Introduction
Colorectal cancer is the third most common cancer in
the world and its annual fatality rate ranks third (1,2). The
main reason leading to the death of patients with colorectal cancer
is distant invasion and metastasis such as liver, lung and bone
metastasis, while liver metastasis of colorectal cancer is the most
common (3). Although the diagnosis
and treatment strategies for liver metastasis of colorectal cancer
have greatly improved in recent years, to date, the overall
survival rate of patients has not significantly improved.
Therefore, exploring the mechanism of liver metastasis of
colorectal cancer can provide effective guidance for potential
clinical interventions, thus improving the long-term prognosis of
patients.
PEA15 is a 15-kDa-sized small molecule protein
containing an NH2 terminal death effector domain and a COOH
terminal irregular structure distributed in the cytoplasm. Its DNA
sequence is located on chromosome 1q21-22 and it is highly
conserved in mammalian gene sequences (4,5).
PEA-15 is widely expressed in human tissues and participates in the
interaction between various proteins. It can regulate cell
apoptosis, proliferation and glucose metabolism by acting on the
key functions of cellular effectors in vivo (6–8). At
present, it has been revealed that PEA-15, through its
death-effector domain (DED) and ERK1/2-targeted binding, inhibited
ERK1/2 phosphorylation and nucleus transfer, thus playing an
anti-apoptotic role (9). Some
studies have demonstrated that PEA15 is highly expressed in tumors
such as liver (10), lung (11), breast (12) and ovarian cancer (13). However, it has a dual role in the
regulation of tumors (14). It has
been revealed that phosphorylation of PEA15 phosphorylated ERK and
promoted the proliferation and invasion of hepatocellular carcinoma
cells (10), whereas the
unphosphorylated state of PEA15 inhibited the phosphorylation of
ERK and EGFR, thus inhibiting proliferation, invasion and
metastasis of breast (12) and
ovarian cancer (13). However,
whether PEA15 plays a role in liver metastasis of colorectal cancer
is still unknown.
In the present study, we revealed that PEA15 was
highly expressed in colorectal cancer tissues and metastatic liver
tissues. High expression of PEA15 was positively correlated with
TNM stage, liver metastasis and indicated a poor prognosis. In
vitro and in vivo experiments confirmed that PEA15
promoted the proliferation, invasion, migration and EMT of
colorectal cancer cells. To determine its mechanism, we used gene
chip analysis and found that PEA15 regulated the ERK/MAPK signaling
pathway to promote the invasion and migration of colorectal cancer
cells. Concurrently, xenograft tumor experiments also confirmed
that PEA15 promoted liver metastasis of colorectal cancer cells
in vivo.
Materials and methods
Experimental reagents and
antibodies
Lipofectamine 2000 liposomes and TRIzol reagent were
purchased from Invitrogen (Thermo Fisher Scientific, Inc., Waltham,
MA, USA). Primary antibodies for PEA15 (cat. no. ab133217), PLD1
(cat. no. ab50695), ERK (cat. no. ab184699), p38 (cat. no.
ab170099), MAP2K6 (cat. no. ab154901), JNK (cat. no. ab47337), PRKX
(cat. no. ab109389) and FLNB (cat. no. ab97457) proteins were
purchased from Abcam (Cambridge, MA, USA). Primary antibodies for
E-cadherin (cat. no. #14472), N-cadherin (cat. no. 13116), vimentin
(cat. no. 5741) and β-actin (cat. no. 3700) were purchased from
Cell Signaling Technology (Danvers, MA, USA). Other reagents were
purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
Patient and samples
The present study included 40 cases of colorectal
cancer and corresponding clinicopathological data, which contained
19 cases of liver metastasis tissue specimens. These specimens were
all paraffin specimens and obtained from the Department of General
Surgery of the Affiliated Hospital of Guilin Medical University
from August 2000 to August 2016 and all the specimens were
clinically diagnosed by the Department of Pathology. All clinical
cases and specimen tissues were reviewed and approved by the Ethics
Committee of Guilin Medical University, and informed consents were
signed by the patients according to the Declaration of Helsinki.
Clinicopathological data included sex, age, tumor diameter, tumor
differentiation, TNM staging and liver metastasis. Follow-up of
patients was performed until March 2018.
Immunohistochemistry
The surgically resected paraffin specimens were
collected from the Department of Pathology of the Affiliated
Hospital of Guilin Medical University and were prepared into 4-mm
thick slices. According to the EnVision two-step procedure
(14), dewaxing and hydration were
carried out, after EDTA buffer high pressure antigen retrieval.
Subsequently a primary antibody against PEA15 (Abcam) with a
dilution of 1:100 was added, and the specimens were incubated at
room temperature for 1 h, then washed and incubated at room
temperature for 0.5 h with a secondary antibody (MaxVision™
HRP-polymer anti-Rabbit IHC Kit, cat. no. KIT-5006, MXB
Biotechnologies, Fuzhou, China) followed by washing with
phosphate-buffered saline (PBS) buffer. A DAB Horseradish
Peroxidase Color Development kit (Beyotime Institute of
Biotechnology, Haimen, China) was used for color development and
scored using the Olympus X71 inverted microscope (Olympus Corp.,
Tokyo, Japan). Based on the staining intensity, samples were
divided into the following grades: 0, <10% of tumor cells were
positively stained; 1+, 11–25% of tumor cells were positively
stained; 2+, 26–50% of tumor cells were positively stained; and 3+,
>50% of tumor cells were positively stained. Immunohistochemical
analysis and scoring were conducted by two researchers,
respectively.
Western blotting
Cell samples were collected and lysis buffer [50 mM
Tris-HCl, 137 mM NaCl, 10% glycerol, 100 mM sodium orthovanadate, 1
mM phenylmethylsulfonyl fluoride (PMSF), 10 mg/ml aprotinin, 10
mg/ml leupeptin, 1% Nonidet P-40, and 5 mM protease inhibitor
cocktail; pH 7.4] was added to extract total proteins. Then the
protein concentration was assessed by BCA (bicinchoninic acid) kit
(Beyotime Institute of Biotechnology) and the appropriate amount of
loading buffer was added. Protein sample (30 µg) was added to 10%
sodium dodecyl sulfate-polyacrylamide gel electrophoresis system
(SDS-PAGE) for electrophoresis separation and then transferred to
polyvinylidene difluoride (PVDF) membranes (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). Then, 5% non-fat milk was used to
incubate the PVDF membranes, and peroxidase-labeled secondary
antibodies (HRP-labeled goat anti-rabbit IgG; cat. no. A0208;
Beyotime Institute of Biotechnolgy) at a 1:10,000 were used for
incubation for 1 h at room temperature. Subsequently,
chemiluminescence was used to display the imprinting, and the
results were analyzed using the Tannon 5200 chemiluminescent
imaging system (Tanon Science and Technology, Shanghai, China).
Quantitative reverse transcription
polymerase chain reaction (qRT-PCR)
The collected cells and tissues were added to TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.) and total RNA was
extracted according to the RNAiso Plus kit instructions (Takara
Bio, Inc., Otsu, Japan). The RNA concentration was assessed using
the NanoDrop spectrophotometer (Thermo Fisher Scientific, Inc.).
The FastQuant cDNA First-Strand Synthesis kit (Tiangen Biotech,
Co., Ltd., Beijing, China) was used for reverse transcription. A
qRT-PCR kit FastStart Universal SYBR Green Master Mix (Rox) (Roche
Diagnostics GmbH, Mannheim, Germany) was used to produce the
reaction system and amplification according to the manufacturer's
instructions. The reaction cycling conditions were performed as
follows: 1 cycle at 95°C for 15 min, followed by 40 cycles at 95°C
for 10 sec and at 60°C for 60 sec, and then relative mRNA
expression was analyzed using the comparative quantification cycle
(Cq) method followed by normalization to β-actin
expression (Applied Biosystems; Thermo Fisher Scientific, Inc.)
(15). The primers used for
amplification were as follows: PEA15 forward,
5′-GTTCTGTAGTCAACCACCAT-3′ and reverse, 5′-ACCAACAACATCACCCTT-3′;
β-actin forward, 5′-TCTTCATGGTGCTGGGAG-3′) and reverse,
5′-AATGAGCGGTTCCGTTGC-3′ which was used as the internal
control.
Cell culture and cell
transfection
Colorectal cancer cells (HT-29, Caco-2, COLO 205,
HCT-116, RKO, SW-620 and SW-480) were purchased from the Cell Bank
of Chinese Academy of Sciences (Shanghai, China). Dulbecco's
modified Eagle's medium (DMEM; Life Technologies; Thermo Fisher
Scientific, Inc.) was used for HT-29, Caco-2, HCT-116 and RKO and
Roswell Park Memorial Institute-1640 medium (RPMI-1640) (Life
Technologies; Thermo Fisher Scientific, Inc.) was used for COLO
205, SW-620 and SW-480. All cell culture media contained 10% fetal
calf serum (FCS; Life Technologies; Thermo Fisher Scientific, Inc.)
and were cultured in a 37°C incubator with 5% CO2/95%
air. The procedure aforementioned was used for the preparation and
transfection of ERK1/2 siRNA and ERK1/2 plasmid (16). For primary colorectal cancer cells,
16 colorectal cancer specimens (aged 35–70 years) were collected
from patients who underwent colorectal cancer resection at the
Affiliated Hospital of Guilin Medical University from October 2014
to October 2016. Specimens were obtained with the approval of the
Medical Ethics Committee of the Affiliated Hospital of Guilin
Medical University. Fresh tumor tissue was cut and incubated with
type IV collagenase (Sigma-Aldrich; Merck KGaA) for 5–10 min at
37°C and a single cell suspension was filtered using a 100-mm cell
strainer (BD Biosciences, Franklin Lakes, NJ, USA). Cell viability
was detected via staining using Trypan blue (Sigma-Aldrich; Merck
KGaA), and CD45+ cells in fresh tumor tissue were
removed using the CD45 removal kit (Depletion kit; Miltenyi Biotec,
Bergisch Gladbach, Germany).
