Introduction
Laryngeal squamous cell carcinoma (LSCC) remains one
of the most common cancers of the upper respiratory tract. Even
though advanced treatment options are available, including surgery,
chemotherapy and radiotherapy, mortality associated with LSCC is
still extremely high (1).
Statistics have revealed that LSCC is one of a few cancers in which
the 5-year survival rate has decreased from 66 to 63% over the past
40 years, while the overall incidence is decreasing (2). Therefore, it is urgent to identify
specific biomarkers that can be used in the early diagnosis and
prognosis of LSCC. Since miRNAs have been reported to be present in
the blood of cancer patients (3),
utilized for targeted cancer therapy (4) and used in cancer prognosis (5), it is universally acknowledged that
deregulated miRNA expression has an important effect on cellular
genes which regulate the proliferation, metastasis and the cell
cycle, leading to the progression of LSCC (6,7).
Notably, due to the inherent heterogeneity of the miRNA sources in
the blood and tendency to degrade, free miRNA in the blood is not
an ideal biomarker in the diagnosis and prognosis of LSCC.
Extracellular vesicles, particularly exosomes, which
are membrane-bound particles ranging from 30–150 nm and not merely
lipid vesicles, also carry cargoes including nucleic acids,
proteins, receptors, lipids, and transcription factors (8). Exosomes are present in biological
fluids and are involved in multiple pathological and physiological
processes (9). They can either be
picked up by adjacent cells or carried to distant sites via
biological fluids and may therefore induce phenotypic modifications
in recipient cells (10). They are
promising candidates in the field of tumor liquid biopsy on account
of cancer-specific expression profiles and reflect cancer-bearing
status. To date, studies focusing on exosomal miRNAs indicated that
exosomal miRNAs are of value for diagnostic and prognostic
significance in head and neck carcinoma, including nasopharyngeal,
oral squamous cell carcinoma and LSCC. Previous studies indicated
that exosomal miR-24-3p impeded T-cell function by targeting
fibroblast growth factor (FGF11) and may serve as a potential
prognostic biomarker for nasopharyngeal carcinoma (11). Li et al demonstrated that
exosomes derived from hypoxic oral squamous cell carcinoma cells
delivered miR-21 to normoxic cells to elicit a prometastatic
phenotype (12). Additionally, Wang
et al found that serum exosomal miR-21 along with HOTAIR
were significantly correlated with clinical parameters of LSCC
(13). These findings indicate that
exosomal miRNAs have a distinctly important effect on the malignant
progression of head and neck carcinoma.
Exosomes contain selected miRNAs that could
contribute to intercellular communication (14). The process by which several miRNAs
are enclosed in exosomes is selective rather than indiscriminate
(15,16). Despite growing interest in studying
the exosomal miRNA difference between cancer cells and normal
cells, we still lack an understanding of the difference between
parental cellular miRNA and exosomal miRNA. Honegger et al
presented the first comprehensive analysis of cellular and exosomal
miRNAs, suggesting that there exists an enormous difference between
them (17). To the best of our
knowledge, the distribution characteristics and comprehensive
expression profile on the RNA content of LSCC-derived exosomes
remains unknown.
The overall goal of this study was to identify and
characterize selective exosomal miRNA expression profiles and
speculate their potential target via bioinformatics analysis. To
achieve this objective, we first isolated the exosomes derived from
the LSCC cell line AMC-HN-8, and then characterized exosome pellets
with transmission electron microscope (TEM), nanoparticle tracking
analysis (NTA) and flow cytometry (FCM). After extraction of total
RNA from cells and exosomes, next generation sequencing was carried
out. Notably, we identified that miR-1246, miR-1290, miR-335-5p,
miR-127-3p and miR-122-5p were upregulated and miR-4521, miR-4483,
miR-30b-5p, miR-29b-3p and miR-374b-5p were downregulated in
exosomes compared with parental cells. Finally, we revealed the
potential targets of these selective exosomal miRNAs via
bioinformatics analysis. Collectively, we speculated that these
selective exosomal miRNAs may play an important role in LSCC, and
shed light on the biological implication of LSCC and provided a
theoretical base for the further research.
Materials and methods
Cell culture and generation of
exosome-depleted FBS
The human laryngeal squamous carcinoma cell line
AMC-HN-8 which was established by Kim et al in 1997
(18) from patients with head and
neck cancer was preserved in our laboratory. Laryngeal squamous
carcinoma cell lines Tu212 and Tu686 were obtained from the Central
South University (Hunan, China). All 3 cell lines are
representative in vitro models for studying the biology of
head and neck carcinoma. All cells were cultured in RPMI-1640
(HyClone; GE Healthcare Life Sciences, Logan, UT, USA), 1%
penicillin-streptomycin (Genom Biotechnology, Hangzhou, China) and
10% exosome-depleted fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) in humidified air with 5%
CO2 at 37°C.
The generation of exosome-depleted FBS was carried
out by ultracentrifugation to reduce contamination from bovine
exosomes. Briefly, centrifuge tubes were loaded with FBS and
centrifuged at 120,000 × g for 6 h at 4°C (Beckman Coulter Optima
L-100XP Ultracentrifuge, SW 32Ti; Beckman Coulter, Inc., Fullerton,
CA, USA). The supernatant of FBS was then filtered using a 0.22-µm
filter (Merck KGaA, Darmstadt, Germany).
Conditioned medium collection and
exosome isolation
Cells were cultured in conditioned medium containing
10% exosome-depleted FBS for 72 h at 90–100% density. The
conditioned medium was harvested and centrifuged at 2,000 × g for
10 min followed by 10,000 × g for 30 min at 4°C. Then, the
supernatant was filtered using a 0.22-µm filter to eliminate
cellular debris thoroughly and concentrated using centrifugal
ultrafiltration (Amicon® Ultra-15 100 KDa; Merck KGaA)
to minimize the potential contamination, respectively.
Exosomes were isolated from processed conditioned
medium with Ribo™ Exosome Isolation Reagent (Guangzhou RiboBio Co.,
Ltd., Guangzhou, China), according to the manufacturer's
instructions. Briefly, processed conditioned medium was mixed with
Ribo™ Exosome Isolation Reagent at a ratio of 3:1 and incubated at
4°C overnight. After centrifugation at 1,500 × g for 30 min,
exosomes pellets were resuspended in appropriate phosphate-buffered
saline (PBS; HyClone; GE Healthcare Life Sciences) for TEM, NTA and
further research. The exosome pellets were used immediately or
stored at −80°C until use.
TEM
The morphology of exosome pellets was examined by
TEM. Briefly, a 20-µl of exosome-PBS solution drop was loaded onto
carbon-coated copper grids and permitted to stand for 1 min. The
filter paper was utilized to remove the excess solution. Then the
exosomes pellets were negatively stained with 20 µl uranyl acetate
dihydrate (2%) and allowed to stand for 1 min. Subsequently, the
filter paper was utilized to remove the excess fluid, again. The
sample was allowed to dry under a lamp for 10 min before viewing on
an FEI Tecnai G2 Spirit transmission electron microscope (FEI
Company, Hillsboro, OR, USA) operated at 80 kV.
NTA and FCM
For the purpose of demonstrating the particle size
distribution, NTA was performed using NanoSight NS300 (Malvern
Instruments, Inc., Westborough, MA, USA), according to the
operating instructions. In addition, FCM (BD Accuri C6 flow
cytometer; BD Biosciences, Franklin Lakes, NJ, USA) was used to
detect exosomal surface markers. Briefly, exosome pellets were
resuspended in 100 µl filtered PBS, and then 20 µl
immunofluorescence antibody CD63 (CD63-Antibody-FITC; cat. no.
