Introduction
Malignant glioma is the most common and aggressive
type of primary brain tumor in humans, accounting for more than 44%
of all brain tumors in China and 50% of all primary central nervous
system (CNS) neoplasms in the US (1). Glioblastoma, grade IV glioma, is
notorious for its rapid proliferation, intensive infiltration,
genetic alteration and increased angiogenesis, which makes it the
most malignant glioma accounting for more than 46% of all malignant
brain cancers (2–5). Despite enormous efforts in multimodal
treatment approaches including typical surgical tumor resection,
radiotherapy and chemotherapy, the prognosis of glioblastoma
patients is undeniably grim with only a median survival time of
12–15 months (6,7). Perhaps the dismal prognosis is mainly
due to the limited understanding of the molecular basis of
glioblastoma development and progression. Thus, further research of
the molecular mechanisms underlying glioblastoma and a thorough
investigation of molecular biomarkers for prognosis and targeted
therapy are crucially needed to improve glioblastoma patient
outcomes.
Human trophoblast cell surface antigen 2 (TROP2), a
36-kDa transmembrane protein expressed primarily in epithelial
cells and encoded by the TACSTD2 gene, was originally
discovered in human placental trophoblastic tissue (8). TROP2, as a cell surface receptor, can
recognize specific ligands and participate in numerous
intracellular signaling pathways of cell proliferation,
self-renewal and survival (9–11).
Recently, it has received attention as a potential target for
cancer therapy since high TROP2 expression was found to be
positively associated with poor patient prognosis (12). Overexpression of TROP2 was found in
several human carcinomas, including colorectal (13), lung cancer (14), glioma (15), gastric carcinoma (16), cervical (17), breast (18) and pancreatic cancer (19), underscoring the potential role of
TROP2 in tumorigenesis. However, the precise mechanism and
biological function of TROP2 expression in relation to glioblastoma
have not been studied.
Interleukin-6 (IL-6), a well-characterized
proinflammatory cytokine, has been shown to play a crucial role in
driving many of the ‘hallmarks’ of cancer including activation of
the signaling of cell proliferation, enhancement of migration and
invasion, resistance to cell death and induction of angiogenesis
through downstream activation of the Janus kinase (JAK)/signal
transducer and activator of transcription 3 (STAT3) signaling
pathway (20–25). Persistent activation of JAK2/STAT3
is frequently documented in human carcinomas and has emerged as an
ideal target for cancer treatment (26–28).
Therefore, tight regulation of the JAK2/STAT3 pathway may be
effective in therapy against cancer.
In the present study, we found that downregulation
of TROP2 efficiently inhibited glioblastoma cell proliferation and
metastasis by regulating the JAK2/STAT3 pathway. These data offer
insights into TROP2 function and suggest TROP2 as a potential
target for glioblastoma patients.
Materials and methods
Cell culture and transfection
Human normal astroglia cells (SVGP12), glioblastoma
cell lines [A172, LN-229, U-87 MG (cat. no. HTB-14 of unknown
origin) and U-118 MG and a retroviral packaging cell line (293FT)
were obtained from the American Type Culture Collection (ATCC;
Manassas, VA, USA) and cultured as previously described (29). All cell lines tested
mycoplasma-negative. The sequences of the TROP2 short hairpin RNA
(shTROP2) and GFP short hairpin RNA (shGFP) were purchased from
GenePharma Co., Ltd. (Shanghai, China). Sequences of shTROP2 are
provided in Table I. A vector
encoding the human full-length TROP2 was cloned by PCR-based
amplification and then subcloned into the pCDH-CMV-EF1-copGFP
vector. Sequences of the primers used are given in Table II. The lentivirus was produced as
previously described (30).
 | Table I.Sequences of the TROP2-specific
shRNAs. |
Table I.
Sequences of the TROP2-specific
shRNAs.
| shTROP2-1-F |
CCGGCTACTTCGAGAGGGACATCAACTCGAGTTGATGTCCCTCTCGAAGTAGTTTTTG |
| shTROP2-1-R |
AATTCAAAAACTACTTCGAGAGGGACATCAACTCGAGTTGATGTCCCTCTCGAAGTAG |
| shTROP2-2-F |
CCGGGAGAAAGGAACCGAGCTTGTACTCGAGTACAAGCTCGGTTCCTTTCTCTTTTTG |
| shTROP2-2-R |
AATTCAAAAAGAGAAAGGAACCGAGCTTGTACTCGAGTACAAGCTCGGTTCCTTTCTC |
| Table II.Primer pairs for human full-length
TROP2. |
Table II.
Primer pairs for human full-length
TROP2.
| TROP2-F |
ATGGCTCGGGGCCCC |
| TROP2-R |
CTACAAGCTCGGTTCCTTTCTCA |
Reagents
Gibco™ fetal bovine serum (FBS) and Dulbecco's
modified Eagle's medium (DMEM) were purchased from Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). Dimethyl sulfoxide (DMSO) and
crystal violet were obtained from Sigma-Aldrich/Merck KGaA
(Darmstadt, Germany). WP1066 was obtained from Selleck (Shanghai,
China). The TROP2 (cat. no. ab214488) antibody, survivin (cat. no.
ab76424) antibody and recombinant human IL-6 protein (cat. no.
ab73197) were purchased from Abcam (Shanghai, China); phospho-JAK2
(Tyr1007/1008; cat. no. 3771), JAK2 (cat. no. 3230), phospho-STAT3
(Tyr705; cat. no. 9145), STAT3 (cat. no. 9139), cyclin D1 (cat. no.
2978), MMP2 (cat. no. 40994) and VEGF (cat. no. 9698) antibodies
were purchased from Cell Signaling Technology (Beverly, MA, USA).
GAPDH antibody was purchased from BD Pharmingen (Shanghai, China).
All antibodies were diluted according to the instructions.
Immunohistochemical staining
Formalin-fixed and paraffin-embedded tumor specimens
from patients were sliced into 5-µm thick sections that were then
deparaffinized and hydrated. The procedure of endogenous antigen
retrieval was performed via microwave heating for 20 min in 10 mM
citric acid buffer (pH 6.0). The sections were incubated
sequentially with primary antibodies overnight at 4°C and secondary
antibodies for 30 min at room temperature followed by
diaminobenzidine (DAB) development. All slides were examined and
assessed from microscopic fields at ×20 magnification (Olympus
CKX41; Olympus Corp., Tokyo, Japan). Immunohistochemical staining
results were reviewed and scored as follows: 0, no staining and no
background; 1+, weak staining in more than 30% of cells; 2+,
moderately intense staining in more than 30% of cells but without
intense staining in some cells; and 3+, staining in more than 30%
of tumor cells with markedly intense staining.
Western blot analysis
Cells and fresh tissues were lysed in RIPA lysis
buffer that contained a protease inhibitor cocktail. Proteins were
harvested and then separated by SDS-PAGE and electrotransferred
onto PVDF membranes. The membranes were incubated sequentially with
primary antibodies overnight at 4°C and HRP-linked secondary
antibody for 2 h at room temperature, and then visualized by a
detection system (Clinx Science Instruments Co., Ltd., Shanghai,
China).
Quantitative real-time PCR
Total RNA was extracted from glioblastoma cell lines
using Invitrogen™ TRIzol reagent (Thermo Fisher Scientific) and
then reverse transcribed into cDNA using GoScript™ Reverse
Transcriptase (Promega, Madison, WI, USA). Real-time PCR was
performed using specific primer pairs, cDNA and SYBR Green PCR
Master Mix (Toyobo Life Science, Inc., Osaka, Japan) according to
the manufacturer's instructions. All primer pairs are listed in
Table III and
glycerol-dehyde-3-phosphate dehydrogenase (GAPDH) served as the
endogenous control. Relative quantification of mRNA expression was
defined based on Cq, and the individual values were normalized to
that of the GAPDH control (31).
| Table III.Primer pairs for real-time PCR. |
Table III.