Cell proliferation experiments
The prepared cell suspension was seeded in a 96-well
plate (100 µl/well) with a cell density of 2×103
cells/well, and 10 µl of Cell Counting kit-8 (CCK-8) solution
(Dojindo Laboratories, Kumamoto, Japan) was added to each well
after the predefined different culture time. The plates were placed
in an incubator for ~1 h and then placed in a microplate reader
(SpectraMax plus 384; Molecular Devices, San Jose, CA, USA) to
assess the absorbance at 450 nm.
Plate cloning experiment
The prepared cell suspension was seeded on a 6-well
plate at 600 cells/well. Cell colonies were formed after being
cultured for ~2 weeks. Cells were fixed with 4% paraformaldehyde
for 20 min and stained overnight with 1% crystal violet (cat. no.
G1062; Solarbio, Tokyo, Japan). After three washes in PBS, the
images of the results were observed and captured using a
fluorescence microscope (IX71; Olympus Corporation) and the number
of colonies/well was calculated using ImageJ (National Institutes
of Health, Bethesda, MD, USA).
Experiments of cell invasion and
migration
A Transwell chamber (8 µm; BD Biosciences) (the
chamber for invasion detection was coated with Matrigel, and the
chamber for migration detection was without Matrigel) was placed
into a 24-well plate with ~600 µl medium containing 10% FBS which
was added to the bottom chamber. The prepared cell suspension was
seeded in a Τranswell chamber at 1,000 cells/well. One or two days
later, the cell preparation in the small chamber was observed under
a fluorescence microscope (IX71; Olympus Corporation). After an
appropriate number of cells had passed through the cell pores, the
cells were fixed with 4% paraformaldehyde and stained with 1%
crystal violet (cat. no. G1062; Solarbio) and images were
captured.
Confocal immunofluorescence
microscopy
Cells were seeded on a special slide (Costar,
Manassas, VA, USA). After 24 h, the cells on the slides were washed
with PBS and 4% paraformaldehyde was added to fix the cells. After
the cell membrane was permeabilized with 0.5% Triton X-100, the
cells were blocked by adding 10% FBS for 30 min at room
temperature. The primary antibody was added for incubation
overnight at 4°C, and then the cells were washed three times with
PBS, followed by the addition of a fluorescein isothiocyanate
(FITC)-labeled secondary antibody (cat. no. A-21063; Invitrogen;
Thermo Fisher Scientific, Inc.) and incubation for 1 h in a dark
chamber. After washing three times with PBS, the cells were stained
with DAPI for 5 min and analyzed with the Carl Zeiss Confocal
Imaging System (LSM 780; Carl Zeiss, Jena, Germany).
Detection of colorectal cancer stem
cell characteristics
A prepared primary colorectal cancer single cell
suspension was counted (Luna II; Logos Biosystems, Anyang, Korea),
and 1×106 cells were added into tumor stem cell culture
medium suspension for culture, and shaken 1 time every 6 h on the
1st and 2nd days to prevent cell adherence. When the cell spheres
became larger and the brightness decreased, the structure was
loosened. Accutase enzyme (Thermo Fisher Scientific, Inc.) was used
for digestion passage. The parental cells and the passaged cells
were cultured in a 6-well plate at 1×105 cells/well for
approximately 6 days, the conglomeration rate was calculated. The
surface antigen (EPCAM and CD133) of the primary colorectal cancer
cells was sorted by flow cytometry, and the proportion of side
population cells was counted and analyzed.
Construction of cell lines of stably
upregulated and downregulated PEA15
Construct retroviruses were coated with vector
pBabe-puro-PEA15 and with vector pSuper-retro- puro-PEA15-shRNA.
The extraction of retroviruses and transfection of tumor cells were
performed as previously described (17) and the expression of PEA15 was
verified using qRT-PCR and western blotting.
In vivo tumor growth and
metastasis
Forty BALB/c 8-week-old male nude mice weighing 18
g-20 g (20 were used for tumor growth while the other 20 were used
for a tumor metastasis model) were purchased from the Animal
Experimental Center of Guilin Medical University and all animals
were used in accordance with the operating instructions and
approved by the Animal Care Committee of Guilin Medical University,
with room temperature atmosphere and ad libitum. Tumor cell lines
in logarithmic growth phase were collected and washed with PBS to
prepare a cell suspension. The prepared cell suspension
(2×106 cells/ml) was subcutaneously injected into the
right groin area of 8-week-old nude mice. The tumor volume was
assessed weekly. Nude mice were sacrificed by cervical dislocation.
Twenty-eight days after surgery, the tumors were excised and the
tumor weights were recorded. For the tumor metastasis model, the
cells were collected and washed with PBS and then prepared into a
cell suspension. The cell suspension with a concentration of
1×107 cells/ml was injected into the hepatic portal vein
of nude mice. All mice were sacrificed by cervical dislocation 60
days after inoculation.
Gene chip analysis
The quality of total RNA was assessed using the
Agilent 2100 Bioanalyzer (Agilent Technologies, Inc., Santa Clara,
CA, USA) and the NanoDrop ND-1000 spectrophotometer. RNA expression
was then analyzed using the Affymetrix HU U133 plus 2.0 array
according to the manufacturer's instructions. The raw data was
normalized by the Robust Multiarray Average (RMA) of expression
control platform software (Affymetrix; Thermo Fisher Scientific,
Inc.). Scatter plot analysis of the gene with significant changes
in PEA15 in the downregulation group and the control group to
screen the genes with upregulation or downregulation of expression
≥3 times was performed. Gene Cluster v3.0 software cluster
(Stanford University, Stanford, CA, USA) was used for cluster
analysis, and Java TreeView v1.1.4r3 software (Alok Saldanha;
http://jtreeview.sourceforge.net/) was
used for thermal image visualization.
Statistical analysis
Experimental data were statistically analyzed using
GraphPad Prism 5 (GraphPad Software, Inc., San Diego, CA, USA) and
SPSS v.18.0 (SPSS Inc., Chicago, IL, USA). The Chi-square test
(χ2) was used to evaluate the association between the
expression of PEA15 and the clinicopathological features of the
patients. Comparisons between different groups were undertaken
using the Student's two-tailed t-test. Correlation between PEA15
expression with BRAF and RAS mutation in colon tissues were
assessed using Spearman's rank correlation test. Multigroup
comparisons of the means were carried out by one-way analysis of
variance (ANOVA) test with post hoc contrasts by
Student-Newman-Keuls test. The Kaplan-Meier method was used to
estimate the correlation between the expression of PEA15 and the
5-year survival rate of patients after diagnosis. The log-rank test
was used to compare the survival curves of different groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
PEA15 is highly expressed in
colorectal cancer and metastatic liver cancer tissues and
positively correlated with tumor TNM stage
We randomly collected 40 cases of colorectal cancer
and the corresponding liver metastasis tissues with
immunohistochemical detection. The results revealed that the
expression level of PEA15 in colorectal cancer and metastatic liver
cancer tissues was significantly higher than the corresponding
paracancerous tissues. In addition, the expression level of PEA15
in liver cancer tissues was significantly higher than that in
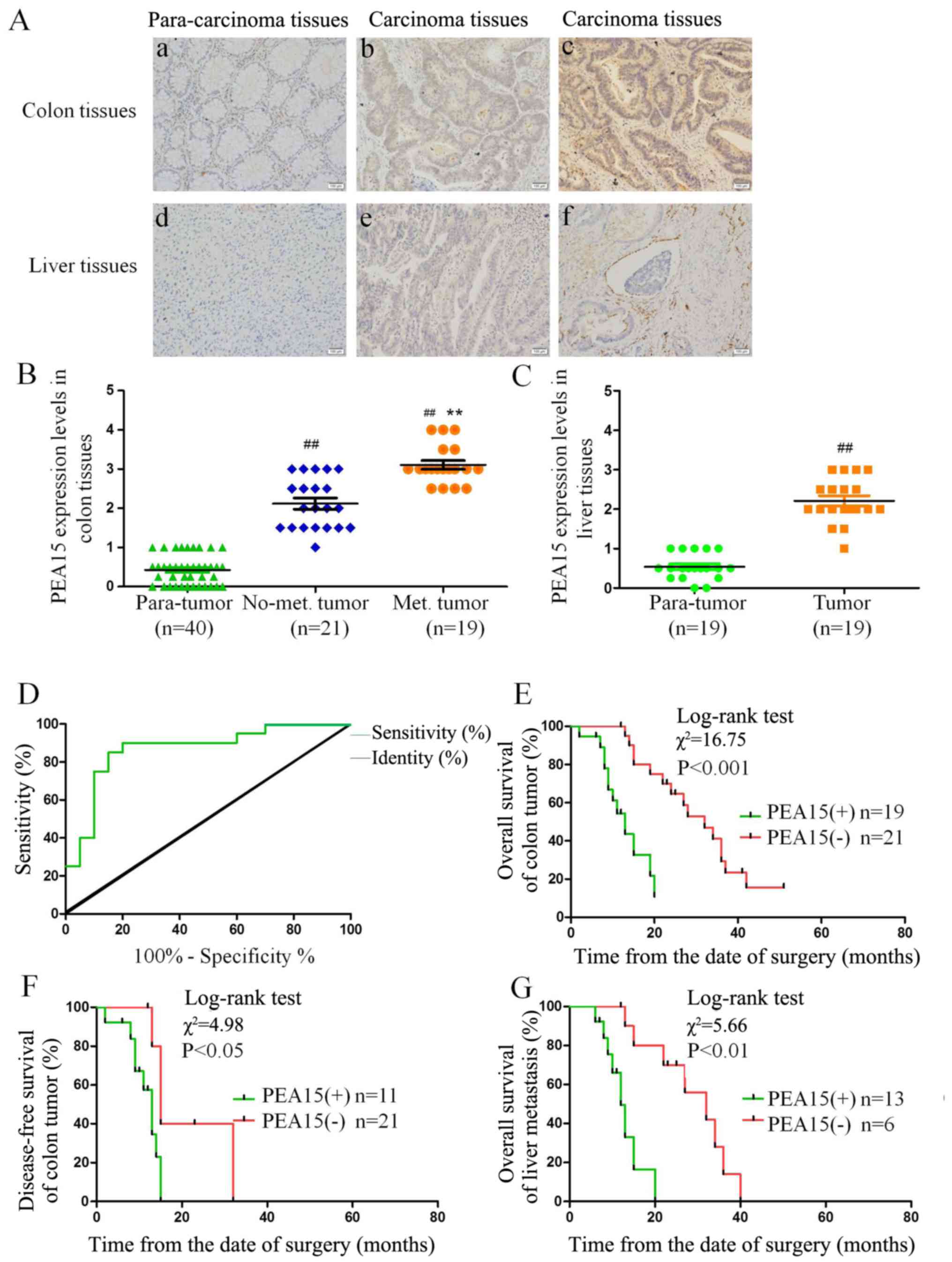
non-metastatic liver cancer tissues (Fig. 1A-C). Notably, we found that the
expression of PEA15 was also significantly increased in endangium
of liver metastasis (Fig. 1Aa-f),
indicating that PEA15 may be involved in the hematogenous
metastasis of tumors. Combined with clinicopathological data of
patients with liver metastasis of colorectal cancer, we analyzed
the correlation between the expression of PEA15 and the
clinicopathological features of patients. After mapping the ROC
curve of PEA15 expression (Fig. 1D)
and determining the division point of high or low PEA15 expression,
we divided the patients into a PEA15-high expression group and a
low expression group. The statistical results revealed that the
expression of PEA15 was correlated with TNM stage (P<0.05) and
liver metastasis (P<0.001) (Table
I).
 | Table I.Correlations between PEA15 expression
and clinicopathological parameters in 40 colorectal carcinoma
patients. |
Table I.
