Introduction
Human genomic DNA is subject to multiple endogenous
and exogenous insults that include deamination, pyrimidine dimer,
mismatches, interstrand crosslink and DNA single or double strand
breaks (SSBs or DSBs), induced by free radical species, chemical
compounds, UV and ionizing radiation (IR) (1). Among many types of DNA lesions, DSBs
are the most lethal form and it is generally believed that a single
unrepaired DSB is sufficient to trigger cell death (2). In mammalian cells, DSBs are primarily
repaired by non-homologous end joining (NHEJ) and homologous
recombination (HR). HR repairs DSBs in an error-free fashion in
S/G2 phases when a sister chromatid is available to be used as a
repair template. NHEJ could be error prone but plays a central role
in repairing DSBs in all cell cycle stages other than the M phase
in dividing cells, and is particularly critical for DSB repair
(DSBR) in post-mitotic cells such as neurons, in which the
HR-mediated repair is not available (3). To ensure that DSBs are repaired prior
to initiation of cell division, DSBs induce cell cycle arrest,
which is known as DNA damage checkpoint activation. The cellular
DNA damage response (DDR) is a kinase-based signal transduction
pathway that involves multiple DSB sensor proteins such as an MRN
complex, transducer proteins such as ATM and ATR, mediator proteins
such as 53BP1 and BRCA1, and effectors such as Chk1 and Chk2, which
protect genome stability and integrity against DSB in a coordinated
way (4,5). Aberrant expression of DDR proteins has
been linked to the initiation and progression of human malignant
tumors. For example, overexpression of Rad51 has been linked to
pancreatic cancer and deficient BRCA1/BRCA2 has been linked to
breast cancer (6,7).
DDR could be a double-edged sword in normal vs.
cancer cells. DSB-inducing agents such as IR and radiomimetics is
an established therapeutic strategy for cancer, which is known as
radiotherapy (RT) and applied in at least 50% of all cancer
patients (8). While normal cells
rely on efficient DDR to maintain genome fidelity, DDR hampers the
RT response in cancer cells. Many cancers display resistance to
standardized irradiation due to intrinsic or acquired
radioresistance. For example, one of leading malignancies in women
worldwide, ~60% of cervical cancer cases are subjected to RT while
in an overall 13% of incidences local recurrence is observed due to
radioresistance (9). One of the
major mechanisms underlying radioresistance is increased DDR
activity and DSBR efficiency in those tumors (10). In addition, cancer stem cells that
survive fractionated irradiation can also be radioresistant and
cause tumor relapse (11). In order
to overcome radioresistance, decades of work has been focused on
the inhibition of DSB response as a viable strategy, and the link
between DDR inhibition and IR-induced cancer cell death has been
well demonstrated (12). Inhibitors
of DSBR factors and DNA damage checkpoint regulators, and a number
of synthetic inhibitors such as BO2 targeting Rad51, AZD7762
targeting Chk1/2, PCI-24781 targeting HDAC, and neutral
radiosensitizers extracted from plants such as quercetin targeting
ATM, genistein targeting cyclin B, have been evaluated (13,14).
However, compared with natural radiosensitizers, synthetic
inhibitors have limited improvement on treatment and more side
effects in general (14).
Medicinal plant extracts have long been known to be
beneficial towards human diseases, and the molecular
characterization of plant-based products is critical for the
discovery of new drug candidates (15). Sinomenium acutum is such a
plant that has been used to treat neuralgia and rheumatoid
arthritis in many Asian countries since antiquity (16). As the active ingredient of the
plant, alkaloid sinomenine (SIN) was subsequently isolated and the
pharmacological effects of SIN on anti-angiogenesis (17), analgesia (18), anti-inflammation (19,20),
immunosuppression (19,21) and anti-nociceptive (22) properties were demonstrated by
studies in vitro or in vivo. Notably, the anticancer
effects of SIN and its water-soluble form, sinomenine hydrochloride
(SH) were also characterized recently. Jiang et al found
that SIN induced apoptosis of a lung cancer cell line by collapsing
the mitochondrial membrane (23);
Lv et al found that SIN inhibited the proliferation of
gastric cancer cells by suppressing cyclooxygenase-2 expression
(24); Lu et al revealed
that SH inhibited hepatocellular carcinoma cell growth by involving
cell cycle arrest and apoptosis (25); Song et al reported that SIN
inhibited breast cancer cell invasion and migration by inactivating
NF-κB (26). However, its
radiosensitizing function in cancer treatment has never been
characterized.
In the present study, we evaluated the sensitizing
efficacy of SH on human cervical cancer cell line HeLa to
irradiation, and demonstrated its potential as a radiosensitizer on
a cellular level and in a mouse model.
Materials and methods
Cell cultures and preparation of
SH
Human HeLa cervical cancer cells and SiHa cervical
cancer cells were cultured with Dulbecco's modified Eagle's medium
(DMEM; HyClone Laboratories; GE Healthcare Life Sciences, Logan,
UT, USA) supplemented with 10% fetal bovine serum (FBS; HyClone
Laboratories; GE Healthcare Life Sciences), 100 U/ml penicillin and
100 µg/ml streptomycin. Cultures were grown in a 5% CO2
incubator at 37°C. SH (Zhengqing Pharmaceutical Group Co., Ltd.,
Hunan, China) was dissolved in phosphate-buffered saline (PBS) to a
concentration of 100 mmol/l, and stored at −20°C for up to 4
weeks.
Methylthiazoltetrazolium (MTT)
assay
HeLa cells were seeded in 96-well plates with a
density of 4,000 cells/well in 200 µl culture medium and incubated
overnight. SH solutions were prepared with DMEM without serum with
final gradient concentrations of 0.5, 1, 1.5, 2 and 5 mmol/l. After
the cells were incubated for 24, 48 and 72 h, 20 µl
3-(4,5-dimethylthiazol-2-y1)-2,5-diphenytetrazolium bromide was
added to each well and the cell cultures were incubated for an
additional 4 h. The colored solution was quantified by a
spectrophotometer at an absorbance of 490 nm. The inhibition rate
of the cells was then calculated.
Colony forming assay
HeLa and SiHa cells were incubated in 10
cm2 flasks overnight, and then divided into 4 groups:
Control, SH (1 mmol/l) alone, radiation alone, and SH combined with
radiation. Cells were treated with SH for 48 h, and then irradiated
by X-ray linear accelerator. Following IR, the medium containing SH
was removed and cells were maintained in normal culture medium. The
cell density of groups was: 300 cells for 0 Gy, 1,000 cells for 2
Gy, 2,000 cells for 4 Gy and 4,000 cells for 6 Gy. Fourteen days
later, the cells were washed and stained with crystal violet. The
colonies containing >50 cells were counted. Cell survival curves
were constructed.
Apoptosis and cell cycle assay
Apoptosis was quantitated using the KGI
Biotechnology Apoptosis Kit (Nanjing, China) following the
manufacturer's instructions. Cells were fixed with 70% ethanol (2
h, 4°C), and stained with propidium iodide and RNase A (30 min,
37°C) for cell cycle analysis. Samples with 10,000 cells/well were
used.
