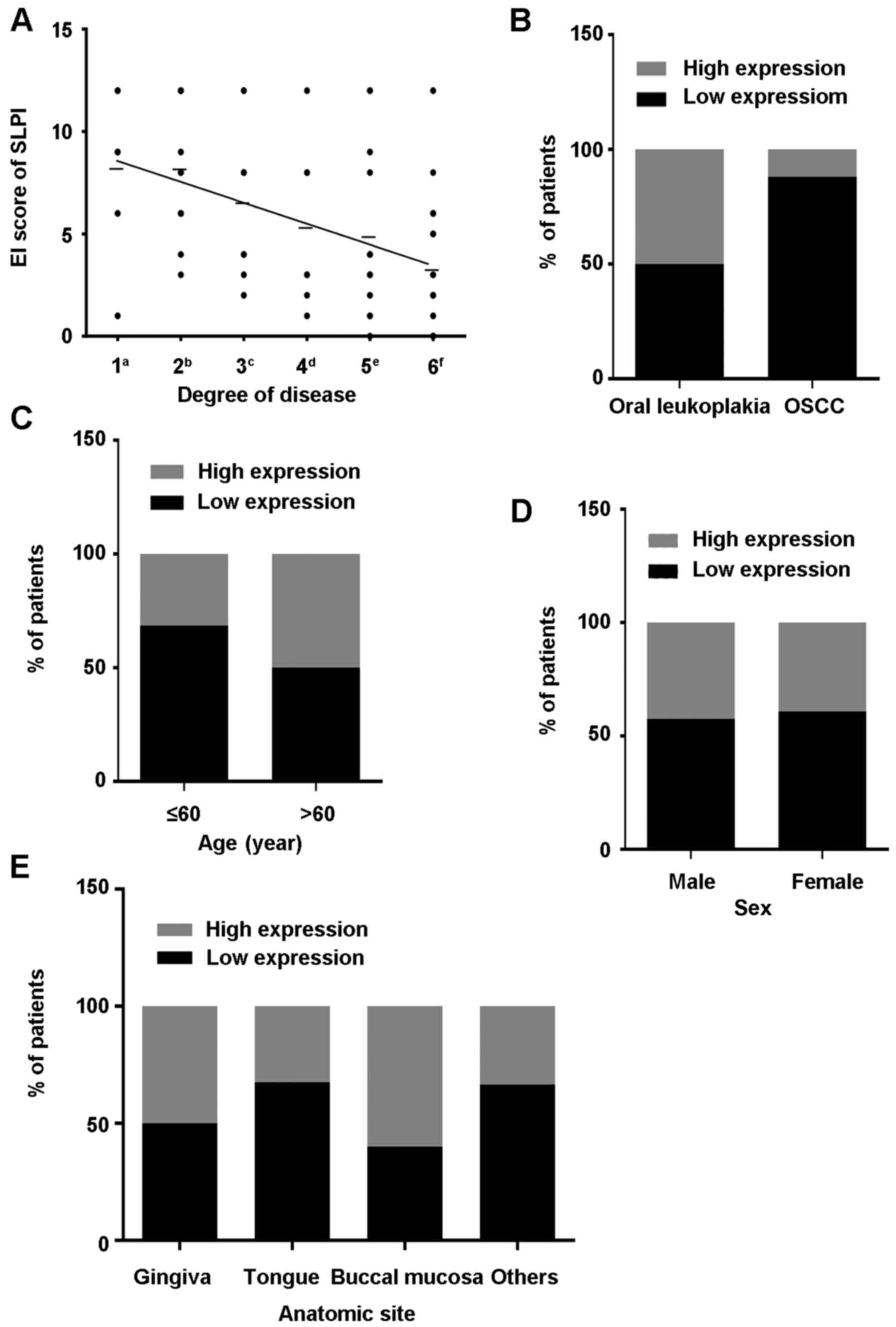
from various stages of OL to OSCC (r=−0.4922, P<0.05; Figs. 1B-H and 2A), whereas no significant associations
were determined between the expression pattern of SLPI and age
(P=0.142), sex (P=0.804) or anatomic site (P=0.286; Table II and Fig. 2C-E). In addition, the expression of
SLPI between OL and OSCC was significantly different (P<0.05;
Fig. 2B).
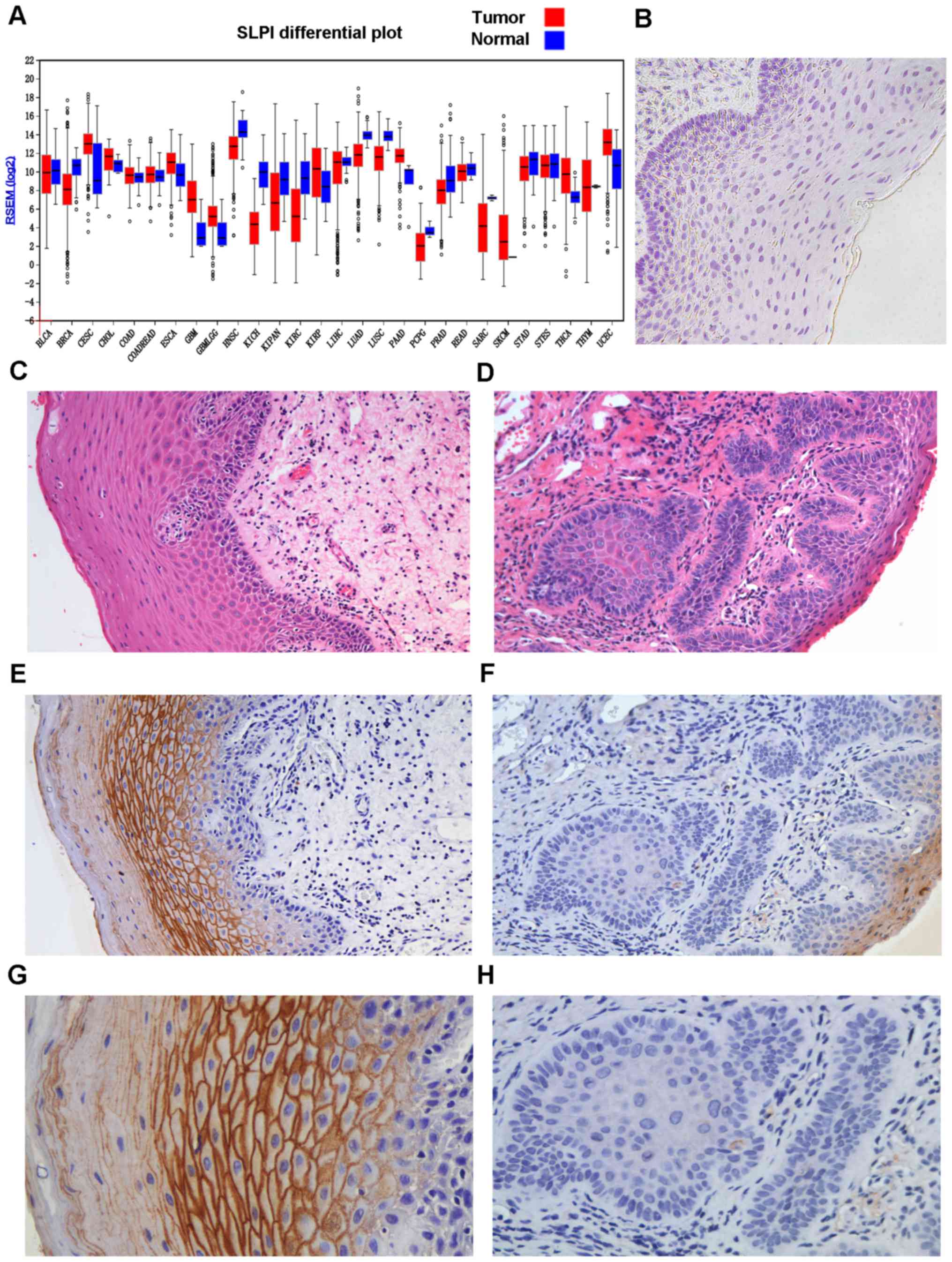 | Figure 1.(A) The mRNA expression of SLPI in
different tumors and protein expression of SLPI in OSCC and OPML by
IHC staining. The different mRNA expression of SLPI in tumor
tissues and matched normal tissues. Data from The Cancer Genome
Atlas (TCGA). Red, tumor tissues; Blue, matched normal tissues. (B)
Negative control (original magnification, ×100). Negative staining
control was phosphate-buffered saline (PBS). (C) The histological
image of H&E staining is from the same tissue sample displayed
in E and G (original magnification, ×200). (D) The histological
image of H&E staining is from the same tissue sample shown in F
and H (original magnification, ×200). (E) Strong SLPI
immunoreactivity is displayed in OL tissue sample with mild
dysplasia (original magnification, ×200). (F) Negative SLPI
immunoreactivity is detected in OSCC (original magnification,
×200). (G) Strong SLPI immunoreactivity in OL tissue sample with
mild dysplasia, which is from the same tissue sample shown in E
(original magnification, ×400). (H) Negative SLPI immunoreactivity
in OSCC, the same tissue sample displayed in Fig. 1F (original magnification, ×400).
BLCA, bladder urothelial carcinoma; BRCA, breast invasive
carcinoma; CESC, cervical and endocervical cancers; CHOL,
cholangiocarcinoma; COAD, colon adenocarcinoma; COADREAD, colon and
rectal adenocarcinoma; ESCA, esophageal carcinoma; GBM,