Primer pairs for real-time PCR.
| TROP2-F |
ACAACGATGGCCTCTACGAC |
| TROP2-R |
GTCCAGGTCTGAGTGGTTGAA |
| CCND1-F |
GCTGCGAAGTGGAAACCATC |
| CCND1-R |
CCTCCTTCTGCACACATTTGAA |
| Survivin-F |
AGGACCACCGCATCTCTACAT |
| Survivin-R |
AAGTCTGGCTCGTTCTCAGTG |
| MMP2-F |
GATACCCCTTTGACGGTAAGGA |
| MMP2-R |
CCTTCTCCCAAGGTCCATAGC |
| VEGF-F |
AGGGCAGAATCATCACGAAGT |
| VEGF-R |
AGGGTCTCGATTGGATGGCA |
| GAPDH-F |
ACAACTTTGGTATCGTGGAAGG |
| GAPDH-R |
GCCATCACGCCACAGTTTC |
Cell proliferation analysis
Cell viability was examined using the MTT assay.
Cells (1×103 cells/well) were seeded in 96-well plates
with three replicates, and then were detected from day 0 to 6. The
purple formazan was dissolved by dimethyl sulfoxide, and the
absorbance was measured at a wavelength of 560 nm. All experiments
were carried out independently in triplicate.
BrdU incorporation assay
Cells (2×104 cells/well) were seeded into
24-well plates and incubated overnight. Then a final concentration
of 10 µg/ml BrdU (Sigma-Aldrich; Merck KGaA) was added into the
media and the plates were incubated for 30 min and then fixed for
15 min. After incubation with 1 mol/l HCl for 15 min and 5% goat
serum for 2 h, the cells were incubated sequentially with primary
antibody overnight at 4°C and secondary antibody for 2 h at room
temperature. Cell nuclei were stained with DAPI for 30 min. The
BrdU-positive cells were observed and calculated from microscopy
fields at ×20 magnification (Nikon 80i; Nikon Corp., Tokyo,
Japan).
Flow cytometry
For cell cycle analysis, cells were harvested and
then fixed in 75% ethanol at 4°C for 24 h, and then incubated with
propidium iodide (PI) and RNaseA at 37°C for 30 min. Cells were
examined by flow cytometry (BD Biosciences, San Jose, CA, USA) and
analyzed by FlowJo software (version FlowJo 7.6; FlowJo LLC,
Ashland, OR, USA).
TUNEL analysis
Cells were grown on coverslips and then stained with
Click-iT plus TUNEL assay kit (Invitrogen; Thermo Fisher
Scientific). After incubation with 4% paraformaldehyde for 20 min
and 0.25% Triton X-100 for 15 min, the cells were incubated
sequentially with TdT reaction buffer, TdT reaction mixture and
TUNEL reaction cocktail according to the manufacturer's
instructions. Cell nuclei were stained with DAPI for 30 min. Cells
were imaged and calculated from randomly chosen microscopic fields
at ×20 magnification (Nikon 80i; Nikon Corp.).
Migration and invasion assays
Boyden chambers (24-well) (8-µm pore size; Corning
Inc., Corning, NY, USA) were used in the migration and invasion
assays. Matrigel (BD Biosciences) was used to coat the membranes in
the invasion assay. Medium with 10% FBS as a chemoattractant was
added to the lower chamber, and cells with serum-free media were
placed in the upper chamber. After incubation for 48 h, the cells
were fixed in 4% paraformaldehyde (PFA) and then stained with
crystal violet. Cells were imaged and calculated from randomly
chosen microscopic fields at ×10 magnification (Nikon 80i; Nikon
Corp.).
Soft agar assay
To evaluate colony-forming ability, Gibco™ DMEM 2X
(Thermo Fisher Scientific) containing 0.6% agarose (Sigma-Aldrich;
Merck KGaA) were added to 12-well plates as a bottom layer. Then
0.4×103 cells/well in medium (10% FBS and DMEM)
containing 0.3% agarose (Sigma-Aldrich; Merck KGaA) was added onto
the bottom layer. After culturing for 14–21 days, colonies were
imaged by the scanner (Epson Perfection V700 Photo; Epson, Beijing,
China) and counted in each well.
Xenograft assay
The detailed procedures were performed as described
previously (7). All studies were
approved by the Animal Care and Use Committee of Southwest
University, and carried out in conformity to the Guide for the Care
and Use of Laboratory Animals (Ministry of Science and Technology
of China, 2006).
Patient data analysis and patient
tumor tissues
Kaplan-Meier analysis and survival curves were
downloaded from the online database R2: microarray analysis and
visualization platform (http://hgserver1.amc.nl/cgi-bin/r2/main.cgi). A total
of 8 patients (5 males and 3 females, age range, 32–82, mean age
53.1) who underwent surgical resection between February 2015 and
July 2015 at Daping Hospital, Chongqing were enrolled and tumor and
adjacent normal tissues were collected. Prior approval was obtained
from the Ethics Committee of Daping Hospital, Chongqing, China.
Tissue analysis was approved by the Ethics Committee of Southwest
University of China. All of the patients provided informed
consent.
Statistical analysis
All experiments were carried out in triplicates, and
the quantitative data are expressed as mean ± SD. Two-tailed
Student's t-test was performed to calculate significance in an
interval of 95% confidence level, following normal distribution
with different but similar SDs. A probability (P) value of <0.05
was considered statistically significant. *P<0.05, **P<0.01,
***P<0.001 indicate different degrees of statistical
significance as noted in the figures.