Immunofluorescence
We monitored the DNA DSBs and DSB repair capacity of
HeLa cells by phospho-H2AX foci immunofluorescence. The cells were
incubated in 10 cm2 flasks overnight and then divided
into 4 groups as aforementioned. Drugs were added 48 h prior to
radiation exposure (6 Gy). After IR, the medium containing SH was
removed and the cells were maintained in normal culturing medium.
Twenty-four hours later, the cells were fixed with PFA at 37°C for
20 min, treated with 0.02% Triton X-100 for 10 min, and then
blocked with a blocking buffer at room temperature for 1 h.
Subsequently, the cells were incubated with the phospho-H2AX S139
antibody (cat. no. 9718; Cell Signaling Technology, Inc., Danvers,
MA, USA) at a 1:1,000 dilution overnight at 4°C, and then with a
goat anti-rabbit IgG fluorescent-conjugated secondary antibody
(cat. no. 8889; Cell Signaling Technology, Inc.) at a 1:400
dilution for 2 h at room temperature. Then, DAPI was used for 15
min to stain the nuclei. The images of the γ-H2AX foci were
obtained by fluorescence microscopy (Olympus Corp., Tokyo,
Japan).
Western blotting and real-time PCR
analyses
The cells were lysed with RIPA lysis buffer (Pioneer
Biotech, Co., Ltd., Shaanxi, China) and the protein concentrations
were quantified using a BCA kit (Roche Applied Science, Penzberg,
Germany). The ATM antibody (cat. no. 2873), phospho-ATM Ser1981
antibody (cat. no. 5883), phospho-H2AX S139 antibody (cat. no.
9718), Ku80 antibody (cat. no. 2180), Rad50 antibody (cat. no.
3427), phospho-Chk2 Thr68 antibody (cat. no. 2661), phospho-Chk1
Ser345 antibody (cat. no. 2348) and anti-rabbit IgG HRP-linked
antibody (cat. no. 7074) were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA) and the dilution used were
1:1,000-1:3,000. Equal amounts of proteins (60 µg per lane) were
separated by 10% SDS-PAGE and transferred to polyvinylidene
difluoride (PVDF) membranes. The membranes were blocked with 5%
non-fat milk in Tris-buffered saline with 0.05% Tween-20 at pH 7.5
for 2 h, and then incubated with primary antibodies at 4°C
overnight, and secondary antibodies for 2 h at 37°C. The membranes
were washed 3 times with Tris-buffered saline with 0.05% Tween-20
and once with Tris-buffered saline. An enhanced chemiluminescence
detection kit (cat. no. WBKLS0500; Millipore, Burlington, MA, USA)
was used to detect the signals of immunoblotted proteins on an
JS-380A automatic gel imaging system (Quantity One Quantitation
software; Bio-Rad Laboratories, Inc., Hercules, CA, USA). RNA was
isolated from cells using RNAfast200 (Pioneer Biotech, Co., Ltd.).
The Ku80 primer/probe sets (F, 5’-ATTTGCTGGAGGACATTGAAAG-3′ and R,
5′-CTGAATCGGCTGCTGAGG-3′); the Rad51 primer/probe sets (F,
5′-CAACACAGACCACCAGACC-3′ and R, 5′-AGAAGCATCCGCAGAAACC-3′); and
the GAPDH primer/probe sets (F, 5′-AAGGCTGTGGGCAAGGTCATC-3′ and R,
5′-GCGTCAAAGGTGGAGGAGTGG-3′) were used. PrimeScript™ RT Master Mix
and the SYBR® Premix Ex Taq™ II were purchased from
Takara Biotechnology, Co., Ltd. (Dalian, China) and used for
real-time PCR. The optimal cycling conditions were 30 sec at 95°C,
and 40 cycles of 5 sec at 95°C, 30 sec at 60°C. All real-time PCR
experiments were performed in triplicate.
Comet assay
HeLa and SiHa cells were incubated in 6
cm2 flasks overnight, and then divided into 4 groups:
Control, SH (1 mmol/l) alone, radiation alone, and SH combined with
radiation for the neutral comet assay. HeLa cells were also divided
into 6 groups: Control, SH, H2O2 (0 min
post-H2O2 treatment for 1 h), SH combined
with H2O2 (0 min
post-H2O2 treatment for 1 h),
H2O2 (180 min post-H2O2
treatment for 1 h), SH combined with H2O2
(180 min post-H2O2 treatment for 1 h) for the
alkaline comet assay. Comet Assay kits (Trevigen, Inc.,
Gaithersburg, MD, USA) were used in the present study. Twenty-four
hours after IR, or 0 min/180 min after H2O2
treatment, the cells were harvested and mixed with agarose for
electrophoresis. The nuclear DNA was stained with SYBR-Green dye
for 10 min and the images were obtained by fluorescence microscopy.
The olive tail moment was assessed using Comet analysis software
(cat. no. 4260-000-CS; Trevigen, Inc.).
Dual-Luciferase reporter assay
The promoter region of the Rad51 gene from 543 bp
upstream to 204 bp downstream was cloned (primers used for PCR,
5′-CGGGGTACCGTCTCACTCTGTCATGAGGC-3′ and
5′-CCGCTCGAGGTCTAATTTGGGTCTTGACC-3′) into the pGL-Basic report
vector (27). Plasmids
pGL-Basic-Rad51-promoter and pRL-TK (as an internal control) were
co-transfected into 293T cells seeded in 12-well plates. The
luciferase activity was assessed using a Dual-Luciferase Assay Kit
(Promega Corp., Madison, WI, USA) 48 h after SH treatment. Reporter
luciferase activity was normalized to Renilla luciferase
activity.
In vivo test
The animal experiments were approved and supervised
by the Laboratory Animal Care Committee of Xi'an Jiaotong
University. Twenty-four mice were housed in sterile cages under
standard conditions (12 h light/dark cycles at 21±2°C and normal
atmosphere) with ad libitum access to disinfected food and
water. Twenty-four 4-week-old female BALB/c nude mice were injected
s.c. in the back with 1.0×106 HeLa cells. When
xenografts reached a volume of 150–200 mm3, they were
randomly divided into 4 groups adjusted by initial tumor volume and
treated with: i) PBS alone; ii) SH 100 mg/kg by intraperitoneal
injection (every day till the mice were sacrificed); iii)
irradiation (3 doses of local irradiation at a dose of 4 Gy at
2-day intervals); iv) SH combined with irradiation (IR starting
after 2 days of SH injections). Animals were weighed and the tumor
volume was assessed every 2 days until the mice were sacrificed 3
weeks later using pentobarbital sodium at a dose of 100 mg/kg. The
tumor volume was calculated by V = 1/2 (length ×
width2).
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 5 (GraphPad Software, Inc., La Jolla, CA, USA). Differences
between the control and treatment groups were determined by
Student's t-test and considered to be significant at P<0.05.