557288) and CD81 (CD81-Antibody-FITC; cat. no. 551108; both from BD
Biosciences) were added to the exosome-PBS solution. Following
incubation, FCM was utilized to detect exosomal surface
markers.
Total RNA extraction
Total RNA in cells and exosomes was extracted using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) as previously described (14). The quality and concentration of RNA
were assessed using NanoDrop 2000 Spectrophotometer (NanoDrop
Technologies; Thermo Fisher Scientific, Inc.). RNA content was
assessed using an Agilent 2200 Bioanalyzer (Agilent Technologies,
Inc., Santa Clara, CA, USA). Total RNA was sequenced or
reverse-transcribed to cDNA immediately for further research.
Small RNA sequencing
Small RNA library preparation and sample sequencing
were accomplished with the assistance of Guangzhou RiboBio Co.,
Ltd., using an Illumina HiSeq™ 2500 device. Total RNA from LSCC
AMC-HN-8 cells (n=3) and exosomes (n=3) were concatenated with 5′-
and 3′-adaptors. After cDNA synthesis and PCR amplification, the
cDNA library (18–40 nt) was obtained using an acrylamide gel
purification method. Then, single end (SE) sequencing was
performed: 1×50 bp.
RT-qPCR validation
To validate the results of sequencing, we selected 6
miRNAs (miR-1246, miR-122-5p, miR-320d, miR-4483, miR-30c-5p and
let-7e-5p) for further research by RT-qPCR in all 3 cell lines and
its exosomes. Total RNA (300 ng) was reversed-transcribed using
PrimeScript™ RT reagent Kit (Takara Biotechnology Co., Ltd.,
Dalian, China) according to the manufacturer's instructions. The
expression level of miRNA was measured using SYBR®
Premix Ex Taq™ (Takara Biotechnology Co., Ltd.) following the
manufacturer's instructions. We used miR-93-5p as an internal
control, which has a stable expression level between cells and
exosomes, to normalize the relative expression ratio of miRNA,
using the 2−ΔΔCq method (19). An ABI 7500 PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.) was used to perform the
RT-qPCR and analyze the data. The thermocycling conditions were as
follows: holding stage: 95°C for 30 sec; cycling stage: 95°C for 5
sec and 60°C for 34 sec, 40 cycles; melt curve stage: 95°C for 15
sec, 60°C for 1 min and 95°C for 15 sec. The primers used are
listed in Table I.
 | Table I.The sequences of primers. |
Table I.
The sequences of primers.
| Sequence name | Primer
sequences |
|---|
| miR-93-5p | Forward:
5′GGCAAAGTGCTGTTCGTG3′ |
|
| Reverse:
5′CAGTGCGTGTCGTGGAGT3′ |
| miR-1246 | Forward:
5′GGGGAATGGATTTTTGG3′ |
|
| Reverse:
5′CAGTGCGTGTCGTGGAG3′ |
| miR-122-5p | Forward:
5′GGGTGGAGTGTGACAATGG3′ |
|
| Reverse:
5′CAGTGCGTGTCGTGGAGT3′ |
| miR-320d | Forward:
5′AAAAGCTGGGTTGAGAGGA3′ |
|
| Reverse:
5′CAGTGCGTGTCGTGGAGT3′ |
| miR-4483 | Forward:
5′GGGAAAAAGGGGTGGTCTG3′ |
|
| Reverse:
5′GTGCGTGTCGTGGAGTCG3′ |
| miR-30c-5p | Forward:
5′GGGGGTGTAAACATCCTAC3′ |
|
| Reverse:
5′GTGCGTGTCGTGGAGTCG3′ |
| let-7e-5p | Forward:
5′GGGGTGAGGTAGGAGGTTGT3′ |
|
| Reverse:
5′GTGCGTGTCGTGGAGTCG3′ |
Bioinformatics analysis
The target genes of selective exosomal miRNAs were
predicted using TargetScan (http://www.targetscan.org/mamm_31/), miRDB (http://www.mirdb.org/), miRanda (http://www.microrna.org/microrna/home.do) and StarBase
(http://starBase.sysu.edu.cn/). Only
target genes that were found by 3 of the 4 tools were identified to
be the target genes of exosomal miRNAs. The comprehensive function
annotations of the potential targets of selective exosomal miRNAs
were performed with Gene Ontology (GO) and Kyoto Encyclopedia of
Genes and Genomes (KEGG) analysis based on the DAVID 6.7 software
(http://david.abcc.ncifcrf.gov/home.jsp). All GO terms
and signaling pathways were analyzed with a threshold of
significance that was defined by a P-value of <0.05.
Results
Isolation and characterization of
exosomes derived from the LSCC cell line AMC-HN-8
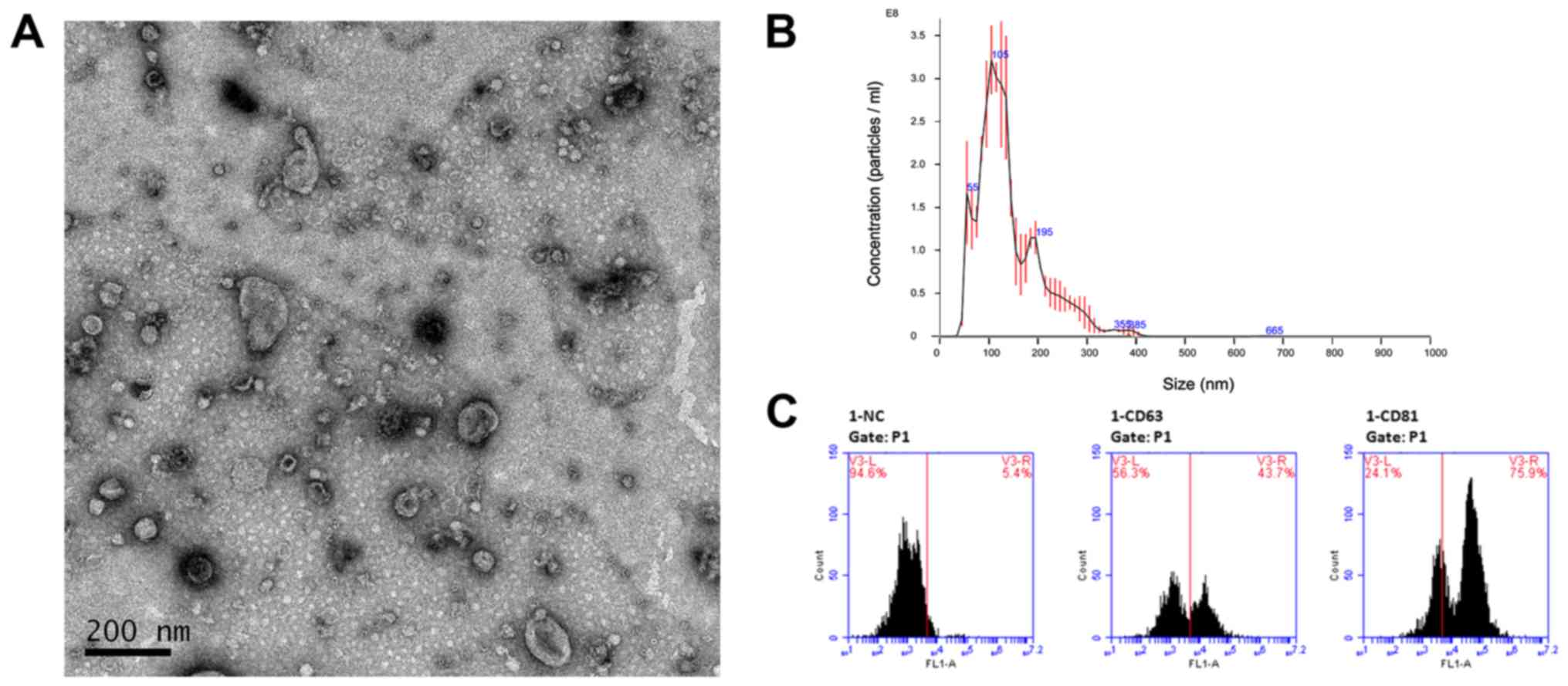
Our first step was to capture and verify the
exosomes derived from LSCC cell line AMC-HN-8. Exosomes were
isolated from conditioned medium of AMC-HN-8 using Ribo™ Exosome
Isolation Reagent. After isolation, the morphology of exosome
pellets was examined by TEM revealing membrane-bound particles that
were homogeneous in appearance (Fig.
