Introduction
Radiotherapy is an important curative treatment
option for patients with metastatic tumors such as prostate cancer
(PCa) (1,2). High tumor recurrence and therapy
resistance rate in patients with PCa (>30%) suggest that
cellular radioresistance is a major obstacle and challenge to
effective radiotherapy for PCa (3,4).
Improved radiotherapy modalities and treatments for PCa are
warranted and being assessed in pre-clinical applications.
Radiotherapy usually induces severe damage in
genomic DNA, including DNA single-strand breaks (SSBs) and
double-strand breaks (DSBs) (5). In
higher eukaryotes, DNA-DSBs are repaired by two major DNA repair
pathways, the homologous recombination (HRR) and the non-homologous
end joining (NHEJ) pathways (9).
Cell cycle arrest, apoptosis and autophagy are typical impairments
induced by radiotherapy (6–8). In response to DNA damage, cell cycles
are mainly interfered by genotoxic stress at two checkpoints: G1/S
and G2/M transitions (9). DNA
damage could be monitored by characterizations of inhibited DNA
replication at G1/S checkpoint and damaged chromosome segregation
at G2/M checkpoint (9,10). Irradiation-mediated genotoxic stress
is caused by sustained DNA damage that overloads DNA repair
capacity by the HRR and NHEJ pathways (5). This DNA repair failure finally results
in increased cell apoptosis, cell cycle arrest and autophagy, thus,
inducing radiosensitivity.
Radiosensitivity of tumor cells could be achieved by
targeting signalings, gene transcriptions and non-coding RNAs
(microRNAs and lncRNAs) which ultimately regulate cell cycle
progression, apoptosis and induce cell cycle checkpoint defects
(4,6,11–14).
Two important tyrosine kinase receptors, insulin-like growth factor
1 receptor (IGF1R) and epidermal growth factor receptor (EGFR),
play essential roles in cancer development and cell cycle
progression, invasion and radiosensitivity via the activation of
downstream pathways (5,15). IGF1R gene silencing was found to
enhance the sensitivity of human PCa cells to DNA-damaging agents
(16).
P13K/Akt/mTOR signaling is responsive to various
stresses and plays a critical role in cell cycle, apoptosis,
autophagy, invasion, metastasis, tumorigenesis and radiosensitivity
of tumor cells (5,17–19).
Genistein is a tyrosine-specific protein kinases inhibitor,
topoisomerase II poison, which inactivates EGFR, IGF1R and
Akt-mediated signalling (20–23).
It was reported that genistein inhibited DSB repair through the
inactivation of DNA-dependent protein kinases (DNA-PKs) and led to
NHEJ and HRR incompleteness (24).
Tyrosine kinase inhibitor (tyrphostin) AG1024 is a specific IGF1R
inhibitor, which has been reported to radiosensitize PCa cells and
breast cancer cells (5,18). However, less information is known
about the radiosensitivity of PCa cells to the combination
treatment with AG1024 and genistein.
In the present study, we investigated the
synergistic effect of combination treatment with AG1024 and
genistein on the radiosensitivity of PCa cells. Before
X-irradiation, PC3 and DU145 PCa cells were treated with genistein,
AG1024 and their combination. The radiosensitivity of PCa cells was
evaluated by changes in DNA damage, cell proliferation, apoptosis,
cell cycle distribution and the inactivation of the NHEJ and HRR
pathways. The synergistic effect of AG1024 and genistein on the
sensitivity of PCa cells to radiotherapy is discussed. This study
provides new insights into the compensation/synergic effect on NHEJ
and HRR pathways for DNA damage repair by AG1024 and genistein.
Materials and methods
Cell lines, culture conditions and
treatments
The human PCa cell lines PC3 and DU145 were
purchased from the American Type Culture Collection (ATCC,
Manassas, VA, USA). Cells were maintained in RPMI-1640 medium
(Gibco-BRL; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS) and 1%
penicillin/streptomycin (Gibco-BRL; Thermo Fisher Scientific) at
37°C under 5% CO2 in a humidified incubator. For cell
treatment, PC3 and DU145 cells were cultured in RPMI-1640 medium
supplemented with genistein (purity >98%, Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) and AG1024 (Sigma-Aldrich; Merck KGaA) at
series concentrations. Genistein (0, 10, 20, 30, 50 and 100 µM) and
AG1024 (10 µM) (5) were dissolved
in dimethyl sulfoxide (DMSO; Sigma-Aldrich; Merck KGaA). X-ray
irradiation was delivered using an X-6 MV photon linear accelerator
(Varian Associates Inc., Palo Alto, CA, USA) at room temperature
with a dose rate of 2 Gy/min (25).
Cells treated with DMSO were considered as negative control for
irradiation or drug treatment.
Cell viability and in vitro
cytotoxicity assay
The optimal concentration of genistein and optimal
dose of X-ray irradiation were selected using a Cell Counting Kit-8
(CCK-8) assay kit (Sigma-Aldrich; Merck KGaA). The cytotoxicity of
genistein and X-irradiation to human PCa cells was assessed using
cell viability analysis. Cells were placed into 96-well plates,
pretreated with DMSO, genistein, AG1024 and combination of
genistein and AG1024 for 1 h, followed by X-irradiation. Then,
cells were incubated in RPMI-1640 medium for another 24 h in
conditions as above. Ten microliters of CCK-8 solution per well was
added into the cell culture for 2 h, and then the cell culture was
subjected to a microplate spectrophotometer (Bio-Rad Laboratories,
Hercules, CA, USA) for determination of the optical density
(OD450nm).
Soft agar colony formation
analysis
Monolayer clonogenic assay was performed to
investigate the effect of genistein, AG1024 and the combination on
colony formation ability. Cells were placed into 6-well plates and
grown to log phase, followed by transferring to medium containing
DMSO, genistein, AG1024 and genistein plus AG1024. After a 1-h
incubation, the cells were irradiated with 4 Gy of X-irradiation
and incubated in RPMI-1640 medium for 24 h. Then, the cells were
treated with trypsin (Gibco-BRL; Thermo Fisher Scientific, Inc.),
transferred to a 6-cm diameter tissue culture dish with fresh
RPMI-1640 medium and incubated for 12 days. Medium was replaced
every third day. For colony detection, dishes were washed with
phosphate-buffered saline (PBS) for two times, fixed with methanol
at 4°C for 15 min, and stained with Giemsa (Sigma-Aldrich; Merck
KGaA) for 30 min. Colony counting was performed on clearly visible
colonies (colony with >50 cells or diameter >50 µm) using a
light microscope (BX51; Olympus Optical Co., Ltd., Tokyo, Japan).
Each experiment was performed in triplicate.
Flow cytometric analysis
The effects of the combination treatment of
genistein and AG1024 and single treatments on cell cycle
distribution and apoptosis were detected using a flow cytometer (BD
Biosciences, San Jose, CA, USA). For cell cycle analysis, cells
under different treatments for 24 h were harvested (trypsin;
Gibco-BRL; Thermo Fisher Scientific, Inc.), fixed (70% ethanol),
stained with 50 µg/ml propidium iodide (PI) solution, and were
subjected to flow cytometry. Cell cycle distribution at G0/G1, S
and G2/M phase was analyzed. For cell apoptosis analysis, fixed
cells were incubated with PI and Annexin V-FITC for 10 min in the
dark, and then subjected to a BD FACSCalibur flow cytometer (BD
Biosciences). Cells undergoing early apoptosis were considered as
apoptotic cells (Annexin V-positive and PI-negative, Annexin
V+/PI−). Each experiment was conducted in
triplicate.
Immunofluorescence
Immunofluorescence analysis was performed to detect
the γH2AX foci formation as described by Wang et al
(5). In brief, 1×106
cells/ml of PC3 and DU145 cells were seeded into 6-well plates with
coverslips and were treated with different treatments combined with
X-irradiation for 24 h. The cells were then fixed with 4%
paraformaldehyde for 20 min, incubated with 0.2% Triton X-100 in
PBS for 5 min, and coverslips were blocked with 5% bovine serum
albumin (BSA; Gibco-BRL; Thermo Fisher Scientific, Inc.) for 30 min
at room temperature. Then slips with fixed cells were incubated
with specific primary antibody against phospho-histone γH2AX
(1:500; cat. no. 2595; Cell Signaling Technology, Inc., Danvers,
MA, USA) at 4°C overnight, followed by incubation with Cy3-labelled
goat anti-rabbit fluorescent secondary antibody (1:2,000; cat. no.