Correlations between PEA15 expression
and clinicopathological parameters in 40 colorectal carcinoma
patients.
|
| PEA15
expression |
|
|
|---|
|
|
|
|
|
|---|
| Variables | Low (21) | High (19) | Total | P-value |
|---|
| Age (years) |
|
|
| 0.548 |
|
<40 | 9 (47) | 10 (53) | 19 |
|
|
≥40 | 12 (57) | 9 (43) | 21 |
|
| Sex |
|
|
| 0.125 |
|
Male | 7 (39) | 11 (61) | 18 |
|
|
Female | 14 (64) | 8 (36) | 22 |
|
| Tumor diameter
(cm) |
|
|
| 0.062 |
|
<5 | 5 (33) | 10 (67) | 15 |
|
| ≥5 | 16 (64) | 9 (36) | 25 |
|
| Tumor
differentiation |
|
|
| 0.292 |
|
Well | 3 (33) | 6 (67) | 9 |
|
|
Moderate | 8 (50) | 8 (50) | 16 |
|
|
Poor | 10 (67) | 5 (33) | 15 |
|
| TNM stage |
|
|
| 0.028a |
|
I–II | 6 (33) | 12 (67) | 18 |
|
|
III–IV | 15 (68) | 7 (32) | 22 |
|
| Liver
metastasis |
|
|
|
<0.001a |
| No | 17 (81) | 4 (19) | 21 |
|
|
Yes | 4 (21) | 15 (79) | 19 |
|
PEA15 is positively correlated with
the clinical prognosis of patients with liver metastasis of
colorectal cancer and the BRAF mutation
Through follow-up of 5-year survival time with
patients after being clinically diagnosed, a Kaplan-Meier survival
curve was used to analyze the correlation between the expression
level of PEA15 and clinical prognosis, and the overall survival
time (median, 13 vs. 32 months; P<0.001) and disease-free
survival time (median, 11 months vs. 28 months; P<0.05) of
patients with liver metastasis of colorectal cancer and low PEA15
expression were significantly better than those of the
PEA15-overexpression group (Fig. 1E and
F). In addition, in patients with liver metastatis, the total
survival time (median, 13 months vs. 15 months, P<0.01) of the
PEA15-low expression group was longer than that of the PEA15-high
expression group (Fig. 1G).
Multivariate Cox regression analysis revealed that PEA15
overexpression (relative risk=3.698; P=0.016) could be used as a
relatively independent predictor of poor prognosis in patients with
liver metastases of colorectal cancer (Table II). Furthermore, we detected and
analyzed the correlation between the high expression of PEA15 and
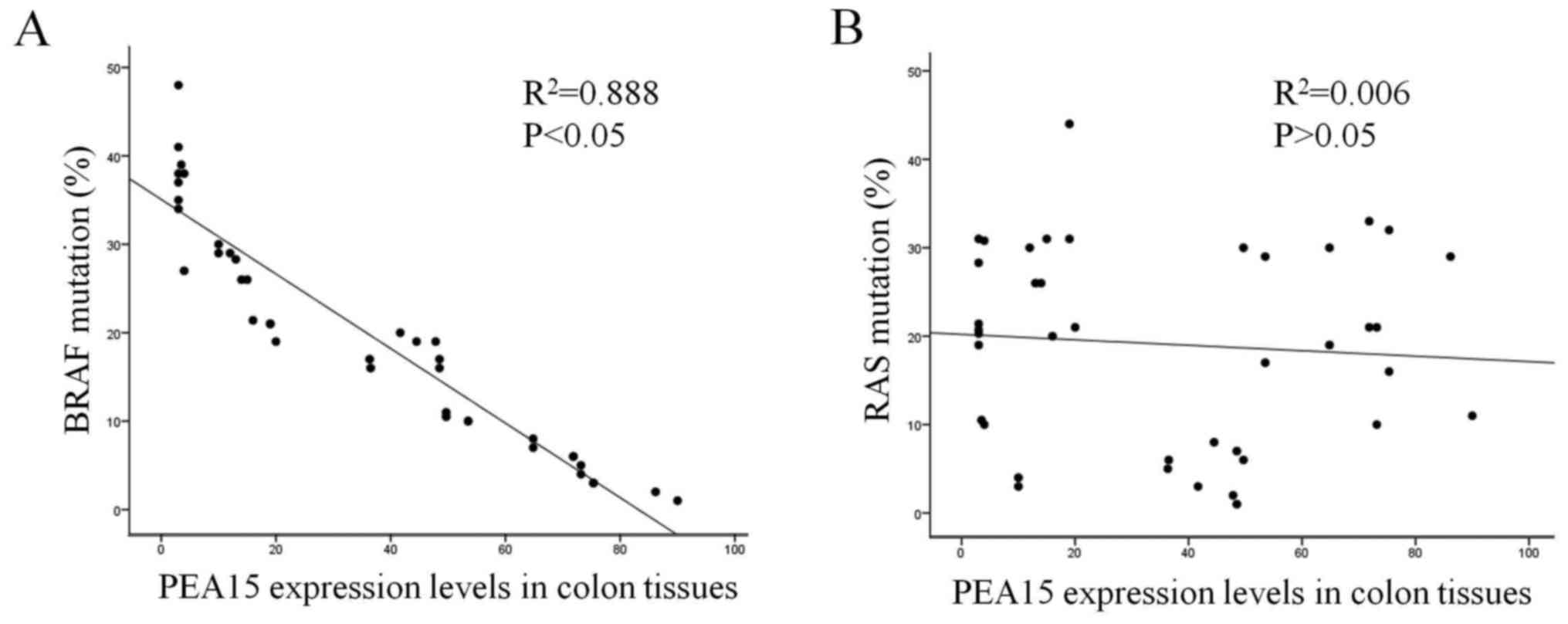
the mutation rates of RAS and BRAF in colorectal cancer tissues.
The results revealed that the high expression of PEA15 was
positively correlated with BRAF mutation but not with RAS mutation
(Fig. 2A and B).
| Table II.Multivariate analysis with a Cox
proportional hazards regression model. |
Table II.
Multivariate analysis with a Cox
proportional hazards regression model.
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Variables | RR | 95% CI | P-value | RR | 95% CI | P-value |
|---|
| Age (years) (<40
vs. >40) | 0.521 | 0.420–0.647 | 0.760 | 0.671 | 0.520–0.864 | 0.078 |
| Sex (male vs.
female) | 1.638 | 1.318–2.034 | 0.782 | 0.753 | 0.592–0.957 | 0.082 |
| Tumor diameter (cm)
(<5 vs. >5) | 1.509 | 1.217–1.872 | 0.325 | 3.337 | 2.599–4.284 | 0.095 |
| Tumor
differentiation (well vs. poor) | 1.144 | 0.616–2.124 | 0.370 | 0.544 | 0.421–0.703 | 0.061 |
| TNM stage (I–II vs.
III–IV) | 0.807 | 0.657–0.991 | 0.041a | 0.633 | 0.499–0.803 | 0.003a |
| Liver metastasis
(No vs. yes) | 7.758 | 5.598–10.75 |
<0.001a | 7.410 | 5.181–10.6 | 0.001a |
| PEA15 (Positive vs.