Results
TROP2 is upregulated in human
glioblastoma and is a prognostic indicator for glioblastoma
patients
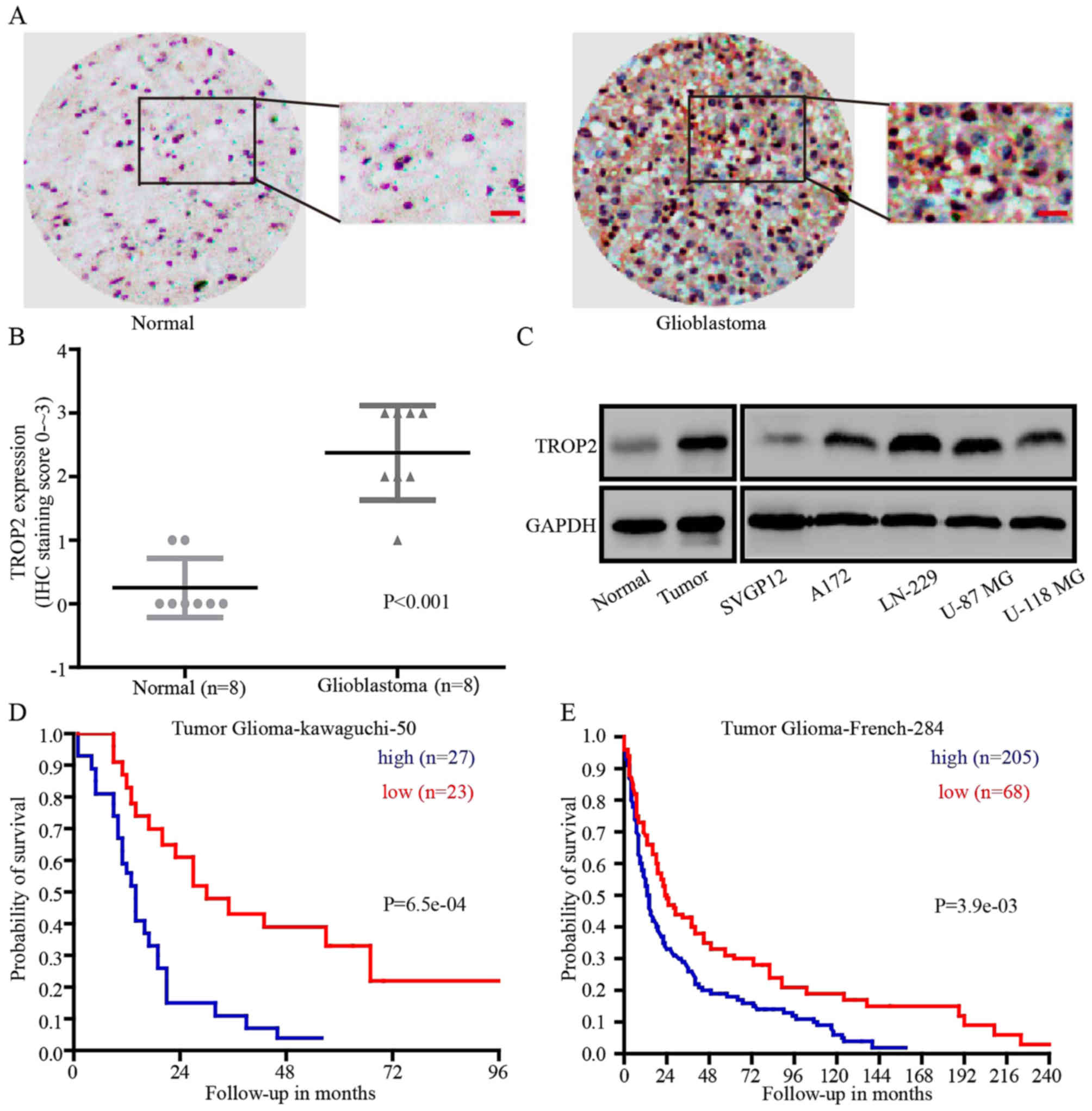
To explore whether TROP2 could be a prognostic
marker for glioblastoma, immunohistochemical analysis (IHC) was
performed. IHC revealed that TROP2 expression was significantly
higher in glioblastoma tissues compared with that noted in the
normal brain tissues in 8 paired samples (Fig. 1A and B). Then, we detected TROP2
expression at the protein level in human normal astroglial cells
(SVGP12), glioblastoma cell lines (A172, LN-229, U-87 MG and U-118
MG), normal tissues and glioblastoma tissues. Compared with SVGP12
cells and normal tissues, TROP2 expression was significantly
increased in all four glioblastoma cell lines and tumor tissues
(Fig. 1C). To determine whether
TROP2 expression level is implicated in the clinical prognosis of
glioblastoma patients, we evaluated the prognostic value of TROP2
in the Tumor Glioma Kawaguchi 50 database and Tumor Glioma French
284 database from the R2 platform (genomics analysis and
visualization platform). The results showed that high TROP2
expression was associated with a poor patient outcome (Fig. 1D and E). Taken together, we found
that TROP2 was upregulated in glioblastoma tumors and glioblastoma
cell lines, and high TROP2 expression was associated with the poor
prognosis of glioblastoma patients.
TROP2 promotes the migration and
invasion of glioblastoma cells
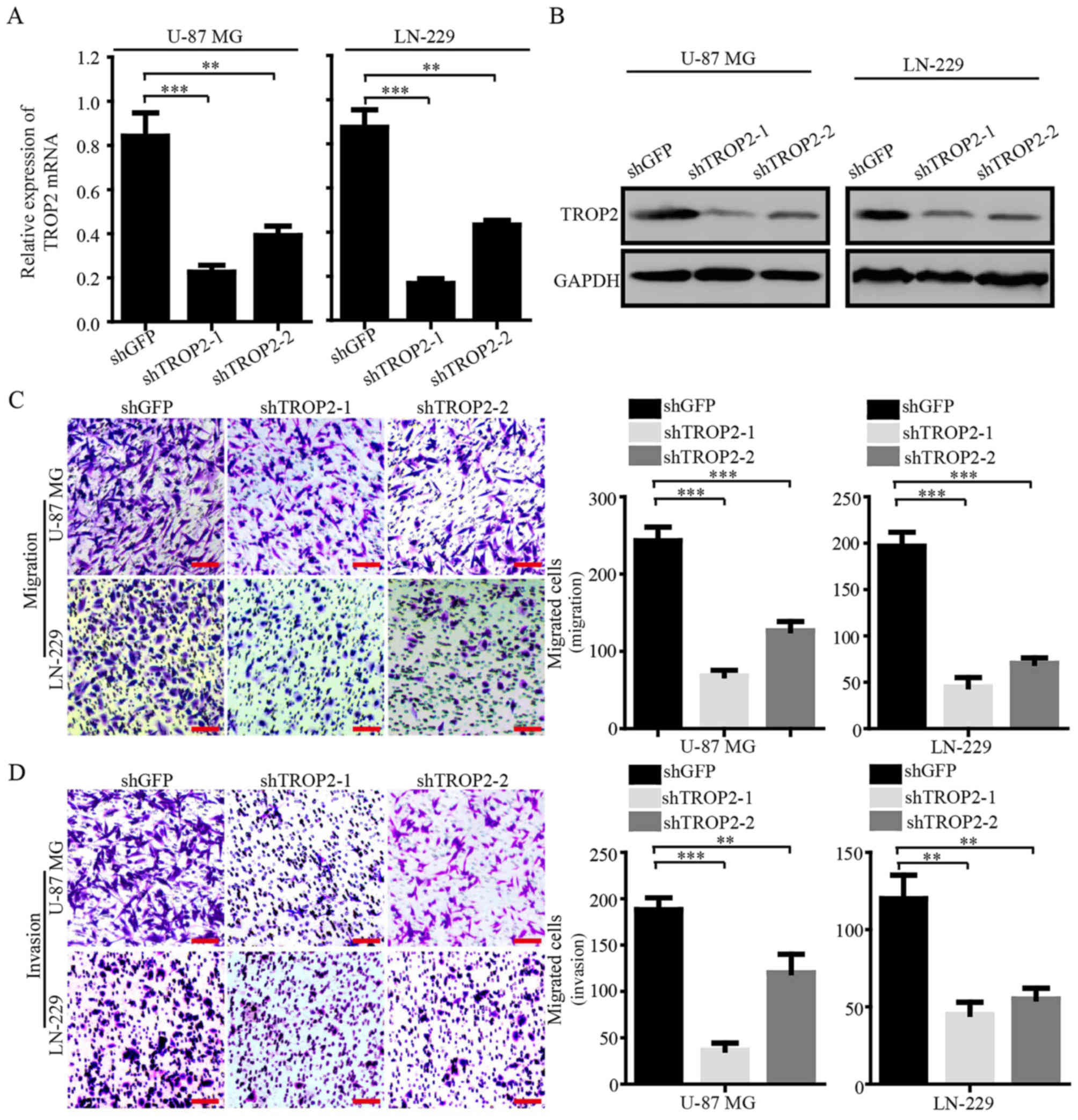
To explore the biological role of TROP2 in
glioblastoma cells, we knocked down TROP2 by using two shRNA
sequences (shTROP2-1 and −2) in glioblastoma cell lines U-87 MG and
LN-229. Quantitative RT-PCR and western blot analysis were
conducted to confirm stable knockdown of TROP2 expression in U-87
MG and LN-229 cells (Fig. 2A and
B). Migration and invasion assays were performed to determine
whether the expression of TROP2 was related to the metastasis of
glioblastoma cells. The results showed that TROP2 knockdown
significantly inhibited cell migration and invasion compared to the
control cells (Fig. 2C and D).
These results demonstrated that TROP2 expression is positively
associated with the cell migration and invasion of glioblastoma
cells.