Results
SH sensitizes HeLa cells to
irradiation
Although SIN was previously implicated in cancer
cell proliferation suppression (23), the radiosensitive effect on cervical
cancer cells has not been characterized. To address this question,
first, we tested the effect of SH on HeLa cells by performing
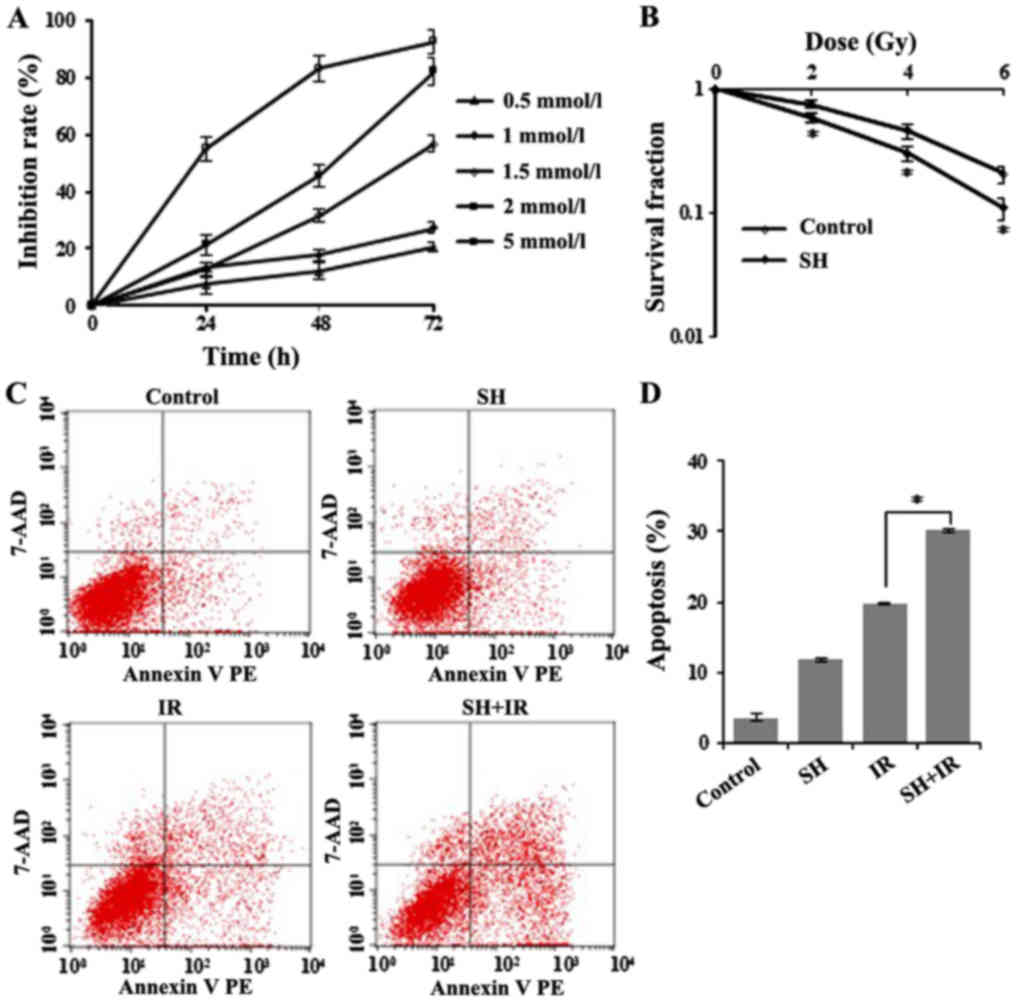
MTT-based cell viability analysis (Fig.
1A). The cell viability inhibition rate was calculated at 24,
48 and 72 h after SH treatment and as shown in Fig. 1A, cellular survival was
significantly inhibited by SH at various concentrations.
Concentrations of 1.5, 2 and 5 mM induced severe cell death (~30 to
90%), while 0.5 and 1 mM SH exhibited a similar and relatively
moderate effect on cell survival compared with higher
concentrations (Fig. 1A). Given
that 1 mM was an already established concentration in previously
published studies for a number of carcinoma cell lines (24,25,28),
we finally chose the 1 mM dose in the following experiments to make
it consistent with previous studies, thus enabling the comparison
of the SH-induced phenotypes across different cell lines and with
other studies. We next examined the radiosensitive effect of SH on
HeLa cells by clonogenic assay. HeLa cells were treated with 1 mM
SH for 48 h followed by IR with 0, 2, 4 or 6 Gy, and we observed a
significant reduction in clonogenic survival of the SH plus IR
group compared with IR alone, as illustrated by the survival curves
in Fig. 1B, which indicated a
sensitization of SH-treated HeLa cells to IR. We also examined
whether SH sensitized a squamous cell carcinoma cell line to IR. We
performed a clonogenic assay in SiHa cells and observed increased
sensitivity of SH-treated SiHa cells to IR (data now shown), which
was consistent with the observation in HeLa cells.
Furthermore, we conducted flow cytometric analysis
to test apoptosis in IR-treated HeLa cells incubated with or
without SH, and observed significantly increased apoptosis in the
group treated by IR in the presence of SH compared with the other
groups (Fig. 1C and D).
Collectively, these data demonstrated a role of SH in sensitizing
HeLa cells to IR.
SH treatment impairs DNA double-strand
break response in HeLa cells
Given that IR triggers cellular apoptosis by
inducing DNA damage, we tested whether SH enhances IR-induced DSB
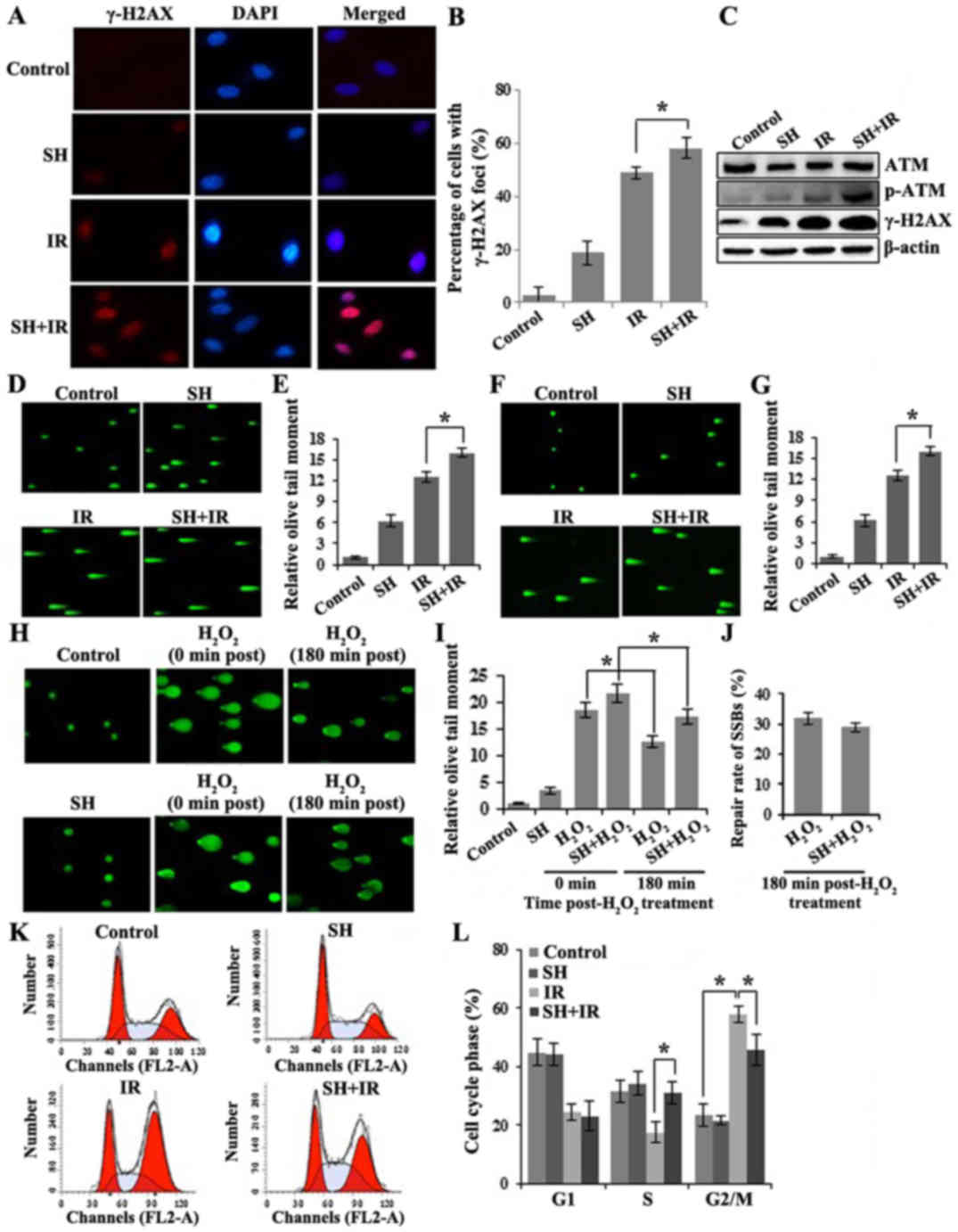
accumulation. We evaluated DSBs by assessing the level of γ-H2AX.