negative) | 4.434 | 3.356–5.858 |
<0.001a | 3.698 | 2.636–5.189 | 0.016a |
PEA15 promotes the proliferation of
colorectal cancer cells in vivo and in vitro
To investigate the regulatory effect of PEA15 on
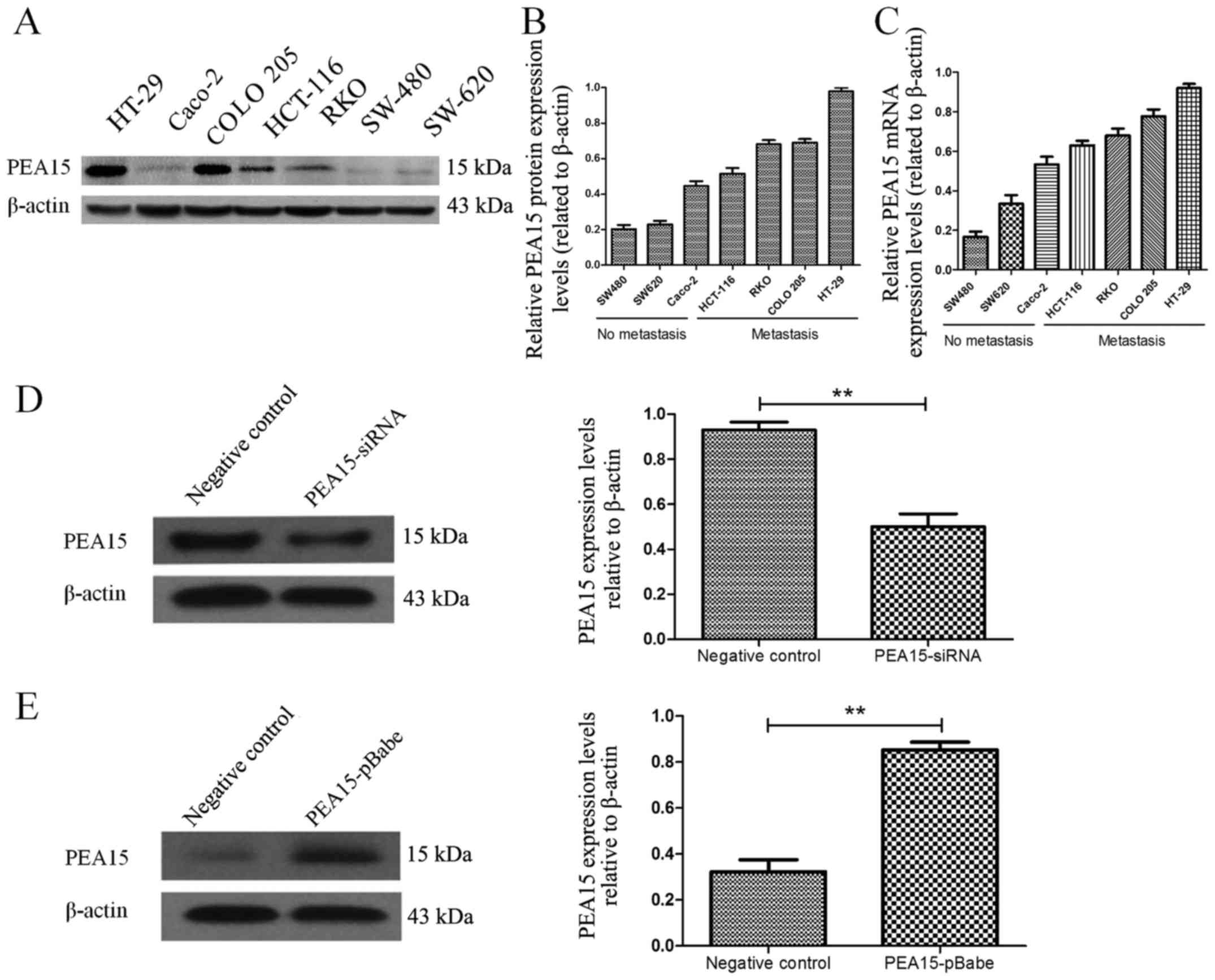
colorectal cancer cell lines, we first selected seven colorectal
cancer cell lines (HT-29, Caco-2, HCT-116, RKO, COLO-205, SW-620
and SW-480). The expression of PEA15 in different cell lines was
detected by PCR and western blotting, respectively. The results
revealed that the expression level of PEA15 was significantly
increased in colorectal cancer cell lines with metastatic potential
(Fig. 3A-C). To evaluate the role
of PEA15 in cell proliferation and tumor growth, we selected the
HT-29 and RKO cell lines that produce PEA15 at high levels and
proceeded to knockdown its expression using short hairpins RNAs
(PEA15-shRNA plasmid). We also selected the SW-480 cell line to
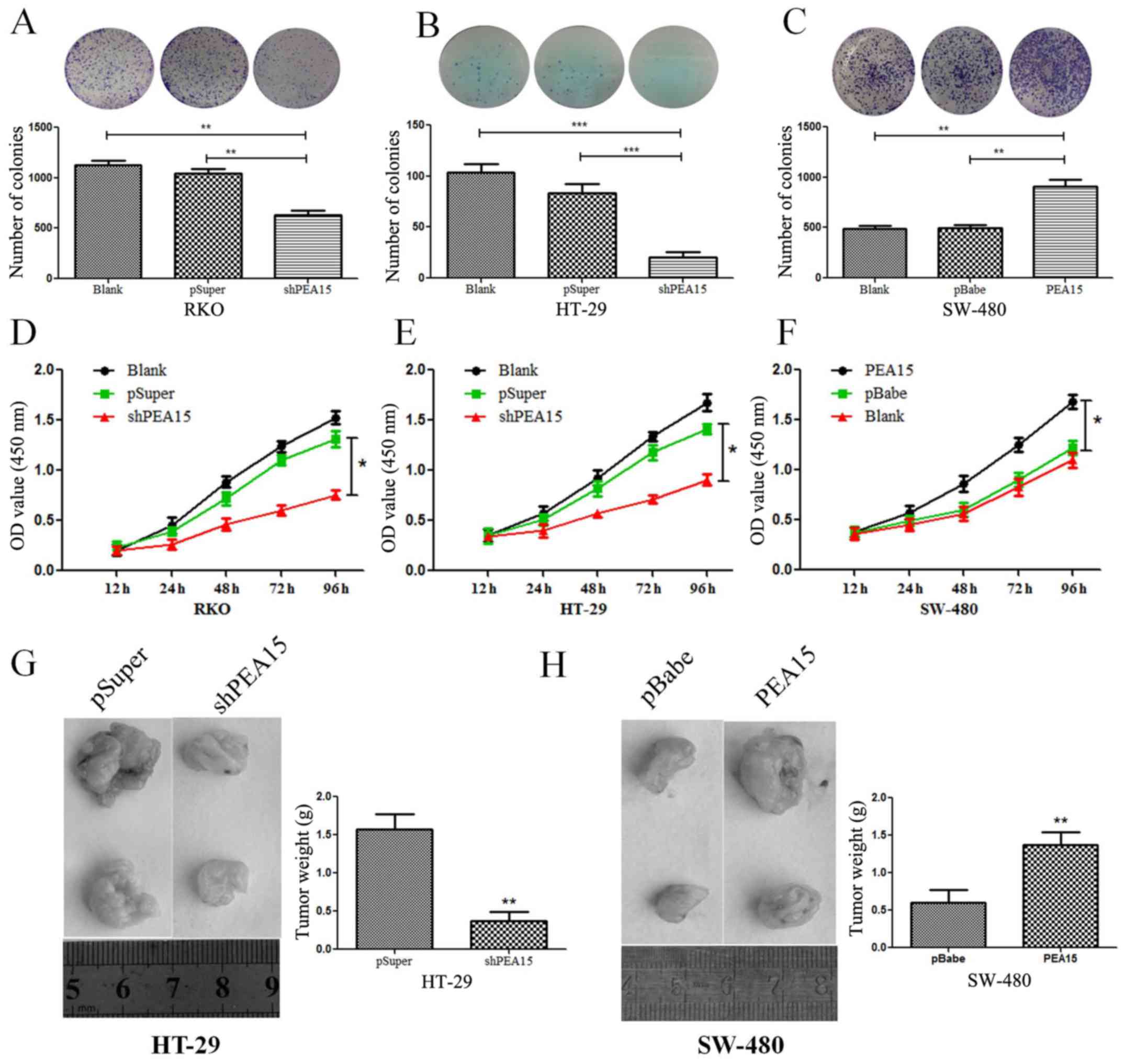
transfect with the PEA15 plasmid as aforementioned (Fig. 3D and E). The plate cloning results
revealed that the colony formation of HT-29 and RKO cells after
PEA15 downregulation was significantly less than that of the normal
and negative control group (Fig. 4A and
B), but the colony formation of SW-480 cells after PEA15
upregulation was significantly more than that of the normal and
negative control group (Fig. 4C).
The CCK-8 kit assay results revealed that the proliferation ability
of HT-29 and RKO cells was significantly weakened after PEA15
downregulation (Fig. 4D and E)
compared with normal group and negative control group, while the
proliferation of SW-480 cells increased after PEA15 upregulation
(Fig. 4F). In vivo, it was
also confirmed that downregulation of PEA15 significantly reduced
the tumor weight in nude mice, whereas upregulation of PEA15
increased the tumor weight (Fig. 4G and
H).
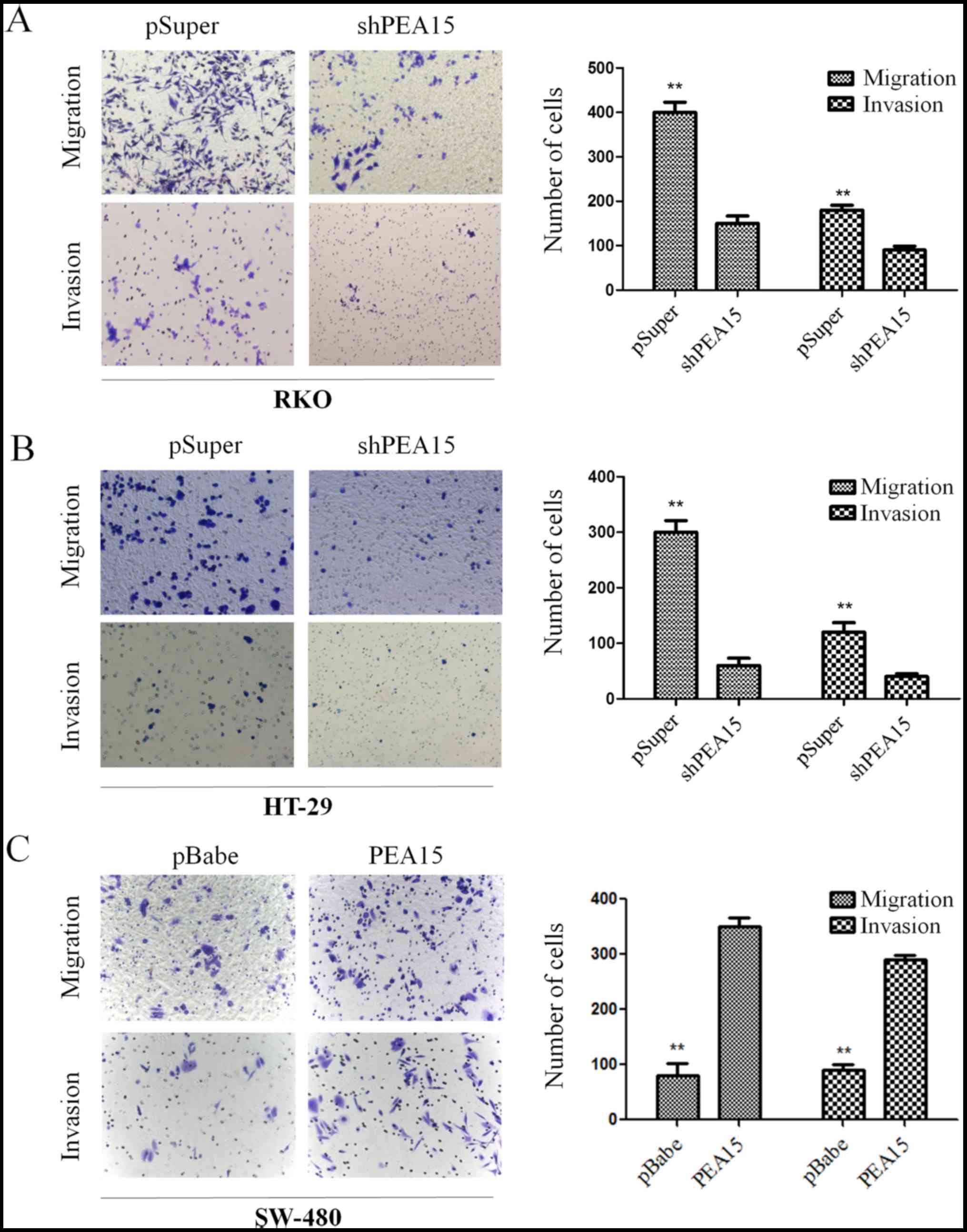
PEA15 promotes the invasion and
migration of colorectal cancer cells in vitro and participates in
the regulation of EMT
According to Transwell chamber invasion and
migration experiments, the invasion and migration abilities of
HT-29 and RKO cells was evidently decreased after downregulation of
PEA15 (Fig. 5A and B), while in
SW-480 cells, these abilities were enhanced after PEA15 was
upregulated (Fig. 5C).