1A). In addition, the size distribution of exosome pellets was
analyzed using NTA and the results revealed prospective diameters
which ranged from 30–150 nm (Fig.
1B). Furthermore, we assessed two well-established surface
markers for exosomes, CD63 and CD81, using flow cytometry (Fig. 1C). The results revealed a
significant difference after incubation with CD63 and CD81
antibodies. These results indicated that these pellets which we
isolated had the size (30–150 nm), specific surface markers and
morphology typical of that of exosomes.
MicroRNA expression profiles are
significantly altered between parental cells and exosomes
To identify the miRNA expression profiles of
parental cells and exosomes to investigate which miRNAs are
selectively enclosed in exosomes, we performed small RNA
sequencing. First, total RNA was extracted from parental cells
(n=3) and exosomes (n=3). A small RNA library preparation and
sample sequencing were conducted under the assistance of Guangzhou
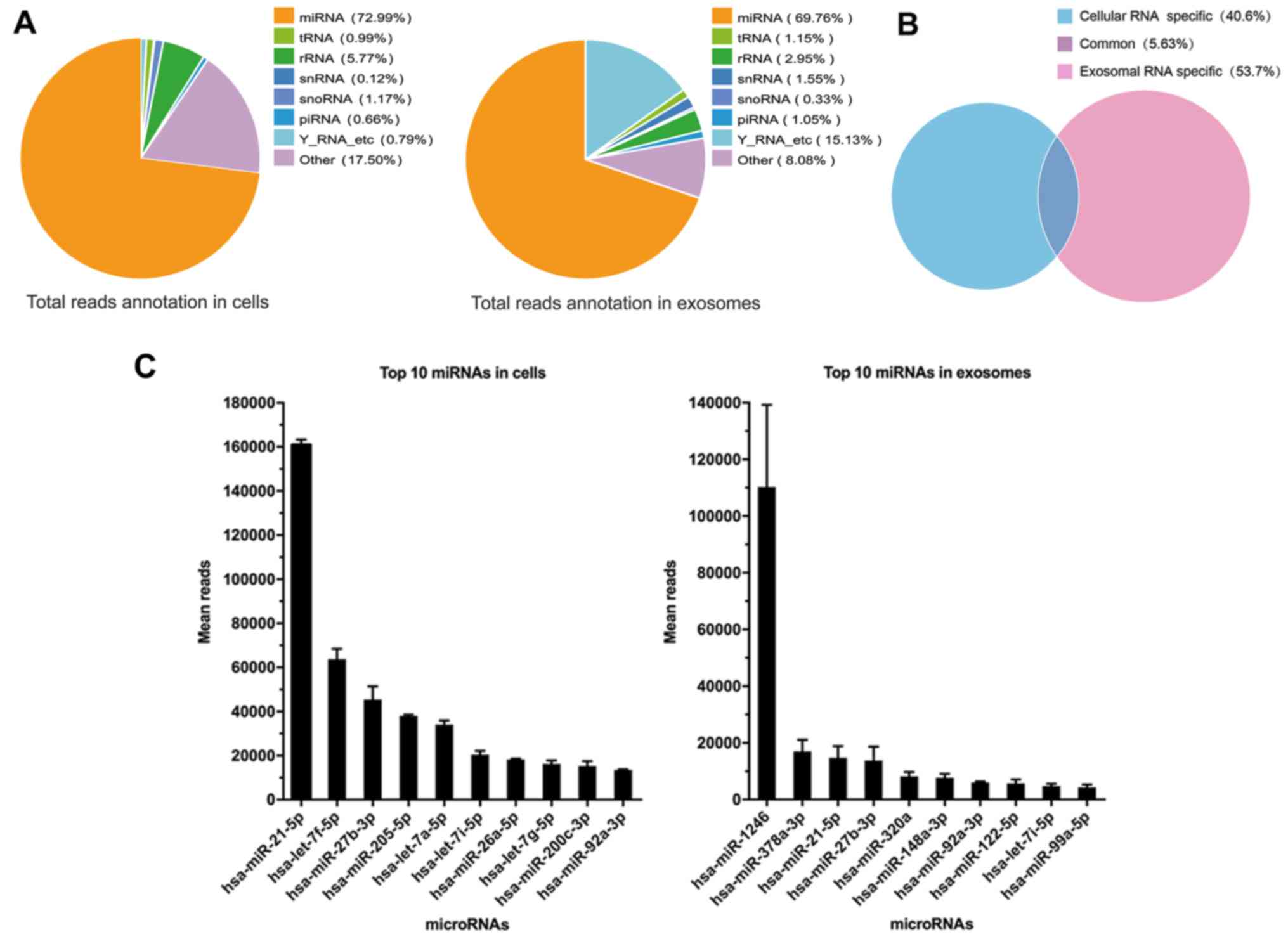
RiboBio Co., Ltd. The miRNAs were the most common among the known
sequences, followed by tRNAs, snRNAs, Y RNAs, piRNAs and other RNAs
which can not be classified. Of all the RNA examined, an average of
72.99% miRNAs were detected in parental cells compared with an
average of 69.76% miRNAs in exosomes derived from parental cells,
which did not exhibit an obvious discrepancy in composition of
small RNAs (Fig. 2A). However, only
5.63% miRNAs were common for both cellular miRNAs and exosomal
miRNAs, which meant that the process of miRNA packing into exosomes
is selective (Fig. 2B). The most
abundant miRNA in cells and exosomes was miR-21-5p and miR-1246,
respectively (Fig. 2C). miR-21-5p
has been previously reported to act as a predictor of recurrence
and novel biomarker of urothelial (20) and gastric carcinoma (21). In addition, exosomal miR-1246
induced cell motility and invasion in oral squamous cell carcinoma
(22), and functioned as a
promising biomarker in peripheral circulation of patients who have
breast and colon cancer (23,24).