γ-H2AX is a phosphorylated core histone variant H2AX,
phosphorylated after DSB induction in an ATM-dependent manner and
quickly forms nuclear foci that can be visualized by
immunofluorescence (IF) assay (29,30).
It has been established that the γ-H2AX-containing foci are
correlated directly with the number of DSBs (31). Four groups of cells including one
untreated group as a control, and another 3 groups treated with SH,
IR or SH plus IR, respectively were fixed for IF following the
treatments. Cells with positive γ-H2AX foci were counted by
microscopy. The IR-induced DNA damage indicated by γ-H2AX foci was
significantly enhanced in the presence of SH (Fig. 2A and B). The results were further
confirmed by immunoblotting (IB). In addition to the γ-H2AX,
another DSB marker phospho-ATM (32) exhibited a similar pattern (Fig. 2C), indicating a persistent
accumulation of IR-induced DSBs in the presence of SH. We further
performed neutral single-cell gel electrophoresis assay (comet
assay), a method that exclusively visualizes cellular DSBs to
confirm the SH-mediated IR-induced DSB accumulation. The higher
mean comet tail moment in the SH plus IR group than the IR group,
indicated that more IR-induced DSBs were accumulated in HeLa cells
when pre-treated with SH (Fig. 2D and
E). We noticed that the DSBs were also slightly increased in
HeLa cells treated with SH alone, likely caused by the impaired
DSBR on endogenous DSBs. Similarly, more IR-induced DSBs were
accumulated in the SH pre-treated SiHa cells as well (Fig. 2F and G).
In addition, we investigated whether SH impaired DNA
single-strand break repair (SSBR). To reach this aim, we conducted
alkaline comet assays, which detect both DSB and SSB. We treated
HeLa cells with H2O2 to induce SSBs and
performed alkaline comet assays in untreated, SH,
H2O2 and SH combined with
H2O2-treated HeLa cells. In the SH-treated
group, we observed a slightly increased tail moment, likely due to
the DSBs that SH caused as we previously demonstrated.
H2O2-treated cells exhibited significantly
higher accumulation of SSBs as expected and ~30% of damages were
repaired after 3-h release from treatment (Fig. 2H and I). Although SH combined with
H2O2-treated cells exhibited a higher DNA
damage level than the H2O2 group, a
comparable repair rate (~30% after 3-h release) was observed in
both groups (Fig. 2J), which
indicated that the SSBR was not impaired by SH.
Cellular DNA damage activates cell cycle
checkpoints. We questioned whether SH alters IR-mediated DNA-damage
checkpoint activation. The cell cycle distribution was examined in
the 4 groups of HeLa cells by flow cytometry. As shown in Fig. 2K and L, in response to IR treatment,
we observed a reduced population in the S phase but a significant
enrichment population in the G2/M phase population, indicating the
activation of the G2/M checkpoint by IR. We did not observe any
changes in cell cycle distribution by SH incubation alone. However,
compared with the IR group, the SH plus IR group displayed a
markedly decreased cell population arrested in G2/M, which
indicated an interfering effect of SH on IR-induced G2/M checkpoint
activation. Notably, the IR-induced S phase reduction was rescued
by SH, which was comparable with the control (Fig. 2K and L). Overall, our results
indicated that SH caused DNA damage accumulation and interfered
with cell-cycle checkpoint activation.
SH suppresses expression of DSBR
protein Ku80 and Rad51 and enhances the IR-induced activition of
Chk1
As aforementioned, IR induces cancer cell death by
generating DSBs, which are mainly repaired by HR and NHEJ. As SH
sensitizes HeLa cells to IR via accumulation of IR-induced DSBs, we
wondered whether SH impairs DSBR pathways. We examined the
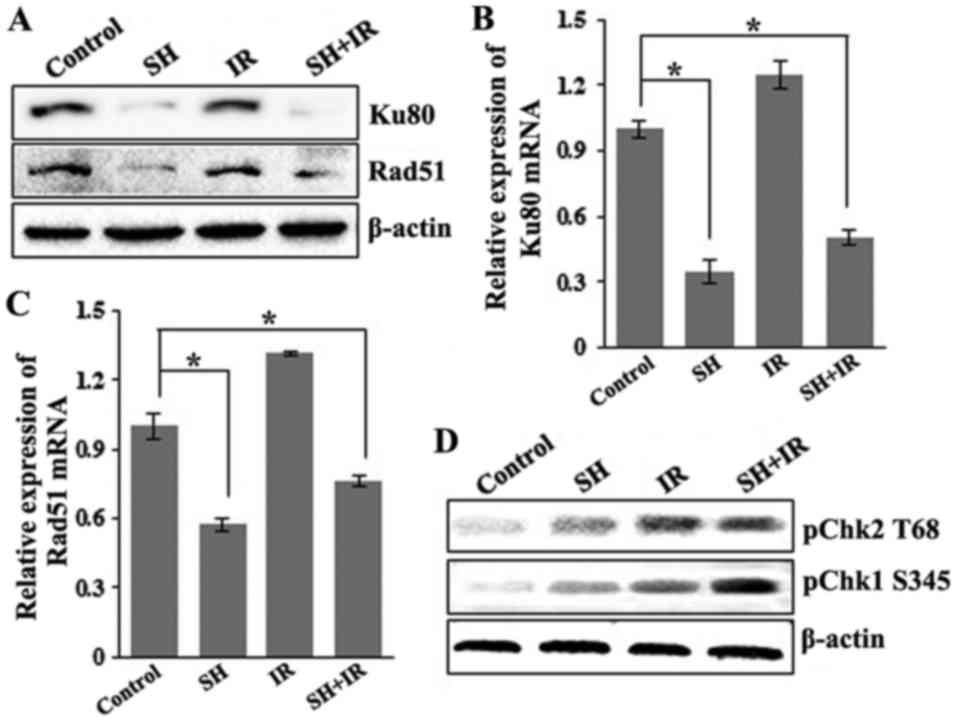
expression levels of a number of key DSBR factors including ATM,
BRCR1, 53BP1, Ku80 and Rad51 by IB, and found that the incubation
with SH resulted in a marked reduction in the levels of Ku80 and
Rad51 in HeLa cells (Fig. 3A).
