Introduction
Colorectal cancer (CRC) is the most frequently
diagnosed malignancy of the gastrointestinal tract. During recent
decades, the incidence and mortality of CRC have been rapidly
rising in China and the burden of this cancer will continue to be
severe in the future (1,2). In spite of treatment combinations
including surgery and adjuvant therapies, more than half of the
cases with resected CRCs will finally result in tumor recurrence
and metastasis. Current clinicopathological biomarkers are still
incapable of accurately predicting tumor progression or further
stratifying CRC patients with the same TNM stage. Therefore, it is
necessary to develop new prognostic biomarkers for improving the
stratification of CRC cases and for exploring original therapeutic
alternatives.
MicroRNAs (miRNAs) are ~22 nucleotide, endogenous,
non-coding RNAs that can induce translational suppression and mRNA
degradation by binding to complementary sequences in the
3′untranslated regions (3′UTR) of the targeted mRNAs. Several
studies revealed abundant miRNA expression abnormalities in CRC
tissues compared with normal tissues. A number of miRNAs were
proven to be involved in diverse biological processes of CRC such
as cell proliferation, apoptosis, angiogenesis, invasion and
metastasis. For example, the miR-143/145 cluster suppresses CRC
cell growth through modulation of the KRAS signaling pathway
(3,4). A positive feedback regulatory network
exists between p53 and miR-34a, and the latter indirectly
upregulates p53-dependent apoptosis in CRC (5,6). The
c-Myc-mediated transcriptional suppression of miR-15-16 in hypoxia
promotes tumor angiogenesis and hematogenous metastasis by
upregulating FGF2 (7). Furthermore,
a double-negative feedback loop between the miR-200 family and
E-cadherin transcriptional suppressors ZEB1 and ZEB2 mediates
epithelial-mesenchymal transition (EMT), subsequently regulating
the metastatic behavior of CRC (8).
Growing evidence suggests miR-197 as a novel
biomarker for various cancers (9).
Several studies revealed that miR-197 may be an oncomiR in cancer
cell invasion and metastasis. For example, miR-197 induces
epithelial-mesenchymal transition in pancreatic cancer cells along
with the downregulation of p120 catenin, an E-cadherin interacting
protein (10). By targeting CD82,
miR-197 may also play a vital role as an invasion and migration
promoter in gastric cancer and hepatocellular cancer via the
EGFR-ERK1/2-MMP7 signaling pathway (11,12).
Our previous research revealed that miR-197 could influence the
sensitivity of CRC cells to fluorouracil (5-FU) by modulating the
expression of thymidylate synthase (TYMS); however it had no impact
on cell proliferation and cell cycle regulation (13). However, it is unclear whether
miR-197 exhibits other biological functions in CRC. Thus, in the
present study, we intended to uncover the role of miR-197 in cell
migration and invasion of CRC and the possible underlying
mechanisms.
Materials and methods
Tumor cell culture
HCT8 (ileocecal colorectal adenocarcinoma), HCT116
(colorectal carcinoma) and SW480 (Dukes' type B, colorectal
adenocarcinoma) cell lines were purchased from the Cell Resource
Center, Institute of Basic Medical Sciences (IBMS), Chinese Academy
of Medical Sciences and Peking Union Medical College (CAMS/PUMC)
(Beijing, China). They were maintained in Gibco™ Dulbecco's
modified Eagle's medium (DMEM) supplemented with 10% fetal calf
serum (FCS) (both from Thermo Fisher Scientific, Inc., Waltham, MA,
USA) in a humidified 5% CO2/95% air atmosphere at
37°C.
miRNA and siRNA transfection
The synthetic miR-197 mimic, miR-197 inhibitor,
mimic control and inhibitor control were purchased from GenePharma
Inc. (Shanghai, China), and their sequences are listed in Table I. The siRNAs for IGFBP3 and siRNA
control were synthesized (Invitrogen; Thermo Fisher Scientific,
Inc.) and the sequences are listed in Table II.
 | Table I.Sequences of microRNAs. |
Table I.
Sequences of microRNAs.
| Name | Sequences |
|---|
| miR-197 mimic |
5′-UUCACCACCUUCUCCACCCAGC-3′ |
|
|
5′-UGGGUGGAGAAGGUGGUGAAUU-3′ |
| Mimic control |
5′-UUCUCCGAACGUGUCACGUTT-3′ |
|
|
5′-ACGUGACACGUUCGGAGAATT-3′ |
| miR-197
inhibitor |
5′-GCUGGGUGGAGAAGGUGGUGGUGAA-3′ |
| Inhibitor
control |
5′-CAGUACUUUUGUGUAGUACAA-3′ |
| Table II.IGFBP3 siRNA sequence. |
Table II.
IGFBP3 siRNA sequence.
| Name | Sense (5′-3′) | Antisense
(5′-3′) |
|---|
| IGFBP3 siRNA |
GUUGACUACGAGUCUCAGATT |
UCUGAGACUCGUAGUCAACTT |
In vitro migration and invasion
assays
Transwell chambers (8-µm pore size; Costar; Corning
Costar, Cambridge, MA, USA) were used in the in vitro
migration assay. Colorectal cancer cells were transfected with the
miR-197 mimic or the mimic control. Following 48 h, cells were
detached with trypsin, washed with PBS and resuspended in
serum-free medium. Two hundred microliters of cell suspension
(1×105 cells/ml for miR-197 mimic and its control,
2×105 cells/ml for miR-197 inhibitor and its control)
was added to the upper chamber, and 500 ml of complete medium was
added to the bottom well. The cells that had not migrated were
removed from the upper surfaces of the filters using cotton swabs,
and the cells that had migrated to the lower surfaces of the
filters were fixed with 4% paraformaldehyde solution and stained
with crystal violet. Images of 3 random fields (×10 magnification)
were captured from each membrane, and the number of migratory cells
was counted as previously described (14). Similar inserts coated with Matrigel
were used to determine the invasive potential.
RNA reverse transcription and
RT-qPCR
Total RNA was extracted using the Invitrogen™ TRIzol
total RNA isolation reagent (Thermo Fisher Scientific, Inc.) and
purified with the Column DNA Erasol kit (Tiangen Biotech Co., Ltd.,
Beijing, China) according to the manufacturer's instructions. mRNA
levels were assessed with RT-qPCR using SYBR-Green I (Takara
Biotechnology Co., Ltd., Dalian, China). The gene expression level
was normalized to the endogenous reference gene GAPDH. The
experiments were performed in triplicate. The primers for RT-qPCR
are listed in Table III. The
primers for miR-197 and U6 were purchased from Qiagen (Duesseldorf,
Germany) (Table IV). Reverse
transcription of miRNAs was performed with a miScript Reverse
Transcription kit (Qiagen). The expression of mature miRNAs was
determined using miRNA-specific quantitative RT-qPCR (Takara
Biotechnology Co., Ltd.). The expression levels were normalized to
the U6 endogenous control and assessed by the comparative Ct (ΔΔCq)
method (15).
| Table III.Primers (mRNA) for real-time PCR. |
Table III.
Primers (mRNA) for real-time PCR.
| Name | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| GAPDH |
GAAGGTGAAGGTCGGAGTC |
AAGATGGTGATGGGATTTC |
| IGFBP3 |
GAGGGCGACACTGCTTTTTC |
CCAGCTCCAGGAAATGCTAG |
| Table IV.Primers (miRNA) for real-time
PCR. |
Table IV.
Primers (miRNA) for real-time
PCR.
| miRNA | Primer sequence
(5′-3′) |
|---|
| hsa-miR-197 |
TTCACCACCTTCTCCACCCAGC |
| U6 |
CAAGGATGACACGCAAATTCG |
Prediction for target genes using
bioinformatics web server
Since miRNAs commonly act on posttranscriptional
regulation by targeting 3′UTRs of mRNAs of target genes, in order
to ascertain the mechanism underlying the involvement of miR-197 in
biological processes, we first searched for possible target genes
of miR-197 in the database on the web server ‘Targetscan’
(http://www.targetscan.org/vert_71/).
Subsequently, we determined the candidate genes for our study from
this repertoire of target genes by carefully reviewing
corresponding literature and selecting the most possible target
genes which had been reported to participate in cell migration,
invasion and tumor metastasis.
Dual-luciferase reporter gene
construct and dual-luciferase reporter assay
Fragments from 3 candidate genes including ADAMTS5,
CBL and IGFBP3 whose 3′-UTRs contained the predicted binding site
for hsa-miR-197, and flanking sequences on each side were
synthesized with a short extension containing cleavage sites for
XhoI (5′-end) and NotI (3′-end) (Table V); second fragments containing
mutated binding site sequences were also synthesized. The two
constructs were termed WT (Gene-wild type) and MT (Gene-mutant).