glioblastoma multiforme; GBMLGG, glioma; HNSC, head and neck
squamous cell carcinoma; KICH, kidney chromophobe; KIPAN,
pan-kidney cohort (KICH+KIRC+KIRP); KIRC, kidney renal clear cell
carcinoma; KIRP, kidney renal papillary cell carcinoma; LIHC, liver
hepatocellular carcinoma; LUAD, lung adenocarcinoma; LUSC, lung
squamous cell carcinoma; PAAD, pancreatic adenocarcinoma; PCPG,
pheochromocytoma and paraganglioma; PRAD, prostate adenocarcinoma;
READ, rectum adenocarcinoma; SARC, sarcoma; SKCM, skin cutaneous
melanoma; STAD, stomach adenocarcinoma; STES, stomach and
esophageal carcinoma; THCA, thyroid carcinoma; THYM, thymoma; UCEC,
uterine corpus endometrial carcinoma. |
| Table II.Correlation between the expression of
SLPI and the clinical parameters in OL and OSCC. |
Table II.
Correlation between the expression of
SLPI and the clinical parameters in OL and OSCC.
|
| Immunoreactivity
score, n (%) |
|
|---|
|
|
|
|
|---|
|
Characteristics | Low (≤6), n=40 | High (>6),
n=27 | P-value |
|---|
| Age (years) |
|
|
|
|
≤60 | 24 (68.57) | 11 (31.43) | 0.142 |
|
>60 | 16 (50.00) | 16 (50.00) |
|
| Sex |
|
|
|
|
Male | 15 (57.69) | 11 (42.31) | 0.804 |
|
Female | 25 (60.98) | 16 (39.02) |
|
| Anatomic site |
|
|
|
|
Gingiva | 3
(50.00) | 3
(50.00) | 0.286 |
|
Tongue | 27 (67.50) | 13 (32.50) |
|
| Buccal
mucosa | 6
(40.00) | 9
(60.00) |
|
|
Others | 4
(66.67) | 2
(33.33) |
|
| Disease |
|
|
|
| OL | 25 (50.00) | 25 (50.00) | 0.009 |
|
OSCC | 15 (88.24) | 2
(11.76) |
|
Different expression of SLPI in Leuk1
and HNSCC cell lines
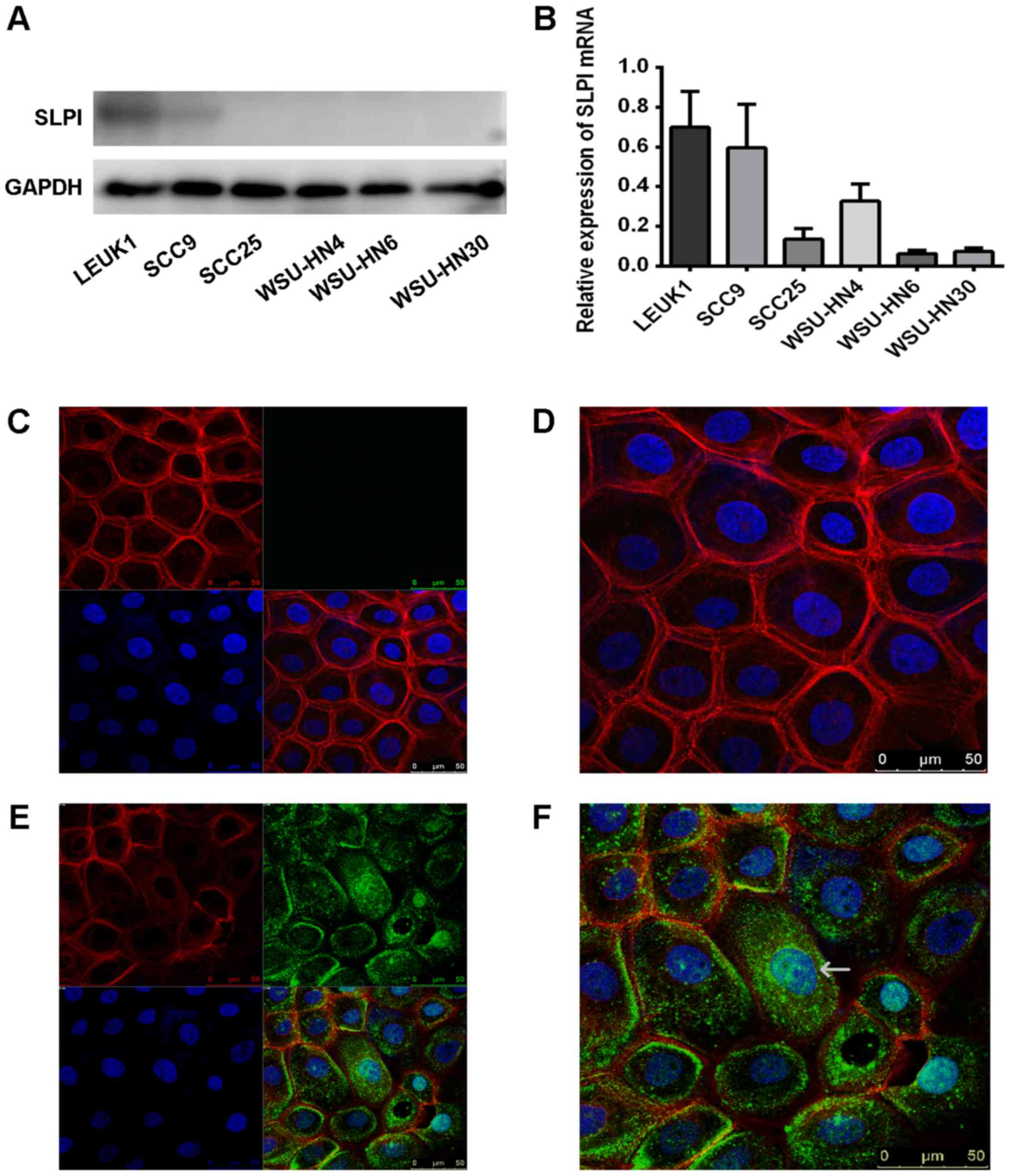
To confirm our previous results (3) and the IHC data, the levels of SLPI
protein in Leuk1 and 5 HNSCC cell lines were researched using
western blot analysis. The protein levels of SLPI in HNSCC cell