TROP2 enhances proliferation and
viability of glioblastoma cells
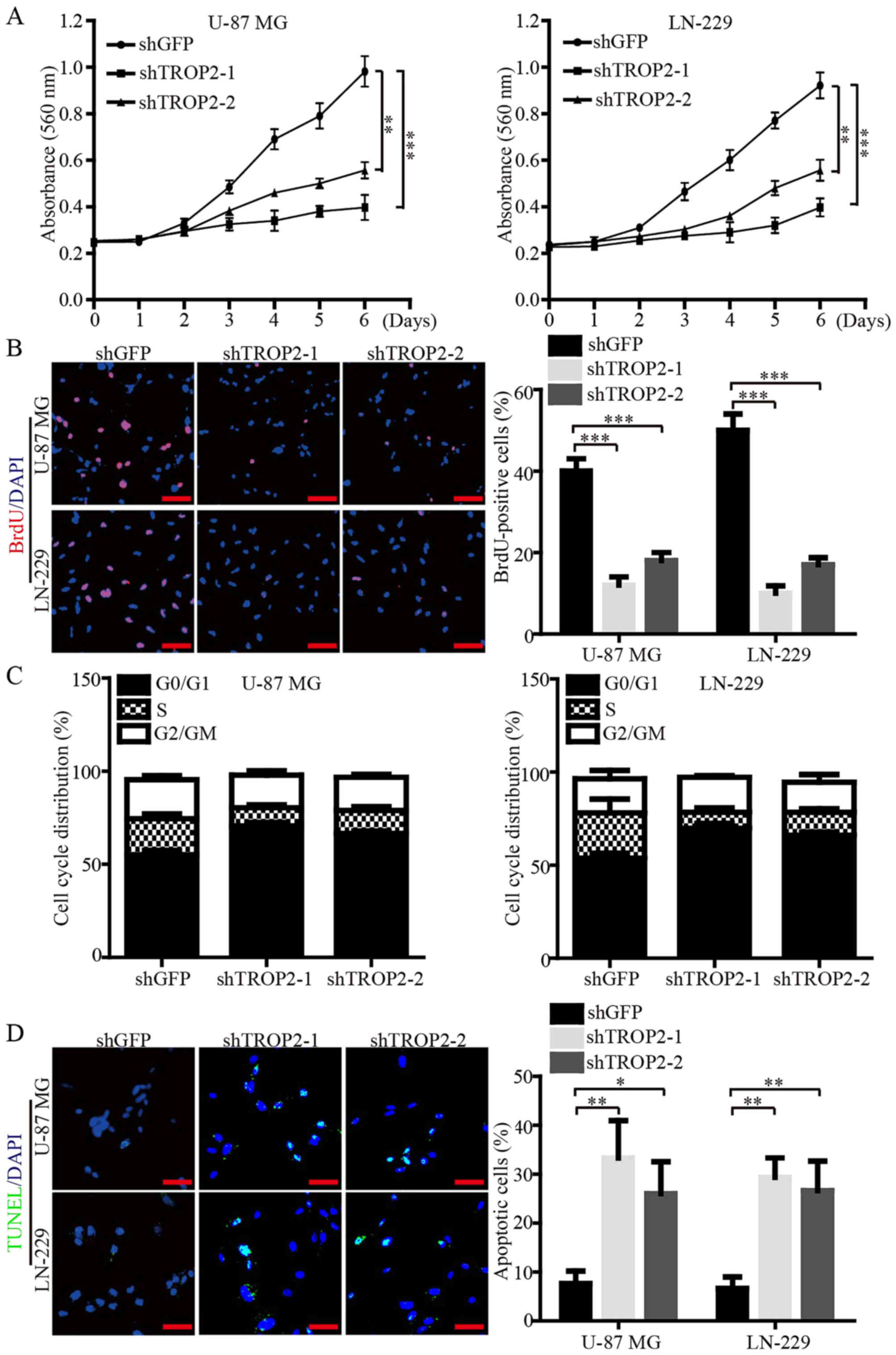
We subsequently investigated the biological role of
TROP2 in cell proliferation and viability of glioblastoma cells via
MTT and BrdU assays. The data demonstrated that the cell growth
curve and DNA synthesis of TROP2-knockdown cells were significantly
reduced when compared to the control cells (Fig. 3A and B). Flow cytometry was
performed to corroborate the data of proliferation, and the results
showed that TROP2 knockdown led to cell cycle arrest at the G1
phase (Fig. 3C). In addition,
immunofluorescence (IF) assays of TUNEL were performed to validate
that TROP2 knockdown significantly increased the cell apoptosis of
both U-87 MG and LN-229 cells (Fig.
3D). Collectively, these data demonstrated that TROP2 potently
promotes proliferation and viability of glioblastoma cells.
TROP2 promotes self-renewal and tumor
growth of glioblastoma cells in vitro and in vivo
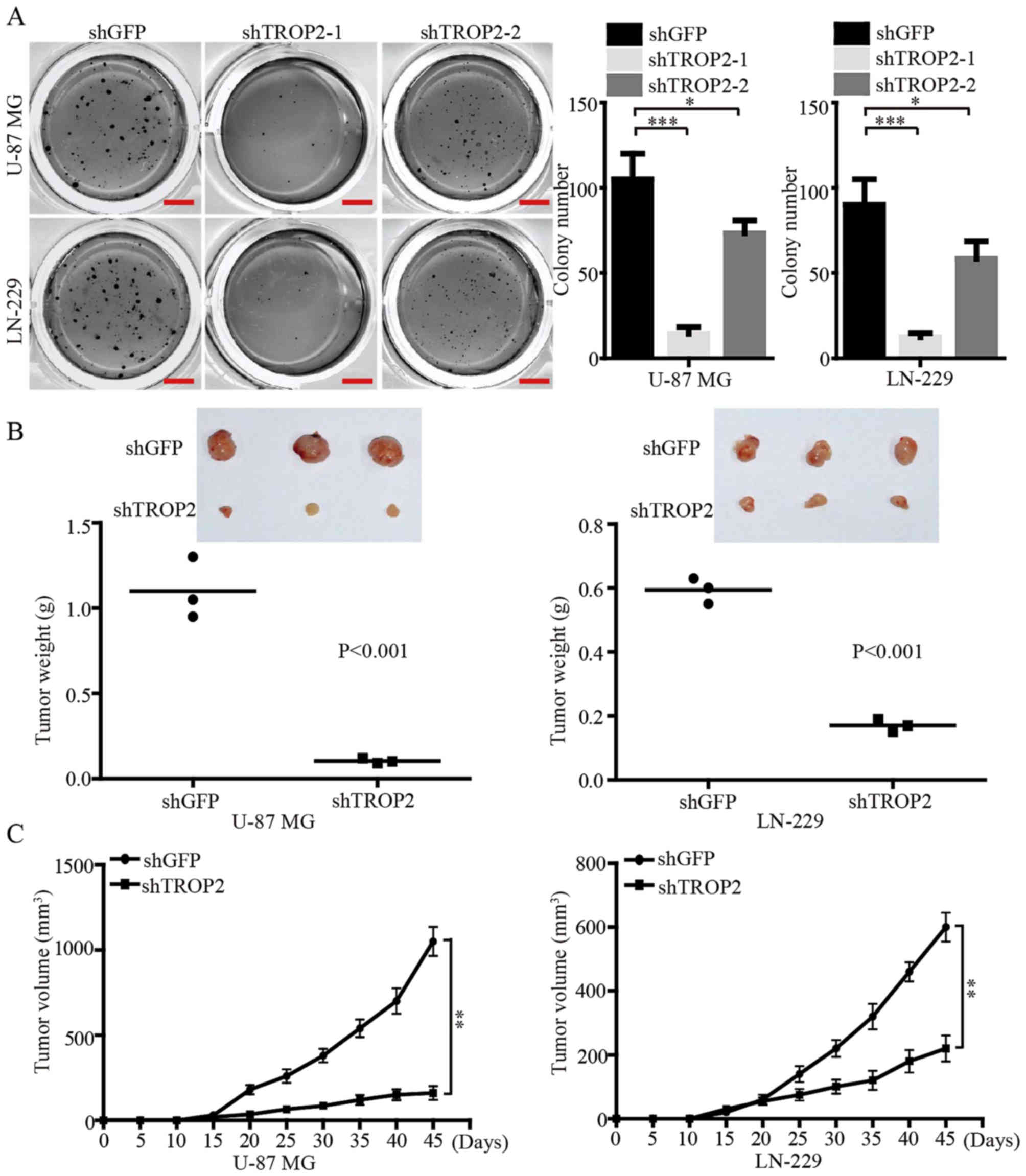
Soft agar colony formation assays were performed to
investigate the effects of TROP2 expression on self-renewal of
glioblastoma cells in vitro. The results demonstrated that
the number and size of colonies in the TROP2-konckdown U-87 MG and
LN-229 cells were reduced compared with the control cells (Fig. 4A). We also found that knockdown of
TROP2 using shTROP2-1 exhibited a more powerful inhibitory effect
on the colony formation of glioblastoma cells. Therefore, the
highly effective shTROP2-1 was used as a representative
TROP2-knockdown means for the following experiments. In order to
confirm the effects of TROP2 in vivo, a subcutaneous
xenograft experiment was performed. Consistent with the in
vitro results, the growth of tumors derived from
TROP2-knockdown cells was significantly retarded compared with the
controls. At the study end point, the knockdown of TROP2 resulted
in significantly decreased tumor weight and volume compared with
the relevant controls (Fig. 4B and
C). Taken together, these data suggest that TROP2 potently
promotes the tumor growth of glioblastoma cells.