The fragments were cloned into the psiCHECK™-2 vector (Promega
Corporation, Madison, WI, USA). Ten nanograms of WT, MT or control
vector and 200 nmol/l miR-197 mimic were transfected into 293T
cells (293T cells were supplied by the Cell Center of Institue of
Basic Medical Sciences, Chinese Academy of Medical Sciences) using
Invitrogen™ Lipofectamine 2000 (Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. The cells were
harvested 24 h following transfection and assayed for
Renilla and firefly luciferase activities using the
Dual-Luciferase Reporter Assay System (Promega Corporation).
| Table V.Sequences of target genes and mutated
target genes. |
Table V.
Sequences of target genes and mutated
target genes.
| Target genes | Sequences |
|---|
| IGFBP3
wild-type | F
5′-TCGAGCAGCTGGCTACAGCCTCGATTTATATTTCTGTTTGTGGTGAACTGATTTTTTTTAAACCAAAGTTTAGAAAGAGGGC-3′ |
|
| R
5′-GGCCGCCCTCTTTCTAAACTTTGGTTTAAAAAAAATCAGTTCACCACAAACAGAAATATAAATCGAGGCTGTAGCCAGCTGC-3′ |
| IGFBP3 mutated | F
5′-TCGAGCAGCTGGCTACAGCCTCGATTTATATTTCTGTTTCTGATTTTTTTTAAACCAAAGTTAGAAAGAGGGC-3′ |
|
| R
5′-GGCCGCCCTCTTTCTAAACTTTGGTTTAAAAAAAATCAGAAACAGAAATATAAATCGAGGCTGTAGCCAGCTGC-3′ |
| ADAMTS5
wild-type | F
5′-TCGAGCAAAGATAACTGGAGGATTCAGCACTGATGCAGTCGTGGTGAACAGGAGGTCTACCTAACGCACAGAAAGTCATGCTTGC-3′ |
|
| R
5′-GGCCGCAAGCATGACTTTCTGTGCGTTAGGTAGACCTCCTGTTCACCACGACTGCATCAGTGCTGAATCCTCCAGTTATCTTTGC-3′ |
| ADAMTS5
mutated | F
5′-TCGAGCAAAGATAACTGGAGGATTCAGCACTGATGCAGTCCAGGAGGTCTACCTAACGCACAGAAAGTCATGCTTGC-3′ |
|
| R
5′-GGCCGCAAGCATGACTTTCTGTGCGTTAGGTAGACCTCCTGGACTGCATCAGTGCTGAATCCTCCAGTTATCTTTGC-3′ |
| CBL wild-type | F
5′-TCGAGAGCTATGAGGCACCTCCTACGTCTGTTTTCTGGCTGTGGTGACTTGGGATTTTTAACCTTATATATCTTTTTCCTTTGC-3′ |
|
| R
5′-GGCCGCAAAGGAAAAAGATATATAAGGTTAAAAATCCCAAGTCACCACAGCCAGAAAACAGACGTAGGAGGTGCCTCATAGCTC-3′ |
| CBL mutated | F
5′-TCGAGAGCTATGAGGCACCTCCTACGTCTGTTTTCTGGCTCTTGGGATTTTTAACCTTATATATCTTTTTCCTTTGC-3′ |
|
| R
5′-GGCCGCAAAGGAAAAAGATATATAAGGTTAAAAATCCCAAGAGCCAGAAAACAGACGTAGGAGGTGCCTCATAGCTC-3′ |
Western blot analysis
Following washing twice with PBS, cells were lysed
in ice-cold Radio Immunoprecipitation Assay (RIPA) lysis buffer
(Beyotime Institute of Biotechnology, Nanjing, China) and manually
scraped from the culture plates. Proteins were separated on 10%
sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) gels, electroblotted onto a polyvinylidene difluoride
(PVDF) membrane and incubated with anti-IGFBP3 antibody (1:1,000
dilution; cat. no. sc-135947; Santa Cruz Biotechnology, Inc., Santa
Cruz, CA, USA), anti-GAPDH antibody (1:1,000 dilution; cat. no.
sc-47724; Santa Cruz Biotechnology, Inc.), followed by incubation
with a secondary anti-rabbit or anti-mouse horseradish
peroxidase-conjugated antibody (1:1,000; Zhongshan Golden Bridge
Biotechnology, Beijing, China). Antibody-antigen complexes were
detected using a chemiluminescent ECL reagent (EMD Millipore,
Billerica, MA, USA).
Clinical samples
Fresh frozen samples of colorectal adenocarcinomas
were collected from 21 patients who underwent surgery at Peking
Union Medical College Hospital (PUMCH) from May 2015 to July 2015.
All patients had signed written informed consents prior to
enrolling in the study before admission. The use of human subjects
in this study was also approved by the Ethics Committee at Peking
Union Medical College Hospital (Beijing, China). This cohort
includes 11 males and 10 females patients with average age of 60.5
(46–72 years). Basic clinicopathological features of these 21
patients are listed in Tables VI
and VII.
| Table VI.Relationship of miR-197 and IGFBP3
expression with clinicopathological factors. |
Table VI.
Relationship of miR-197 and IGFBP3
expression with clinicopathological factors.
| Clinicopathological
features | miR-197 high
expression ΔCT level (≤7.64) n (%) | miR-197 low
expression ΔCT level (>7.64) n (%) | P-value | IGFBP3 Negative n
(%) | IGFBP3 Positive n
(%) | P-value |
|---|
| Tumor invasion |
|
| 0.090 |
|
| 0.131 |
|
T2-3 | 7 (41.2) | 10 (58.8) |
| 9 (52.9) | 8 (47.1) |
|
| T4 | 4 (100) | 0 (0) |
| 4 (100.0) | 0 (0%) |
|
| N staging |
|
| 0.635 |
|
| 0.336 |
| N1 | 4 (66.7) | 2 (33.3) |
| 5 (83.3) | 1 (16.7) |
|
| N2 | 7 (46.7) | 8 (53.3) |
| 8 (53.3) | 7 (46.7) |
|
| Histology |
|
| 0.149 |
|
| 0.325 |
| Good
and moderate | 10 (62.5) | 6 (37.5) |
| 11 (66.8) | 5 (31.2) |
|
|
Poor | 1 (20.0) | 4 (80.0) |
| 2 (40.0) | 3 (60.0) |
|
| Lymphovascular
invasion |
|
| 0.890 |
|
| 0.776 |
| No | 8 (53.3) | 7 (46.7) |
| 9 (60.0) | 6 (40.0) |
|
|
Yes | 3 (50.0) | 3 (50.0) |
| 4 (66.7) | 2 (33.3) |
|
| Site |
|
| 0.659 |
|
| 0.965 |
|
Colon | 6 (46.2) | 7 (53.8) |
| 8 (61.5) | 5 (38.5) |
|
|
Rectum | 5 (62.5) | 3 (37.5) |
| 5 (63.5) | 3 (37.5) |
|
| Table VII.Clinicopathological features of the
CRC patients. |
Table VII.
Clinicopathological features of the
CRC patients.
| No. | Histology | Site | Stage | miR-197 (ΔCt) | IGFBP3 (−/+) |
|---|
| 1 | Adenocarcinoma | Rectum | IIIB (pT3N1) | 7.69 | Positive |
| 2 | Adenocarcinoma | Colon | IIIB (pT3N1) | 7.69 | Positive |
| 3 | Adenocarcinoma | Rectum | IIIC (pT4N2b) | 7.38 | Negative |
| 4 | Adenocarcinoma | Rectum | IIIB (pT3N1) | 6.57 | Negative |
| 5 | Adenocarcinoma | Colon | IIIB (pT3N1) | 8.68 | Positive |
| 6 | Adenocarcinoma | Rectum | IIIB (pT3N1) | 7.72 | Positive |
| 7 | Adenocarcinoma | Colon | IIIB (pT3N2a) | 8.77 | Negative |
| 8 | Adenocarcinoma | Colon | IIIB (pT4aN1) | 2.20 | Negative |
| 9 | Adenocarcinoma | Rectum | IIIA (pT2N1) | 7.84 | Positive |
| 10 | Adenocarcinoma | Colon | IIIB (pT3N1) | 5.70 | Negative |
| 11 | Adenocarcinoma | Colon | IIIB (pT3N2a) | 4.34 | Negative |
| 12 | Adenocarcinoma | Colon | IIIB (pT3N1) | 7.94 | Positive |
| 13 | Adenocarcinoma | Rectum | IIIB (pT3N1) | 6.54 | Negative |
| 14 | Adenocarcinoma | Colon | IIIC (pT3N2b) | 2.37 | Negative |
| 15 | Adenocarcinoma | Colon | IIIB (pT3N1) | 8.75 | Positive |
| 16 | Adenocarcinoma | Colon | IIIB (pT3N2a) | 8.47 | Negative |
| 17 | Adenocarcinoma | Rectum | IIIB (pT4aN1) | 6.95 | Negative |
| 18 | Adenocarcinoma | Rectum | IIIB (pT4aN1) | 7.64 | Negative |
| 19 | Adenocarcinoma | Colon | IIIB (pT3N1) | 7.99 | Negative |
| 20 | Adenocarcinoma | Colon | IIIB (pT3N2a) | 7.22 | Positive |
| 21 | Adenocarcinoma | Colon | IIIB (pT3N1) | 7.00 | Negative |
Immunohistochemical staining
(IHC)
IGFBP3 expression was evaluated by IHC staining.