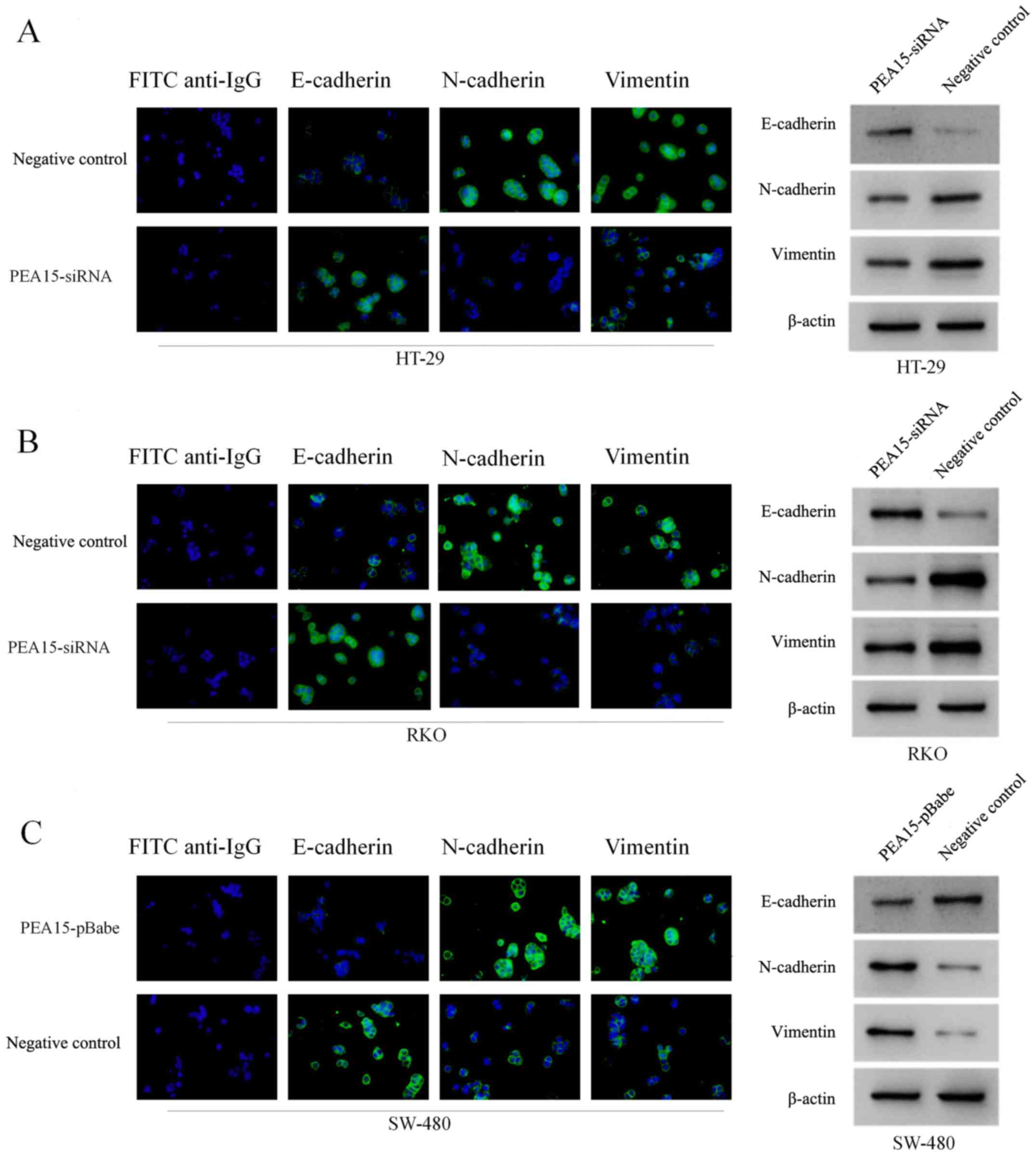
Subsequently, we examined the effect of PEA15 on the expression of
EMT-related proteins in colorectal cancer cells by
immunofluorescence confocal microscopy and western blotting. The
results revealed that after PEA15 downregulation, the expression of
E-cadherin in HT-29 and RKO cells increased, while the expression
of N-cadherin and vimentin decreased (Fig. 6A and B); while in SW-480 cells, the
opposite was observed after PEA15 was downregulated (Fig. 6C). The aforementioned results
indicated that PEA15 promoted the invasion and migration of tumor
cells by regulating the EMT of colorectal cancer cells.
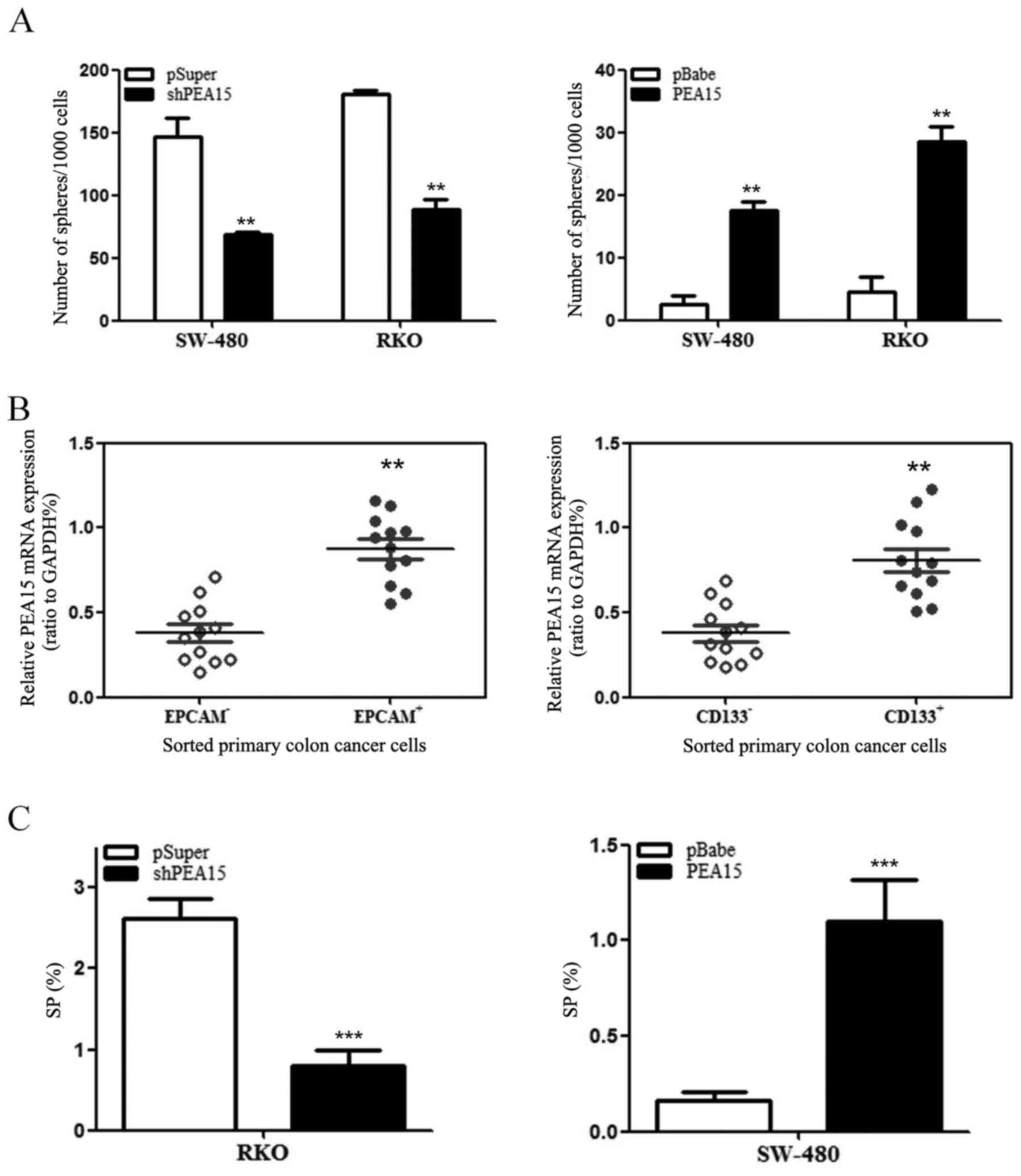
PEA15 induces stem cell-like features
of colorectal cancer cells
In order to investigate whether PEA15 regulated the
stem cell characteristics of colorectal cancer cells, we detected
the ability of spheroid formation of SW-480 cells and RKO cells by
upregulation or downregulation of PEA15. The results revealed that
with the upregulation of PEA15, the ability of spheroid formation
of SW-480 cells and RKO cells was enhanced. In contrast, the
ability of spheroid formation of both cells weakened after PEA15
downregulated (Fig. 7A).