Furthermore, we examined the mRNA levels of the two proteins by
performing RT-PCR and found that the mRNA levels of Ku80 and Rad51
were consistently downregulated in response to SH incubation
(Fig. 3B and C), indicating a
negative-regulatory role of SH on Ku80 and Rad51 at the
transcriptional level. We questioned whether the promotor activity
of the genes was affected by SH treatment. Given that the promotor
region of the Rad51 gene has been well characterized (27), we cloned Rad51 promoter fragment
(from 543 bp upstream to 204 bp downstream) into a pGL-Basic
luciferase reporter vector and performed luciferase reporter
assays. The results revealed that the promoter activity of the
Rad51 gene was significantly inhibited by incubation with SH (data
not shown). To explore the underlying mechanism by which
pre-treatment with SH promoted IR-dependent S-phase arrest as
displayed in Fig. 2K and L, we
detected the activation of Chk1 and Chk2, downstream targets of
ATR/ATM kinase, which is necessary for DNA damage checkpoint
signaling. Upon genotoxic injuries, Chk1 and Chk2 are
phosphorylated at Serine 345 (S345) and Threonine 68 (T68) by ATR
and ATM, respectively, facilitating G1/S, intra-S and G2/M cell
cycle checkpoint activition (33–37).
We thus assessed the phosphorylation level of Chk1 S345 and Chk2
T68 by western blotting and found that SH treatment specifically
increased IR-induced phosphorylation of Chk1 at S345 (Fig. 3D). Our data indicated that SH may
regulate the DSBR pathway by impairing the expression and
activition of key DDR proteins.
Radiosensitizing effect of SH in a
cervical cancer xenograft model
Our in-cellulo observations that SH sensitizes HeLa
cells and accumulates IR-induced DSBs by impairing DSBR proteins
Ku80 and Rad51, led us to evaluate its therapeutic role in RT of
cervical cancer, using a nude-mouse tumor xenograft model to
investigate SH-mediated radiosensitivity in vivo. HeLa cells
(1.0×106) were subcutaneously injected into 4-week
old-nude mice, which were randomly divided into 4 groups and
treated by PBS injection (control), SH injection, IR and SH
injection plus IR, respectively. In the IR and SH plus IR groups,
the mice received experimental fractionated radiotherapy (38) when xenografts reached a volume of
150–200 mm3 (39). The
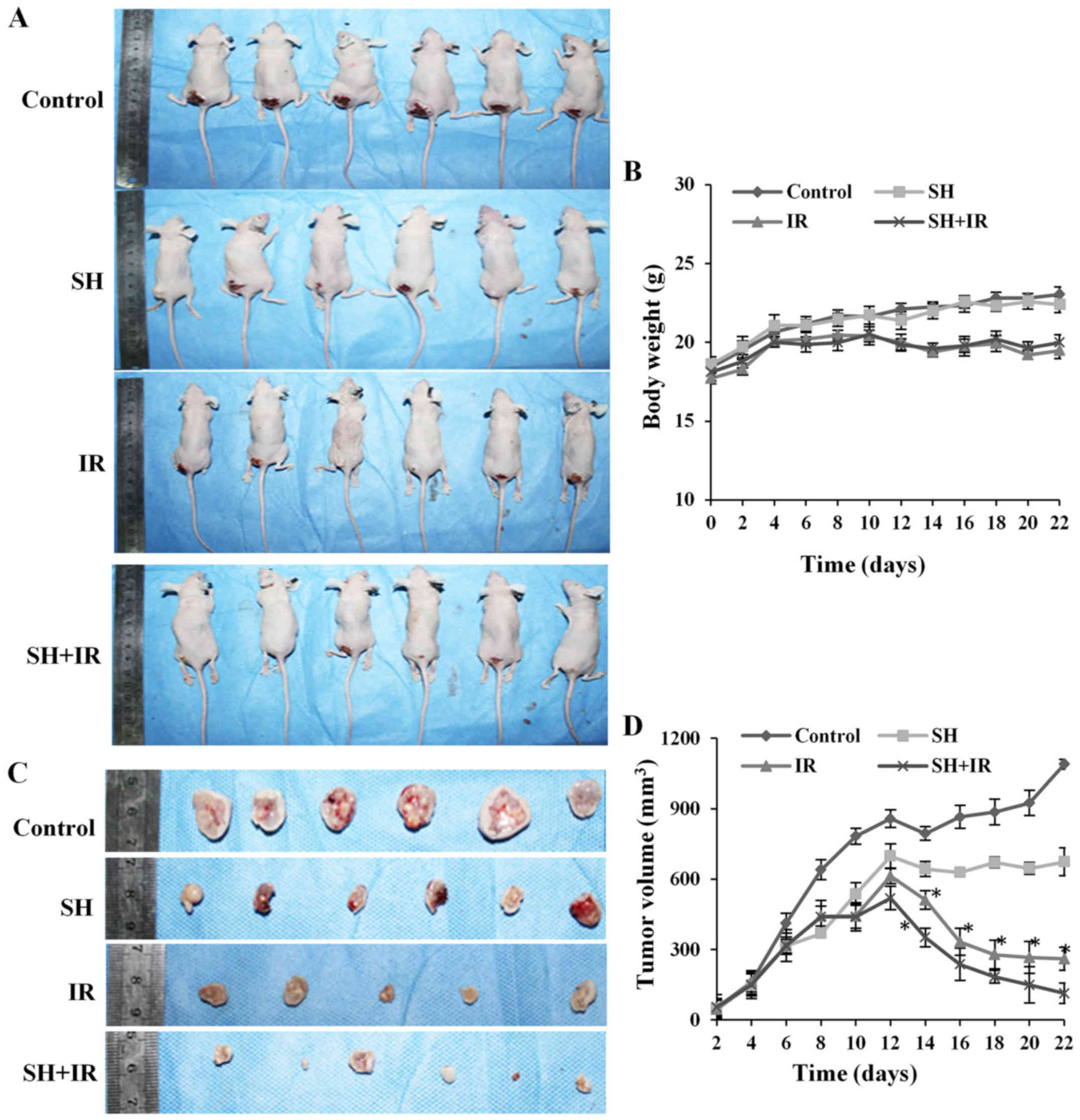
mice weight and tumor volume were assessed every 2 days and the
mice were sacrificed 3 weeks later. We did not observe any
significant difference in the weight of the mice between the
control group and SH injection group, while a marked reduced weight
was observed in the IR-treated groups compared with the
non-irradiated groups (Fig. 4A and
B), whereas the SH plus IR group exhibited a similar weight
pattern with the IR group (Fig. 4A and
B). The tumor volume was assessed every 2 days by vernier
calipers according to the formula: Volume = 1/2 (length ×
width2). As shown in Fig. 4C
and D, a large tumor was observed in the control group 3 weeks
after subcutaneous injection but the tumor was suppressed by IR,
and as anticipated, the suppression was significantly enhanced by
SH injection (Fig. 4C and D).