111-165-003; Jackson ImmunoResearch Laboratories, West Grove, PA,
USA) for 1 h at room temperature and 1 µg/ml DAPI (Invitrogen;
Thermo Fisher Scientific, Inc.) for additional 10 min in the dark.
Images were captured using an Olympus laser scanning confocal
microscopy (LEXT 3100; Olympus Corp., Tokyo, Japan).
Western blot analysis
Cells were placed into 6-well plates and incubated
using the different treatments as above. Cells were harvested at 24
h post X-irradiation. Cellular and nuclear protein was isolated
using RIPA buffer (Pierce Inc., Beijing, China). Proteins were
prepared as described by Liu et al (26). Western blot analysis was performed
according to the standard methods. Specific primary antibodies of
anti-phospho (p)-IGF1R (Tyr1135), -IGF1R, -ATM, -ATM(Ser1981),
-Bax, -Bcl2, -cleaved caspase-3, -Ku70, -Rad51, -DNA-PKcs and
-GAPDH were purchased from Cell Signaling Technology, Inc. Primary
antibody p-DNA-PKcs (Thr2609) was purchased from Santa Cruz
Biotechnology Inc., (Santa Cruz, CA, USA; cat. no. sc-101664).
In vivo tumor radiation protocol
The in vivo subcutaneous mouse tumors were
produced by subcutaneously injecting 5×106 DU145 cells,
mixed with BD Matrigel (BD Biosciences), into the flank of male
nude mice (6–7 weeks old, 18–20 g, n=60) provided by the
Experimental Animal Center of the Fourth Military Medical
University (5). Animals were
maintained with ad libitum access to food and water for 5
days at 25±1°C in environmental chambers, with 40–50% humidity and
12 h light: 12 h dark cycle. A digital Vernier caliper was used for
measuring tumor volume [V = 0.5 × tumor length (mm) × tumor
width2 (mm2)]. Twenty days later, mice were
randomly divided into four groups (n=15 in each group): the DMSO +
IR (control) group received X-irradiation every three days for 5
times (15-day treatment course), with orally intubated with 200
mg/kg/day DMSO; the genistein + IR group received 100 mg/kg/day
genistein, 100 mg/kg/day DMSO and X-irradiation for 5 times; the
AG1024 + IR group received 100 mg/kg/day AG1024, 100 mg/kg/day DMSO
and X-irradiation for 5 times; the Combination (genistein + AG1024)
+ IR group received 100 mg/kg/day genistein, 100 mg/kg/day AG1024,
plus with X-irradiation for 5 times. The in vivo therapeutic
efficacy of the different treatments on in vivo tumors was
evaluated using changes in tumor volume and proliferation index
(PI, PI=Vtreatment/Vcontrol) (5). Body weight (g) of experimental animals
were recorded. Multiple nodes in one mouse were circled into one
circle and the accumulated volume was calculated as above. All mice
were sacrificed by anesthesia and the tumors were removed on day 15
after the 1st administration of genistein, AG1024 and the
combination treatment. The animal experiment protocols were
approved by the Ethics Committee of the Fourth Military Medical
University (Xi'an, China).
Statistical analyses
Each cellular experiment was performed in
triplicate. All quantitative data and continuous variables are
expressed as mean ± standard deviation (SD). Statistical analysis
was performed using the unpaired two-tailed Student's t-test
in SPSS software 17.0 (SPSS, Inc., Chicago, IL, USA). GraphPad
Prism 6 (Graphpad Software, Inc., San Diego, CA, USA) was used for
plotting. Statistical significant P-value is given as P≤0.05.
Results
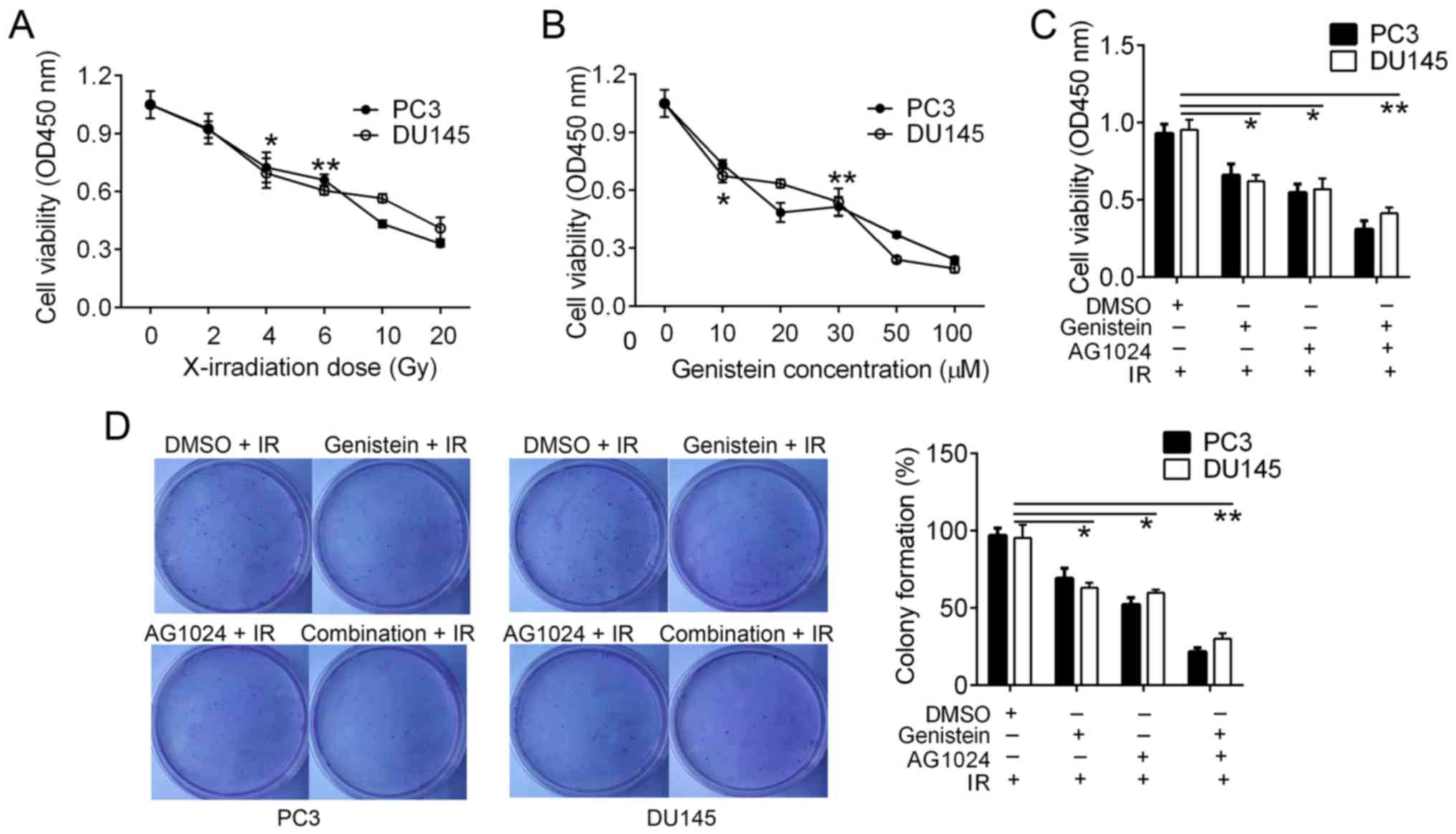
Cytotoxicity of irradiation and
genistein in PCa cells
The frequently used concentration of AG1024 is 10 µM
for cancer cells (5,18). In order to select suitable doses of
X-irradiation and genistein, we treated PC3 and DU145 cells with
different doses of X-irradiation (0, 2, 4, 6, 10 and 20 Gy) and
genistein (0, 10, 20, 30, 50 and 100 µM). Cell cytotoxicity was
detected using the CCK-8 assay. In comparison analysis, we found
that cell viability was significantly inhibited by ≥4-Gy
X-irradiation (P<0.05; Fig. 1A)
and ≥10 µM genistein (P<0.05; Fig.