lines were significantly lower than the expression in the Leuk1
cell line (Fig. 3A). Furthermore,
compared with the mRNA level of SLPI in Leuk1 cells, the levels
were remarkably downregulated in most of the HNSCC cell lines by
real-time PCR (Fig. 3B).
Exogenous SLPI can be absorbed in
cells
According to previous research, the invasion effect
of SLPI in WSU-HN12 cell lines has been investigated (18). It is well-known (30) that the WSU-HN4 and WSU-HN12 cell
lines were derived from the same patient. Furthermore, the WSU-HN4
cell line was derived from the primary squamous cell carcinoma of
the tongue and it has an epithelial phenotype and low invasive
capacity. The WSU-HN12 cell line exhibits a mesenchymal phenotype
and has high invasive capacity. It was derived from a nodal
metastasis in the patient from whom the HN-4 cells originated. It
is considered an appropriate choice to research the other
biological effects of SLPI in the low invasive WSU-HN4 cell lines.
By comparing a blank group with the exogenous SLPI group in the
WSU-HN4 cells, we found that exogenous SLPI protein was absorbed in
cells. In some cells, partial proteins were also localized in the
nucleus (Fig. 3C-F). A previous
research (31), has demonstrated
that SLPI can enter the cytoplasmic and nuclear fractions of
macrophages and it was hypothesized that it may be the
arginine-rich nature of SLPI that enables it to interact with the
predominantly negatively-charged cell membrane and be internalized;
and SLPI is a small molecule (~11 kDa). Researchers have found that
it prevents the interaction of p65 with the NF-κB consensus region
by binding directly to the NF-κB binding sites in macrophages.
Thus, it may exhibit similar regulatory mechanisms in OPML and
OSCC. Future studies are needed to clarify this hypothesis.