Subsequently, we used flow cytometry to detect the proportion of
surface cancer antigen marker molecules (EPCAM and CD133) of the
colorectal cancer cells and the ratio of side population cells. The
experimental results revealed that the high expression of PEA15 in
colorectal cancer cells exhibited more of EPCAM+ and
CD133+ (Fig. 7B). Side
population (SP) cell ratio analysis revealed that the proportion of
SP increased in SW-480 cells after PEA15 was upregulated, whereas
the proportion of SP in RKO cells decreased after PEA15 was
downregulated (Fig. 7C). Therefore,
the aforementioned results indicated that PEA15 could regulate the
stem cell characteristics of colorectal cancer cells.
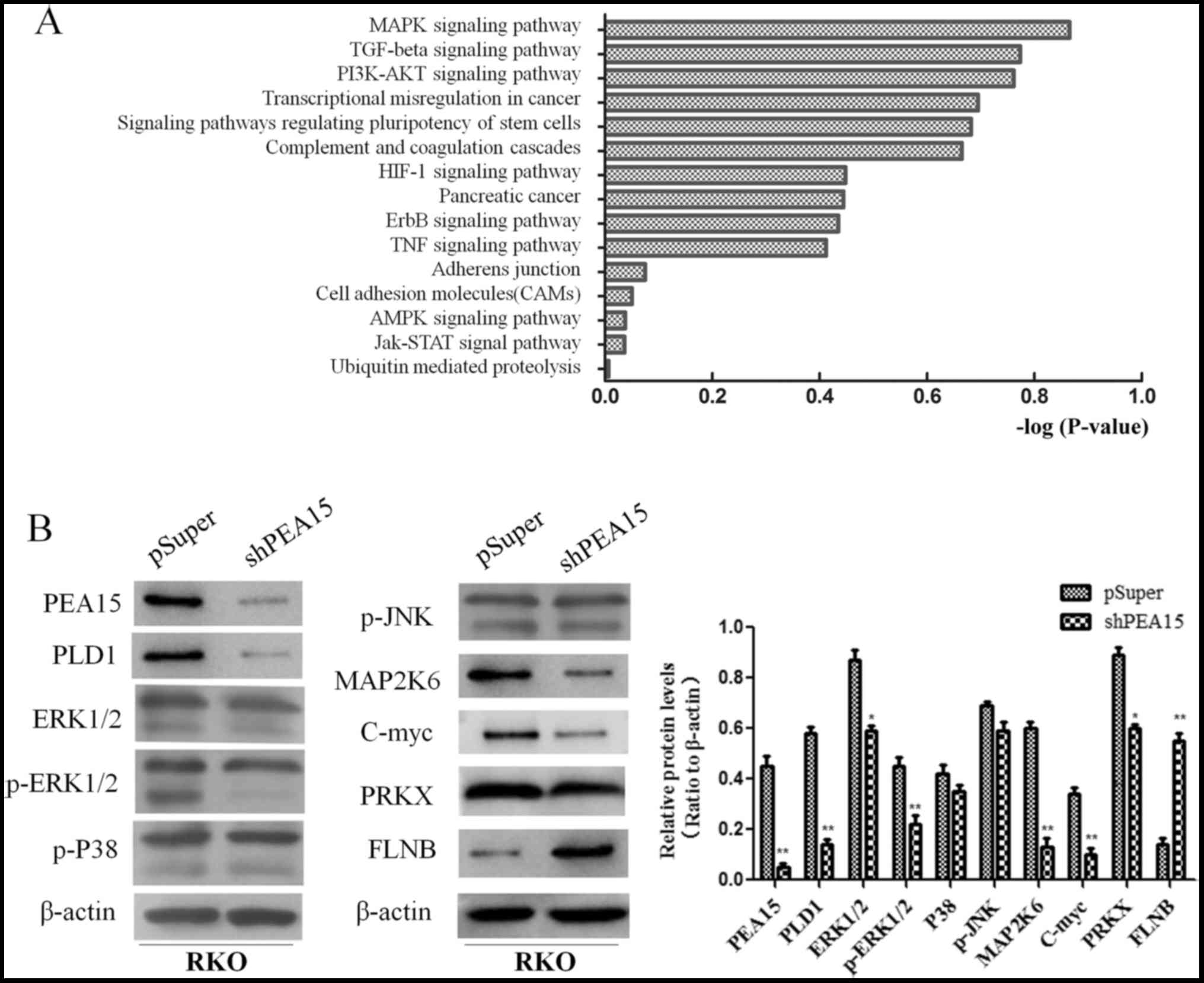
PEA15 promotes the development of
colorectal cancer by regulating the MAPK signaling pathway
In order to further explore the mechanism of PEA15
in the regulation of liver metastasis of colorectal cancer, we
compared the gene expression differences of HT-29 cells between the
control group and the PEA15-downregulation group by gene chip
analysis (the data is not currently available in GEO). Scatter
plots were used to analyze the significantly changed genes in the
PEA15-downregulation group and the control group. More than 400
genes with upregulation or downregulation change of more than 3
times were selected. Gene microarray analysis revealed that these
genes were involved in a variety of signaling pathways, including
the MAPK signaling pathway, the TGF-β signaling pathway, the
PI3K/AKt signaling pathway, the stem cell regulatory pathway and
the HIF-1 signaling pathway, while the most noticeable difference
gene change was in the MAPK signaling pathway (Fig. 8A). These results indicated that
PEA15 affected the development of colorectal cancer cells mainly
through the regulation of the MAPK signaling pathway. We further
detected the expression of MAPK signaling pathway-related proteins
before and after PEA15 downregulation by western blotting. The
results revealed that the expression of PEA15, PLD1, ERK, MAP2K6,
c-Myc and PRKX proteins decreased and the expression of FLNB
protein increased in RKO cells after downregulation of PEA15, while
the expression of p38 and JNK proteins exhibited no significant
changes (Fig. 8B). Furthermore, we
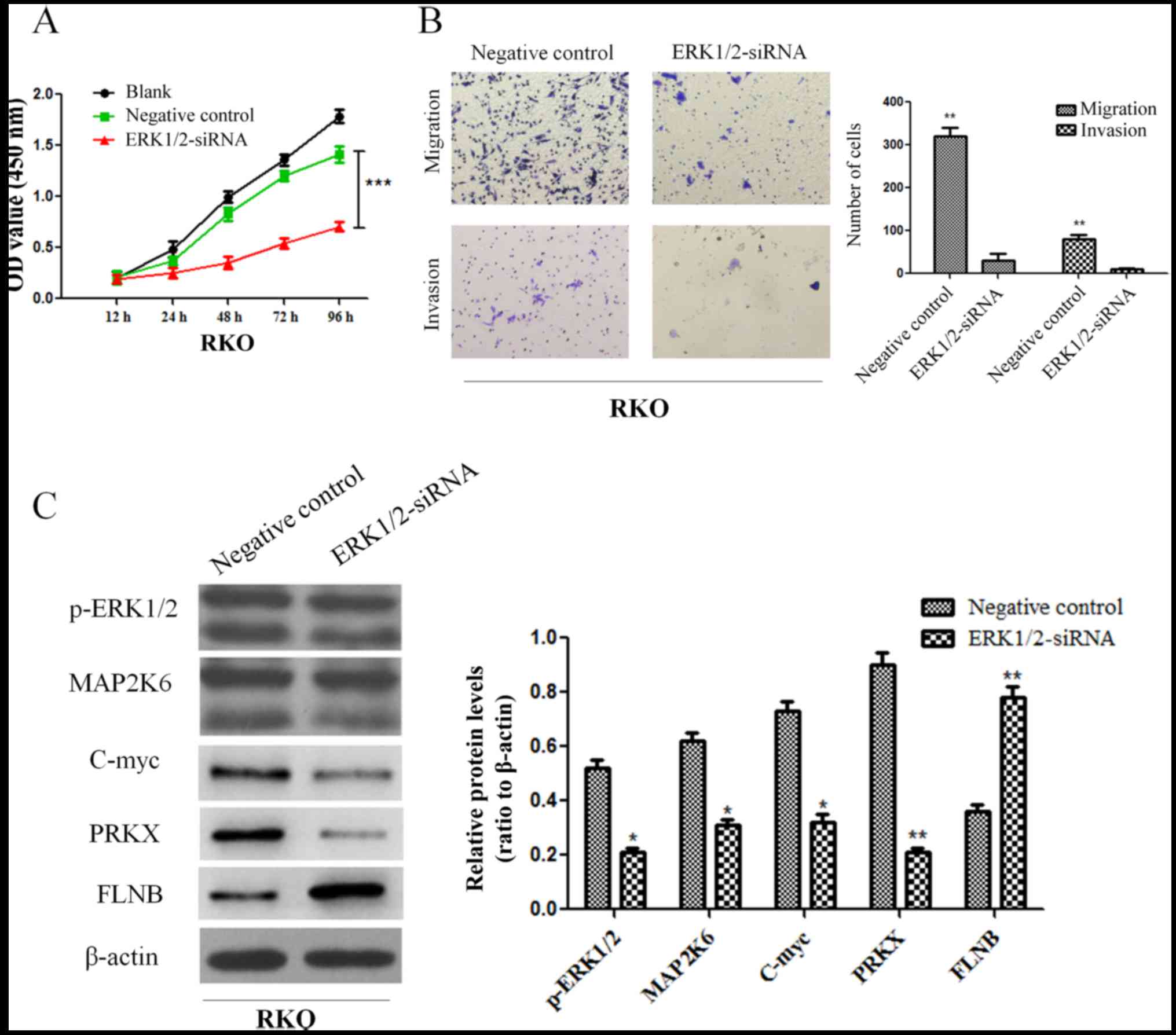
downregulated ERK1/2 and found that the proliferation, invasion and
migration abilities of RKO cells were all suppressed (Fig. 9A and B). Moreover, the results
revealed that the expression of MAP2K6, c-Myc and PRKX proteins
decreased while the expression of FLNB protein increased in RKO
cells after downregulation of ERK1/2 (Fig. 9C). The aforementioned results
indicated that PEA15 regulated the development of colorectal cancer
by regulating the ERK/MAPK signaling pathway.