Briefly, after 5-µm sections were deparaffinized, antigen retrieval
was performed by use of heat-induced epitoperetrieval with 10 mM
citrate buffer. Sections were incubated with a monoclonal anitbody
against IGFBP3 (cat. no. sc-135947 Santa Cruz Biotechnology, Inc.)
at 1:250 dilution. The IGFBP3 antibody was detected using the
avidin-biotin-peroxidase technique (Dako LSAB kit; Dako,
Carpinteria, CA, USA). The expression levels of IGFBP3 were
determined by a pathologist. The classification of ‘- and +’ was
defined by the percentage of IGFBP3 positive cells at the level of
<5 and ≥5%, respectively.
Statistical analysis
Comparisons between different groups in migration
and invasion assays were analyzed using t-tests (two-sided). ANOVA
followed by Dunnette's test was applied to compare intergroup
differences in luciferase reporter assays. Correlation between
miR-197 and IGFBP3 expression was determined by correlate bivariate
Spearman assay. Chi-square Fisher's exact test were used to
calculate differences of miR-197 expression between different
groups of clinicopathological factors. All data were calculated
using SPSS 19.0 (IBM Corp., Armonk, NY, USA) and two-sided P-value
<0.05 was considered to indicate a statistically significant
difference.
Results
miR-197 enhances migration and
invasion in CRC cell lines in vitro
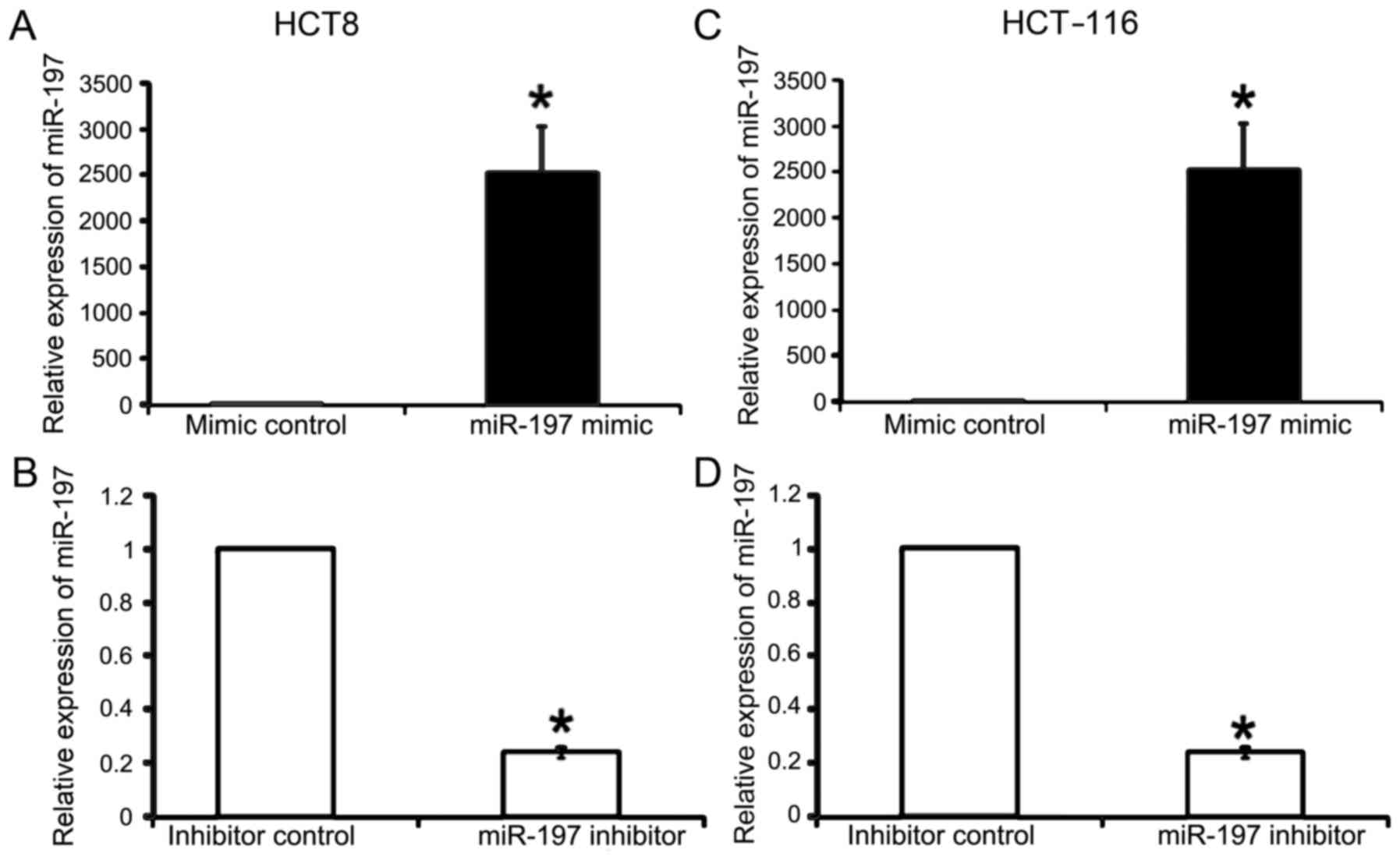
We first evaluated the efficacy of miR-197
overexpression and inhibition using synthetic miR-197 mimic,
miR-197 inhibitor and corresponding controls in CRC cell lines
(Fig. 1). We then investigated the
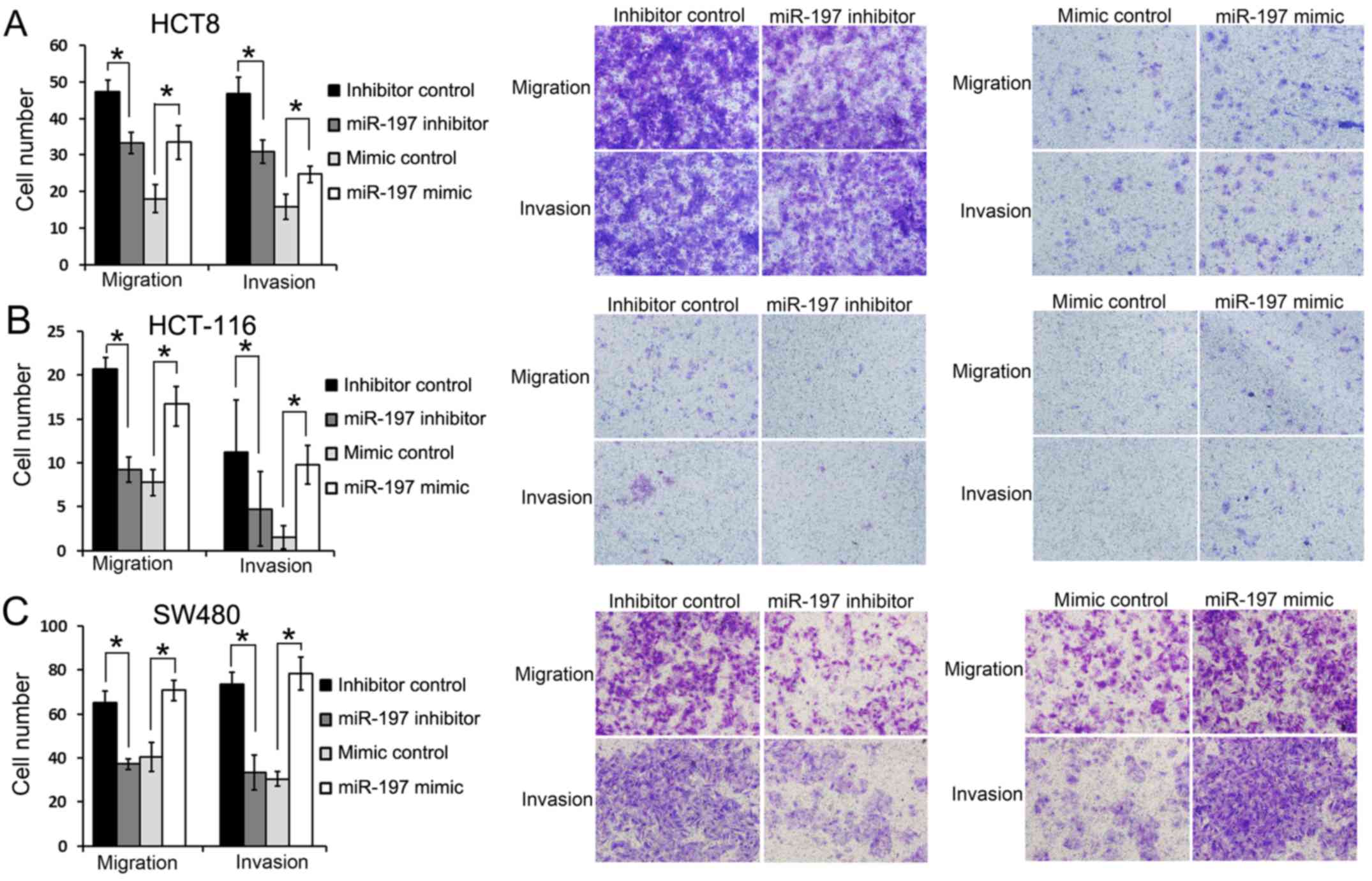
effect of miR-197 on the migration and invasion in colorectal cell
lines. HCT8, HCT116 and SW480 cells were transfected with either
miR-197 mimic, mimic control, miR-197 inhibitor or inhibitor
control and then subjected to cell migration and invasion assays
(Fig. 2). In the HCT8 cell line,
significantly more tumor cells in the miR-197 mimic group had
migrated to the lower surface of the filter compared with the
miR-197 mimic control group (33.50±4.65 vs. 18.00±3.91, P<0.05)
in the migration assay and similarly, significantly more cells
invaded to the lower surface in the miR-197 mimic group than its
control group in the invasion assay (24.75±2.21 vs. 18.00±3.91,
P<0.05, Fig. 2A). On the
contrary, significant less HCT8 cells in miR-197 inhibitor group
were found on the lower surface of the filter compared with its
control group in the migration assay (32.25±2.98 vs. 47.25±3.20,
P<0.05) and also in the invasion assay (31.00±3.16 vs.