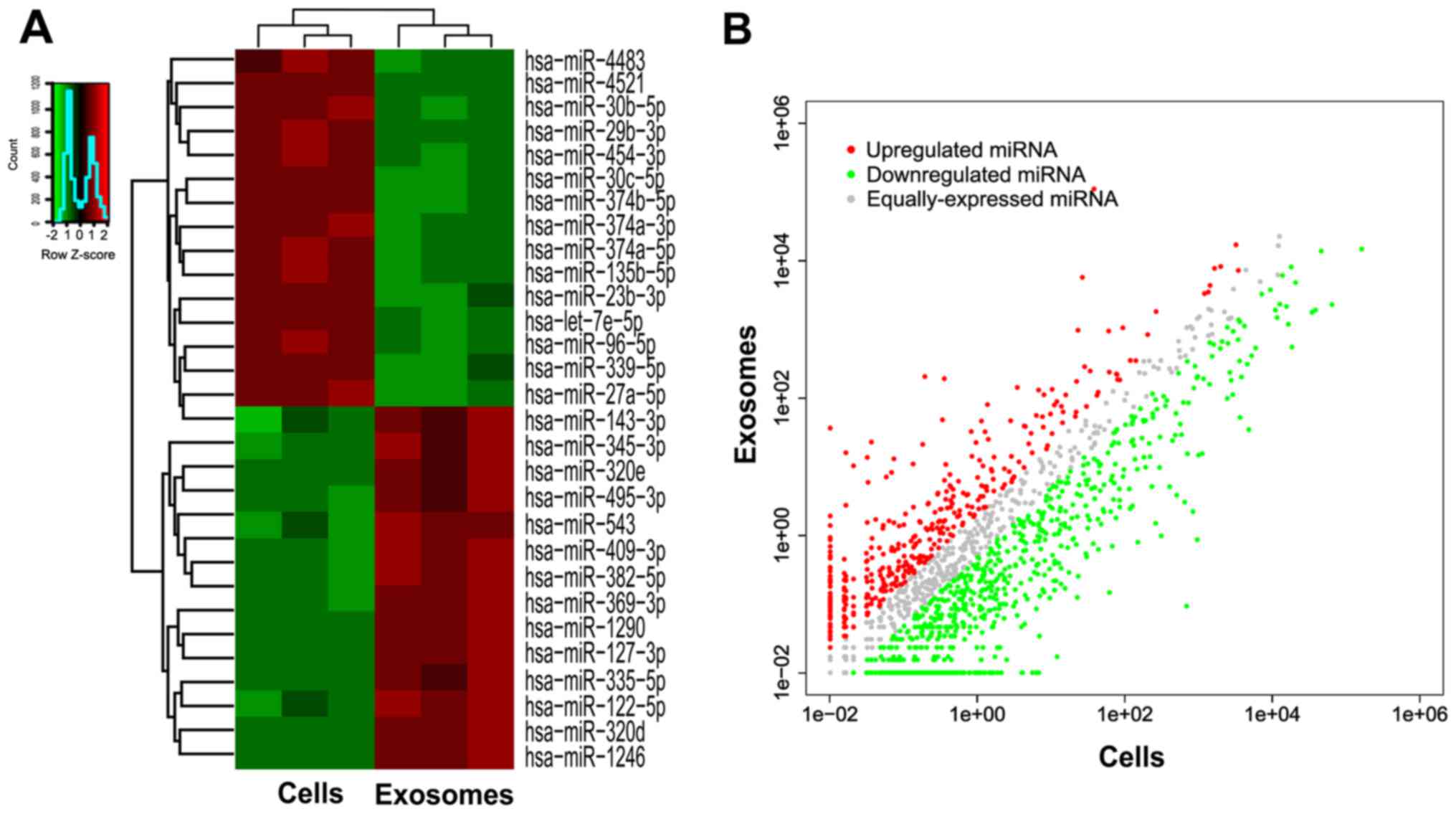
To explore the functions of selective miRNAs in exosomes, we then
detected 15 miRNAs downregulated in exosomes and 14 miRNAs
upregulated in exosomes as compared to parental cells, with both a
P-value of ≤0.05 and >5-fold-changes after filtering (Fig. 3). Detailed data are listed in
Table II.
| Table II.Expression of miRNAs in LSCC cell
line AMC-HN-8 and in exosomes as compared to parental cells. |
Table II.
Expression of miRNAs in LSCC cell
line AMC-HN-8 and in exosomes as compared to parental cells.
| miRNA | Mean reads in
cells | Mean reads in
exosomes | Log2
(fold-change) |
Upregulated/downregulated |
|---|
| miR-1246 | 37.75 | 110304.50 | 11.43 | Up |
| miR-1290 | 0.19 | 205.06 | 10.57 | Up |
| miR-335-5p | 0.04 | 22.78 | 9.43 | Up |
| miR-127-3p | 0.35 | 191.25 | 9.04 | Up |
| miR-122-5p | 26.24 | 5701.51 | 7.91 | Up |
| miR-369-3p | 0.33 | 48.14 | 7.17 | Up |
| miR-382-5p | 0.18 | 21.11 | 7.07 | Up |
| miR-409-3p | 1.35 | 80.08 | 5.89 | Up |
| miR-543 | 3.44 | 142.28 | 5.60 | Up |
| miR-495-3p | 0.82 | 32.45 | 5.31 | Up |
| miR-320d | 22.95 | 967.48 | 5.31 | Up |
| miR-320e | 1.12 | 46.15 | 5.25 | Up |
| miR-345-3p | 1.26 | 32.17 | 5.01 | Up |
| miR-143-3p | 6.70 | 130.52 | 5.01 | Up |
| miR-27a-5p | 699.66 | 15.40 | −5.57 | Down |
| miR-339-5p | 230.24 | 5.14 | −5.71 | Down |
| miR-96-5p | 988.10 | 14.73 | −6.05 | Down |
| let-7e-5p | 1109.58 | 15.48 | −6.26 | Down |
| miR-23b-3p | 3611.89 | 51.95 | −6.26 | Down |
| miR-135b-5p | 401.53 | 5.11 | −6.28 | Down |
| miR-374a-5p | 500.32 | 3.85 | −7.10 | Down |
| miR-30c-5p | 4737.26 | 34.67 | −7.17 | Down |
| miR-374a-3p | 259.11 | 1.50 | −7.49 | Down |
| miR-454-3p | 148.35 | 0.80 | −7.59 | Down |
| miR-374b-5p | 639.38 | 3.05 | −7.82 | Down |
| miR-29b-3p | 308.53 | 0.98 | −8.08 | Down |
| miR-30b-5p | 831.74 | 2.22 | −8.57 | Down |
| miR-4483 | 948.75 | 0.87 | −9.96 | Down |
| miR-4521 | 680.15 | 0.10 | −14.45 | Down |
All of these data revealed altered miRNA expression
profiles between parental cells and exosomes and indicated that
miRNAs are selectively enriched or encapsulated in exosome pellets
secreted from AMC-HN-8 cells rather than indiscriminate. These
selective miRNAs in exosomes may promote malignant biological
properties of tumors and play a crucial role in the cell-cell
communication, including tumor cells-tumor cells or tumor
cells-stromal cells crosstalk.