Reduced cellular proliferation in
Leuk1 and WSU-HN4 cell lines with exogenous SLPI
To investigate the biological effects of SLPI
protein in OPML and OSCC, Leuk1 and WSU-HN4 cell lines were
incubated with exogenous SLPI protein for 0, 24, 48 and 72 h and
the proliferation rate was analyzed using CCK-8 assay. The
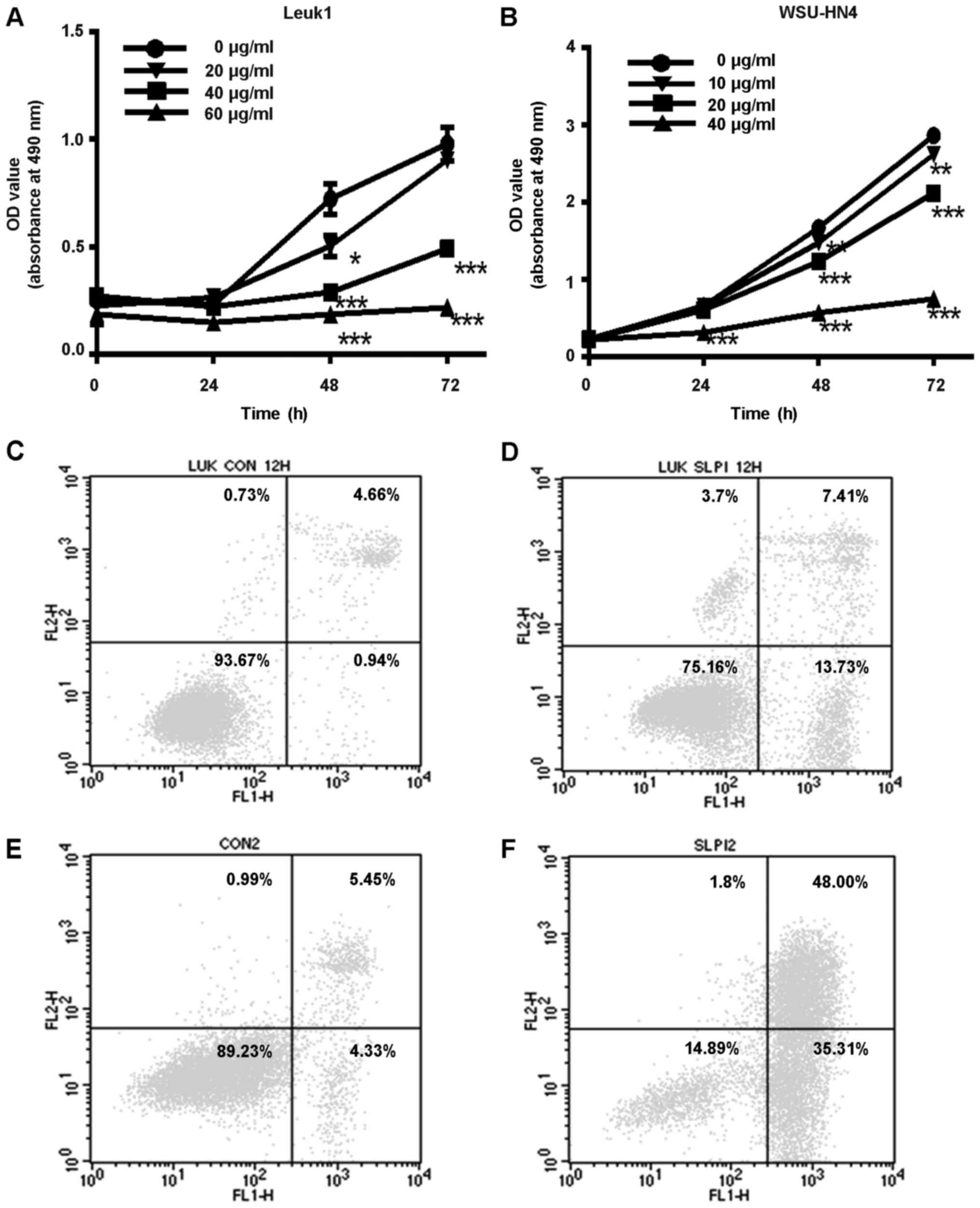
proliferation rates of Leuk1 cells significantly decreased both in
the 40-µg/ml exogenous SLPI protein group and in the 60-µg/ml group
(P<0.05, Fig. 4A). Due to the
fact that most cells were killed at a concentration of 60 µg/ml
SLPI protein, we reduced the concentration to 10, 20 or 40 µg/ml in
the WSU-HN4 cells. Similarly, compared with the control group, the
proliferation rate of the WSU-HN4 cells was significantly slower in
the 40-µg/ml exogenous SLPI protein group (P<0.05, Fig. 4B). These data revealed that SLPI
inhibited cell growth in Leuk1 and WSU-HN4 cell lines, especially
at the concentration of 40 µg/ml.
Exogenous SLPI promotes apoptosis in
the Leuk1 and WSU-HN4 cell lines
We investigated whether SLPI could inhibit cell
growth by arresting the cell cycle at some point, however the
evidence was not sufficient (data not shown). To analyze whether
the suppressive effect of SLPI protein was caused by pro-apoptotic
effect, Leuk1 and WSU-HN4 cells with exogenous SLPI or
ddH2O were assessed using PI and Annexin V. The
proportion of early apoptotic cells in Leuk1 and WSU-HN4 cells
incubated with exogenous SLPI was 13.73 and 35.31%, respectively,
which was notably higher than the ratio of early apoptotic cells in
the control cells with ddH2O (0.94 and 4.33%) (Fig. 4C-F). It is a new discovery that SLPI
has the pro-apoptotic biological effect both in OPML and in
OSCC.
SLPI protein activates caspase-3/-7 in
Leuk1 and WSU-HN4 cell lines
To verify that the SLPI protein can induce apoptosis
in Leuk1 and WSU-HN4 cell lines, we performed a quantitative
caspase-activity assay. Caspase-3 and −7 are known as caspase
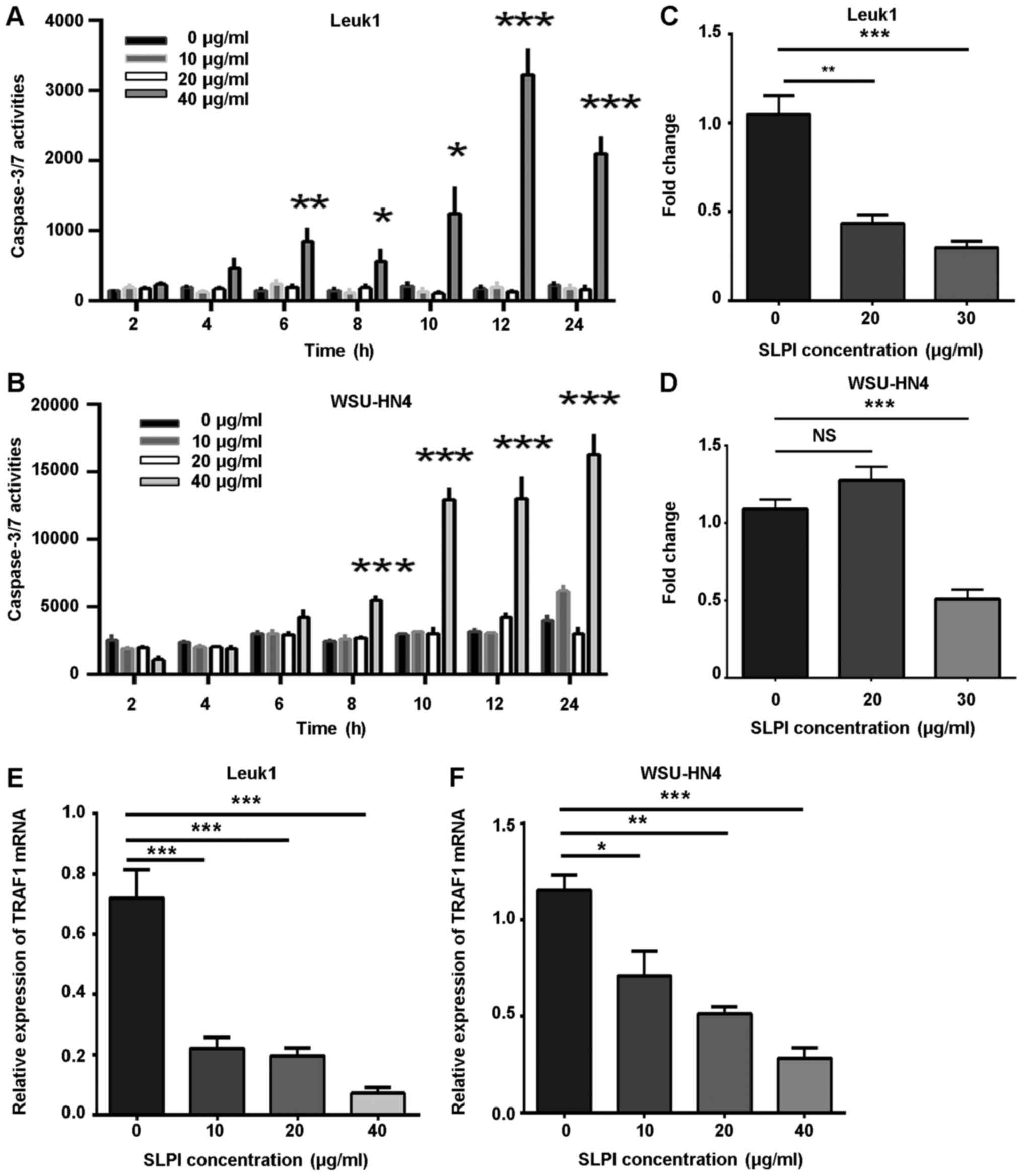
effectors and induce apoptosis. Our results revealed that the
activation of caspase-3/-7 was increased in a dose- and
time-dependent manner (Fig. 5A and
B). In addition, after 6 h, caspase-3/-7 was constantly
activated and lasted for >24 h.