PEA15 promotes the formation of
metastases of colorectal cancer cells in vivo
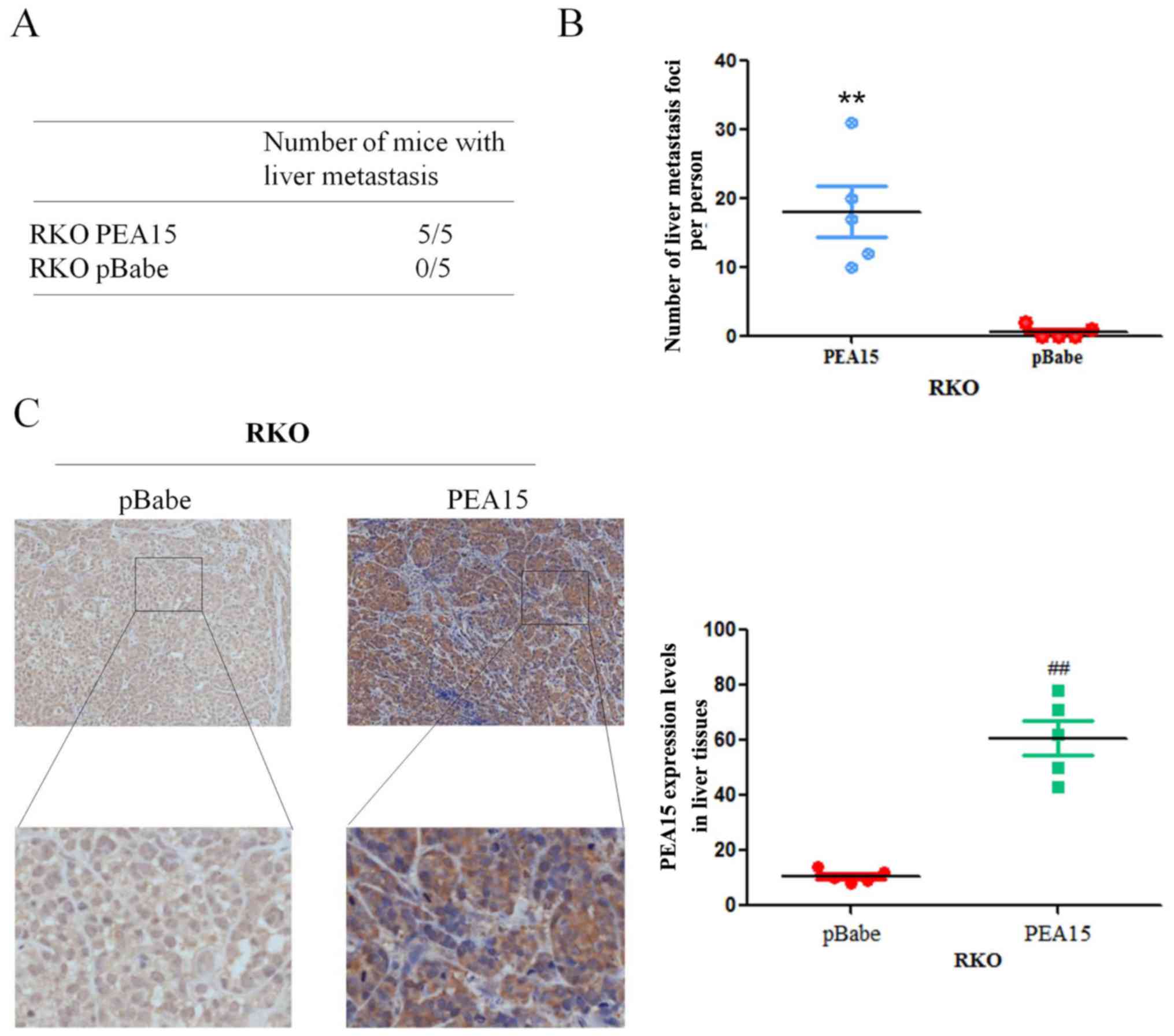
In order to verify the effect of PEA15 on the
metastatic potential of colorectal cancer cells in vivo, we
upregulated PEA15 in the RKO cell line and injected it into the
hepatic portal vein of nude mice. After 60 days of inoculation,
nude mice were sacrificed by cervical dislocation, and the liver
metastasis foci were recorded. The results revealed that the foci
number of liver metastases significantly increased after PEA15
upregulation (Fig. 10A and B).
Meanwhile, immunohistochemistry revealed that the expression of
PEA15 in liver tissues was significantly higher in the
PEA15-upregulated colorectal cancer cell group (Fig. 10C). The aforementioned results
confirmed that PEA15 promoted the development of liver metastasis
of colorectal cancer by regulating the ERK/MAPK signaling pathway
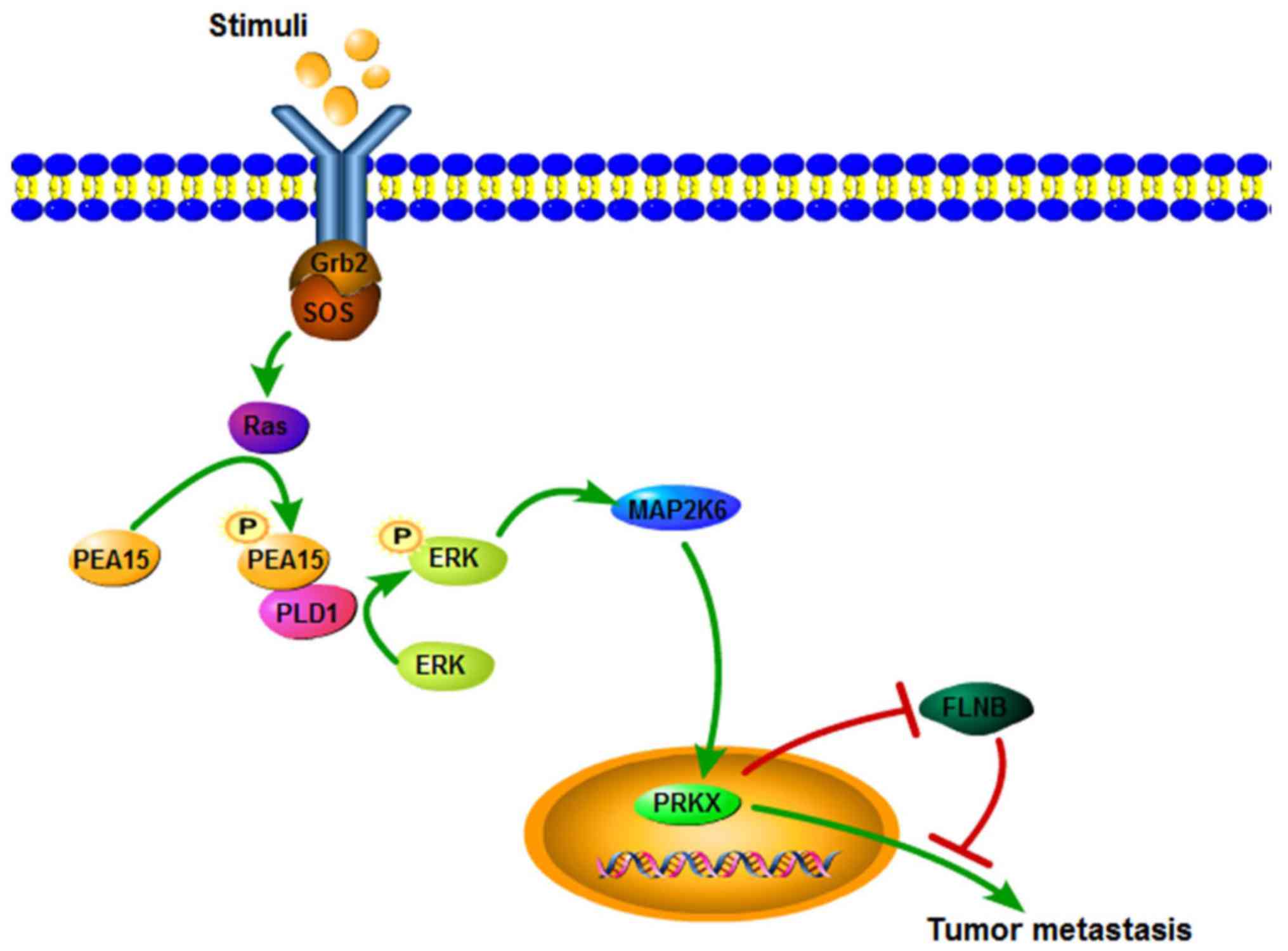
(Fig. 11).
Discussion
A previous study revealed that PEA15 is a key member
of the anti-apoptotic family that inhibits apoptosis by restraining
the formation of the death-inducing signal complex (DISC) and
blocking the cascade activation of caspases (18). The present study confirmed that
PEA15 promoted the proliferation of colorectal cancer cells in
vitro and in vivo. Notably, we also found that the
expression of PEA15 in colorectal cancer with liver metastasis was
significantly higher than that without metastasis. In addition, the
clinical TNM stage and BRAF gene mutation were positively
correlated with PEA15 expression. These results indicated that
PEA15 may be involved in the regulation of liver metastasis of
colorectal cancer.
Current studies have revealed that EMT of colorectal
cancer cells is a key factor in the distant metastasis of
colorectal cancer (19). The
present study revealed that when PEA15 was downregulated, the
expression of E-cadherin in colorectal cancer cells increased,
however, V-cadherin and vimentin decreased, and the invasion and
migration capacities of colorectal cancer cells were significantly
weakened. Tumor stem cell characterization test results also
revealed that with downregulation of PEA15, the ability of cell
formation became weaker and the proportion of side population cells
decreased. While with upregulation of PEA15, the opposite results
were obtained. Therefore, we conclude that PEA15 regulated EMT to
promote liver metastasis of colorectal cancer and induced stem cell
characteristics of colorectal cancer cells.
Current studies have also found that the MAPK
signaling pathway may be involved in the regulation of colorectal
cancer of liver metastasis (20).