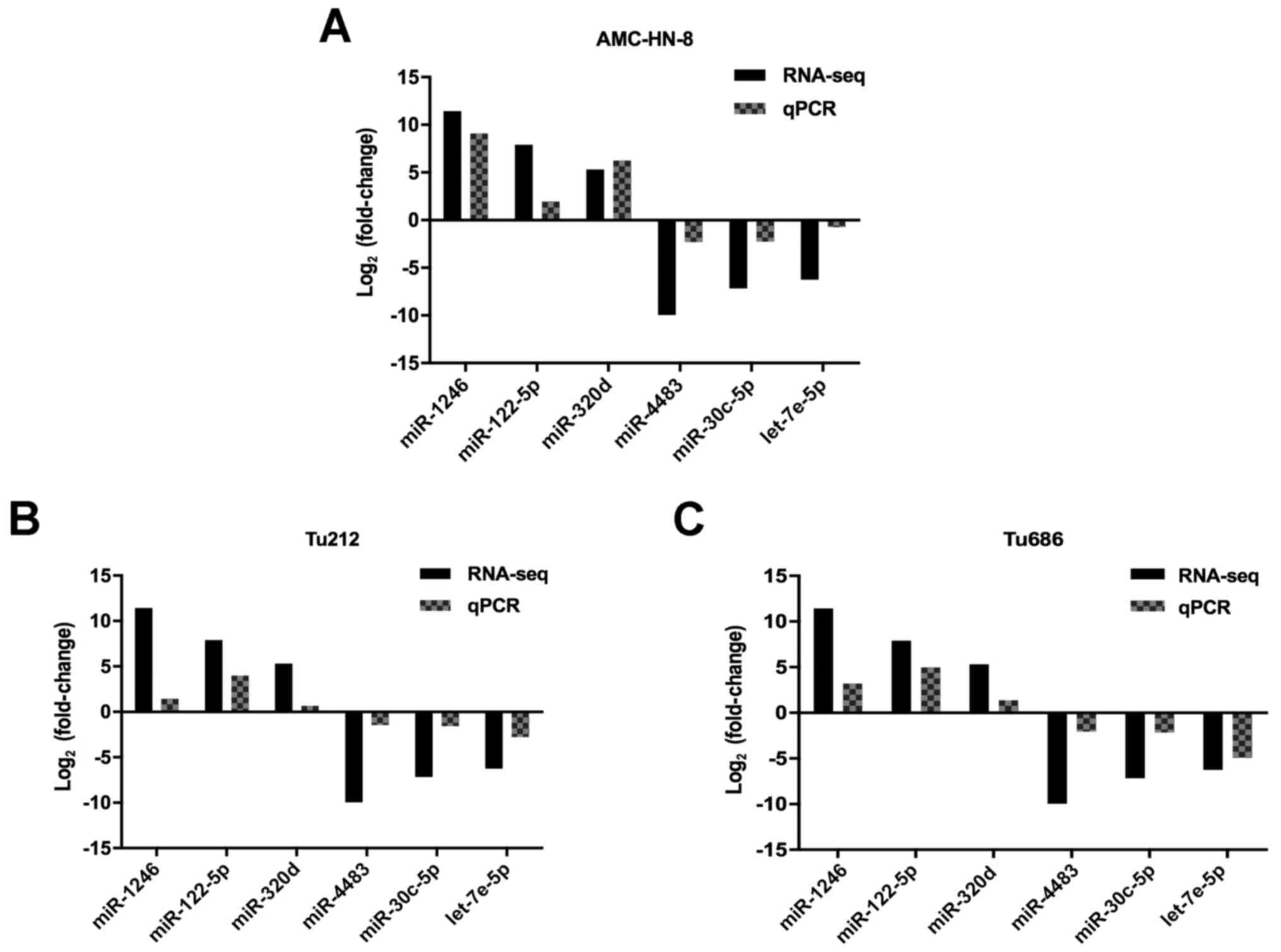
RT-qPCR reveals the same trend as the
sequencing results
To validate the results of small RNA sequencing,
RT-qPCR was carried out to detect 3 upregulated miRNAs (miR-320d,
miR-1246 and miR-122-5p) and 3 downregulated miRNAs (miR-4483,
miR-30c-5p and let-7e-5p) in exosomes derived from 3 cell lines.
Since no internal controls for exosomal microRNA analysis have been
established, we used miR-93-5p due to its stable expression level
between cells and exosomes. As anticipated, the results revealed
that the expression level of miR-320d, miR-1246 and miR-122-5p was
significantly upregulated, and miR-4483, miR-30c-5p and let-7e-5p
was significantly downregulated in exosomes derived from all 3 cell
lines, respectively (Fig. 4). These
results were consistent with the small RNA sequencing data, which
signified that the results of the small RNA sequencing were
credible.
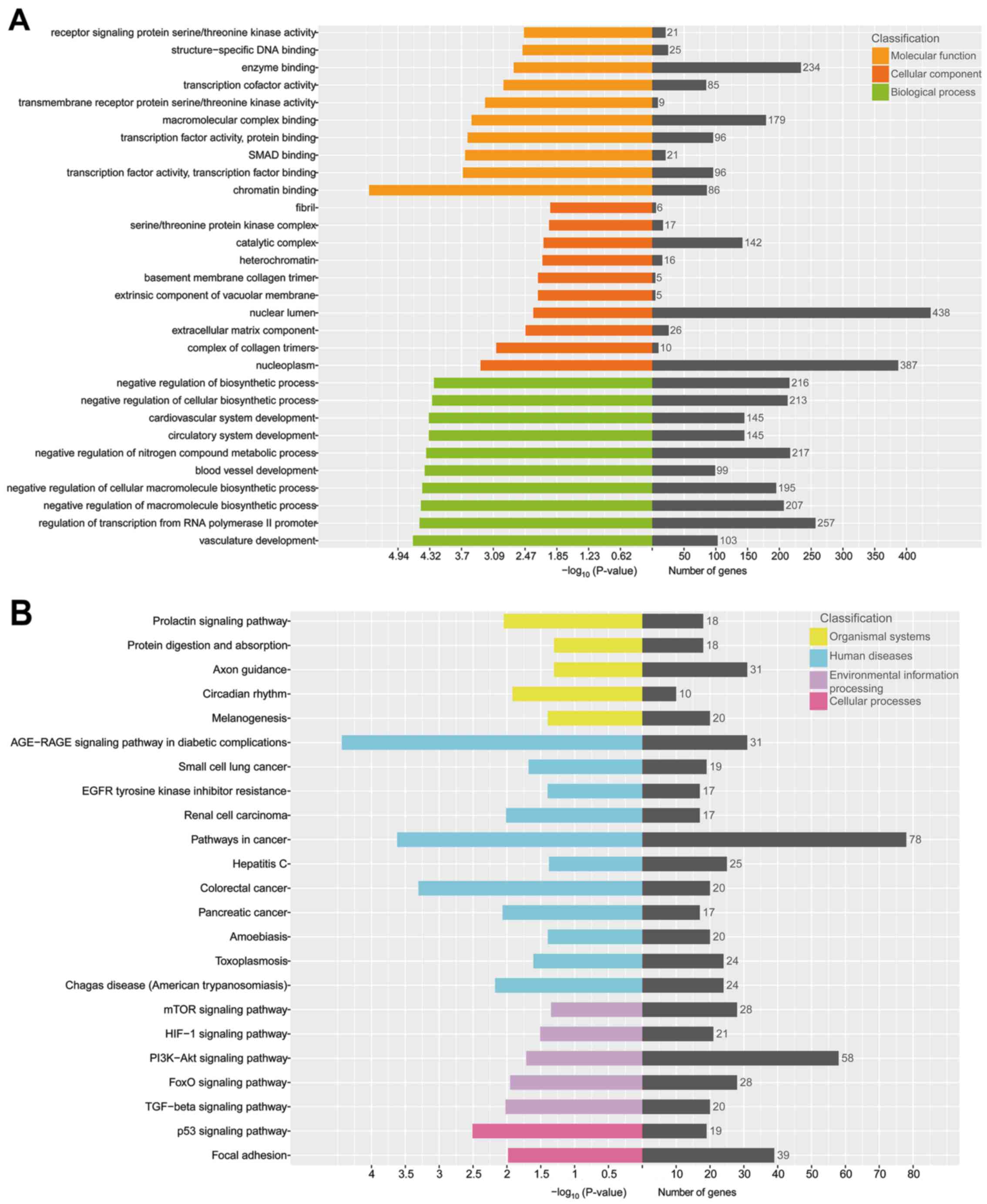
GO and KEGG pathway enrichment
analysis of selective exosomal miRNAs
Furthermore, GO annotation and KEGG pathway
enrichment analysis were performed to explore the potential targets
of these 29 selective exosomal miRNAs. The results revealed that a
total of 485 GO terms were involved in biological processes, with
P<0.05 (Fig. 5A). The top 3
biological processes with the lowest P-value were related to
vasculature development, regulation of transcription from RNA
polymerase II promoter and negative regulation of macromolecule
biosynthetic process. Up to 51 GO terms were in relation to
cellular components, with P<0.05. The top 3 cellular components
with the lowest P-value were related to nucleoplasm, complex of
collagen trimers and extracellular matrix component. There were 88
GO terms related to molecular function in all, with P<0.05. The
top 3 molecular functions with the lowest P-value were related to
chromatin binding, transcription factor activity, transcription
factor binding and SMAD binding.