TROP2 mediates oncogenic effects by
promoting the activation of the JAK2/STAT3 pathway
To explore the mechanisms underlying changes in
TROP2-knockdown glioblastoma cells, we performed quantitative
RT-PCR assays to compare the mRNA expression pattern of
TROP2-knockdown glioblastoma cells with the control cells. TROP2
knockdown attenuated the expression of several genes involved in
cell proliferation, apoptosis, migration and invasion, including
cyclin D1 (CCND1), survivin, matrix metalloproteinase 2 (MMP2) and
vascular endothelial growth factor (VEGF), which are generally
considered as downstream molecules of the JAK2/STAT3 signaling
pathway (Fig. 5A). Next, western
blot analysis showed decreased phosphorylation of
JAK2Tyr1007/1008 and STAT3Tyr705 in the
TROP2-knockdown glioblastoma cells and xenograft tumors. CCND1,
survivin, MMP2 and VEGF expression levels were observably decreased
in the TROP2-knockdown glioblastoma cells and xenograft tumors
(Fig. 5B and C). To determine the
clinical relevance, we carried out immunohistochemical analysis for
level of TROP2 and phosphorylation of JAK2Tyr1007/1008
and STAT3Tyr705 in human GBM specimens and normal brain
tissues. The GBM samples had high TROP2 expression, which
correlated with high JAK2 and STAT3 activation (Fig. 5D). These results suggest that TROP2
promotes cell proliferation and metastasis of glioblastoma cells by
activating the JAK2/STAT3 pathway.
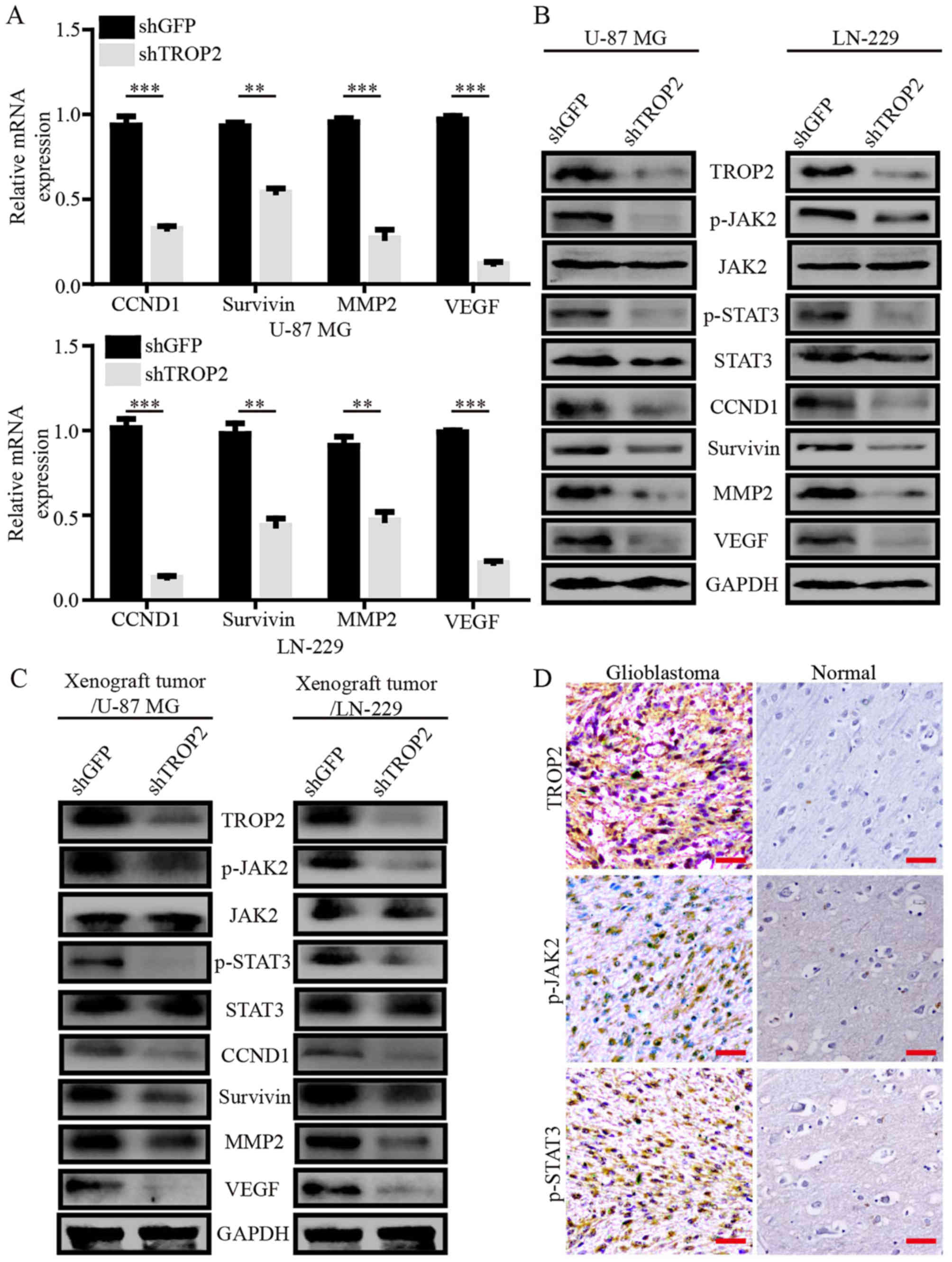 | Figure 5.TROP2 mediates oncogenic effects by
promoting the activation of the JAK2/STAT3 pathway. (A) Relative
mRNA expression of STAT3 target genes in TROP2-knockdown or control
U-87 MG cells and LN-229 cells. Transcript levels were normalized
to GAPDH. (B and C) Western blot analysis of
JAK2Tyr1007/1008, total JAK2, STAT3Tyr705,
total STAT3, CCND1, survivin, MMP2 and VEGF levels in
TROP2-silenced U-87 MG and LN-229 cells and xenograft tumors, GAPDH
was used as loading control. (D) Representative IHC staining for
TROP2, p-JAK2, and p-STAT3 in GBM samples from patients. All data
are shown as the means ± SD; **P<0.01, ***P<0.001. All
P-values are based on control vs. treatment group. TROP2,
trophoblast cell surface antigen 2; GBM, glioblastoma. |