Inhibition of SLPI on NF-κB activity
in Leuk1 and WSU-HN4 cell lines
NF-κB, an early response gene associated with SCC
progression, is implicated in the regulation of proinflammatory and
anti-apoptosis pathways. A previous study (31) has revealed that SLPI inhibited the
activation of NF-κB in macrophages. In the present study, in order
to explore the hypothesis that SLPI may promote oral precancerous
lesion and OSCC cell apoptosis by hindering the NF-κB-mediated
anti-apoptosis pathway, we used the NF-κB luciferase reporter assay
to investigate whether the SLPI-protein treatment decreased the
activation of the NF-κB gene in vitro. Our results
demonstrated that exogenous SLPI protein downregulated the
NF-κB-gene activity in Leuk1 and WSU-HN4 cell lines, in a
relatively dose-dependent fashion, as displayed in Fig. 5C and D. In addition, the Leuk1 cell
lines were more sensitive to SLPI protein. This initial data
indicated that SLPI may promote apoptosis in OPML and OSCC cells by
inhibiting the NF-κB anti-apoptosis pathway.
SLPI suppresses the mRNA expression
level of TRAF1
It is well-known that the transcription factor,
NF-κB, regulates numerous antiapoptotic proteins. Therefore, in the
present study, after incubating the Leuk1 and WSU-HN4 cell lines
with different concentrations of SLPI, we collected the total RNA
and determined the mRNA expression levels of XIAP, BIRC2, BIRC3,
TRAF1, TRAF2, BCL2L1 and BCL2, which are the downstream genes of
the NF-κB pathway. Compared with the control group, the mRNA
expression of TRAF1 in exogenous SLPI groups significantly
decreased (P<0.05; Fig. 5E and
F); the mRNA expression levels of the other genes were not
statistically significant (data not shown). To some extent, these
results may indicate that TRAF1 is likely to participate in the
apoptosis regulation of SLPI in OPML and OSCC. In the future, we
need to further investigate the relationship between the expression
of SLPI and TRAF1 and explore whether the NF-κB pathway is the main
pathway involved in the regulation of apoptosis.
Discussion
In our previous study (3), using non-invasive oral brush biopsies,
it was revealed that SLPI was markedly reduced in the OPML tissues
compared with normal oral mucosa. The abundance of SLPI protein in
the saliva of patients with OPML revealed the same changing
pattern. In addition, another study (18) reported that the content of SLPI
protein in the OSCC tissues was less than its expression in the
normal oral tissues. SLPI protein in well- and
moderate-differentiated OSCCs (32)
is more abundant than that in poorly differentiated ones. Based on
these findings, we aimed to study the correlation between the
abundance of SLPI protein and different histological stages of OL
by IHC. Our results indicated that along with the progressive
severity of dysplasia in OL tissues, the abundance of SLPI protein
gradually decreased. Furthermore, there is a statistically
significant difference in the expression of the SLPI protein
between OPML and OSCC (P<0.05). According to these results, SLPI
protein may be a potential tool to predict the possibility of
malignant transformation of oral precancerous lesions and perhaps
provide reliable evidence for early diagnosis and timely treatment
to reduce the incidence of OSCC. To verify these results, more
in vivo experiments are needed in the future.
The biological effects of the SLPI protein in OPML
and OSCC cells are not yet fully understood. According to previous
studies, SLPI has distinct functions in various types of tumors.
For instance, it was reported that SLPI promoted cell proliferation
and antagonized paclitaxel in ovarian cancer cells (17,33).
In the Lewis lung carcinoma 3LL-S cell line (34), SLPI promoted cell proliferation and
played an important role in the malignant progress. In addition,
SLPI promoted cell survival and growth of castration-resistant
prostate (35) and pancreatic
cancer (8). Paradoxically, Nakamura
et al observed that SLPI inhibited cell growth and induced
apoptosis in ovarian cancer cells (36). Furthermore, previous studies
(16,37,38)
also reported that SLPI induced apoptosis of mammary tumor cells
in vitro and decreased mammary tumor growth in vivo.
Thus, the role of this protein in cancer metastasis is dual. It can
promote metastasis in gastric (39)
and ovarian cancer (40), however,
in OSCC it may have a protective effect on the process of
metastasis (5,18). It is well-known that abnormal
apoptosis may induce the occurrence of cancer. The present study
revealed for the first time, that SLPI promoted apoptosis in OPML
and OSCC cell lines. Combined with its inhibition effect of
metastasis in OSCC, SLPI may have a potential protective role in
OSCC. In addition, many studies (41,42)
have revealed that proteins which inhibit cell proliferation and
induce apoptosis in OL can participate in the prevention of
malignant progression. SLPI may be a potential therapeutic drug for
patients with OPML and OSCC. In the present study, exogenous SLPI
significantly inhibited cell proliferation at the concentration of
40 µg/ml. Using exogenous SLPI in Leuk1 and WSU-HN4 cell lines, the
activation of caspase-3/-7 was increased in a dose- and
time-dependent manner, especially at the concentration of 40 µg/ml
and after 6 h and the activation lasted for >24 h. The changing
patterns of SLPI expression and the dual effects of SLPI in various
tumors are worth being further researched in depth.