46.75±4.42, P<0.05, Fig. 2A).
Furthermore, similar results were also achieved in the HCT116 and
SW480 cells (Fig. 2B and C),
indicating that miR-197 could regulate migration and invasion in
these 3 CRC cell lines in vitro.
miR-197 directly suppresses the
expression of IGFBP3 by targeting its 3′UTR
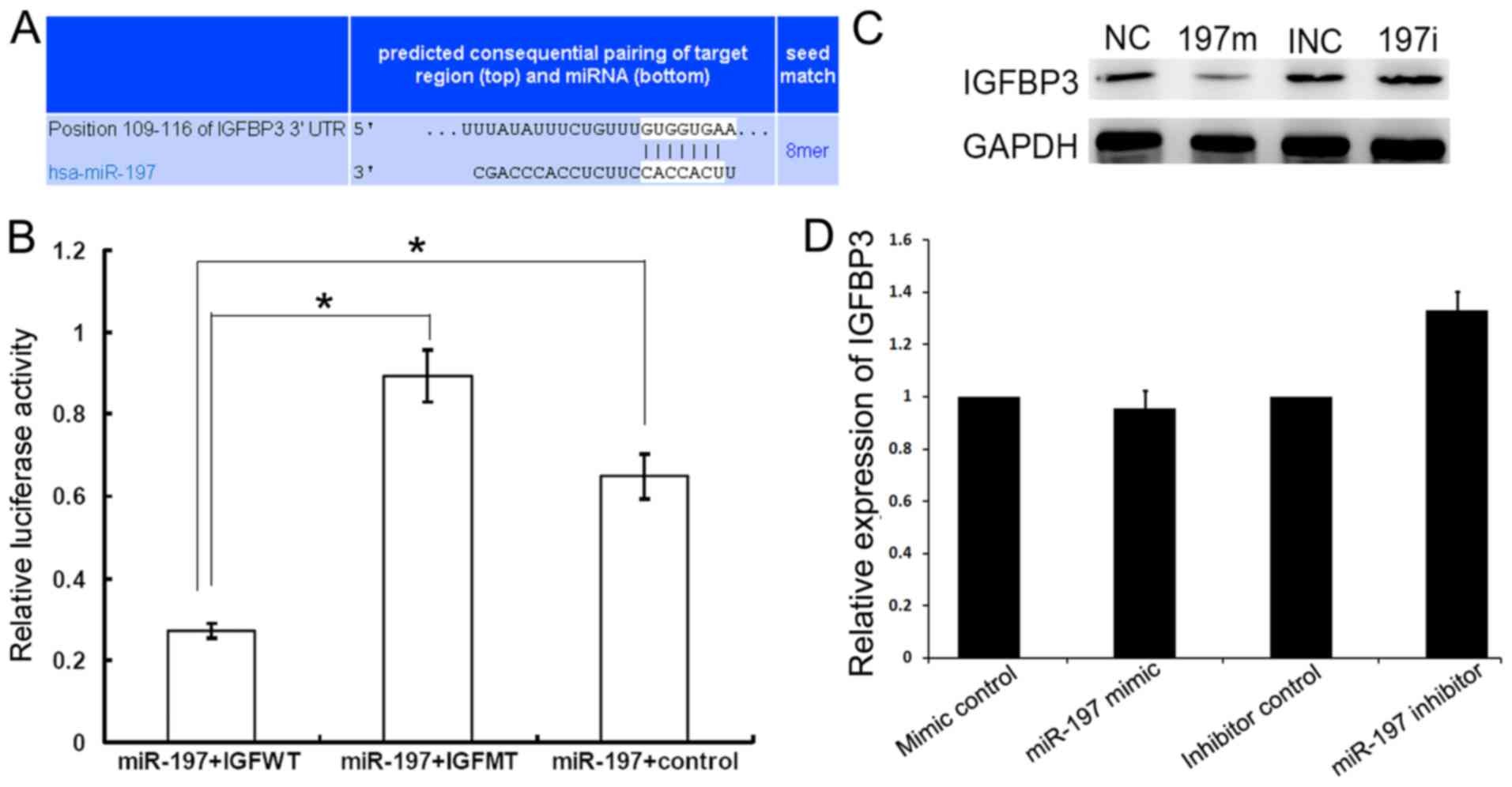
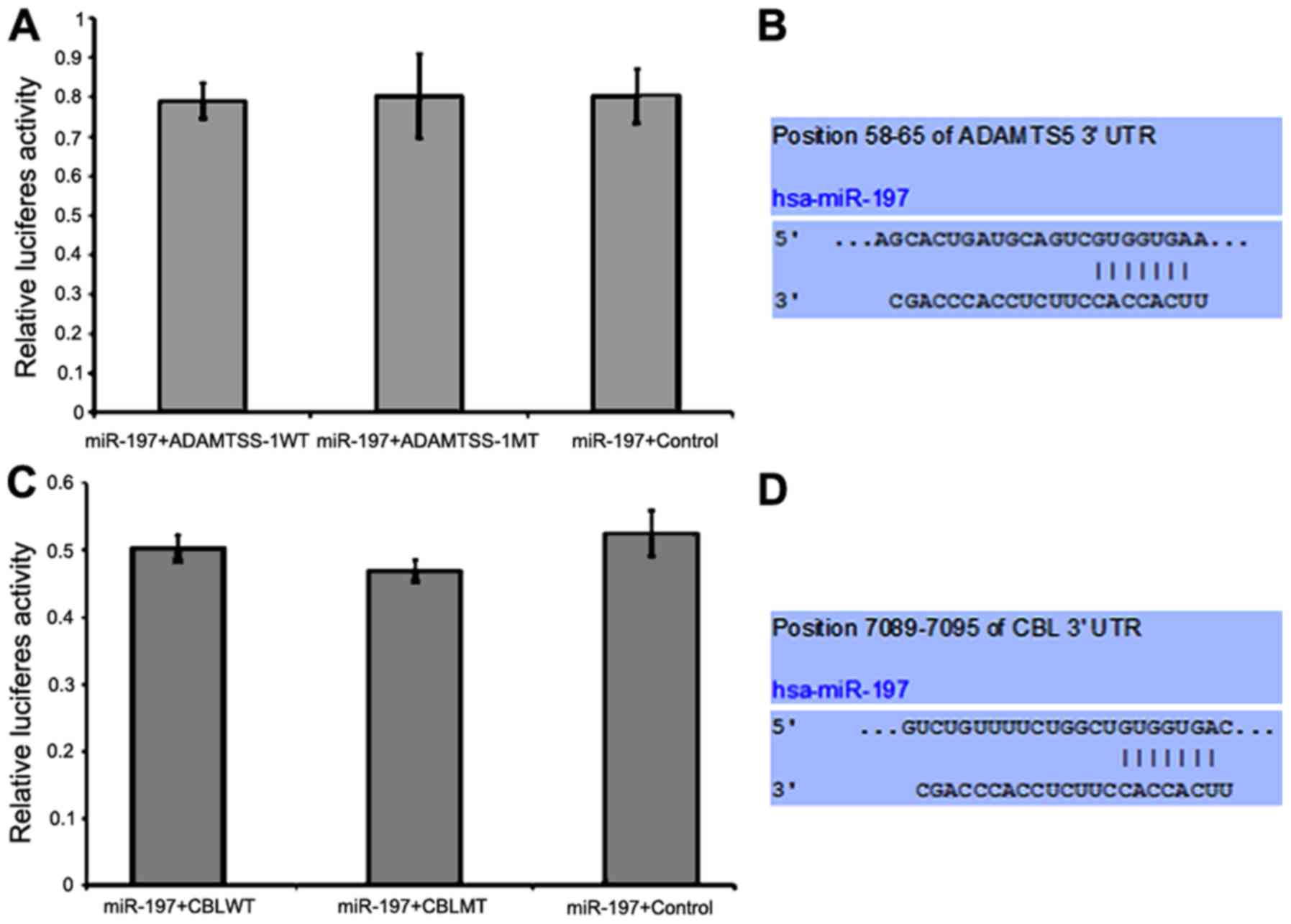
The mRNAs which may be targeted by miR-197 were
predicted, and genes associated with tumor invasion and metastasis
were selected. According to this principle, we screened out 3
candidate target genes including IGFBP3, ADAMTS5 and CBL. Predicted
miR-197 binding sites within the 3′UTR of mRNAs of these 3 genes
are listed in Figs. 3A, 4B and D, respectively.