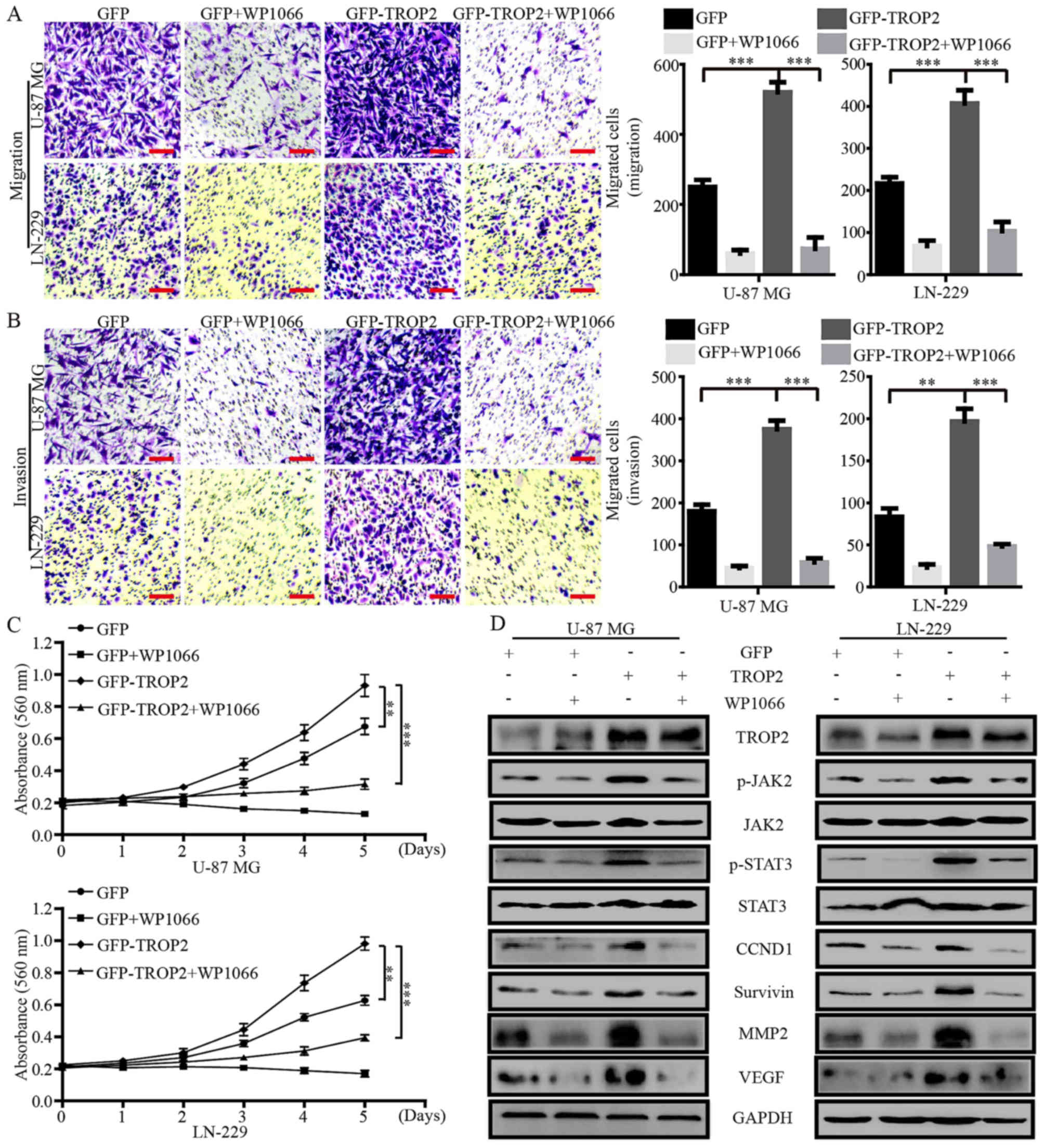
Blocking the activation of JAK2/STAT3
signaling by WP1066 negates the effects of TROP2
overexpression
WP1066 is a potent JAK2/STAT3 inhibitor that
inhibits the proliferation and metastasis of cancer cells. To
confirm whether the effects of TROP2 on glioblastoma cells were
JAK2/STAT3 dependent, we treated TROP2-overexpressing glioblastoma
cells and their controls cells with WP1066 (10 µM). Migration and
invasion assays showed that WP1066 significantly decreased the
accelerative effects of TROP2 on cell migration and invasion
(Fig. 6A and B). MTT assays showed
that WP1066 significantly reduced the promotive effects of TROP2 on
cell proliferation (Fig. 6C).
Western blot analysis demonstrated that the levels of JAK2 and
STAT3 phosphorylation (JAK2Tyr1007/1008 and
STAT3Tyr705) were altered in proportion to the
alteration of total JAK2 and STAT3 protein levels in glioblastoma
cells. In addition, CCND1, survivin, MMP2 and VEGF levels were
markedly increased by TROP2 overexpression and were reduced by
WP1066 treatment (Fig. 6D).
Collectively, these data demonstrated that TROP2 promotes the
activation of the JAK2/STAT3 signaling pathway.
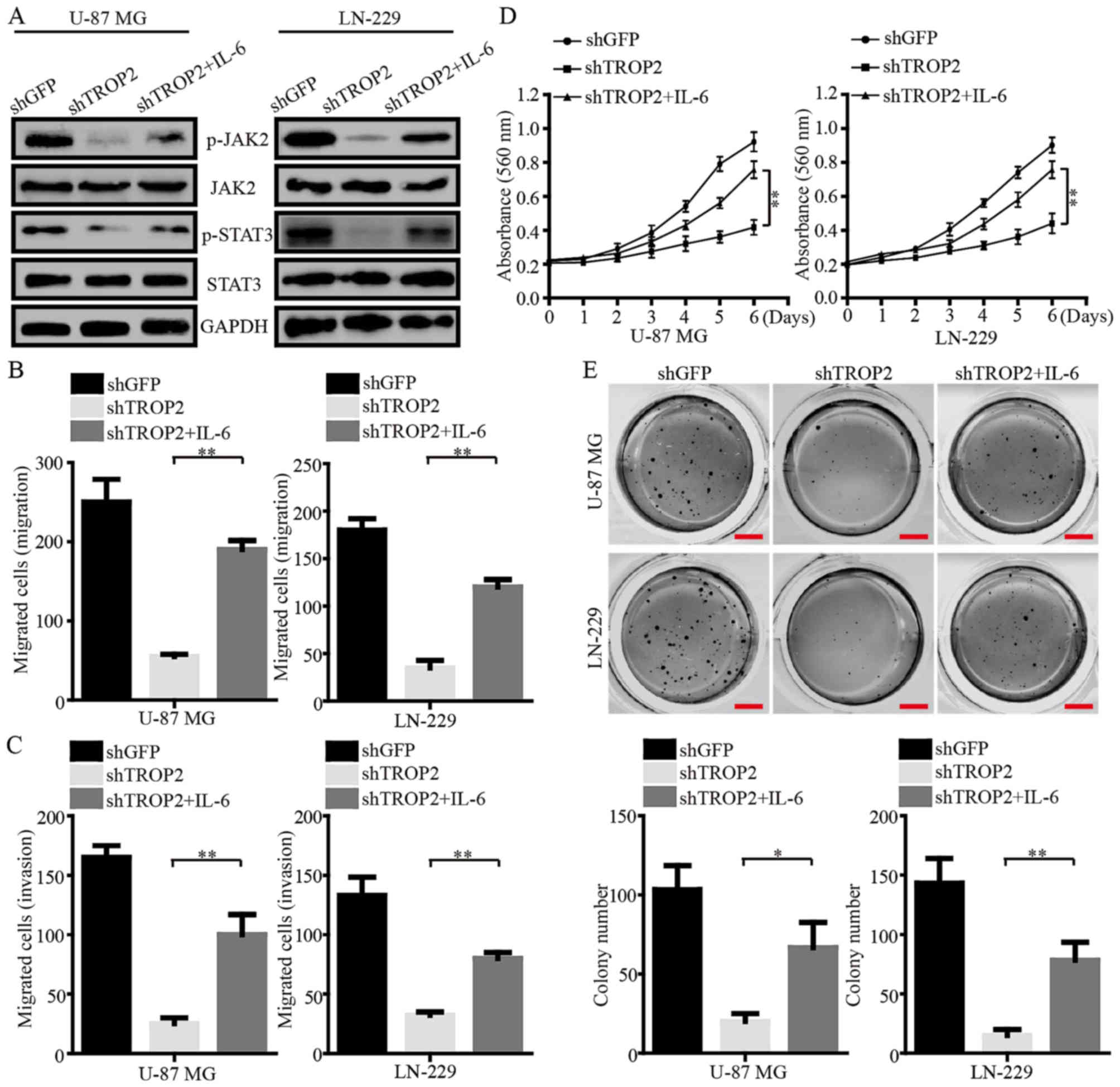
Exogenous IL-6 reverses phenotype
changes of TROP2-silencing glioblastoma cells
IL-6, a potent activator of JAK2/STAT3, was used to
further determine the role of TROP2 in the regulation of the
JAK2/STAT3 pathway. Western blot assays demonstrated that the
phosphorylation of JAK2Tyr1007/1008 and
STAT3Tyr705 were increased in TROP2-knockdown
glioblastoma cells after the cells were exposed to IL-6 (Fig. 7A). The migration and invasion of
TROP2-knockdown cells were clearly increased after IL-6 exposure
(Fig. 7B and C). In addition, MTT
assays showed that cell viability was increased after IL-6 exposure
(Fig. 7D). Furthermore, IL-6
exposure led to larger and more numerous colonies formed by
TROP2-silenced U-87 MG and LN-229 cells than the colonies formed by
these cells without exposure to IL-6 (Fig. 7E). Collectively, these data strongly
suggest that TROP2 functions in an oncogenic role in glioblastoma
by activating the JAK2/STAT3 signaling pathway.