Many pathways are involved in the regulation of cell
apoptosis, such as the P13K/Akt, NF-κB, p53 and MAPK signaling
pathways. Previous studies (31,43)
revealed that SLPI inhibits LPS-induced NF-κB activation in
monocytic cells, alveolar epithelial cells (20) and gingival squamous cell
carcinoma-derived Ca9-22 cells (21). One underlying mechanism is that SLPI
inhibited the degradation of IκBα without affecting the
phosphorylation and ubiquitination of IκBα in monocytic cells. SLPI
suppressed the NF-κB activation by preventing the IκBβ degradation
in Ca9-22 cells. Another theory is that SLPI can enter cell nucleus
and prevent p65 interaction with the NF-κB consensus region by
binding directly to the NF-κB binding sites. In the present study,
SLPI was localized in the nucleus of some cells and found in the
cytoplasm in more cells. We hypothesized that SLPI inhibited the
activation of NF-κB through the two ways aforementioned, especially
by inhibiting the IκBα or IκBβ degradation. However, this
hypothesis needs to be further investigated. Previous studies
(23–25) found that constitutive activation of
the NF-κB signaling pathway in HNSCC promoted survival, growth,
migration and the secretion of pro-inflammatory and pro-angiogenic
cytokines. According to these results, we assumed that SLPI may
regulate cell apoptosis in OPML and OSCC by inhibiting the NF-κB
signaling pathway. In the present study, due to the apoptosis
effect of SLPI, it was used at a concentration of <40 µg/ml to
ensure that sufficient cells could be collected to detect the
activation of the NF-κB reporter gene. We observed that, in a
relatively dose-dependent manner, SLPI significantly decreased the
activation of the NF-κB reporter gene in oral pre-malignant cells
and oral squamous carcinoma cells. And Leuk1 cell lines were more
sensitive to SLPI irritation. In previous research it was
demonstrated that the expression level of NF-κB in OSCC cells was
significantly higher than that in OPML and normal oral epithelial
tissues (44,45) and that NF-κB may be a valid target
for therapeutic approaches in patients with OSCC and potentially
malignant oral lesions. This may explain why SLPI is more
pronounced in Leuk1 cells and may have the ability to prevent OL
cancerization process. Subsequently, we analyzed the changes of the
downstream anti-apoptotic genes of the NF-κB pathway. We observed
that the level of TRAF1 mRNA in Leuk1 and WSU-HN4 cells obviously
decreased in a dose-dependent manner. Previous studies have
demonstrated that TRAF1 (46,47)
exerts a direct anti-apoptotic effect. For instance, along with
BIRC2 and BIRC3, the NF-kB-controlled expression of TRAF1 and TRAF2
(48) can provide maximum
protection against the TNF induced-apoptosis. TRAF1 transgenic mice
(49) have a defect in
antigen-induced apoptosis of the CD8+ T cells. Previous
research revealed that the expression of TRAF1 in response to H.
pylori infection in gastric epithelial cells was mainly
regulated by the activation of NF-kB. In addition, upregulation of
TRAF1 plays an anti-apoptotic role in H. pylori-infected
gastric cells (50). Using the
Kyoto Encyclopedia of Genes and Genomes (KEGG) database, we found
that TRAF1 also promoted anti-apoptosis activity via inducing the
NF-κB signaling pathway. These findings indicated that they can
regulate each other. We hypothesized that by using exogenous SLPI
protein, the NF-κB signaling pathway, which is suppressed by SLPI,
inhibits the expression of TRAF1. In turn, the reduced TRAF1 may
further inhibit NF-κB signaling pathway. However it is also
possible that SLPI firstly inhibits TRAF1 and then TRAF1 regulates
NF-κB. To verify this assumption, we should use TRAF1 and NF-κB
mutant types to observe the relevant changes of NF-κB and TRAF1
after using SLPI. In brief, it tentatively explored that SLPI
protein may promote cell apoptosis through the inhibition of the
NF-κB pathway. However, the detailed apoptosis regulatory mechanism
of the SLPI protein in OPML and OSCC is not sufficiently clarified
in the present study. This will be the focus of our next
experimental study.
In conclusion, this is the first time to find that
the abundance of SLPI protein is negatively correlated with the
grade of OL and it may have a potential prediction role for the
malignant transformation of OPML. The present study further
revealed that the SLPI protein exerted a pro-apoptotic effect in
OPML and OSCC cell lines. It promoted apoptosis, to some extent,
through suppression of the NF-κB pathway. The present study about
the apoptosis regulatory mechanism of SLPI is a tentative research.
In the future, more research is needed to elucidate the detailed
mechanism.
Acknowledgements
We would like to thank the patients who donated
valuable tissue samples. We would also like to thank the teachers
from the Shanghai Key Laboratory of Stomatology for their valuable
help. The present study was supported by grants from the Shanghai
Pujiang Program (no. 14pjd023).
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ding YM, Dong JH, Chen LL and Zhang HD:
Increased expression of galectin-1 is associated with human oral
squamous cell carcinoma development. Oncol Rep. 21:983–987.
2009.PubMed/NCBI
|
|
3
|
Yang Y, Rhodus NL, Ondrey FG, Wuertz BR,
Chen X, Zhu Y and Griffin TJ: Quantitative proteomic analysis of
oral brush biopsies identifies secretory leukocyte protease
inhibitor as a promising, mechanism-based oral cancer biomarker.
PLoS One. 9:e953892014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bouchard D, Morisset D, Bourbonnais Y and
Tremblay GM: Proteins with whey-acidic-protein motifs and cancer.
Lancet Oncol. 7:167–174. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cordes C, Häsler R, Werner C, Görögh T,
Röcken C, Hebebrand L, Kast WM, Hoffmann M, Schreiber S and
Ambrosch P: The level of secretory leukocyte protease inhibitor is
decreased in metastatic head and neck squamous cell carcinoma. Int
J Oncol. 39:185–191. 2011.PubMed/NCBI
|
|
6
|
Scott A, Weldon S and Taggart CC: SLPI and
elafin: Multifunctional antiproteases of the WFDC family. Biochem
Soc Trans. 39:1437–1440. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Williams SE, Brown TI, Roghanian A and
Sallenave JM: SLPI and elafin: One glove, many fingers. Clin Sci.