1B). Thus, we selected 4-Gy X-irradiation and 30 µM genistein
as the optimal conditions for cell cytotoxicity in the PCa
cells.
Combination of genistein and AG1024
enhances X-irradiation-reduced PC3 and DU145 cell
proliferation
We pretreated PC3 and DU145 cells with genistein (30
µM), AG1024 (10 µM) and the combination before X-irradiation (4
Gy). Twenty-four hours later, we confirmed that the combination of
genistein and AG1024 significantly decreased cell proliferation
(P<0.01), followed by genistein (30 µM, P<0.05) and AG1024
(10 µM, P<0.05) as compared with that of the control cells
(Fig. 1C). Using colony formation
assay, we detected that the colony numbers of PC3 and DU145 cells
were reduced by treatments of genistein (30 µM), AG1024 (10 µM) and
the combination treatment (P<0.05; Fig. 1D). PC3 and DU145 cells pretreated
with the combination of genistein (30 µM) and AG1024 (10 µM) before
X-irradiation (4 Gy) showed the lowest frequency of colony
formation (P<0.01), followed by cells pretreated with either
genistein (30 µM) or AG1024 (10 µM) before X-irradiation (4 Gy,
P<0.05; Fig. 1D). These results
showed that the combination treatment of genistein and AG1024 and
single treatments enhanced PCa cell cytotoxicity by X-irradiation.
In addition, the combination of genistein and AG1024 indicated
higher efficacy in the inhibition of cell proliferation than either
genistein or AG1024 single treatment, suggesting a synergistic
inhibition effect on PCa cell proliferation.
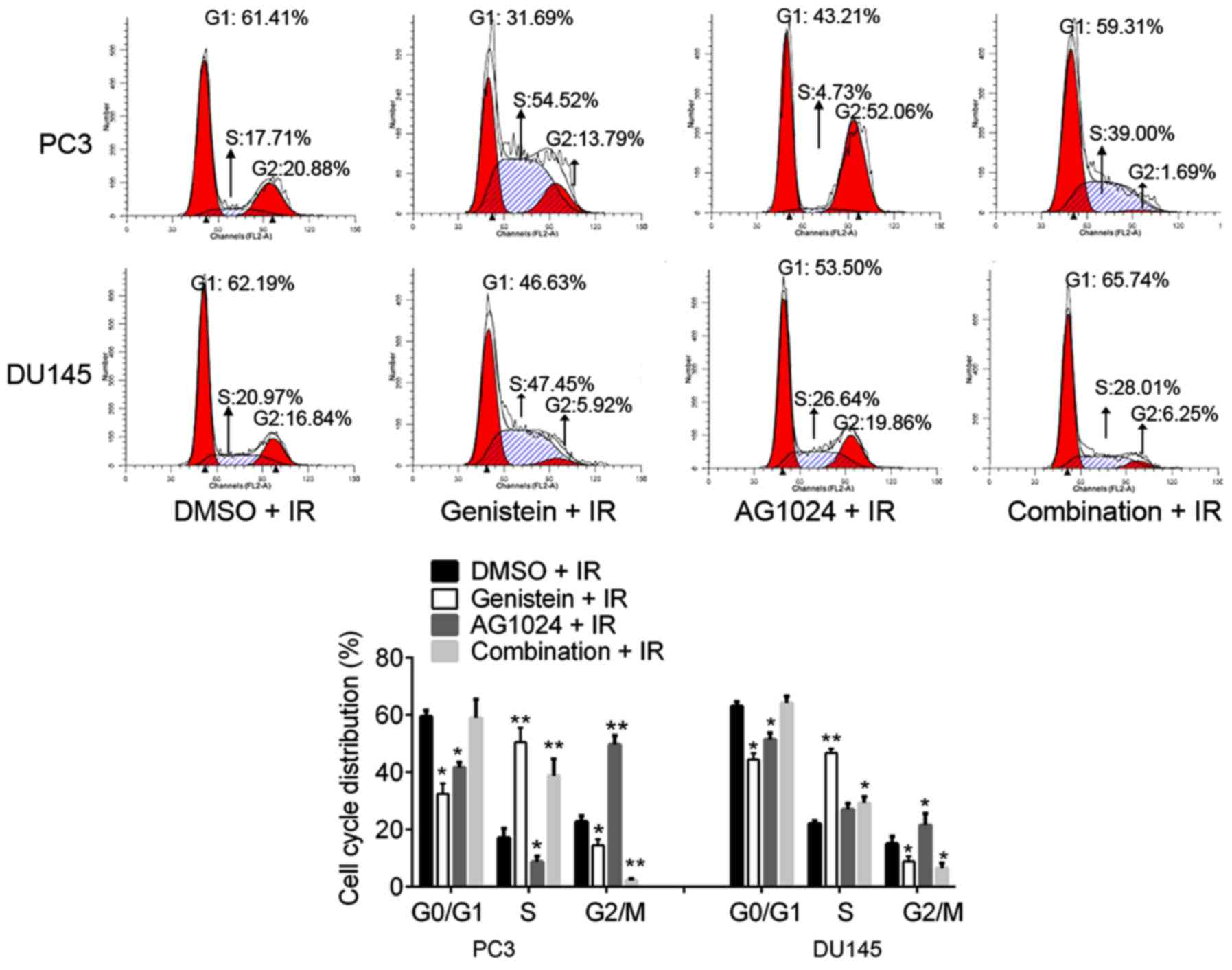
Combination treatment of genistein and
AG1024 and single treatments induce cell cycle arrest
Cell cycle distribution analysis showed the
percentages of cells at G0/G1, S and G2/M phases were significantly
disturbed by genistein (30 µM), AG1024 (10 µM) and the combination
treatment. As compared with the control (DMSO + irradiation),
genistein (30 µM) treatment significantly decreased the percentages
of cells at G0/G1 and G2/M phases (P<0.05), and increased the
percentages of cells at S phases (P<0.01), revealing that
genistein induced S cell cycle arrest (Fig. 2). In addition, we observed that
AG1024 treatment induced G2/M cell cycle arrest in PC3 and DU145
cells. AG1024 treatment significantly increased the percentages of
cells at the G2/M phases (P<0.01 for PC3 cells; P<0.05 for
DU145 cells) and reduced the percentages of cells at G0/G1 and/or S
phases (P<0.05).
Notably, the combination treatment with genistein
(30 µM) and AG1024 (10 µM) significantly increased the percentages
of cells at the S phase (P<0.01 for PC3 cells; P<0.05 for
DU145 cells) and decreased the percentages of cells at G2/M phases
(P<0.01 for PC3 cells; P<0.05 for DU145 cells). The
percentages of cells at G0/G1 phases were concomitant with those of
the control cells (DMSO + IR; P>0.05). Taken together, we found
that genistein (30 µM) induced S cell cycle arrest, AG1024 (10 µM)
induced G2/M cell cycle arrest, and the combination of genistein
(30 µM) and AG1024 (10 µM) induced S cell cycle arrest. In
addition, the combination treatment with genistein and AG1024
showed a synergistic inhibitory effect on chromosome segregation at
the G2/M phase, but not DNA replication at the G0/G1 phase in PCa
cells.