Collectively, the results demonstrated that SH injection enhanced
IR-induced inhibition of cervical xenograft growth, which indicated
that SH could be a potential radiosensitizer to RT for cervical
cancer.
Discussion
Cervical cancer is the second leading cause of
cancer-related deaths in women worldwide (40). Treatments for cervical cancer
include surgery, chemotherapy and RT. Compared to surgery,
chemotherapy and RT are mainly used in more advanced stages. RT
greatly benefits cervical cancer treatment whereas the survival
rate is largely limited by radioresistance developed in patients.
Multiple molecular signaling pathways including DDR have been
demonstrated to be associated with radioresistance in cervical
cancer (41–43). With the use of high throughput
screening, independent studies have also indicated DDR gene
expression pattern changes in cervical carcinoma tissues or cell
lines resisted to IR (41,44,45).
For example, radioresistant cervical cancer exhibits upregulated
NHEJ proteins (44), which sheds
light on the molecular mechanisms underlying DDR-related
radioresistance. In addition, recent efforts have been focused on
the identification/development and evaluation of radiosensitizers
for cervical cancer treatment by targeting DDR factors (46–50).
SIN that is found in the root of the climbing plant
Sinomenium acutum is an alkaloid used in traditional herbal
medicine in Asian countries, and its antitumor effects in various
types of cancers have been subsequently investigated. In the
present study, we demonstrated a novel role of SH benefiting
cervical cancer treatment as a radiosensitizer. The
radiosensitizing effects of SH were examined both in cells and in a
xenograft mouse model. In the mouse model, we developed a large
tumor through subcutaneous injection of HeLa cells and observed a
significant regression by IR. However, IR-induced tumor volume
reduction was enhanced by SH pre-treatment, which was consistent
with our in vitro experimental results that pre-treatment
with SH sensitizes HeLa cells to IR, and indicates it
radiosensitizing capacity. Mice treated with SH alone also
exhibited a lower tumor volume compared with the control, likely
due to its apoptotic effects that were documented in previous
publications.
Efficient DSBR is responsible for the RT failure and
rapid tumor cell recurrence, and targeting DDR is the key to
successful radiosensitization. We also reasoned that SH may impair
IR-induced DDR. We observed that HeLa cells pre-treated with SH
maintained significantly higher levels of IR-induced DSBs than
untreated cells, which was confirmed by either elevated DSB marker
levels or long moment tails as revealed by comet assays, likely due
to DSBR defects. In addition to HeLa, we also demonstrated the
SH-induced sensitivity to IR and DSBR deficiency in squamous cell
carcinoma cell line, SiHa. To explore the impairment of SH on DSBR,
we screened a number of key DDR proteins and finally, we found that
the expression of Rad51 and Ku80 were markedly inhibited by SH
incubation at the transcriptional level. Rad51 is a central factor
involved in HR-mediated DSBR. When mammalian cells are exposed to
genotoxic agents like irradiation, Rad51 is recruited to DSB sites
and mediates homologous sequence searching during HR. Rad51 has
been established as a radiosensitization target in multiple cancers
(51–56). A recent study revealed that Rad51
was highly expressed in glioblastoma stem cells, and inhibition of
Rad51 caused significant radiosensitization (51). Notably, the protein level of Rad51
has also been observed to be upregulated ~4 folds in cervical
cancer HeLa cells compared with primary cells (57), which indicates its therapeutic
applications as a sensitizer target to IR in cervical cancer
treatment. Similar to Rad51, Ku80 also plays a crucial role in DSBR
but through the NHEJ pathway. Ku80 is one subunit of the Ku80/Ku70
heterodimer, and the Ku heterodimer recruits key DDR proteins like
DNA-PK to the DSB sites when DSB occurs and facilitates the DNA
damage repair. A number of studies have suggested Ku80 as a
molecular target of radiosensitization (58–60).
For example, one study demonstrated that inhibition of Ku80
promotes IR combined cisplatin-induced HeLa cell death (58), and another study revealed the
relationship between Ku80 suppression and radiosensitivity in human
osteosarcoma cells (60). Based on
our results, we confirmed the cellular radiosensitization efficacy
of inhibiting Rad51 and Ku80 to IR, and identified the potential of
SH as a natural radiosensitizer by impairing both HR- and
NHEJ-mediated DSBR by inhibiting the two targets. Our observation
that SH inhibited promoter activity of the Rad51 gene provided an
insight into how SH impairs DSBR. However, why SH selectively
regulates the Rad51 and Ku80 genes remain unknown. One of the
possibilities is that SH alters histone modification, for example,
methylation that specifically regulates transcription of certain
genes (61), however further
investigation is warranted. In addition to the DSBR defects, SH
also disturbed DNA-damage checkpoint activation. Although our data
did not reveal any cell cycle disturbance by SH alone, we observed
a significantly decreased IR-induced G2/M arrest and increased
S-phase arrest by SH pre-incubation, likely due to the elevated
phosphorylation level of Chk1 S345, which is important for DNA
damage-mediated S-phase checkpoint activation (36).
Overall, our data indicated that SH contributed to
radiosensitization through dual pathways, DNA repair and cell cycle
checkpoint regulation. A comprehensive investigation of its
applications on clinic treatment as radiosensitizer will be a focus
of our future studies.
Acknowledgements
The authors thank the members of Han Laboratory in
the First Affiliated Hospital of Xi'an Jiaotong University (Xi'an,
China) for their technical support.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (nos. 81272488 and
81472795) to SH, (no. 81572968) to HW and NINDS-NIH R01 NS088645 to
MLH.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
DZ designed and performed the majority of the
experiments in the study and co-wrote the manuscript. YD and CZ
contributed to procuring the reagents/materials/analysis tools and
analyzed the data. YZ and YQ performed partial experiments. HW and
SH designed and supervised the study, and co-wrote and prepared the
final manuscript. MLH analyzed and interpreted the data and revised
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The animal experiments were approved and supervised
by the Laboratory Animal Care Committee of Xi'an Jiaotong
University (Shaanxi, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
SH
|
sinomenine hydrochloride
|
|
IR
|
ionizing radiation
|
|
DSBs
|
double-strand breaks
|
|
DDR
|
DNA damage response
|
|
NHEJ
|
non-homologous end joining
|
|
HR
|
homologous recombination
|
References
|
1
|
Helleday T, Eshtad S and Nik-Zainal S:
Mechanisms underlying mutational signatures in human cancers. Nat
Rev Genet. 15:585–598. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kanungo J: DNA-dependent protein kinase
and DNA repair: Relevance to Alzheimer's disease. Alzheimers Res
Ther. 5:132013. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Guerrero EN, Wang H, Mitra J, Hegde PM,
Stowell SE, Liachko NF, Kraemer BC, Garruto RM, Rao KS and Hegde
ML: TDP-43/FUS in motor neuron disease: Complexity and challenges.
Prog Neurobiol. 145–146:78–97. 2016. View Article : Google Scholar
|
|
4
|
Polo SE and Jackson SP: Dynamics of DNA
damage response proteins at DNA breaks: A focus on protein
modifications. Genes Dev. 25:409–433. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Freeman AK and Monteiro AN: Phosphatases
in the cellular response to DNA damage. Cell Commun Signal.