110:21–35. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zuo J, Zhang C, Ren C, Pang D, Li Y, Xie
X, Tang Z and Jiang X: Secretory leukocyte protease inhibitor is a
proliferation and survival factor for pancreatic cancer cells. Clin
Transl Oncol. 17:314–321. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cheng WL, Wang CS, Huang YH, Liang Y, Lin
PY, Hsueh C, Wu YC, Chen WJ, Yu CJ, Lin SR and Lin KH:
Overexpression of a secretory leukocyte protease inhibitor in human
gastric cancer. Int J Cancer. 123:1787–1796. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Brodt P, Fallavollita L, Khatib AM, Samani
AA and Zhang D: Cooperative regulation of the invasive and
metastatic phenotypes by different domains of the type I
insulin-like growth factor receptor beta subunit. J Biol Chem.
276:33608–33615. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jan Treda C, Fukuhara T, Suzuki T,
Nakamura A, Zaini J, Kikuchi T, Ebina M and Nukiwa T: Secretory
leukocyte protease inhibitor modulates urethane-induced lung
carcinogenesis. Carcinogenesis. 35:896–904. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Velarde MC, Parisek SI, Eason RR, Simmen
FA and Simmen RC: The secretory leukocyte protease inhibitor gene
is a target of epidermal growth factor receptor action in
endometrial epithelial cells. J Endocrinol. 184:141–151. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tsukishiro S, Suzumori N, Nishikawa H,
Arakawa A and Suzumori K: Use of serum secretory leukocyte protease
inhibitor levels in patients to improve specificity of ovarian
cancer diagnosis. Gynecol Oncol. 96:516–519. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tse KP, Wu CS, Hsueh C, Chang KP, Hao SP,
Chang YS and Tsang NM: The relationship between secretory leukocyte
protease inhibitor expression and Epstein-Barr virus status among
patients with nasopharyngeal carcinoma. Anticancer Res.
32:1299–1307. 2012.PubMed/NCBI
|
|
15
|
Quabius ES, Görögh T, Fischer GS, Hoffmann
AS, Gebhard M, Evert M, Beule A, Maune S, Knecht R, Óvári A, et al:
The antileukoprotease secretory leukocyte protease inhibitor (SLPI)
and its role in the prevention of HPV-infections in head and neck
squamous cell carcinoma. Cancer Lett. 357:339–345. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rosso M, Lapyckyj L, Amiano N, Besso MJ,
Sánchez M, Chuluyan E and Vazquez-Levin MH: Secretory Leukocyte
Protease Inhibitor (SLPI) expression downregulates E-cadherin,
induces β-catenin re-localisation and triggers apoptosis-related
events in breast cancer cells. Biol Cell. 106:308–322. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Simpkins FA, Devoogdt NM, Rasool N, Tchabo
NE, Alejandro EU, Kamrava MM and Kohn EC: The alarm anti-protease,
secretory leukocyte protease inhibitor, is a proliferation and
survival factor for ovarian cancer cells. Carcinogenesis.
29:466–472. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wen J, Nikitakis NG, Chaisuparat R,
Greenwell-Wild T, Gliozzi M, Jin W, Adli A, Moutsopoulos N, Wu T,
Warburton G and Wahl SM: Secretory leukocyte protease inhibitor
(SLPI) expression and tumor invasion in oral squamous cell
carcinoma. Am J Pathol. 178:2866–2878. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sano C, Shimizu T and Tomioka H: Effects
of secretory leukocyte protease inhibitor on the tumor necrosis
factor-alpha production and NF-kappaB activation of
lipopolysaccharide-stimulated macrophages. Cytokine. 21:38–42.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lentsch AB, Jordan JA, Czermak BJ, Diehl
KM, Younkin EM, Sarma V and Ward PA: Inhibition of NF-kappaB
activation and augmentation of IkappaBbeta by secretory leukocyte
protease inhibitor during lung inflammation. Am J Pathol.
154:239–247. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mikami Y, Iwase T, Komiyama Y, Matsumoto
N, Oki H and Komiyama K: Secretory leukocyte protease inhibitor
inhibits expression of polymeric immunoglobulin receptor via the
NF-κB signaling pathway. Mol Immunol. 67:568–574. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Barkett M and Gilmore TD: Control of
apoptosis by Rel/NF-kappaB transcription factors. Oncogene.
18:6910–6924. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Duffey DC, Chen Z, Dong G, Ondrey FG, Wolf
JS, Brown K, Siebenlist U and Van Waes C: Expression of a
dominant-negative mutant inhibitor-kappaBalpha of nuclear
factor-kappaB in human head and neck squamous cell carcinoma
inhibits survival, proinflammatory cytokine expression, and tumor
growth in vivo. Cancer Res. 59:3468–3474. 1999.PubMed/NCBI
|
|
24
|
Nottingham LK, Yan CH, Yang X, Si H,
Coupar J, Bian Y, Cheng TF, Allen C, Arun P, Gius D, et al:
Aberrant IKKα and IKKβ cooperatively activate NF-κB and induce
EGFR/AP1 signaling to promote survival and migration of head and
neck cancer. Oncogene. 33:1135–1147. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ondrey FG, Dong G, Sunwoo J, Chen Z, Wolf
JS, Crowl-Bancroft CV, Mukaida N and Van Waes C: Constitutive
activation of transcription factors NF-(kappa)B, AP-1, and NF-IL6
in human head and neck squamous cell carcinoma cell lines that
express pro-inflammatory and pro-angiogenic cytokines. Mol
Carcinog. 26:119–129. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu H, Zhu R, Liu C, Ma R, Wang L, Chen B,
Li L, Niu J, Zhao D, Mo F, et al: Evaluation of decalcification
techniques for rat femurs using HE and immunohistochemical
staining. Biomed Res Int. 2017:90507542017.PubMed/NCBI
|
|
27
|
Cai B, Miao Y and Liu Y, Xu X, Guan S, Wu
J and Liu Y: Nuclear multidrug-resistance related protein 1
contributes to multidrug-resistance of mucoepidermoid carcinoma
mainly via regulating multidrug-resistance protein 1: A human
mucoepidermoid carcinoma cells model and Spearman's rank
correlation analysis. PLoS One. 8:e696112013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dalley AJ, Pitty LP, Major AG, Abdulmajeed
AA and Farah CS: Expression of ABCG2 and Bmi-1 in oral potentially
malignant lesions and oral squamous cell carcinoma. Cancer Med.