The KEGG pathway enrichment analysis indicated that
279 pathways were involved in the current small RNA sequencing
data, in which many of these pathways were related to organismal
systems, cellular processes, environmental information processing
and human diseases (Fig. 5B). The
top 5 pathways, with the lowest P-value, were the AGE-RAGE
signaling pathway in diabetic complications, pathways in cancer,
colorectal cancer, the p53 signaling pathway and signaling pathways
regulating pluripotency of stem cells.
These results indicated that selective exosomal
miRNAs play a distinctly important role in the process of cell
metabolism, cell cycle and tumorigenesis.
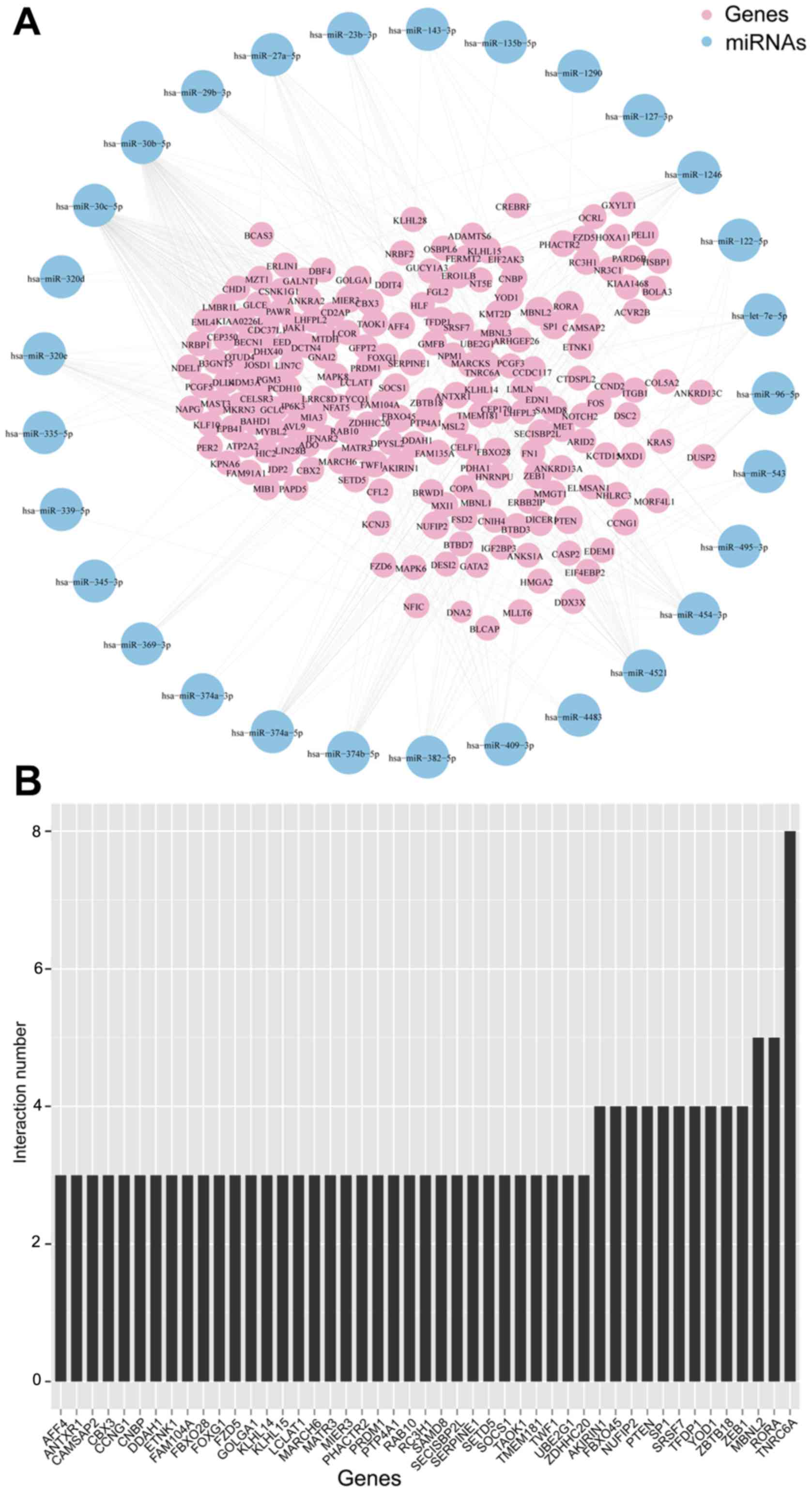
Selective exosomal miRNA-gene
co-expression network
We then constructed the co-expression network of
selective exosomal miRNAs and genes (Fig. 6A). The most common overlay was the
gene ‘TNRC6A’ (Fig. 6B), which was
previously reported that along with Ago2, was overexpressed in the
tissue of prostate and esophageal cancers (25). This network indicated that one
single miRNA was bound up with a couple of genes and vice
versa.
Discussion
Exosomes are nanometer-scale membrane vesicles
(ranging from 30–150 nm) of endocytic origin which are released
into the extracellular environment. Since exosomes are found in the
supernatant of maturing sheep reticulocytes (26,27),
studies have increasingly demonstrated that almost all types of
mammalian cells can release exosomes (8) and exosomes act as messengers carrying
bioinformation in the process of cell-cell communication rather
than a ‘waste bin’ for excreting useless content (28). MicroRNAs (miRNAs) are short (18–24
nt) ncRNAs that are associated with post-transcriptional regulation
of gene expression by affecting both the stability and translation
of mRNAs (29). Exosomal miRNAs
have gained much attention since Valadi et al (10) demonstrated that exosomes contain
miRNAs and these so-called ‘exosomal miRNAs’ can be delivered into
the extracellular environment, taken up by adjacent cells or
carried to distant sites via biological fluids. These exosomal
miRNAs may therefore induce the phenotypic modifications in
recipient cells, indicating that exosomes are important mediators
of intercellular communication and may play a significant role on
the malignant process of tumors.