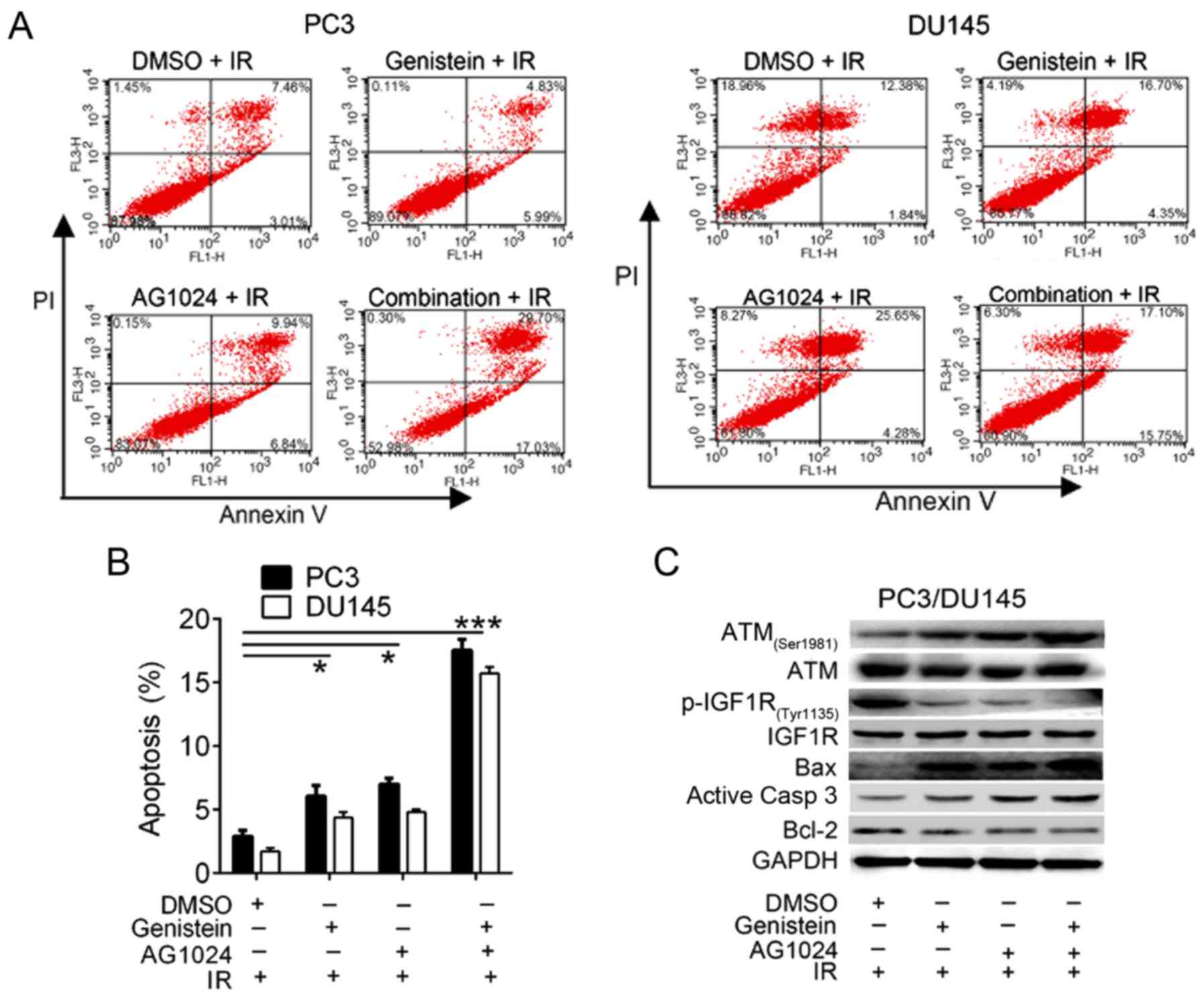
Combination treatment with genistein
and AG1024 enhances X-irradiation-induced apoptosis in PC3 and
DU145 cells
We then confirmed that the treatment of genistein
(30 µM), AG1024 (10 µM) and their combination promote cell
apoptosis using flow cytometric analysis. Significant enhancement
of cell apoptosis was observed in cells treated with genistein (30
µM, P<0.05), AG1024 (10 µM, P<0.05), and both of them
(P<0.01) combined with X-irradiation (Fig. 3A and B). These data demonstrated
that genistein (30 µM) and/or AG1024 (10 µM) treatment enhanced
X-irradiation-induced PCa cell apoptosis, and the combination
treatment with genistein and AG1024 synergistically promoted cell
radiosensitivity.
Using western blot analysis, we detected the
expression of cell apoptosis-related proteins and pathway. In
comparison with the control cells (DMSO + irradiation), the
expression of ATM (Ser1981), Bax and active caspase-3 were
increased, whereas the expression of p-IGF1R (Tyr1135) and Bcl-2
were decreased in PC3 and DU145 cells treated with genistein (30
µM) and/or AG1024 (10 µM) plus X-irradiation (Fig. 3C). Thus, we suggested that both
genistein and AG1024 induced PCa cell apoptosis via the activation
of apoptosis-related pathways, which may be associated with the
inactivation of IGF1R. In addition, combination of genistein and
AG1024 synergistically modulated the expression of these proteins,
showing synergistic effect on irradiation-induced cell apoptosis of
PCa cells.
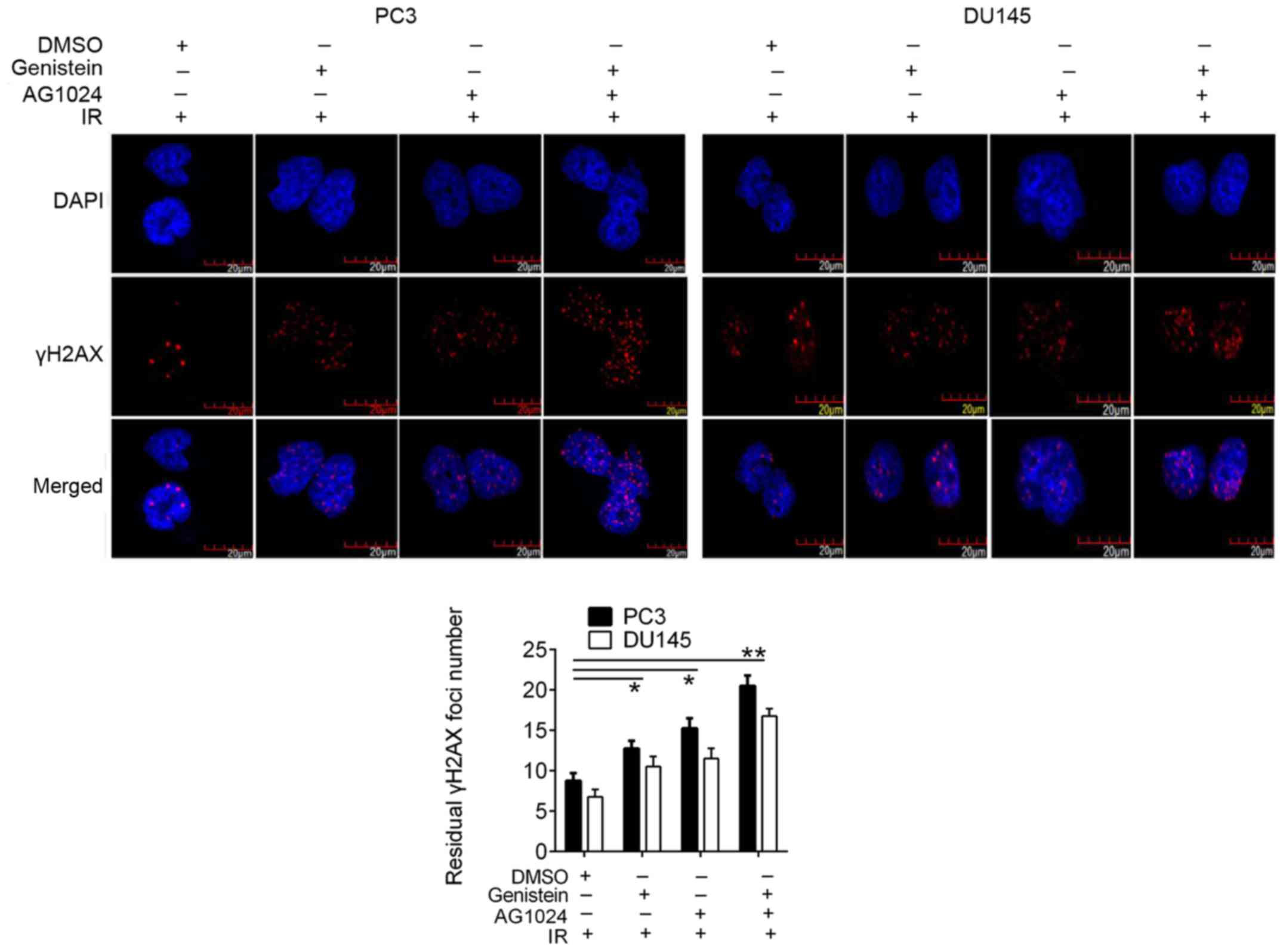
Genistein and/or AG1024 enhances
irradiation-induced DSB in PCa cells
We then revealed the increased γH2AX foci formation,
a DSB marker, in genistein (30 µM) and/or AG1024 (10 µM) treated
PCa cells (Fig. 4), suggesting the
elevation of DNA damage by pretreatment with genistein, AG1024, and
combination before X-irradiation. In PC3 and DU145 cells pretreated
with different treatments before X-irradiation, the numbers of
residual γH2AX foci were significantly elevated as compared with
that in the control cells (P<0.05). Moreover, PCa cells treated
with the combined treatment with genistein and AG1024 demonstrated
a higher number of γH2AX foci than cells treated with single
treatments of either genistein or AG1024, showing synergistic
enhancement of γH2AX foci formation by genistein and AG1024.