The MAPK signaling pathway is one of the most important regulatory
pathways, and its signal-related proteins are widely distributed in
the nucleus, cytoplasm, mitochondria and Golgi participating in
regulating the various life activities of the body (21). The ERK/MAPK signaling pathway can
regulate cell proliferation and invasion, promote the EMT process
and finally promote the development of colorectal cancer (22,23).
However, phosphorylation of two sites, Ser-104 and Ser-116 in PEA15
can promote tumor invasion and migration (24). The present study revealed that the
expression of PEA15, PLD1, ERK, MAP2K6 and PRKX was downregulated,
and the expression of FLNB was upregulated, while the expression of
p38 and JNK was not markedly changed after PEA15 downregulation. In
addition, downregulation of ERK1/2 supressed the proliferation,
invasion and migration abilities of RKO cells. Therefore, we
concluded that PEA15 regulated the ERK/MAPK signaling pathway.
Research has shown that phosphorylated PEA15 can phosphorylate ERK
in combination with PLD1, thereby exerting a tumor-promoting effect
(25). In gliomas, Ser116
phosphorylation of PEA-15 regulates the ERK/MAPK signaling axis and
antagonizes glioma cell death due to glucose deficiency (26). In addition, PEA15 could regulate
PRKX to promote tumor epithelial cell proliferation, invasion and
vascular structure formation (27).
Our results also revealed that the expression of PLD1 and PRKX
proteins decreased when PEA15 was downregulated, and PEA15
expression was positive in vascular endothelium in metastatic liver
tissues. These results confirmed that PEA15 promoted the invasion
and migration of colorectal cancer cells through the ERK/MAPK
signal axis, and promoted EMT of colorectal cancer cells and
generation of stem cell characteristics, thereby contributing to
the occurrence of liver metastasis of colorectal cancer.
In summary, the present study confirmed that PEA15
is highly expressed in colorectal and metastatic liver tissues. We
also determined for the first time that high expression of PEA15
was positively correlated with tumor TNM stage and liver
metastasis, and acted as a relatively independent predictor of poor
prognosis in patients with liver metastasis of colorectal cancer.
Concurrently, the present study revealed that PEA15 not only
regulated proliferation ability, but also the invasion and
migration capacities, as well as EMT characteristics, and induced
the stem cell characteristics of cancer cells. In relation to its
mechanism, we determined for the first time that the PEA15/ERK/MAPK
signaling pathway induced invasion and migration by regulating the
EMT of colorectal cancer cells and induced tumor stem cell
characteristics.
Acknowledgements
We wish to particularly acknowledge the patients
enrolled in this study for their participation, and the Department
of Pathology and Physiopathology, Guilin Medical University, for
its collaboration in providing the human samples and the clinical
information used in this study with the appropriate ethics
approval.
Funding
The present study was supported in part by The
National Natural Science Foundation of China (no. 81560393), the
Guangxi Science Fund for Distinguished Young Scholars Program (no.
2016GXNSFFA380003), the Natural Science Foundation of Guangxi (no.
2015jjDA40010) and the Scientific Research and Technology
Development Project for Guilin (no. 20140310-2-2).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
BT, YW and CY conceived and designed the
experiments. BT and WL performed the experiments. BT, WL, YL, and
ZL analyzed the data. YW, CY supervised the whole experimental work
and revised the manuscript. BT and WL wrote the paper. All authors
read and approved the manuscript and agree to be accountable for
all aspects of the research in ensuring that the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
All clinical cases and specimen tissues were
reviewed and approved by the Ethics Committee of Guilin Medical
University, and informed consents were signed by the patients
according to the Declaration of Helsinki. The animal experiments
were approved by the Animal Care Committee of Guilin Medical
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wu Z, Wei D, Gao W, Xu Y, Hu Z, Ma Z, Gao
C, Zhu X and Li Q: TPO-induced metabolic reprogramming drives liver
metastasis of colorectal cancer CD110+ tumor-initiating cells. Cell
Stem Cell. 17:47–59. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Renault F, Formstecher E, Callebaut I,
Junier MP and Chneiweiss H: The multifunctional protein PEA-15 is
involved in the control of apoptosis and cell cycle in astrocytes.
Biochem Pharmacol. 66:1581–1588. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Danziger N, Yokoyama M, Jay T, Cordier J,
Glowinski J and Chneiweiss H: Cellular expression, developmental
regulation, and phylogenic conservation of PEA-15, the astrocytic
major phosphoprotein and protein kinase C substrate. J Neurochem.
64:1016–1025. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Farina B, Doti N, Pirone L, Malgieri G,
Pedone EM, Ruvo M and Fattorusso R: Molecular basis of the
PED/PEA15 interaction with the C-terminal fragment of phospholipase
D1 revealed by NMR spectroscopy. Biochim Biophys Acta.
1834:1572–1580. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nagarajan A, Dogra SK, Liu AY, Green MR
and Wajapeyee N: PEA15 regulates the DNA damage-induced cell cycle
checkpoint and oncogene-directed transformation. Mol Cell Biol.
34:2264–2282. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fiory F, Formisano P, Perruolo G and
Beguinot F: Frontiers: PED/PEA-15, a multifunctional protein
controlling cell survival and glucose metabolism. Am J Physiol
Endocrinol Metab. 297:E592–E601. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mace PD, Wallez Y, Egger MF, Dobaczewska
MK, Robinson H, Pasquale EB and Riedl SJ: Structure of ERK2 bound
to PEA-15 reveals a mechanism for rapid release of activated MAPK.
Nat Commun. 4:16812013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Quintavalle C, Hindupur SK, Quagliata L,
Pallante P, Nigro C, Condorelli G, Andersen JB, Tagscherer KE, Roth
W, Beguinot F, et al: Phosphoprotein enriched in diabetes
(PED/PEA15) promotes migration in hepatocellular carcinoma and
confers resistance to sorafenib. Cell Death Dis. 8:e31382017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zanca C, Garofalo M, Quintavalle C, Romano
G, Acunzo M, Ragno P, Montuori N, Incoronato M, Tornillo L,
Baumhoer D, et al: PED is overexpressed and mediates TRAIL
resistance in human non-small cell lung cancer. J Cell Mol Med.
12:2416–2426. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shin M, Lee KE, Yang EG, Jeon H and Song
HK: PEA-15 facilitates EGFR dephosphorylation via ERK sequestration
at increased ER-PM contacts in TNBC cells. FEBS Lett.
589:1033–1039. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bartholomeusz C, Rosen D, Wei C, Kazansky
A, Yamasaki F, Takahashi T, Itamochi H, Kondo S, Liu J and Ueno NT:
PEA-15 induces autophagy in human ovarian cancer cells and is
associated with prolonged overall survival. Cancer Res.
68:9302–9310. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pinato DJ, Tan TM, Toussi ST, Ramachandran
R, Martin N, Meeran K, Ngo N, Dina R and Sharma R: An expression
signature of the angiogenic response in gastrointestinal
neuroendocrine tumours: Correlation with tumour phenotype and
survival outcomes. Br J Cancer. 110:115–122. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression date using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Method. 25:402–408. 2001.
View Article : Google Scholar
|
|
16
|
Li F and Mahato RI: RNA interference for
improving the outcome of islet transplantation. Adv Drug Deliv Rev.
63:47–68. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sotelo J, Esposito D, Duhagon MA, Banfield
K, Mehalko J, Liao H, Stephens RM, Harris TJ, Munroe DJ and Wu X:
Long-range enhancers on 8q24 regulate c-Myc. Proc Natl Acad Sci
USA. 107:3001–3005. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Urosevic J, Garcia-Albéniz X, Planet E,
Real S, Céspedes MV, Guiu M, Fernandez E, Bellmunt A, Gawrzak S,
Pavlovic M, et al: Colon cancer cells colonize the lung from
established liver metastases through p38 MAPK signalling and PTHLH.
Nat Cell Biol. 16:685–694. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Loboda A, Nebozhyn MV, Watters JW, Buser
CA, Shaw PM, Huang PS, Van't Veer L, Tollenaar RA, Jackson DB,
Agrawal D, et al: EMT is the dominant program in human colon
cancer. BMC Med Genomics. 4:92011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Formisano P, Ragno P, Pesapane A, Alfano
D, Alberobello AT, Rea VE, Giusto R, Rossi FW, Beguinot F, Rossi G
and Montuori N: PED/PEA-15 interacts with the 67 kD laminin
receptor and regulates cell adhesion, migration, proliferation and
apoptosis. J Cell Mol Med. 16:1435–1446. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mor A and Philips MR: Compartmentalized
Ras/MAPK signaling. Annu Rev Immunol. 24:771–800. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fang JY and Richardson BC: The MAPK
signalling pathways and colorectal cancer. Lancet Oncol. 6:322–327.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gu Y, Wang Q, Guo K, Qin W, Liao W, Wang
S, Ding Y and Lin J: TUSC3 promotes colorectal cancer progression
and epithelial-mesenchymal transition (EMT) through WNT/β-catenin
and MAPK signalling. J Pathol. 239:60–71. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee J, Bartholomeusz C, Krishnamurthy S,
Liu P, Saso H, Lafortune TA, Hortobagyi GN and Ueno NT: PEA-15
unphosphorylated at both serine 104 and serine 116 inhibits ovarian
cancer cell tumorigenicity and progression through blocking
β-catenin. Oncogenesis. 1:e222012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sulzmaier F, Opoku-Ansah J and Ramos JW:
Phosphorylation is the switch that turns PEA-15 from tumor
suppressor to tumor promoter. Small GTPases. 3:173–177. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Eckert A, Böck BC, Tagscherer KE, Haas TL,
Grund K, Sykora J, Herold-Mende C, Ehemann V, Hollstein M,
Chneiweiss H, et al: The PEA-15/PED protein protects glioblastoma
cells from glucose deprivation-induced apoptosis via the ERK/MAP
kinase pathway. Oncogene. 27:1155–1166. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li X, Iomini C, Hyink D and Wilson PD:
PRKX critically regulates endothelial cell proliferation,
migration, and vascular-like structure formation. Dev Biol.
356:475–485. 2011. View Article : Google Scholar : PubMed/NCBI
|