Compared to the control group, miR-197 mimic
significantly decreased the luciferase activity when co-transfected
with IGFBP3-WT reporter plasmid (Fig.
3B, P<0.05). However, this inhibitory effect on luciferase
activity mediated by miR-197 mimic was abolished by mutant reporter
plasmid (Fig. 3B, P<0.05). In
addition, miR-197 overexpression had no effect on the luciferase
activities of ADAMTS5 (P=0.974, Fig.
4A) and CBL (P=0.098, Fig. 4C).
We decided to choose IGFBP3 as research target of our study due to
its tight connection with CRC according to multiple studies.
To demonstrate whether miR-197 could exert a similar
influence on IGFBP3 expression in CRC, we transfected HCT8 cell
line again with the miR-197 mimic, miR-197 inhibitor or their
corresponding controls, and then determined IGFBP3 expression at
the RNA and protein levels, respectively. The protein level of
IGFBP3 in HCT8 cells was obviously decreased following transfection
with miR-197 mimic and increased following transfection with
miR-197 inhibitor (Fig. 3C), while
the alterations of IGFBP3 mRNA did not reach statistical
significance upon overexpression and inhibition of miR-197
(P>0.05, Fig. 3D). These
findings revealed that miR-197 suppressed the protein output of
IGFBP3 rather than degrading its mRNA, indicating that IGFBP3 was
regulated by miR-197 in CRC cells at the post-transcriptional
level.
Downregulation of IGFBP3 boosts
migration and invasion of CRC cell lines
We then investigated the role of IGFBP3 in CRC cell
migration and metastasis. Small interfering RNA (siRNA) designed to
target IGFBP3 was employed. The transfection of siRNA resulted in a
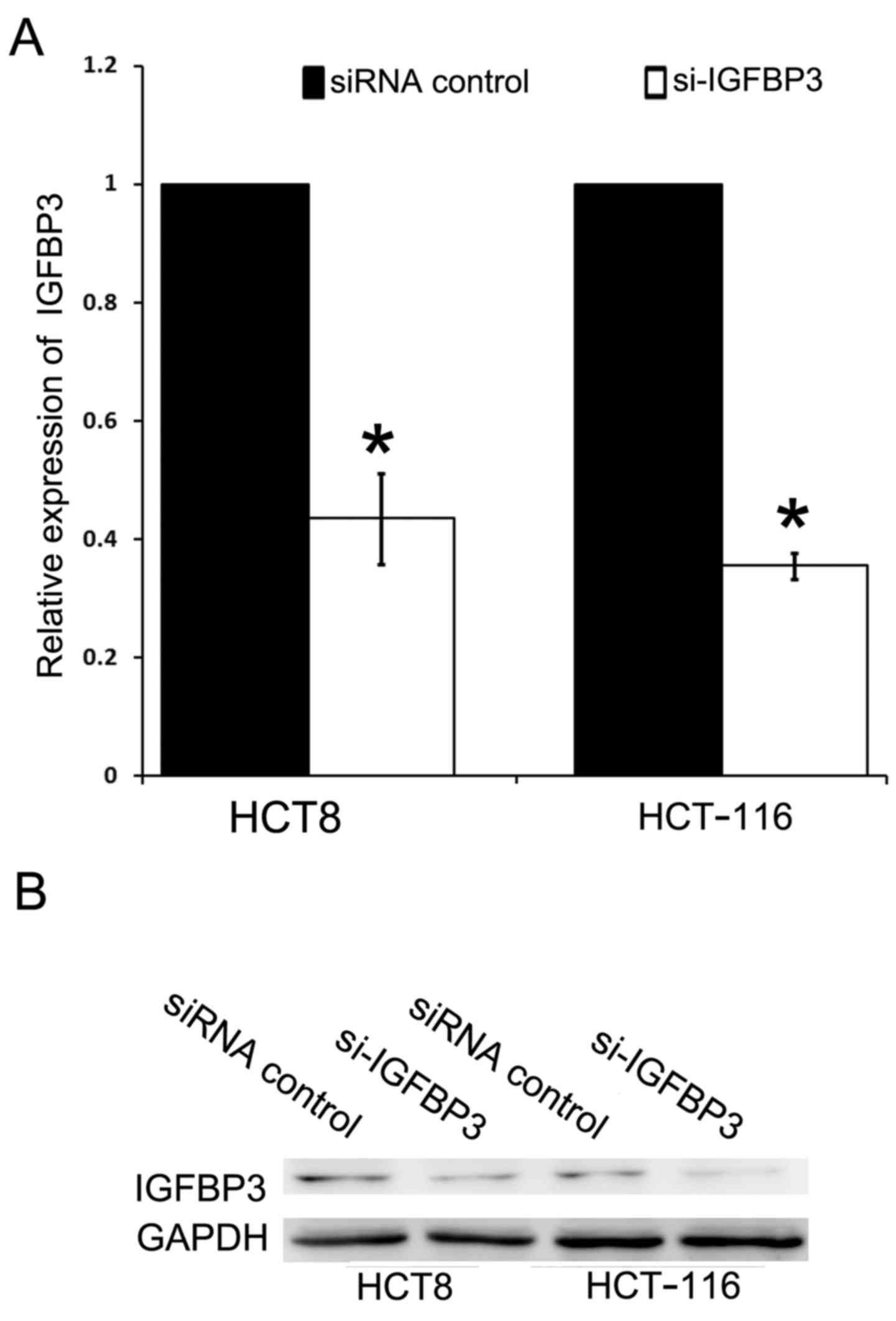
significant reduction in IGFBP3 mRNA and protein level in HCT8 and
HCT-116 cells compared to the siRNA-control (Fig. 5). Both cell lines were transfected
with either IGFBP3 siRNA (si-IGFBP3) or siRNA-control, and then
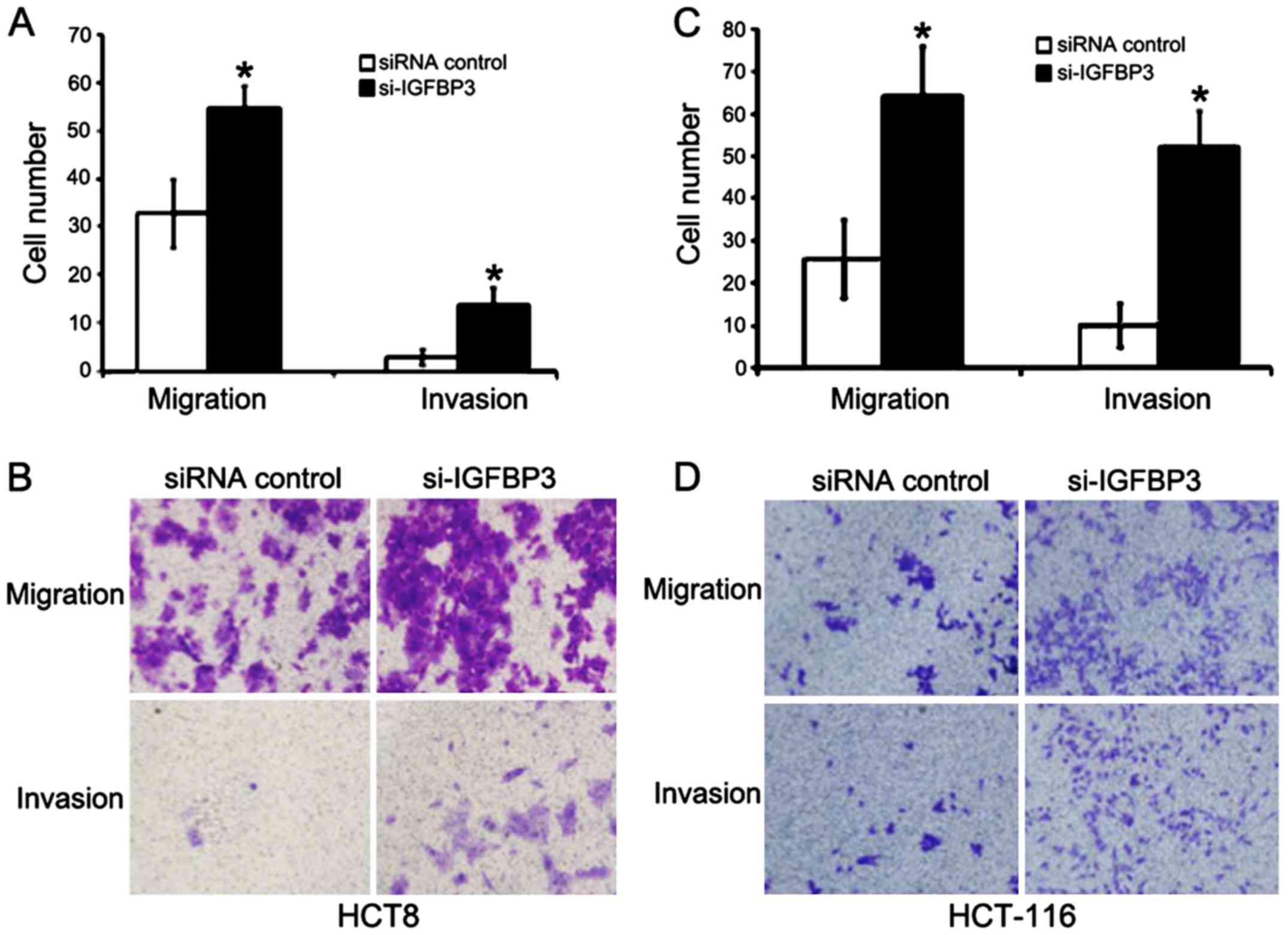
subjected to cell migration and cell invasion assay. In the
migration assay, the number of migratory HCT8 cells in the
si-IGFBP3 group was significantly greater than the migratory HCT8
cells in the si-control group (54.75±4.57 vs. 32.75±7.18,
P<0.05, Fig. 6A and B). In
addition, significantly more HCT8 cells in the si-IGFBP3 group had
invaded to the lower surface of filters compared with those in