Discussion
Glioblastoma is the most common and aggressive
malignant cancer, exhibiting high rates of proliferation and
infiltration. Despite advances in multimodal treatments involving
surgical resection, chemotherapy and radiotherapy, patients with
glioblastoma have a median overall survival (OS) that rarely
exceeds two years, and the rate of 5-year survival is <5%
(32). Molecular-targeted therapy
has become a promising method in cancer treatment, especially for
gastric and lung cancer (33,34).
However, a lack of suitable molecular-targeted drugs is perhaps the
key challenge for combating glioblastoma. TROP2, a single
transmembrane domain protein, has been found to be highly expressed
in various human carcinomas, including breast cancer, lung cancer,
gastric cancer, cervical cancer, ovarian cancer and nasopharyngeal
carcinoma (35). Previous studies
concerning the biological roles of TROP2 in human carcinomas were
mainly focused on the adhesion between cancer cells, and were also
involved in cell signal transduction and the growth of cancer cells
(11). However, the biological
function and molecular mechanism of TROP2 in glioblastoma have not
been fully elucidated, especially in regards to the cell
proliferation and metastasis of glioblastoma cells.
In the present study, we first used
immunohistochemistry and western blot analysis to assess the TROP2
protein pattern in glioblastoma tissues and glioblastoma cell
lines. We observed that TROP2 was highly expressed in human
glioblastoma samples and the levels of TROP2 were positively
associated with the overall survival of glioblastoma patients. In
addition, TROP2 was also overexpressed in glioblastoma cell lines
compared with that noted in human normal astroglial cells (SVGP12).
As to the biological study, we observed that silencing of TROP2 in
glioblastoma cells inhibited cell proliferation and metastasis of
glioblastoma cells. These results suggest that TROP2 functions as
an oncogene in glioblastoma cells.
The molecular mechanisms of TROP2 in carcinogenesis
and tumor progression remain largely unknown. TROP2 has been shown
to regulate the activation of several cancer-related factors, such
as retinoblastoma (RB) protein, Jun and nuclear factor (NF)-κB
(36). Our results demonstrated
that TROP2 knockdown attenuated the expression of various genes
involved in cell proliferation (CCND1), apoptosis (survivin),
metastasis (MMP2) and angiogenesis (VEGF), all of which are
downstream molecules of the JAK2/STAT3 signaling pathway. We found
that TROP2 knockdown was associated with the inhibition of JAK2 and
STAT3 phosphorylation in glioblastoma cells. In addition, the
effects of TROP2 overexpression on glioblastoma cells was negated
by JAK2/STAT3 inhibitor WP1066. IL-6, as a potent activator of
JAK2/STAT3 signaling, increased the phosphorylation of JAK2 and
STAT3 in TROP2-knockdown glioblastoma cells and regulated
phenotypic changes in these cells. Thus, we demonstrated for the
first time that TROP2 could promote the activation of the JAK2/
STAT3 signaling pathway.
In conclusion, we discovered a novel mechanism by
which TROP2 activates the JAK2/STAT3 signaling pathway to promote
glioblastoma growth and metastasis. These results provide new
insights into the functions of TROP2 and indicate that TROP2 is a
promising biomarker and therapeutic target for glioblastoma.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Key
Research and Development Program of China (nos. 2016YFC1302204 and
2017YFC1308600), the Fundamental Research Funds for the Central
Universities (XDJK2017D186), the Graduate Scientific Research
Foundation of Chongqing (CYB17068), the National Natural Science
Foundation of China (81672502) and the Chongqing University
Innovation Team Building Program funded projects
(CXTDX201601010).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JH was responsible for the collection and assembly
of data, data analysis and interpretation, and manuscript writing.
AL and QD carried out the collection and assembly of data and the
data analysis. GZ was responsible for the collection and assembly
of data, and revised the manuscript. XH carried out the collection
and assembly of data, wrote and reviewed the manuscript. HC was
responsible for the conception and design of the study, data
analysis and interpretation, manuscript revision and financial
support. All authors read and approved the final manuscript and
agree to be accountable for all aspects of the research in ensuring
that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
All animal studies were approved by the Animal Care
and Use Committee of Southwest University, and carried out in
conformity to the Guide for the Care and Use of Laboratory Animals
(Ministry of Science and Technology of China, 2006). Primary human
tumor specimen collection was granted prior approval from the
Ethics Committee of Daping Hospital, Chongqing, China. Tissue
analysis was approved by the Ethics Committee of Southwest
University of China, Chongqing, China. All of the patients provided
informed consent to participate.
Patient consent for publication
Not applicable.
Competing interests
The authors declare no competing interests, and all
authors confirm its accuracy.
References
|
1
|
Fuller GN and Scheithauer BW: The 2007
Revised World Health Organization (WHO) Classification of Tumours
of the Central Nervous System: Newly codified entities. Brain
Pathol. 17:304–307. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ostrom QT, Gittleman H, Fulop J, Liu M,
Blanda R, Kromer C, Wolinsky Y, Kruchko C and Barnholtz-Sloan JS:
CBTRUS Statistical Report: Primary brain and central nervous system
tumors diagnosed in the United States in 2008–2012. Neuro-oncol. 17
Suppl 4:iv1–iv62. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lee I: Advances in surgical approaches in
glioblastoma (GBM). Linchuang Zhongliuxue Zazhi. 6:422017.
|
|
4
|
Sanai N, Polley MY, McDermott MW, Parsa AT
and Berger MS: An extent of resection threshold for newly diagnosed
glioblastomas. J Neurosurg. 115:3–8. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lacroix M, Abi-Said D, Fourney DR,
Gokaslan ZL, Shi W, DeMonte F, Lang FF, McCutcheon IE, Hassenbusch
SJ, Holland E, et al: A multivariate analysis of 416 patients with
glioblastoma multiforme: Prognosis, extent of resection, and
survival. J Neurosurg. 95:190–198. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wen PY and Kesari S: Malignant gliomas in
adults. N Engl J Med. 359:492–507. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hou J, Deng Q, Zhou J, Zou J, Zhang Y, Tan
P, Zhang W and Cui H: CSN6 controls the proliferation and
metastasis of glioblastoma by CHIP-mediated degradation of EGFR.