Combination of genistein and AG1024
enhances radiosensitivity of PCa cells via the inactivation of HRR
and NHEJ pathways
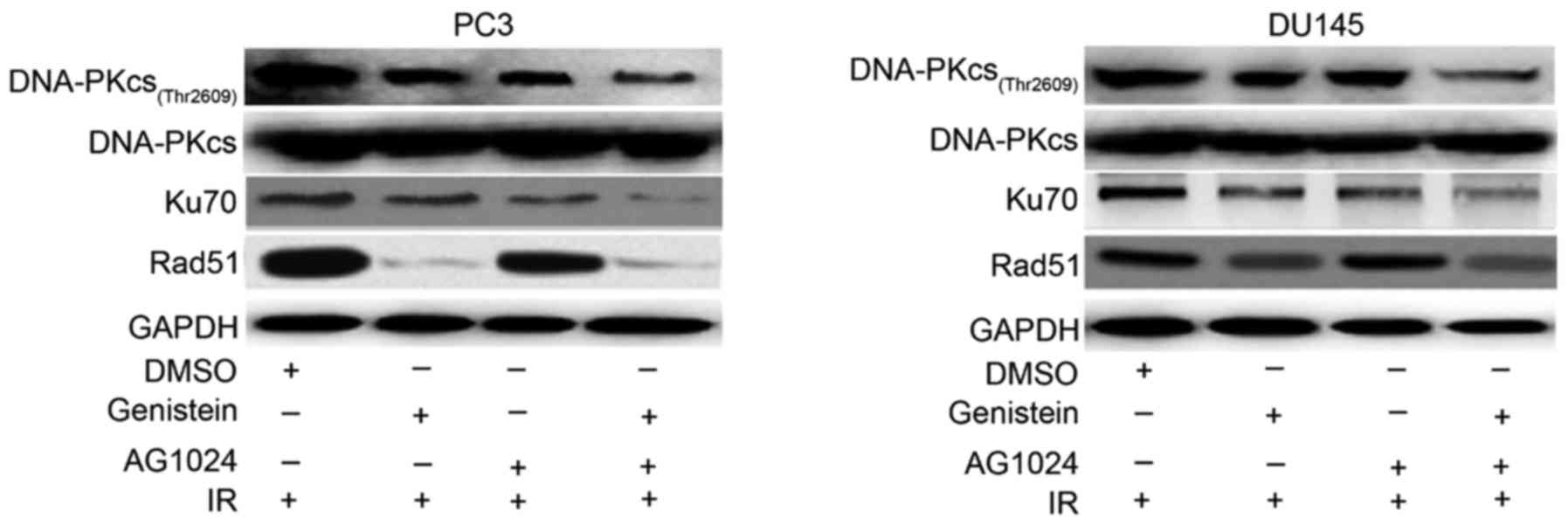
Using western blot analysis, we detected the
decreased expression of NHEJ-related DNA repair proteins DNA-PKcs
(Thr2609) and Ku70 in PCa cells pretreated with genistein (30 µM),
AG1024 (10 µM) and their combination as compared with the control
(Fig. 5). PCa cells pretreated with
the combination of genistein and AG1024 before X-irradiation showed
the lowest expression levels of nuclear DNA-PKcs and Ku70, followed
by PCa cells pretreated with either genistein (30 µM) or AG1024 (10
µM). However, the expression of nuclear Rad51, an enzymatic
component of HRR, was decreased by genistein alone and in
combination with AG1024, but not AG1024 alone. These results
revealed that genistein and the combination of genistein and AG1024
enhanced the radiosensitivity of PCa cells via the inhibition of
both HRR and NHEJ pathways, while AG1024 promoted the
radiosensitivity of PCa cells only by inactivating the NHEJ
pathway.
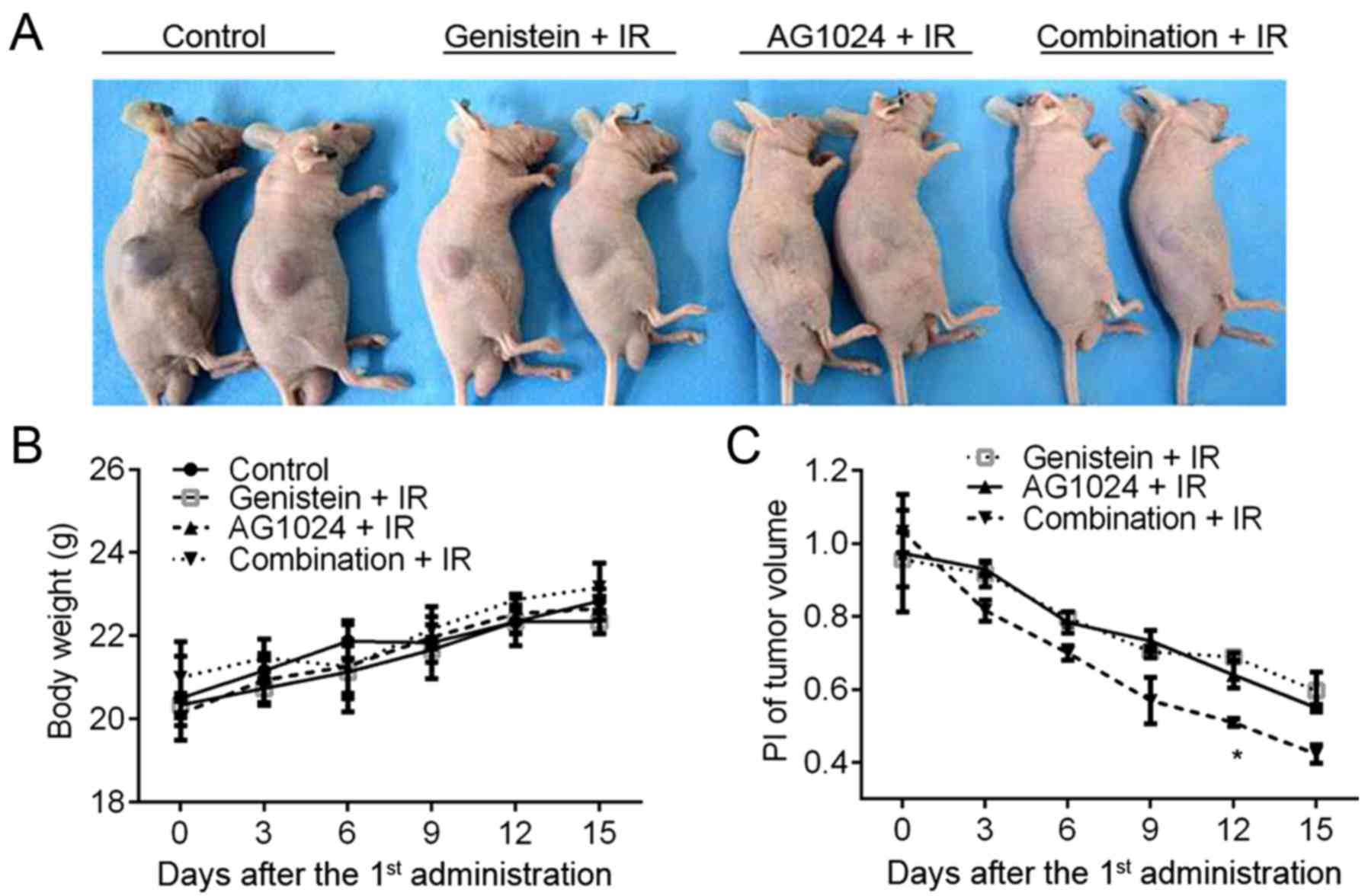
Genistein and/or AG1024 enhance cancer
radiotherapy in a preclinical model of PCa tumors
The in vitro cellular experiments
demonstrated the single treatment with either genistein (30 µM) or
AG1024 (10 µM) and the combination treatment radio-sensitized PCa
cells to X-irradiation. To investigate the in vivo effect of
genistein and AG1024 on cancer radiotherapy, we constructed the
in vivo subcutaneous tumor model using DU145 cells. Before
X-irradiation, mice were orally intubated with DMSO, genistein (100
mg/kg/day), AG1024 (100 mg/kg/day), and the combination of
genistein and AG1024. The body weight of the mouse model increased
from 18–20 g at the onset (6–7 weeks of age) to 20–24 g at the end
of this study (11–12 weeks of age). At the day 15 post the 1st
administration of genistein and/or AG1024, the tumor volume ranged
from 1,697.20 mm3 in control mice to 252.01
mm3 in mice treated with the combination of genistein
and AG1024 (Fig. 6A). No difference
was observed in body weight between the groups (Fig. 6B). As compared with the control mice
(DMSO + IR), significantly decreased PI of tumor volume was
observed in mice treated with genistein, AG1024, and the
combination (P<0.05; Fig. 6C).