3:273–283. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Okamoto A, Higo M, Shiiba M, Nakashima D,
Koyama T, Miyamoto I, Kasama H, Kasamatsu A, Ogawara K, Yokoe H, et
al: Down-regulation of nucleolar and spindle-associated protein 1
(NUSAP1) expression suppresses tumor and cell proliferation and
enhances anti-tumor effect of paclitaxel in oral squamous cell
carcinoma. PLoS One. 10:e01422522015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu S, Ye D, Guo W, Yu W, He Y, Hu J, Wang
Y, Zhang L, Liao Y, Song H, et al: G9a is essential for
EMT-mediated metastasis and maintenance of cancer stem cell-like
characters in head and neck squamous cell carcinoma. Oncotarget.
6:6887–6901. 2015.PubMed/NCBI
|
|
31
|
Taggart CC, Cryan SA, Weldon S, Gibbons A,
Greene CM, Kelly E, Low TB, O'neill SJ and McElvaney NG: Secretory
leucoprotease inhibitor binds to NF-kappaB binding sites in
monocytes and inhibits p65 binding. J Exp Med. 202:1659–1668. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Westin U, Nyström M, Ljungcrantz I,
Eriksson B and Ohlsson K: The presence of elafin, SLPI, IL1-RA and
STNFalpha RI in head and neck squamous cell carcinomas and their
relation to the degree of tumour differentiation. Mediators
Inflamm. 11:7–12. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rasool N, LaRochelle W, Zhong H, Ara G,
Cohen J and Kohn EC: Secretory leukocyte protease inhibitor
antagonizes paclitaxel in ovarian cancer cells. Clin Cancer Res.
16:600–609. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Devoogdt N, Hassanzadeh Ghassabeh G, Zhang
J, Brys L, De Baetselier P and Revets H: Secretory leukocyte
protease inhibitor promotes the tumorigenic and metastatic
potential of cancer cells. Proc Natl Acad Sci USA. 100:pp.
5778–5782. 2003; View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zheng D, Gui B, Gray KP, Tinay I, Rafiei
S, Huang Q, Sweeney CJ, Kibel AS and Jia L: Secretory leukocyte
protease inhibitor is a survival and proliferation factor for
castration-resistant prostate cancer. Oncogene. 35:4807–4815. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nakamura K, Takamoto N, Hongo A, Kodama J,
Abrzua F, Nasu Y, Kumon H and Hiramatsu Y: Secretory leukoprotease
inhibitor inhibits cell growth through apoptotic pathway on ovarian
cancer. Oncol Rep. 19:1085–1091. 2008.PubMed/NCBI
|
|
37
|
Amiano NO, Costa MJ, Reiteri RM, Payés C,
Guerrieri D, Tateosian NL, Sánchez ML, Maffia PC, Diament M, Karas
R, et al: Anti-tumor effect of SLPI on mammary but not colon tumor
growth. J Cell Physiol. 228:469–475. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Amiano N, Reiteri RM, Costa MJ, Tateosian
N and Chuluyan HE: Immunotherapy with SLPI over-expressing mammary
tumor cells decreases tumor growth. Cancer Immunol Immunother.
60:895–900. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Choi BD, Jeong SJ, Wang G, Park JJ, Lim
DS, Kim BH, Cho YI, Kim CS and Jeong MJ: Secretory leukocyte
protease inhibitor is associated with MMP-2 and MMP-9 to promote
migration and invasion in SNU638 gastric cancer cells. Int J Mol
Med. 28:527–534. 2011.PubMed/NCBI
|
|
40
|
Devoogdt N, Rasool N, Hoskins E, Simpkins
F, Tchabo N and Kohn EC: Overexpression of protease inhibitor-dead
secretory leukocyte protease inhibitor causes more aggressive
ovarian cancer in vitro and in vivo. Cancer Sci.
100:434–440. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li Y, Li LJ, Zhang ST, Wang LJ, Zhang Z,
Gao N, Zhang YY and Chen QM: In vitro and clinical studies of gene
therapy with recombinant human adenovirus-p53 injection for oral
leukoplakia. Clin Cancer Res. 15:6724–6731. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tsuzuki H, Fujieda S, Sunaga H, Narita N,
Tokuriki M and Saito H: Expression of p27 and apoptosis in oral
leukoplakia. Anticancer Res. 23:1265–1270. 2003.PubMed/NCBI
|
|
43
|
Taggart CC, Greene CM, McElvaney NG and
O'Neill S: Secretory leucoprotease inhibitor prevents
lipopolysaccharide-induced IkappaBalpha degradation without
affecting phosphorylation or ubiquitination. J Biol Chem.
277:33648–33653. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bindhu OS, Ramadas K, Sebastian P and
Pillai MR: High expression levels of nuclear factor kappa B and
gelatinase in the tumorigenesis of oral squamous cell carcinoma.
Head Neck. 28:916–925. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Pontes HA, Pontes FS, Fonseca FP, de
Carvalho PL, Pereira EM, de Abreu MC, de Freitas Silva BS and dos
Santos Pinto D Jr: Nuclear factor κB and cyclooxygenase-2
immunoexpression in oral dysplasia and oral squamous cell
carcinoma. Ann Diagn Pathol. 17:45–50. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Irmler M, Steiner V, Ruegg C, Wajant H and
Tschopp J: Caspase-induced inactivation of the anti-apoptotic TRAF1
during Fas ligand-mediated apoptosis. Febs Lett. 468:129–133. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lee SY and Choi Y: TRAF1 and its
biological functions. Adv Exp Med Biol. 597:25–31. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wang CY, Mayo MW, Korneluk RG, Goeddel DV
and Baldwin AS Jr: NF-kappaB antiapoptosis: Induction of TRAF1 and
TRAF2 and c-IAP1 and c-IAP2 to suppress caspase-8 activation.
Science. 281:1680–1683. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Speiser DE, Lee SY, Wong B, Arron J,
Santana A, Kong YY, Ohashi PS and Choi Y: A regulatory role for
TRAF1 in antigen-induced apoptosis of T cells. J Exp Med.
185:1777–1783. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wan XK, Yuan SL, Tao HX, Diao LP, Wang YC,
Cao C and Liu CJ: The upregulation of TRAF1 induced by helicobacter
pylori plays an antiapoptotic effect on the infected cells.
Helicobacter. 21:554–564. 2016. View Article : Google Scholar : PubMed/NCBI
|