Oncogene. 36:1134–1144. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lipinski M, Parks DR, Rouse RV and
Herzenberg LA: Human trophoblast cell-surface antigens defined by
monoclonal antibodies. Proc Natl Acad Sci USA. 78:5147–5150. 1981.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lin JC, Wu YY, Wu JY, Lin TC, Wu CT, Chang
YL, Jou YS, Hong TM and Yang PC: TROP2 is epigenetically
inactivated and modulates IGF-1R signalling in lung adenocarcinoma.
EMBO Mol Med. 4:472–485. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ripani E, Sacchetti A, Corda D and Alberti
S: Human Trop-2 is a tumor-associated calcium signal transducer.
Int J Cancer. 76:671–676. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shvartsur A and Bonavida B: Trop2 and its
overexpression in cancers: Regulation and clinical/therapeutic
implications. Genes Cancer. 6:84–105. 2015.PubMed/NCBI
|
|
12
|
Cubas R, Li M, Chen C and Yao Q: Trop2: A
possible therapeutic target for late stage epithelial carcinomas.
Biochim Biophys Acta. 1796:309–314. 2009.PubMed/NCBI
|
|
13
|
Ohmachi T, Tanaka F, Mimori K, Inoue H,
Yanaga K and Mori M: Clinical significance of TROP2 expression in
colorectal cancer. Clin Cancer Res. 12:3057–3063. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zheng Z and Dong XJ: Clinical value of
serum trophoblast cell surface protein 2 (TROP2) antibody in
non-small-cell lung cancer patients. Biomarkers. 21:1–4. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ning S, Liang N, Liu B, Chen X, Pang Q and
Xin T: TROP2 expression and its correlation with tumor
proliferation and angiogenesis in human gliomas. Neurol Sci.
34:1745–1750. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao W, Zhu H, Zhang S, Yong H, Wang W,
Zhou Y, Wang B, Wen J, Qiu Z, Ding G, et al: Trop2 is overexpressed
in gastric cancer and predicts poor prognosis. Oncotarget.
7:6136–6145. 2016.PubMed/NCBI
|
|
17
|
Liu X, Li S and Yi F: Trop2 gene: A novel
target for cervical cancer treatment. J Cancer Res Clin Oncol.
140:1331–1341. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Son S, Shin S, Rao NV, Um W, Jeon J, Ko H,
Deepagan VG, Kwon S, Lee JY and Park JH: Anti-Trop2
antibody-conjugated bioreducible nanoparticles for targeted triple
negative breast cancer therapy. Int J Biol Macromol. 110:406–415.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mao Y, Wang X, Zheng F, Wang C, Tang Q,
Tang X, Xu N, Zhang H, Zhang D, Xiong L, et al: The
tumor-inhibitory effectiveness of a novel anti-Trop2 Fab conjugate
in pancreatic cancer. Oncotarget. 7:24810–24823. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chang Q, Bournazou E, Sansone P, Berishaj
M, Gao SP, Daly L, Wels J, Theilen T, Granitto S, Zhang X, et al:
The IL-6/JAK/Stat3 feed-forward loop drives tumorigenesis and
metastasis. Neoplasia. 15:848–862. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Heinrich PC, Behrmann I, Haan S, Hermanns
HM, Müller-Newen G and Schaper F: Principles of interleukin
(IL)-6-type cytokine signalling and its regulation. Biochem J.
374:1–20. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kanda N, Seno H, Konda Y, Marusawa H,
Kanai M, Nakajima T, Kawashima T, Nanakin A, Sawabu T, Uenoyama Y,
et al: STAT3 is constitutively activated and supports cell survival
in association with survivin expression in gastric cancer cells.
Oncogene. 23:4921–4929. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hajimoradi M, Mohammad Hassan Z, Ebrahimi
M, Soleimani M, Bakhshi M, Firouzi J and Samani FS: STAT3 is
overactivated in gastric cancer stem-like cells. Cell J.
17:617–628. 2016.PubMed/NCBI
|
|
24
|
Rokavec M, Öner MG, Li H, Jackstadt R,
Jiang L, Lodygin D, Kaller M, Horst D, Ziegler PK, Schwitalla S, et
al: Corrigendum. IL-6R/STAT3/miR-34a feedback loop promotes
EMT-mediated colorectal cancer invasion and metastasis. J Clin
Invest. 125:13622015. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chung SS, Giehl N, Wu Y and Vadgama JV:
STAT3 activation in HER2-overexpressing breast cancer promotes
epithelial-mesenchymal transition and cancer stem cell traits. Int
J Oncol. 44:403–411. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cutolo M and Meroni M: Clinical utility of
the oral JAK inhibitor tofacitinib in the treatment of rheumatoid
arthritis. J Inflamm Res. 6:129–137. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vyas D, O'Dell KM, Bandy JL and Boyce EG:
Tofacitinib: The First Janus Kinase (JAK) inhibitor for the
treatment of rheumatoid arthritis. Ann Pharmacother. 47:1524–1531.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hashizume M, Tan SL, Takano J, Ohsawa K,
Hasada I, Hanasaki A, Ito I, Mihara M and Nishida K: Tocilizumab, a
humanized anti-IL-6R antibody, as an emerging therapeutic option
for rheumatoid arthritis: Molecular and cellular mechanistic
insights. Int Rev Immunol. 34:265–279. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang D, Wang F, Pang Y, Zhao E, Zhu S,
Chen F and Cui H: ALG2 regulates glioblastoma cell proliferation,
migration and tumorigenicity. Biochem Biophys Res Commun.
486:300–306. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huang MY, Xuan F, Liu W and Cui HJ: MINA
controls proliferation and tumorigenesis of glioblastoma by
epigenetically regulating cyclins and CDKs via H3K9me3
demethylation. Oncogene. 36:387–396. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Stupp R, Hegi ME, Mason WP, van den Bent
MJ, Taphoorn MJ, Janzer RC, Ludwin SK, Allgeier A, Fisher B,
Belanger K, et al: European Organisation for Research and Treatment
of Cancer Brain Tumour and Radiation Oncology Groups; National
Cancer Institute of Canada Clinical Trials Group: Effects of
radiotherapy with concomitant and adjuvant temozolomide versus
radiotherapy alone on survival in glioblastoma in a randomised
phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet
Oncol. 10:459–466. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang L, Yuan H, Li Y and Han Y: The role
of HER3 in gastric cancer. Biomed Pharmacother. 68:809–812. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Smith J: Erlotinib: Small-molecule
targeted therapy in the treatment of non-small-cell lung cancer.
Clin Ther. 27:1513–1534. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu P, Zhao Y, Liu K, Lin S, Liu X, Wang M,
Yang P, Tian T, Zhu YY and Dai Z: Prognostic role and clinical
significance of trophoblast cell surface antigen 2 in various
carcinomas. Cancer Manag Res. 9:821–837. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Guerra E, Trerotola M, Aloisi AL, Tripaldi
R, Vacca G, La Sorda R, Lattanzio R, Piantelli M and Alberti S: The
Trop-2 signalling network in cancer growth. Oncogene. 32:1594–1600.
2013. View Article : Google Scholar : PubMed/NCBI
|