Moreover, at day 12 post the 1st administration of the agent, the
PI of the tumor volume in mice treated with the combination of
genistein and AG1024 was significantly lower than those in the mice
treated with either genistein or AG1024 (P<0.05). The in
vivo experiments showed that the treatment with either
genistein or AG1024 radiosensitized PCa tumors, and the combination
of them showed a synergistic effect on inhibiting tumor growth and
sensitizing PCa to radiotherapy in vivo.
Discussion
The present study investigated whether the
combination of genistein and AG1024 could enhance the
radiosensitivity of PCa cells to X-irradiation. The data confirmed
that the single and combination of genistein and AG1024
radiosensitized PCa cells to X-irradiation by arresting the cell
cycle, promoting cell apoptosis, and inhibiting DNA repair via the
inactivation of the NHEJ pathway and/or HRR pathway.
Radiotherapy to cancer cells induces cell damage by
causing genomic DNA damage overloading DNA repair capacity, which
finally results into mitotic catastrophe in tumor cells (5). Cell cycle arrest, apoptosis and
autophagy are typical impairments induced by radiotherapy (6–8). Both
genistein and AG1024 regulate DNA damage and enhance the
radiosensitivity of tumor cells (5,17,28).
Genistein has been proven to induce DNA damage, apoptosis, and cell
cycle arrest at the G2/M phase via the ATM-p53-dependent pathway
(Fig. 7), p38 MAPK activation,
NF-κB and other pathways (12,27–29).
In addition, genistein inhibited IGF-1 signaling pathway and
inactivated Akt signaling pathways (30). In addition, genistein has been
reported to arrest cell cycle at the G0/G1 phase in MCF-7, HB4a and
BG-1 cells (31,32). Tyrphostin AG1024 is an inhibitor of
IGF1R. The inhibition of IGF1R enhances tumor cell radiosensitivity
(13). In this study, we confirmed
that the pretreatment of genistein and AG1024 before X-irradiation
promoted DNA damage and cell apoptosis in PC3 and DU145 cells,
thus, enhancing the radiosensitivity of PCa cells to
X-irradiation.
The cell cycle is regulated by check point proteins,
including G0/G1 phase checkpoints, S phase checkpoints and G2/M
phase checkpoints. These checkpoints are responsible for the
genomic instability of tumor cells and are responsive to
irradiation-induced DNA damage (14,33).
These checkpoints control cell cycle arrest and the
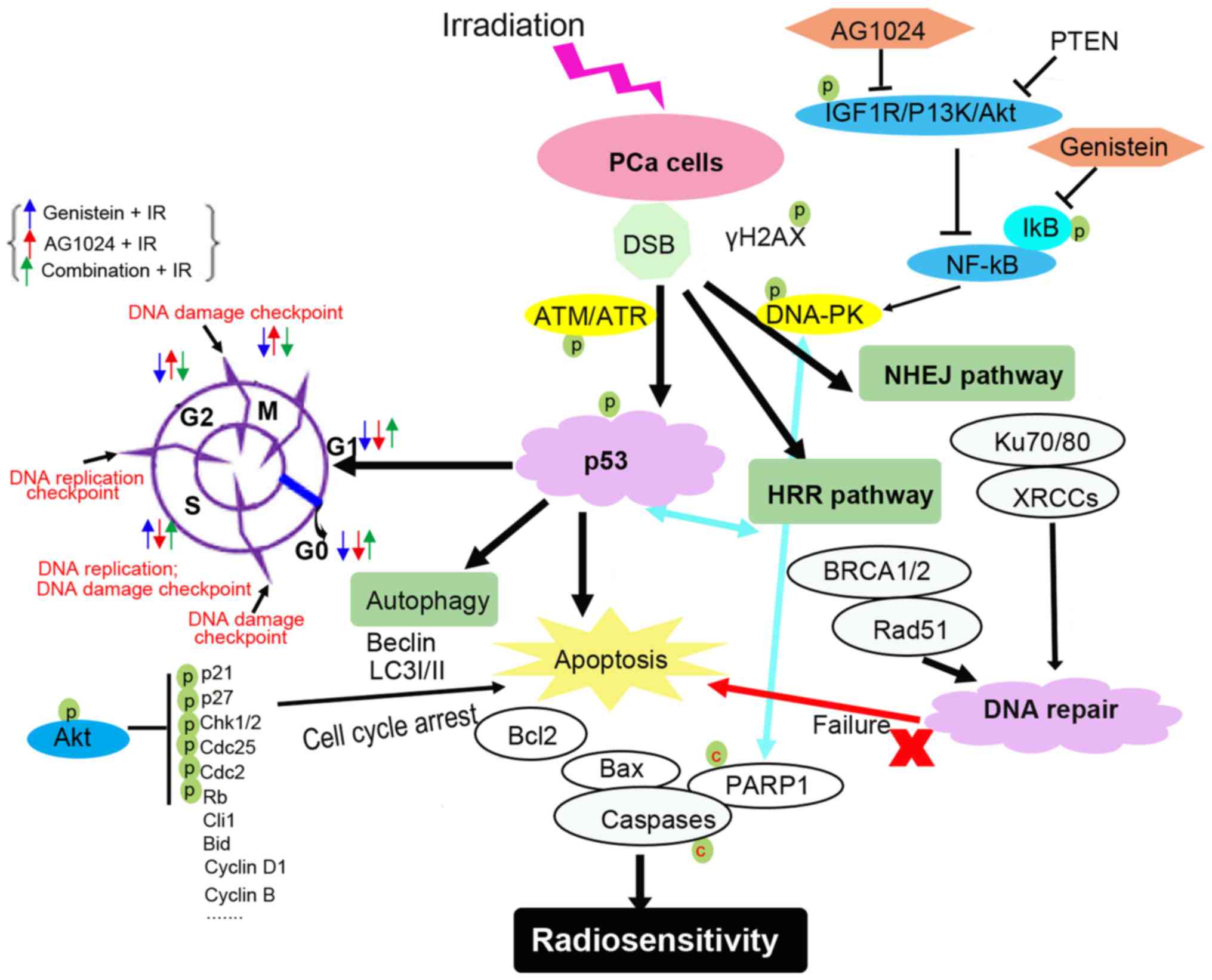
radiosensitivity of tumor cells (Fig.