8:272010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Srivastava M and Raghavan SC: DNA
double-strand break repair inhibitors as cancer therapeutics. Chem
Biol. 22:17–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Davies H, Glodzik D, Morganella S, Yates
LR, Staaf J, Zou X, Ramakrishna M, Martin S, Boyault S, Sieuwerts
AM, et al: HRDetect is a predictor of BRCA1 and BRCA2
deficiency based on mutational signatures. Nat Med. 23:517–525.
2017. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Delaney G, Jacob S, Featherstone C and
Barton M: The role of radiotherapy in cancer treatment: Estimating
optimal utilization from a review of evidence-based clinical
guidelines. Cancer. 104:1129–1137. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang B, Chen J, Ren Z, Chen Y, Li J, Miao
X, Song Y, Zhao T, Li Y, Shi Y, et al: A specific miRNA signature
promotes radioresistance of human cervical cancer cells. Cancer
Cell Int. 13:1182013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Begg AC, Stewart FA and Vens C: Strategies
to improve radiotherapy with targeted drugs. Nat Rev Cancer.
11:239–253. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Krause M, Dubrovska A, Linge A and Baumann
M: Cancer stem cells: Radioresistance, prediction of radiotherapy
outcome and specific targets for combined treatments. Adv Drug
Deliv Rev. 109:63–73. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Thoms J and Bristow RG: DNA repair
targeting and radiotherapy: A focus on the therapeutic ratio. Semin
Radiat Oncol. 20:217–222. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mladenova V, Mladenov E and Iliakis G:
Novel biological approaches for testing the contributions of single
DSBs and DSB clusters to the biological effects of high LET
radiation. Front Oncol. 6:1632016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Malik A, Sultana M, Qazi A, Qazi MH,
Parveen G, Waquar S, Ashraf AB and Rasool M: Role of natural
radiosensitizers and cancer cell radioresistance: An update. Anal
Cell Pathol. 2016:61465952016. View Article : Google Scholar
|
|
15
|
Hegde ML: Molecular characterization of
neuroprotective activities of plant based products could revive
their utilization and lead discovery of new drug candidates for
brain diseases. J Pharm Bioallied Sci. 6:63–64. 2014.PubMed/NCBI
|
|
16
|
Yamasaki H: Pharmacology of sinomenine, an
anti-rheumatic alkaloid from sinomenium acutum. Acta Med Okayama.
30:1–20. 1976.PubMed/NCBI
|
|
17
|
Kok TW, Yue PY, Mak NK, Fan TP, Liu L and
Wong RN: The anti-angiogenic effect of sinomenine. Angiogenesis.
8:3–12. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ju XD, Deng M, Ao YF, Yu CL, Wang JQ, Yu
JK, Cui GQ and Hu YL: Protective effect of sinomenine on cartilage
degradation and chondrocytes apoptosis. Yakugaku Zasshi.
130:1053–1060. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Q and Li XK: Immunosuppressive and
anti-inflammatory activities of sinomenine. Int Immunopharmacol.
11:373–376. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Teng P, Liu HL, Zhang L, Feng LL, Huai Y,
Deng ZS, Sun Y, Xu Q and Li JX: Synthesis and biological evaluation
of novel sinomenine derivatives as anti-inflammatory agents. Eur J
Med Chem. 50:63–74. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shu L, Yin W, Zhang J, Tang B, Kang YX,
Ding F and Hua ZC: Sinomenine inhibits primary CD4+
T-cell proliferation via apoptosis. Cell Biol Int. 31:784–789.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhu Q, Sun Y, Zhu J, Fang T, Zhang W and
Li JX: Antinociceptive effects of sinomenine in a rat model of
neuropathic pain. Sci Rep. 4:72702014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiang T, Zhou L, Zhang W, Qu D, Xu X, Yang
Y and Li S: Effects of sinomenine on proliferation and apoptosis in
human lung cancer cell line NCI-H460 in vitro. Mol Med Rep.
3:51–56. 2010.PubMed/NCBI
|
|
24
|
Lv Y, Li C, Li S and Hao Z: Sinomenine
inhibits proliferation of SGC-7901 gastric adenocarcinoma cells via
suppression of cyclooxygenase-2 expression. Oncol Lett. 2:741–745.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lu XL, Zeng J, Chen YL, He PM, Wen MX, Ren
MD, Hu YN, Lu GF and He SX: Sinomenine hydrochloride inhibits human
hepatocellular carcinoma cell growth in vitro and in vivo:
Involvement of cell cycle arrest and apoptosis induction. Int J
Oncol. 42:229–238. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Song L, Liu D, Zhao Y, He J, Kang H, Dai
Z, Wang X, Zhang S and Zan Y: Sinomenine inhibits breast cancer
cell invasion and migration by suppressing NF-κB activation
mediated by IL-4/miR-324-5p/CUEDC2 axis. Biochem Biophys Res
Commun. 464:705–710. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hasselbach L, Haase S, Fischer D, Kolberg
HC and Sturzbecher HW: Characterisation of the promoter region of
the human DNA-repair gene rad51. Eur J Gynaecol Oncol. 26:589–598.
2005.PubMed/NCBI
|
|
28
|
Li X, Wang K, Ren Y, Zhang L, Tang XJ,
Zhang HM, Zhao CQ, Liu PJ, Zhang JM and He JJ: MAPK signaling
mediates sinomenine hydrochloride-induced human breast cancer cell
death via both reactive oxygen species-dependent and -independent
pathways: An in vitro and in vivo study. Cell Death Dis.
5:e13562014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rogakou EP, Pilch DR, Orr AH, Ivanova VS
and Bonner WM: DNA double-stranded breaks induce histone H2AX
phosphorylation on serine 139. J Biol Chem. 273:5858–5868. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang H, Adhikari S, Butler BE, Pandita TK,
Mitra S and Hegde ML: A perspective on chromosomal double strand
break markers in mammalian cells. Jacobs J Radiat Oncol.
1:0032014.PubMed/NCBI
|
|
31
|
Sancar A, Lindsey-Boltz LA, Unsal-Kacmaz K
and Linn S: Molecular mechanisms of mammalian DNA repair and the
DNA damage checkpoints. Annu Rev Biochem. 73:39–85. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang Y, Cheng J, Li D, Duan H, Yang H, Bin
P, Dai Y, Huang C, Liang X, Leng S, et al: Modulation of DNA repair
capacity by ataxia telangiectasia mutated gene polymorphisms among
polycyclic aromatic hydrocarbons-exposed studyers. Toxicol Sci.
124:99–108. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wilson KA and Stern DF: NFBD1/MDC1, 53BP1
and BRCA1 have both redundant and unique roles in the ATM pathway.
Cell Cycle. 7:3584–3594. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Carlessi L, Buscemi G, Fontanella E and
Delia D: A protein phosphatase feedback mechanism regulates the
basal phosphorylation of Chk2 kinase in the absence of DNA damage.
Biochim Biophys Acta. 1803:1213–1223. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Choi KS, Kim JY, Lim SK, Choi YW, Kim YH,
Kang SY, Park TJ and Lim IK: TIS21/BTG2/PC3 accelerates
the repair of DNA double strand breaks by enhancing Mre11
methylation and blocking damage signal transfer to the
Chk2T68-p53S20 pathway. DNA Repair (Amst).