si-control group (12.75±3.50 vs. 2.75±1.70, P<0.05) in the
invasion assay (Fig. 6A and B).
Similar results were also achieved in HCT-116 cells (Fig. 6C and D). Cell motility of both
assays was significantly enhanced following IGFBP3 downregulation
compared with the control in HCT-116 cells, with similar results in
HCT8 cells, which indicated that IGFBP3 was a suppressor protein in
the process of migration and invasion in CRC cells.
Correlation of miR-197 with IGFBP3 in
CRC patients
We assessed the miR-197 quantity of cancer tissues
using RT-PCR in fresh frozen samples and IGFBP3 expression using
IHC in corresponding paraffin samples in order to analyze the
correlation between miR-197 and IGFBP3 expression in CRC patients.
In this cohort of 21 stage III CRC patients, there were one stage
IIIA cancer, 2 stage IIIC cancers, and 18 patients with stage IIIB
cancer; 13 patients had colon cancer and 8 patients had rectum
cancer. The median ΔΔCq value of miR-197 was 7.64 (2.2–8.77). We
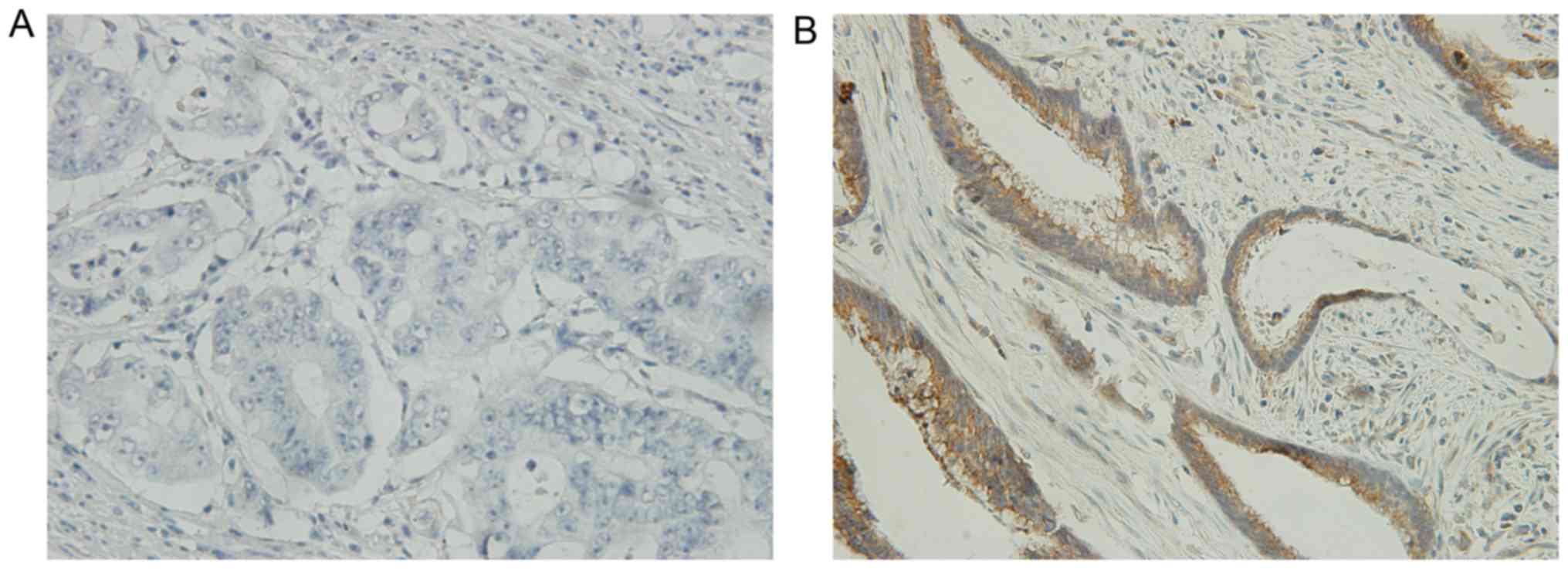
dichotomized IGFBP3 protein expression using ‘positive’ or
‘negative’ as displayed in Fig. 7.
There were 8 IGFBP3-positive and 13 IGFBP3-negative tumors. We
revealed that the miR-197 ΔΔCq level was significantly associated
with the expression of IGFBP3 with statistical significance
(P=0.026), so that miR-197 expression was negatively related to
IGFBP3 protein expression in the CRC tumor tissues.
Furthermore, we divided miR-197 expression level
into two groups using the median ΔΔCq value (≤7.64 vs. >7.64),
and explored whether there was any difference in miR-197 expression
between the different groups of clinicopathological factors.
Although we discovered a trend that there were more miR-197 high
expression tumors in the T4 group compared to the T2-3 group, it
was not statistically significant (P=0.09), and thus were other
factors such as lymph node stage, histology, lymphovascular
involvement and tumor site (Table
VI). In addition we did not detect any differences in the
expression of IGFBP3 between different groups of pathological
features (Table VI). Finally, we
aimed to clarify the association between prognosis and differential
expression of miR-197 plus IGFBP3, but we failed due to the limited
events of recurrence and death. Basic clinical information of this
cohort of CRC patients is listed in Table VII.