7). Under irradiation-mediated genotoxic stress, increased
DNA-DSB challenges the stability of genomic DNA and the DNA repair
capacity determines the cell survival and the radiosensitivity of
cancer cells (5). Under
irradiation, DNA damage is sensed by DNA-PK/ATM and ATM-related
kinase (ATR)-mediated signaling. The activation of this signaling
promotes p53 phosphorylation and activates p53-mediated responses
(34,35). The DNA damage checkpoints, including
G2/M phase, S phase and G0-S transition checkpoints, are sensitive
to and responsible for irradiation-mediated DNA damage (36). The p53-dependent p21 phosphorylation
promoted cell cycle arrest at the G1 phase by inhibiting cyclin D1,
cyclin E and CDK2 (32,37), while G2/M cell cycle arrest was
induced by decreased cyclin B and Chk2/cell division cycle 25C
(Cdc25C)/Cdc2, which was negatively regulated by p53-dependent
GADD45 expression (8,9,27,8).
Genistein and AG1024-induced G2/M cell cycle arrest has been
reported in human colon cancer cells and breast cancer cells
(28,29,40),
whereas genistein and AG1024-induced G1 cell cycle arrest has also
been confirmed in BG-1 ovarian cancer and DU145 PCa cells (18,32).
These results demonstrated that genistein or AG1024-mediated cell
arrest is dependent on cell types or other unknown conditions. Our
current data demonstrated that genistein induced PCa cell cycle
arrest at the S phase, and AG1024 induced PCa cell cycle arrest at
the G2/M phase. In addition, the combination of genistein and
AG1024 arrested the cell cycle at the S phase, showing a
synergistic effect on cell cycle distribution with a genistein
tendency.
In tumor cells, the NHEJ and HRR pathways enhance
DNA repair capacity and modulate cell sensitivity and resistance to
radiotherapy (5). The inhibition of
IGF1R enhances radiosensitivity and delays DSB repair by
inactivating NHEJ and HRR pathways (13). Previous reports have shown that the
expression of the Ku70/80 gene and DNA-PKcs activity did not
correlate with DSB repair capacity and cellular radiosensitivity in
normal human fibroblasts (41). In
tumor cells, the NHEJ pathway is mediated by DNA-PK-dependent
phosphorylation of Ku70/80 and the HRR pathway is mediated by
BRCA1/Rad51 expression (4,41). In comparison with normal tumor
cells, increased NHEJ and HRR pathways were detected in
radio-resistant PCa cells (4). On
contrast, Chang et al indicated that the combination of
P13K/mTOR inhibitor and 6-Gy radiation inhibited both NHEJ and HRR
pathways, thus, radiosensitizing PCa cells (4). It was reported that genistein
radiosensitized breast cancer cells via inactivating the HRR repair
pathway by inhibiting Rad51 expression (28). IGF1R inhibitor AZ12253801
radiosensitized PCa cells through inactivating NHEJ repair pathway
(13). Our data showed that
genistein radiosensitized PCa cells via inhibiting both NHEJ and
HRR pathways, and AG1024 enhanced the radiosensitivity of PCa cells
to X-irradiation via the inhibition of the NHEJ pathway.
Additionally, the combination of AG1024 and genistein showed a
synergistic effect on the inhibition of NHEJ and HRR pathways and
the radiosensitivity of PCa cells to X-irradiation, showing the
therapeutic potential of the combination treatment with AG1024 and
genistein for PCa.
In summary, we confirmed that both AG1024 and
genistein radiosensitized PCa cells by arresting the cell cycle at
the G2/M and S phase, respectively. In addition, the
genistein-induced radiosensitivity of PCa cells was mediated by the
inhibition of both HRR and NHEJ repair pathways, while AG1024
induced radiosensitivity of PCa cells only through the inhibition
of the NHEJ pathway. Both in vitro and in vivo
experiments indicated that the combination of AG1024 and genistein
synergistically enhanced the radiosensitivity of PCa cells. Even
so, we are still not entirely clear concerning the crosstalk
between AG1024 and genistein. The mechanisms and differences in
cell cycle arrest and DNA repair pathways between AG1024 and
genistein were not explored in the present study. We hypothesize
that the investigation of cyclin expression may be helpful for
uncovering the differences in genistein and AG1024-mediated cell
cycle arrest and the radiosensitivity of PCa cells. More
experiments should be performed before the clinical therapy of PCa
by using the combination of AG1024 and genistein.
Acknowledgements
Not applicable.
Funding
The present study was supported by the grants from
the National Natural Science Foundation of China (grant no.
81672535) and the National Natural Science Foundation of Shaanxi
Province (grant no. 2016JM8145).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
QT and JM conceived and designed the study. QT, JM,
JS, LY, FY, WZ, RL and LW performed the experiments. QT and JM
analysed the data. QT and JM wrote the manuscript. YW and HW
reviewed and checked the manuscript. All authors read and approved
the manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
The animal experiment protocols were approved by the
Ethics Committee of the Fourth Military Medical University (Xi'an,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cojoc M, Peitzsch C, Kurth I, Trautmann F,
Kunz-Schughart LA, Telegeev GD, Stakhovsky EA, Walker JR, Simin K,
Lyle S, et al: Aldehyde dehydrogenase is regulated by β-catenin/TCF
and promotes radioresistance in prostate cancer progenitor cells.
Cancer Res. 75:1482–1494. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rycaj K and Tang DG: Cancer stem cells and
radioresistance. Int J Radiat Biol. 90:615–621. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shimura T, Noma N, Sano Y, Ochiai Y,
Oikawa T, Fukumoto M and Kunugita N: AKT-mediated enhanced aerobic
glycolysis causes acquired radioresistance by human tumor cells.
Radiother Oncol. 112:302–307. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chang L, Graham PH, Hao J, Ni J, Bucci J,
Cozzi PJ, Kearsley JH and Li Y: PI3K/Akt/mTOR pathway inhibitors
enhance radiosensitivity in radioresistant prostate cancer cells
through inducing apoptosis, reducing autophagy, suppressing NHEJ
and HR repair pathways. Cell Death Dis. 5:e14372014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang Y, Yuan JL, Zhang YT, Ma JJ, Xu P,
Shi CH, Zhang W, Li YM, Fu Q, Zhu GF, et al: Inhibition of both
EGFR and IGF1R sensitized prostate cancer cells to radiation by
synergistic suppression of DNA homologous recombination repair.
PLoS One. 8:e687842013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kang KB, Zhu C, Yong SK, Gao Q and Wong
MC: Enhanced sensitivity of celecoxib in human glioblastoma cells:
Induction of DNA damage leading to p53-dependent G1 cell cycle
arrest and autophagy. Mol Cancer. 8:662009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ito H, Daido S, Kanzawa T, Kondo S and
Kondo Y: Radiation-induced autophagy is associated with LC3 and its
inhibition sensitizes malignant glioma cells. Int J Oncol.
26:1401–1410. 2005.PubMed/NCBI
|
|
8
|
Raffoul JJ, Wang Y, Kucuk O, Forman JD,
Sarkar FH and Hillman GG: Genistein inhibits radiation-induced
activation of NF-κB in prostate cancer cells promoting apoptosis
and G2/M cell cycle arrest. BMC Cancer. 6:1072006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang J, Wu LJ, Tashino S, Onodera S and
Ikejima T: Protein tyrosine kinase pathway-derived ROS/NO
productions contribute to G2/M cell cycle arrest in
evodiamine-treated human cervix carcinoma HeLa cells. Free Radic
Res. 44:792–802. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hartwell LH and Kastan MB: Cell cycle
control and cancer. Science. 266:1821–1828. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Su H, Jin X, Shen L, Fang Y, Fei Z, Zhang
X, Xie C and Chen X: Inhibition of cyclin D1 enhances sensitivity
to radiotherapy and reverses epithelial to mesenchymal transition
for esophageal cancer cells. Tumor Biol. 37:5355–5363. 2016.
View Article : Google Scholar
|
|
12
|
Cui S, Wienhoefer N and Bilitewski U:
Genistein induces morphology change and G2/M cell cycle arrest by
inducing p38 MAPK activation in macrophages. Int Immunopharmacol.