11:965–975. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sorensen CS, Syljuasen RG, Falck J,
Schroeder T, Ronnstrand L, Khanna KK, Zhou BB, Bartek J and Lukas
J: Chk1 regulates the S phase checkpoint by coupling the
physiological turnover and ionizing radiation-induced accelerated
proteolysis of Cdc25A. Cancer Cell. 3:247–258. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gamper AM, Rofougaran R, Watkins SC,
Greenberger JS, Beumer JH and Bakkenist CJ: ATR kinase activation
in G1 phase facilitates the repair of ionizing radiation-induced
DNA damage. Nucleic Acids Res. 41:10334–10344. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang H, Li LW, Shi M, Wang JH, Xiao F,
Zhou B, Diao LQ, Long XL, Liu XL and Xu L: In vivo study of breast
carcinoma radiosensitization by targeting eIF4E. Biochem Biophys
Res Commun. 423:878–883. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kim JS, Amorino GP, Pyo H, Cao Q, Price JO
and Choy H: The novel taxane analogs, BMS-184476 and BMS-188797,
potentiate the effects of radiation therapy in vitro and in vivo
against human lung cancer cells. Int J Radiat Oncol Biol Phys.
51:525–534. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Torre LA, Islami F, Siegel RL, Ward EM and
Jemal A: Global cancer in women: Burden and trends. Cancer
Epidemiol Biomarkers Prev. 26:444–457. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kitahara O, Katagiri T, Tsunoda T, Harima
Y and Nakamura Y: Classification of sensitivity or resistance of
cervical cancers to ionizing radiation according to expression
profiles of 62 genes selected by cDNA microarray analysis.
Neoplasia. 4:295–303. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Qin C, Chen X, Bai Q, Davis MR and Fang Y:
Factors associated with radiosensitivity of cervical cancer.
Anticancer Res. 34:4649–4656. 2014.PubMed/NCBI
|
|
43
|
Jeon YT, Song YC, Kim SH, Wu HG, Kim IH,
Park IA, Kim JW, Park NH, Kang SB, Lee HP, et al: Influences of
cyclooxygenase-1 and −2 expression on the radiosensitivities of
human cervical cancer cell lines. Cancer Lett. 256:33–38. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Beskow C, Skikuniene J, Holgersson A,
Nilsson B, Lewensohn R, Kanter L and Viktorsson K: Radioresistant
cervical cancer shows upregulation of the NHEJ proteins DNA-PKcs,
Ku70 and Ku86. Br J Cancer. 101:816–821. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Qing Y, Yang XQ, Zhong ZY, Lei X, Xie JY,
Li MX, Xiang DB, Li ZP, Yang ZZ, Wang G, et al: Microarray analysis
of DNA damage repair gene expression profiles in cervical cancer
cells radioresistant to 252Cf neutron and X-rays. BMC
Cancer. 10:712010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kunos CA, Colussi VC, Pink J,
Radivoyevitch T and Oleinick NL: Radiosensitization of human
cervical cancer cells by inhibiting ribonucleotide reductase:
Enhanced radiation response at low-dose rates. Int J Radiat Oncol
Biol Phys. 80:1198–1204. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wieringa HW, van der Zee AG, de Vries EG
and van Vugt MA: Breaking the DNA damage response to improve
cervical cancer treatment. Cancer Treat Rev. 42:30–40. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Mountzios G, Soultati A, Pectasides D,
Dimopoulos MA and Papadimitriou CA: Novel approaches for concurrent
irradiation in locally advanced cervical cancer: Platinum
combinations, non-platinum-containing regimens, and molecular
targeted agents. Obstet Gynecol Int. 2013:5367652013.PubMed/NCBI
|
|
49
|
Candelaria M, Garcia-Arias A, Cetina L and
Duenas-Gonzalez A: Radiosensitizers in cervical cancer. Cisplatin
and beyond. Radiat Oncol. 1:152006. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Fuhrman CB, Kilgore J, LaCoursiere YD, Lee
CM, Milash BA, Soisson AP and Zempolich KA: Radiosensitization of
cervical cancer cells via double-strand DNA break repair
inhibition. Gynecol Oncol. 110:93–98. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
King HO, Brend T, Payne HL, Wright A, Ward
TA, Patel K, Egnuni T, Stead LF, Patel A, Wurdak H, et al: RAD51 is
a selective DNA repair target to radiosensitize glioma stem cells.
Stem Cell Reports. 8:125–139. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Balbous A, Cortes U, Guilloteau K, Rivet
P, Pinel B, Duchesne M, Godet J, Boissonnade O, Wager M, Bensadoun
RJ, et al: A radiosensitizing effect of RAD51 inhibition in
glioblastoma stem-like cells. BMC Cancer. 16:6042016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Chen X, Wong P, Radany EH, Stark JM,
Laulier C and Wong JY: Suberoylanilide hydroxamic acid as a
radiosensitizer through modulation of RAD51 protein and inhibition
of homology-directed repair in multiple myeloma. Mol Cancer Res.
10:1052–1064. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kobashigawa S, Morikawa K, Mori H and
Kashino G: Gemcitabine induces radiosensitization through
inhibition of RAD51-dependent repair for DNA double-strand breaks.
Anticancer Res. 35:2731–2737. 2015.PubMed/NCBI
|
|
55
|
Liu Q, Jiang H, Liu Z, Wang Y, Zhao M, Hao
C, Feng S, Guo H, Xu B, Yang Q, et al: Berberine radiosensitizes
human esophageal cancer cells by downregulating homologous
recombination repair protein RAD51. PLoS One. 6:e234272011.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Gasparini P, Lovat F, Fassan M, Casadei L,
Cascione L, Jacob NK, Carasi S, Palmieri D, Costinean S, Shapiro
CL, et al: Protective role of miR-155 in breast cancer through
RAD51 targeting impairs homologous recombination after
irradiation. Proc Natl Acad Sci USA. 111:4536–4541. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Raderschall E, Stout K, Freier S, Suckow
V, Schweiger S and Haaf T: Elevated levels of Rad51 recombination
protein in tumor cells. Cancer Res. 62:219–225. 2002.PubMed/NCBI
|
|
58
|
Zhuang L, Liu F, Peng P, Xiong H, Qiu H,
Fu X, Xiao Z and Huang X: Effect of Ku80 on the radiosensitization
of cisplatin in the cervical carcinoma cell line HeLa. Oncol Lett.
15:147–154. 2018.PubMed/NCBI
|
|
59
|
Petera J, Sirak I, Beranek M, Vosmik M,
Drastikova M, Paulikova S and Soumarova R: Molecular predictive
factors of outcome of radiotherapy in cervical cancer. Neoplasma.
58:469–475. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Hu L, Wu QQ, Wang WB, Jiang HG, Yang L,
Liu Y, Yu HJ, Xie CH, Zhou YF and Zhou FX: Suppression of Ku80
correlates with radiosensitivity and telomere shortening in the
U2OS telomerase-negative osteosarcoma cell line. Asian Pac J Cancer
Prev. 14:795–799. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zhang Y and Reinberg D: Transcription
regulation by histone methylation: Interplay between different
covalent modifications of the core histone tails. Genes Dev.
15:2343–2360. 2001. View Article : Google Scholar : PubMed/NCBI
|