Discussion
The functions of miR-197 in tumorigenesis are
complicated. Several studies have found that miR-197 plays
controversial roles in the initiation and progression of different
malignancies. It decreases apoptosis in p53 wild-type non-small
cell lung cancer (NSCLC) (16), but
has the opposite function in multiple myeloma (17). It inhibits proliferation in
glioblastoma (18) but promotes
invasion and metastasis in gastric (11), pancreatic (10) and hepatocellular cancer (HCC)
(12). In colorectal cancer, our
previous studies revealed that miR-197 had no influence on
proliferation and cell cycle distribution, but it mediated the 5-FU
response by targeting thymidylate synthase in stage IV CRC
(13). In the present study, we
observed a unanimous phenomenon in 3 different CRC cell lines that
miR-197 upregulation promoted migration and invasion, while its
downregulation hindered the same process. This phenomenon in CRC
parallels that observed in other malignancies arising from the
epithelium. To the best of our knowledge, we revealed for the first
time the biological activities of miR-197 and the mechanism
underlying its function in CRC.
Several studies have revealed that miR-197, alone or
grouped with other miRNAs, may be a novel prognostic biomarker for
several human tumors that arise from epithelial tissues. For
example, miR-197 was found to be an unfavorable prognostic or
metastatic predictor in NSCLC, and to be associated with larger
tumors and squamous histology (19). Finally, meta-analysis displayed that
a panel of multiple miRNAs may deliver much greater predicting
efficacy than a single miRNA assay (20) and recently, 7 miRNA signatures
including miR-197 have been presented to be a feasible survival
indicator for CRC patients (21),
which collaborates our assumption about the role of miR-197 in CRC
prognosis. In order to clarify whether miR-197 may also regulate
IGFBP3 in CRC tissues similar to the results we achieved in
vitro, we detected the miR-197 level and protein expression of
IGFBP3 in tumor tissues of 21 CRC patients, and discovered a
negative association between miR-197 and IGFBP3. However, we failed
to detect any differences in the miR-197 expression level between
the different groups of clinicopathological characteristics. This
may be due to the fact that our cohort was small and highly
homogenous with stage III cancers. However a tendency of a greater
proportion of miR-197 high expression tumors was observed in the T4
group compared to the T2-3 group, which may hint that the deeper
the tumor invades, the more miR-197 is expressed. However, direct
evidence of the association of miR-197 with disease-free survival
and overall survival was not obtained, due to the fact that the
majority of these CRC cases were not presented with disease
recurrence and cancer-related death; therefore, there were not
enough data to come to a conclusion about the effect of miR-197 on
prognosis.
In the present study, we confirmed IGFBP3 as one of
the targets of miR-197 and their inverse correlation in CRC patient
samples. IGFBP3 is the most abundant IGFBP in circulation. It
attenuates IGF1-induced IGFR1 signaling events by sequestering IGF1
away from IGF1R (22). To date,
accumulating evidence revealed that the IGF1/IGF1R axis stimulates
epithelial to mesenchymal transition and contributes to tumor
metastasis including CRC (23,24).
In addition, IGFBP3 may also affect migration and invasion in a
complex intrinsic network, an IGFR-independent manner. For example,
IGFBP3 inhibits intracellular adhesion in head and neck squamous
cell carcinoma through blocking c-jun and c-fos transcription and
integrin β4 expression (25). It
also suppresses prostate cancer progression through degrading NF-κB
signaling (26). In CRC, IGFBP3 has
long been considered as a tumor suppressor (27). However, the biological role and
mechanism of IGFBP3 in the progression and metastasis in CRC are
poorly understood. We used in vitro assays and found that
IGFBP3 decreased migration and invasion in the HCT8 and HCT-116
cell lines. It is not known yet whether this inhibitory effect
correlates with disease progression, resembling the situation in
breast cancer (28). Thus far, the
cellular effects of IGFBPs were reported to be mediated
post-translationally. Other than methylation and proteolysis, miRNA
regulation may be another mode for IGFBP3 inactivation. For
example, miR-21 enhances glioblastoma tumorigenesis by
downregulating IGFBP3 (29);
onco-miRs miR-155 and miR-125b also target IGFBP3, promoting
migration and invasion in HCC and NSCLC through the IGF-II/IGF-1R
axis (30) and downstream PI3K/AKT
activation (31). miRNAs regulate
complementary mRNAs by inducing translational suppression and mRNA
destablization. Although IGFBP3 mRNA decay accounts for the
majority of miRNA-mediated regulations in the above-mentioned
studies, we speculated that miR-197 targets IGFBP3 to promote
invasion and migration in CRC through a translational suppression
manner, since the IGFBP3 mRNA level was only slightly altered while
its protein expression was significantly suppressed upon miR-197
expression. One comparable scenario is the regulation of
proliferation and invasion in CRC by miR-21 through the
PTEN/PI-3K/Akt signaling pathway (32). Dissociation of translation
initiation complex eIF4F was thought to be a mechanism underlying
miRNA-mediated translational suppression (33). Reactivation of suppressed targets by
degradation of RISC under certain contexts could also explain why
target mRNA does not have to end up in degradation (34). However, future studies are needed to
uncover how two silencing modes of miRNAs contribute to the overall
silencing of target mRNAs.
Due to its critical role in the IGF1/IGF1R axis and
various bioactivity, emerging clinical studies have attempted to
apply the serum IGFBP3 level and IGFBP3 methylation status for
prognostic prediction in CRC. An elevated serum IGFBP3 level was
previously reported to be correlated with a significant reduction
of CRC mortality (35). Recently,
high IGFBP3 gene methylation in the primary tumor has been
identified as an independent predictor for recurrence risk in stage
II CRC (36). However, other
studies using the same methodologies revealed quite contrary or
insignificant results (37,38). These data indicated that the
aforementioned IGFBP3 assessments may not be accurate reflections
of the local situation or IGFBP3 may be regulated by an intricate
system. We used IHC staining to evaluate local IGFBP3 protein
expression in tumor tissues but still no associations were found
with clinicopathological factors. Nevertheless, it is too soon to
exclude local IGFBP3 protein expression from being a biomarker. In
gastric cancer, IGFBP3 deficiency in IHC was found to be correlated
with advanced lymph node stage and poor survival (39). It is still necessary and promising
to probe the relationship of local IGFBP3 expression with other
clinicopathological factors and CRC patient prognosis in a larger
cohort with mature survival data.
In addition, we also explored two other possible
genes targeted by miR-197 using luciferase reporter assay and found
that miR-197 overexpression had no inhibitory effect on the
luciferase activity of CBL and ADAMTS5. Therefore, we excluded
these two genes as targets of miR-197. Attention should be drawn to
the fact that we did not succeed in cloning and expressing
full-length IGFBP3 because it is too large, which may compromise
the role of IGFBP3 as a tumor suppressor in CRC cell lines and its
relationship with miR-197 from the opposite angle.
In conclusion, in the present study, we revealed
that miR-197 overexpression could boost migration and invasion in
CRC cell lines. miR-197 directly regressed the expression of IGFBP3
in CRC cells through targeting its 3′UTR.
Acknowledgements
The authors thank all the patients who participated
in this study.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (nos. 81472785 and
61435001) and grants from The 2016 PUMCH Science Fund for Junior
Faculty (Pumch-2016-1.13).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
NZ and ZS co-conceived the study, managed its design
and drafted the manuscript. NL and YG participated in acquisition
and analysis of clinical data. JZ and QH contributed to the design
of the study and carried out the molecular biological assay. LZ and
CB participated in revising the manuscript critically for important
intellectrual content. All authors read and approved the manuscript
and agree to be accountable for all aspects of the research in
ensuring that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee at Peking Union Medical College Hospital (Beijing,
China). All patients signed written informed consents prior to
enrolling in the present study before admission.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang Y, Shi J, Huang H, Ren J, Li N and
Dai M: Burden of colorectal cancer in China. Zhonghua Liu Xing Bing
Xue Za Zhi. 36:709–914. 2015.(In Chinese). PubMed/NCBI
|
|
3
|
Chen X, Guo X, Zhang H, Xiang Y, Chen J,
Yin Y, Cai X, Wang K, Wang G, Ba Y, et al: Role of miR-143
targeting KRAS in colorectal tumorigenesis. Oncogene. 28:1385–1392.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mendell JT and Olson EN: MicroRNAs in
stress signaling and human disease. Cell. 148:1172–1187. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yamakuchi M, Ferlito M and Lowenstein CJ:
miR-34a repression of SIRT1 regulates apoptosis. Proc Natl Acad Sci
USA. 105:13421–13426. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bommer GT, Gerin I, Feng Y, Kaczorowski
AJ, Kuick R, Love RE, Zhai Y, Giordano TJ, Qin ZS, Moore BB, et al:
p53-mediated activation of miRNA34 candidate tumor-suppressor
genes. Curr Biol. 17:1298–1307. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xue G, Yan HL, Zhang Y, Hao LQ, Zhu XT,
Mei Q and Sun SH: c-Myc-mediated repression of miR-15-16 in hypoxia
is induced by increased HIF-2α and promotes tumor angiogenesis and
metastasis by upregulating FGF2. Oncogene. 34:1393–1406. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bracken CP, Gregory PA, Kolesnikoff N,
Bert AG, Wang J, Shannon MF and Goodall GJ: A double-negative
feedback loop between ZEB1-SIP1 and the microRNA-200 family
regulates epithelial-mesenchymal transition. Cancer Res.