18:142–150. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chitnis MM, Lodhia KA, Aleksic T, Gao S,
Protheroe AS and Macaulay VM: IGF-1R inhibition enhances
radiosensitivity and delays double-strand break repair by both
non-homologous end-joining and homologous recombination. Oncogene.
33:5262–5273. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hartwell L: Defects in a cell cycle
checkpoint may be responsible for the genomic instability of cancer
cells. Cell. 71:543–546. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Stegeman H, Kaanders JH, van der Kogel AJ,
Iida M, Wheeler DL, Span PN and Bussink J: Predictive value of
hypoxia, proliferation and tyrosine kinase receptors for
EGFR-inhibition and radiotherapy sensitivity in head and neck
cancer models. Radiother Oncol. 106:383–389. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rochester MA, Riedemann J, Hellawell GO,
Brewster SF and Macaulay VM: Silencing of the IGF1R gene
enhances sensitivity to DNA-damaging agents in both PTEN wild-type
and mutant human prostate cancer. Cancer Gene Ther. 12:90–100.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wen B, Deutsch E, Marangoni E, Frascona V,
Maggiorella L, Abdulkarim B, Chavaudra N and Bourhis J: Tyrphostin
AG 1024 modulates radiosensitivity in human breast cancer cells. Br
J Cancer. 85:2017–2021. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kisielewska J, Ligeza J and Klein A: The
effect of tyrosine kinase inhibitors, tyrphostins: AG1024 and
SU1498, on autocrine growth of prostate cancer cells (DU145). Folia
Histochem Cytobiol. 46:185–191. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shukla V, Chandra V, Sankhwar P, Popli P,
Kaushal JB, Sirohi VK and Dwivedi A: Phytoestrogen genistein
inhibits EGFR/PI3K/NF-κB activation and induces apoptosis in human
endometrial hyperplasial cells. RSC Advances. 5:56075–56085. 2015.
View Article : Google Scholar
|
|
20
|
Hari S, Vasudevan V, Kasibhotla S, Reddy
D, Venkatappa M and Devaiah D: Anti-inflammatory dietary
supplements in the chemoprevention of oral cancer. Cancer Res
Front. 2:380–395. 2016. View Article : Google Scholar
|
|
21
|
Akiyama T, Ishida J, Nakagawa S, Ogawara
H, Watanabe S, Itoh N, Shibuya M and Fukami Y: Genistein, a
specific inhibitor of tyrosine-specific protein kinases. J Biol
Chem. 262:5592–5595. 1987.PubMed/NCBI
|
|
22
|
Akiyama T and Ogawara H: Use and
specificity of genistein as inhibitor of protein-tyrosine kinases.
Methods Enzymol. 201:362–370. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Markovits J, Linassier C, Fossé P, Couprie
J, Pierre J, Jacquemin-Sablon A, Saucier JM, Le Pecq JB and Larsen
AK: Inhibitory effects of the tyrosine kinase inhibitor genistein
on mammalian DNA topoisomerase II. Cancer Res. 49:5111–5117.
1989.PubMed/NCBI
|
|
24
|
Liu XX, Sun C, Jin XD, Li P, Zheng XG,
Zhao T and Li Q: Genistein sensitizes sarcoma cells in vitro and in
vivo by enhancing apoptosis and by inhibiting DSB repair pathways.
J Radiat Res. 57:227–237. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Marampon F, Gravina G, Ju X, Vetuschi A,
Sferra R, Casimiro M, Pompili S, Festuccia C, Colapietro A, Gaudio
E, et al: Cyclin D1 silencing suppresses tumorigenicity, impairs
DNA double strand break repair and thus radiosensitizes
androgen-independent prostate cancer cells to DNA damage.
Oncotarget. 7:5383–5400. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu Y, Majumder S, McCall W, Sartor CI,
Mohler JL, Gregory CW, Earp HS and Whang YE: Inhibition of
HER-2/neu kinase impairs androgen receptor recruitment to the
androgen responsive enhancer. Cancer Res. 65:3404–3409. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fang Y, Zhang Q, Wang X, Yang X, Wang X,
Huang Z, Jiao Y and Wang J: Quantitative phosphoproteomics reveals
genistein as a modulator of cell cycle and DNA damage response
pathways in triple-negative breast cancer cells. Int J Oncol.
48:1016–1028. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu X, Sun C, Jin X, Li P, Ye F, Zhao T,
Gong L and Li Q: Genistein enhances the radiosensitivity of breast
cancer cells via G2/M cell cycle arrest and apoptosis. Molecules.
18:13200–13217. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang Z, Wang CZ, Du GJ, Qi LW, Calway T,
He TC, Du W and Yuan CS: Genistein induces G2/M cell cycle arrest
and apoptosis via ATM/p53-dependent pathway in human colon cancer
cells. Int J Oncol. 43:289–296. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee J, Ju J, Park S, Hong SJ and Yoon S:
Inhibition of IGF-1 signaling by genistein: Modulation of
E-cadherin expression and downregulation of β-catenin signaling in
hormone refractory PC-3 prostate cancer cells. Nutr Cancer.
64:153–162. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tsuboy MS, Marcarini JC, de Souza AO, de
Paula NA, Dorta DJ, Mantovani MS and Ribeiro LR: Genistein at
maximal physiologic serum levels induces G0/G1 arrest in MCF-7 and
HB4a cells, but not apoptosis. J Med Food. 17:218–225. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hwang KA, Kang NH, Yi BR, Lee HR, Park MA
and Choi KC: Genistein, a soy phytoestrogen, prevents the growth of
BG-1 ovarian cancer cells induced by 17β-estradiol or bisphenol A
via the inhibition of cell cycle progression. Int J Oncol.
42:733–740. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kuerbitz SJ, Plunkett BS, Walsh WV and
Kastan MB: Wild-type p53 is a cell cycle checkpoint determinant
following irradiation. Proc Natl Acad Sci USA. 89:7491–7495. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Maréchal A and Zou L: DNA damage sensing
by the ATM and ATR kinases. Cold Spring Harb Perspect Biol.
5:a0127162013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rao F, Cha J, Xu J, Xu R, Vandiver MS,
Tyagi R, Tokhunts R, Koldobskiy MA, Fu C, Barrow R, et al: Inositol
pyrophosphates mediate the DNA-PK/ATM-p53 cell death pathway by
regulating CK2 phosphorylation of Tti1/Tel2. Mol Cell. 54:119–132.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Elledge SJ: Cell cycle checkpoints:
Preventing an identity crisis. Science. 274:1664–1672. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Du WW, Yang W, Liu E, Yang Z, Dhaliwal P
and Yang BB: Foxo3 circular RNA retards cell cycle progression via
forming ternary complexes with p21 and CDK2. Nucleic Acids Res.
44:2846–2858. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lee JJ, Lee JH, Gu MJ, Han JH, Cho WK and
Ma JY: Agastache rugosa Kuntze extract, containing the active
component rosmarinic acid, prevents atherosclerosis through
up-regulation of the cyclin-dependent kinase inhibitors p21
WAF1/CIP1 and p27 KIP1. J Functional Food. 30:30–38. 2017.
View Article : Google Scholar
|
|
39
|
Oki T, Sowa Y, Hirose T, Takagaki N,
Horinaka M, Nakanishi R, Yasuda C, Yoshida T, Kanazawa M, Satomi Y,
et al: Genistein induces Gadd45 gene and G2/M cell cycle
arrest in the DU145 human prostate cancer cell line. FEBS Lett.
577:55–59. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li P, Veldwijk MR, Zhang Q, Li ZB, Xu WC
and Fu S: Co-inhibition of epidermal growth factor receptor and
insulin-like growth factor receptor 1 enhances radiosensitivity in
human breast cancer cells. BMC Cancer. 13:2972013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kasten U, Plottner N, Johansen J,
Overgaard J and Dikomey E: Ku70/80 gene expression and
DNA-dependent protein kinase (DNA-PK) activity do not correlate
with double-strand break (dsb) repair capacity and cellular
radiosensitivity in normal human fibroblasts. Br J Cancer.
79:1037–1041. 1999. View Article : Google Scholar : PubMed/NCBI
|