Previous studies (30,31)
demonstrated that some specific miRNAs may be excreted or retained
in the cells preferentially, which means the process by which
several miRNAs are enclosed in exosomes is selective rather than
indiscriminate. These results implied that these selective exosomal
miRNAs may play a significant role in tumor malignant behaviors.
Therefore, we conducted small RNA sequencing to detect the
difference between LSCC AMC-HN-8 cells and exosomes which were
secreted from this cell line.
The LSCC cell line AMC-HN-8 was cultured in the
conditioned medium containing 10% exosome-depleted FBS (excluding
the contamination from bovine exosomes) for 72 h at 90–100%
density. After preprocessing the conditioned medium by
centrifugation, filtration and concentration, exosomes were
isolated with Ribo™ Exosome Isolation Reagent (32). To characterize the exosomes pellets,
we performed TEM and NTA that are universally recognized (33), and membrane-bound particles that
were homogeneous in appearance and ranging from 30–150 nm in size
were observed. Additionally, we detected two well-established
exosome surface markers CD63 and CD81 with flow cytometry. After
staining with the specific antibody, the positive rate ranged from
43.7–75.9%, which exhibited a significant difference. Specific
combining capacity of antibodies and distribution of antigens may
contribute to the slight difference of a positive rate of two
antibodies in one sample. Now that we successfully isolated the
exosome pellets derived from LSCC, small RNA sequencing was
performed.
Then, we identified the miRNA expression profiles of
parental cells and exosomes to investigate which miRNAs are
selectively enclosed in exosomes via small RNA sequencing. As the
most common known sequences in the cell, miRNAs occupied the
largest portion which was 72.99% followed by snRNAs, snoRNAs,
piRNAs and Y RNAs (Fig. 2A), which
acknowledged that miRNAs were the pivotal mediators in the vital
movement. Similar with the content of cells, exosomal miRNAs
derived from AMC-HN-8 cells made up to 69.76% of whole small RNAs.
However, only 5.63% miRNAs were detected commonly both in cells and
exosomes and 53.7% miRNAs were exclusively detected in exosomes
suggesting a very notable enrichment in exosomes compared to
parental cells (14). According to
the results, miR-21, let-7f-5p and miR-27-3p were identified as the
3 most abundant miRNA types in cells, and these miRNAs had been
previously reported to take part in tumor development, including
gastric (21), laryngeal (34) and esophageal carcinoma (35). Largely different from cellular
miRNAs, miR-1246 ranked first among exosomes which was consistent
with previous research (22,23,36)
indicating that exosomal miR-1246 may take part in tumor
development regardless of the type of tumor. Subsequently,
miR-378a-3p ranked second among exosomes, and miR-378a-3p was
reported to also exist in the exosomes derived from HPV-positive
cervical carcinoma cells (17).
miR-21-5p, consistent with cellular content, was also enriched in
exosomes and ranked third.
Subsequently, we investigated miRNAs with
log2 (fold-change) >2 and revealed their expression
level with heat map and volcano plots (Fig. 3). We identified miRNAs that were
enriched in cells or exosomes. To further narrow the research
scope, we selected 15 miRNAs that were downregulated in exosomes
and 14 miRNAs that were upregulated in exosomes as compared to
parental cells, with a P-value of both ≤0.05 and
>5-fold-changes. These miRNAs are listed in Table II. To validate the results of
sequencing, RT-qPCR was performed to detect 6 randomly selected
miRNAs (miR-320d, miR-1246, miR-122-5p, miR-4483, miR-30c-5p and
let-7e-5p) in 3 cell lines (AMC-HN-8, Tu212 and Tu686). From the
results obtained, we concluded that the qPCR results were in
agreement with the sequencing results, and that the sequencing
results were reliable.
Next, we predicted the potential targets of
selective exosomal miRNAs through TargetScan, miRDB, miRanda and
StarBase databases. Then, the comprehensive function annotations of
the potential targets were performed with GO and KEGG pathway
enrichment analysis. The GO analysis results revealed that the
potential targets of selective exosomal miRNAs were involved in
biological processes, cellular components and molecular functions,
including vasculature development, nucleoplasm and chromatin
binding. KEGG pathway enrichment analysis revealed that the
AGE-RAGE signaling pathway and p53 signaling pathway may be
involved in the process and development of these selective exosomal
miRNAs.
We then constructed the co-expression network of
selective exosomal miRNAs and genes (Fig. 5). In this network, we acquired
specific information about the interrelation between these
selective exosomal miRNAs and genes.
In conclusion, we successfully isolated the exosomes
derived from LSCC cell line AMC-HN-8, and characterized the
exosomes with TEM, NTA and flow cytometry. Through the assistance
of small RNA sequencing combined with RT-qPCR analysis, we revealed
a significant difference between cellular miRNAs and exosomal
miRNAs, and that there is a very notable enrichment in exosomes
compared to parental cells. After GO annotation and KEGG pathway
enrichment analysis, we suggest that these selective exosomal
miRNAs may contribute to tumor development from various
perspectives.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and
Technology Commission of Shanghai Municipality, China (grant nos.
12J1402100 and 16411950101), and Shanghai ShenKang Hospital
Development Center, China (grant no. SHDC12015114). The sponsors or
funding organizations had no role in the design or execution of
this research.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
QH and LZ conceived and designed the study. QH, JY,
JZ, CH and YG performed the experiments. QH and LZ wrote the paper.
QH, LZ, JY, JZ, CH and YG reviewed and edited the manuscript. All
authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
miRNA
|
microRNA
|
|
LSCC
|
laryngeal squamous cell carcinoma
|
|
RT-qPCR
|
reverse transcription quantitative
PCR
|
|
GO
|
Gene Ontology
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
TEM
|
transmission electron microscope
|
|
NTA
|
nanoparticle tracking analysis
|
|
FCM
|
flow cytometry
|
References
|
1
|
Steuer CE, El-Deiry M, Parks JR, Higgins
KA and Saba NF: An update on larynx cancer. CA Cancer J Clin.
67:31–50. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Foekens JA, Sieuwerts AM, Smid M, Look MP,
de Weerd V, Boersma AW, Klijn JG, Wiemer EA and Martens JW: Four
miRNAs associated with aggressiveness of lymph node-negative,
estrogen receptor-positive human breast cancer. Proc Natl Acad Sci
USA. 105:13021–13026. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kelly EJ, Hadac EM, Greiner S and Russell
SJ: Engineering microRNA responsiveness to decrease virus
pathogenicity. Nat Med. 14:1278–1283. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ji J, Shi J, Budhu A, Yu Z, Forgues M,
Roessler S, Ambs S, Chen Y, Meltzer PS, Croce CM, et al: MicroRNA
expression, survival, and response to interferon in liver cancer. N
Engl J Med. 361:1437–1447. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cao P, Zhou L, Zhang J, Zheng F, Wang H,
Ma D and Tian J: Comprehensive expression profiling of microRNAs in
laryngeal squamous cell carcinoma. Head Neck. 35:720–728. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hui AB, Lenarduzzi M, Krushel T, Waldron
L, Pintilie M, Shi W, Perez-Ordonez B, Jurisica I, O'Sullivan B,
Waldron J, et al: Comprehensive MicroRNA profiling for head and
neck squamous cell carcinomas. Clin Cancer Res. 16:1129–1139. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tkach M and Théry C: Communication by
extracellular vesicles: Where we are and where we need to go. Cell.