68:7846–7854. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang DD, Chen X, Yu DD, Yang SJ, Shen HY,
Sha HH, Zhong SL, Zhao JH and Tang JH: miR-197: A novel biomarker
for cancers. Gene. 591:313–319. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hamada S, Satoh K, Miura S, Hirota M,
Kanno A, Masamune A, Kikuta K, Kume K, Unno J, Egawa S, et al:
MiR-197 induces epithelial-mesenchymal transition in pancreatic
cancer cells by targeting p120 catenin. J Cell Physiol.
228:1255–1263. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xu L, Hou Y, Tu G, Chen Y, Du YE, Zhang H,
Wen S, Tang X, Yin J, Lang L, et al: Nuclear Drosha enhances cell
invasion via an EGFR-ERK1/2-MMP7 signaling pathway induced by
dysregulated miRNA-622/197 and their targets LAMC2 and
CD82 in gastric cancer. Cell Death Dis. 8:e26422017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dai W, Wang C, Wang F, Wang Y, Shen M,
Chen K, Cheng P, Zhang Y, Yang J, Zhu R, et al: Anti-miR-197
inhibits migration in HCC cells by targeting KAI 1/CD82. Biochem
Biophys Res Commun. 446:541–548. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sun Z, Zhou N, Han Q, Zhao L, Bai C, Chen
Y, Zhou J and Zhao RC: MicroRNA-197 influences 5-fluorouracil
resistance via thymidylate synthase in colorectal cancer. Clin
Transl Oncol. 17:876–883. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sun Z, Han Q, Zhou N, Wang S, Lu S, Bai C
and Zhao RC: Micro RNA-9 enhances migration and invasion through
KLF17 in hepatocellular carcinoma. Mol Oncol. 7:884–894. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fiori ME, Barbini C, Haas TL, Marroncelli
N, Patrizii M, Biffoni M and De Maria R: Antitumor effect of
miR-197 targeting in p53 wild-type lung cancer. Cell Death Differ.
21:774–782. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang Y, Li F, Saha MN, Abdi J, Qiu L and
Chang H: miR-137 and miR-197 induce apoptosis and suppress
tumorigenicity by targeting MCL-1 in multiple myeloma. Clin Cancer
Res. 21:2399–2411. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tian LQ, Liu EQ, Zhu XD, Wang XG, Li J and
Xu GM: MicroRNA-197 inhibits cell proliferation by targeting GAB2
in glioblastoma. Mol Med Rep. 13:4279–4288. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mavridis K, Gueugnon F, Petit-Courty A,
Courty Y, Barascu A, Guyetant S and Scorilas A: The oncomiR miR-197
is a novel prognostic indicator for non-small cell lung cancer
patients. Br J Cancer. 112:1527–1535. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang R, Wen H, Xu Y, Chen Q, Luo Y, Lin Y,
Luo Y and Xu A: Circulating microRNAs as a novel class of
diagnostic biomarkers in gastrointestinal tumors detection: A
meta-analysis based on 42 articles. PLoS One. 9:e1134012014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu M, Kuang Y, Wang M, Han X and Yang Q: A
microRNA expression signature as a predictor of survival for colon
adenocarcinoma. Neoplasma. 64:56–64. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Firth SM and Baxter RC: Cellular actions
of the insulin-like growth factor binding proteins. Endocr Rev.
23:824–854. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li H, Batth IS, Qu X, Xu L, Song N, Wang R
and Liu Y: IGF-IR signaling in epithelial to mesenchymal transition
and targeting IGF-IR therapy: Overview and new insights. Mol
Cancer. 16:62017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yao C, Su L, Shan J, Zhu C, Liu L, Liu C,
Xu Y, Yang Z, Bian X, Shao J, et al: IGF/STAT3/NANOG/Slug signaling
axis simultaneously controls epithelial-mesenchymal transition and
stemness maintenance in colorectal cancer. Stem Cells. 34:820–831.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lee HJ, Lee JS, Hwang SJ and Lee HY:
Insulin-like growth factor binding protein-3 inhibits cell adhesion
via suppression of integrin β4 expression. Oncotarget.
6:15150–15163. 2015.PubMed/NCBI
|
|
26
|
Han J, Jogie-Brahim S, Harada A and Oh Y:
Insulin-like growth factor-binding protein-3 suppresses tumor
growth via activation of caspase-dependent apoptosis and cross-talk
with NF-κB signaling. Cancer Lett. 307:200–210. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Buckbinder L, Talbott R, Velasco-Miguel S,
Takenaka I, Faha B, Seizinger BR and Kley N: Induction of the
growth inhibitor IGF-binding protein 3 by p53. Nature. 377:646–649.
1995. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zielinska HA, Bahl A, Holly JM and Perks
CM: Epithelial-to-mesenchymal transition in breast cancer: A role
for insulin-like growth factor I and insulin-like growth
factor-binding protein 3? Breast Cancer. 7:9–19. 2015.PubMed/NCBI
|
|
29
|
Yang CH, Yue J, Pfeffer SR, Fan M, Paulus
E, Hosni-Ahmed A, Sims M, Qayyum S, Davidoff AM, Handorf CR and
Pfeffer LM: MicroRNA-21 promotes glioblastoma tumorigenesis by
down-regulating insulin-like growth factor-binding protein-3
(IGFBP3). J Biol Chem. 289:25079–25087. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
El Tayebi HM, Waly AA, Assal RA, Hosny KA,
Esmat G and Abdelaziz AI: Transcriptional activation of the
IGF-II/IGF-1R axis and inhibition of IGFBP-3 by miR-155 in
hepatocellular carcinoma. Oncol Lett. 10:3206–3212. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang HH, Wang YC, Wu DW, Hung CS, Chen CY
and Lee H: Targeting insulin-like growth factor-binding protein-3
by microRNA-125b promotes tumor invasion and poor outcomes in
non-small-cell lung cancer. Tumour Biol.
39:10104283176943162017.PubMed/NCBI
|
|
32
|
Xiong B, Cheng Y, Ma L and Zhang C: MiR-21
regulates biological behavior thourgh the PTEN/PI-3K/Akt signalling
pathway in human colorectal cancer cells. Int J Oncol. 42:219–228.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Iwakawa HO and Tomari Y: The functions of
microRNAs: mRNA decay and translational respression. Trends Cell
Biol. 25:651–665. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kundu P, Fabian M, Sonenberg N,
Bhattacharyya SN and Filipowicz W: HuR protein attenuates
miRNA-mediated repression by promoting miRISC dissociation from the
target RNA. Nucleic Acids Res. 40:5088–5100. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Haydon AM, Macinnis RJ, English DR, Morris
H and Giles GG: Physical activity, insulin-like growth factor 1,
insulin-like growth factor binding protein 3, and survival from
colorectal cancer. Gut. 55:689–694. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Fu T, Pappou EP, Guzzetta AA, Mde Calmon
F, Sun L, Herrera A, Li F, Wolfgang CL, Baylin SB,
Iacobuzio-Donahue CA, et al: IGFBP-3 gene methylation in
primary tumor predicts recurrence of stage II colorectal cancers.
Ann Surg. 263:337–344. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Perez-Carbonell L, Balaguer F, Toiyama Y,
Egoavil C, Rojas E, Guarinos C, Andreu M, Llor X, Castells A, Jover
R, et al: IGFBP3 methylation is a novel diagnostic and
predictive biomarker in colorectal cancer. PLoS One. 9:e1042852014.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wolpin BM, Meyerhardt JA, Chan AT, Ng K,
Chan JA, Wu K, Pollak MN, Giovannucci EL and Fuchs CS: Insulin, the
insulin-like growth factor axis, and mortality in patients with
nonmetastatic colorectal cancer. J Clin Oncol. 27:176–185. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xue M, Fang Y, Sun G, Zhuo W, Zhong J,
Qian C, Wang L, Wang L, Si J and Chen S: IGFBP3, a transcriptional
target of homeobox D10, is correlated with the prognosis of gastric
cancer. PLoS One. 8:e814232013. View Article : Google Scholar : PubMed/NCBI
|