164:1226–1232. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Van Niel G, D'Angelo G and Raposo G:
Shedding light on the cell biology of extracellular vesicles. Nat
Rev Mol Cell Biol. 19:213–228. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Valadi H, Ekström K, Bossios A, Sjöstrand
M, Lee JJ and Lötvall JO: Exosome-mediated transfer of mRNAs and
microRNAs is a novel mechanism of genetic exchange between cells.
Nat Cell Biol. 9:654–659. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ye SB, Zhang H, Cai TT, Liu YN, Ni JJ, He
J, Peng JY, Chen QY, Mo HY, Jun-Cui, et al: Exosomal miR-24-3p
impedes T-cell function by targeting FGF11 and serves as a
potential prognostic biomarker for nasopharyngeal carcinoma. J
Pathol. 240:329–340. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li L, Li C, Wang S, Wang Z, Jiang J, Wang
W, Li X, Chen J, Liu K, Li C, et al: Exosomes derived from hypoxic
oral squamous cell carcinoma cells deliver miR-21 to normoxic cells
to elicit a prometastatic phenotype. Cancer Res. 76:1770–1780.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang J, Zhou Y, Lu J, Sun Y, Xiao H, Liu M
and Tian L: Combined detection of serum exosomal miR-21 and HOTAIR
as diagnostic and prognostic biomarkers for laryngeal squamous cell
carcinoma. Med Oncol. 31:1482014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kogure T, Lin WL, Yan IK, Braconi C and
Patel T: Intercellular nanovesicle-mediated microRNA transfer: A
mechanism of environmental modulation of hepatocellular cancer cell
growth. Hepatology. 54:1237–1248. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dickman CT, Lawson J, Jabalee J, MacLellan
SA, LePard NE, Bennewith KL and Garnis C: Selective extracellular
vesicle exclusion of miR-142-3p by oral cancer cells promotes both
internal and extracellular malignant phenotypes. Oncotarget.
8:15252–15266. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lawson J, Dickman C, MacLellan S, Towle R,
Jabalee J, Lam S and Garnis C: Selective secretion of microRNAs
from lung cancer cells via extracellular vesicles promotes
CAMK1D-mediated tube formation in endothelial cells. Oncotarget.
8:83913–83924. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Honegger A, Schilling D, Bastian S,
Sponagel J, Kuryshev V, Sültmann H, Scheffner M, Hoppe-Seyler K and
Hoppe-Seyler F: Dependence of intracellular and exosomal microRNAs
on viral E6/E7 oncogene expression in HPV-positive tumor cells.
PLoS Pathog. 11:e10047122015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim SY, Chu KC, Lee HR, Lee KS and Carey
TE: Establishment and characterization of nine new head and neck
cancer cell lines. Acta Otolaryngol. 117:775–784. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Matsuzaki K, Fujita K, Jingushi K,
Kawashima A, Ujike T, Nagahara A, Ueda Y, Tanigawa G, Yoshioka I,
Ueda K, et al: MiR-21-5p in urinary extracellular vesicles is a
novel biomarker of urothelial carcinoma. Oncotarget. 8:24668–24678.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Park SK, Park YS, Ahn JY, Do EJ, Kim D,
Kim JE, Jung K, Byeon JS, Ye BD, Yang DH, et al: MiR 21-5p as a
predictor of recurrence in young gastric cancer patients. J
Gastroenterol Hepatol. 31:1429–1435. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sakha S, Muramatsu T, Ueda K and Inazawa
J: Exosomal microRNA miR-1246 induces cell motility and invasion
through the regulation of DENND2D in oral squamous cell carcinoma.
Sci Rep. 6:387502016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hannafon BN, Trigoso YD, Calloway CL, Zhao
YD, Lum DH, Welm AL, Zhao ZJ, Blick KE, Dooley WC and Ding WQ:
Plasma exosome microRNAs are indicative of breast cancer. Breast
Cancer Res. 18:902016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ogata-Kawata H, Izumiya M, Kurioka D,
Honma Y, Yamada Y, Furuta K, Gunji T, Ohta H, Okamoto H, Sonoda H,
et al: Circulating exosomal microRNAs as biomarkers of colon
cancer. PLoS One. 9:e929212014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yoo NJ, Hur SY, Kim MS, Lee JY and Lee SH:
Immunohistochemical analysis of RNA-induced silencing
complex-related proteins AGO2 and TNRC6A in prostate and esophageal
cancers. APMIS. 118:271–276. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pan BT, Teng K, Wu C, Adam M and Johnstone
RM: Electron microscopic evidence for externalization of the
transferrin receptor in vesicular form in sheep reticulocytes. J
Cell Biol. 101:942–948. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Johnstone RM, Adam M, Hammond JR, Orr L
and Turbide C: Vesicle formation during reticulocyte maturation.
Association of plasma membrane activities with released vesicles
(exosomes). J Biol Chem. 262:9412–9420. 1987.PubMed/NCBI
|
|
28
|
Becker A, Thakur BK, Weiss JM, Kim HS,
Peinado H and Lyden D: Extracellular vesicles in cancer:
Cell-to-Cell mediators of metastasis. Cancer Cell. 30:836–848.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guo H, Ingolia NT, Weissman JS and Bartel
DP: Mammalian microRNAs predominantly act to decrease target mRNA
levels. Nature. 466:835–840. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Baglio SR, Rooijers K, Koppers-Lalic D,
Verweij FJ, Pérez Lanzón M, Zini N, Naaijkens B, Perut F, Niessen
HW, Baldini N, et al: Human bone marrow- and adipose-mesenchymal
stem cells secrete exosomes enriched in distinctive miRNA and tRNA
species. Stem Cell Res Ther. 6:1272015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Endzeliņš E, Berger A, Melne V,
Bajo-Santos C, Soboļevska K, Ābols A, Rodriguez M, Šantare D,
Rudņickiha A, Lietuvietis V, et al: Detection of circulating
miRNAs: Comparative analysis of extracellular vesicle-incorporated
miRNAs and cell-free miRNAs in whole plasma of prostate cancer
patients. BMC Cancer. 17:7302017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen J, Yu Y, Li S, Liu Y, Zhou S, Cao S,
Yin J and Li G: MicroRNA-30a ameliorates hepatic fibrosis by
inhibiting Beclin1-mediated autophagy. J Cell Mol Med.
21:3679–3692. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Théry C, Amigorena S, Raposo G and Clayton
A: Isolation and characterization of exosomes from cell culture
supernatants and biological fluids. Curr Protoc Cell Biol Chapter.
3:Unit 3.22. 2006.
|
|
34
|
Yao XD, Li P and Wang JS: MicroRNA
differential expression spectrum and microRNA-125a-5p inhibition of
laryngeal cancer cell proliferation. Exp Ther Med. 14:1699–1705.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hummel R, Sie C, Watson DI, Wang T, Ansar
A, Michael MZ, Van der Hoek M, Haier J and Hussey DJ: MicroRNA
signatures in chemotherapy resistant esophageal cancer cell lines.
World J Gastroenterol. 20:14904–14912. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xu YF, Hannafon BN, Zhao YD, Postier RG
and Ding WQ: Plasma exosome miR-196a and miR-1246 are potential
indicators of localized pancreatic cancer. Oncotarget.
8:77028–77040. 2017.PubMed/NCBI
|