Introduction
Lung cancer is a malignant tumor with the highest
morbidity and mortality in the world. It is a major disease
threatening human life and health (1). The world is undergoing an increasing
cancer burden (1). This was
suggested in World Cancer Reports from the International Agency for
Research on Cancer (IARC) of WTO released in 2014. Lung cancer
cases ranked first among all newly diagnosed cancers in 2012
(2). This figure is approximately
1.8 million. It accounts for 13% of the total number of common
cancer cases. In addition, lung cancer is also the leading cause of
cancer-related mortality. This number was approximately 1.6 million
in 2015, which accounted for 19.4% of the total number of deaths
(2). Of them, over 1/3 were Chinese
cases (2).
It was discovered in recent years that circulatory
miRNAs can serve as disease markers (3). This has aroused the interests of
numerous scientific researchers. In particular, close attention has
been paid to miRNAs as tumor diagnostic markers. miRNAs are a class
of endogenous non-coding small molecule RNAs. They are highly
conserved in evolution. They consist of approximately 18–23 basic
groups in length (4). miRNAs have
been verified in reports to play a certain role in cancer
pathogenesis, genesis and development. In addition, they exert
functions that are similar to oncogenes or tumor-suppressor genes
(5).
Epidermal growth factor receptor (EGFR) is a
transmembrane tyrosine kinase receptor. It is approximately 170 kb
in molecular weight (6). It
consists of three parts, namely, the extracellular domain, the
transmembrane domain and the intracellular domain. The
intracellular domain is also the EGFR-tyrosine kinase domain
(7). It is an important component
for regulating tumor cell proliferation, invasion, angiogenesis,
adhesion, metastasis and apoptosis. EGFR binds with its ligand in
the extracellular domain (8). This
can produce an EGFR dimer and induce intracellular phosphorylation
(8). Thus, it can activate a series
of downstream signaling pathways (9). Of these, the phosphoinositide 3-kinase
(PI3K) pathway is a canonical pathway. PI3K is the major downstream
effector of receptor tyrosine kinase and G-protein coupled receptor
(9). It can produce activated
serine/threonine protein kinase AKT and its downstream effector
phospholipid. Consequently, it can transduce signals from all
growth factors and cytokines to intracellular messenger (9). This is closely associated with tumor
cell proliferation and survival. EGFR is notably correlated with
NSCLC. The downstream PI3K/AKT pathway that it activates is
abnormally expressed in numerous tumors including breast cancer,
prostate cancer and gastric cancer (10). It was demonstrated that miRNA-223
suppressed cervical cancer cell growth via targeting the
EGFR/AKT2/CCND1 pathway (11).
Herein, our study further explored the molecular mechanisms of
miRNA-223 in NSCLC.
Materials and methods
Patients
We analyzed the prospectively collected data of
NSCLC patients and healthy volunteers between March 2016 and May
2016 at the Tsinghua Changgung Hospital. The study was approved by
Jining First People's Hospital. Basic data of all patients with
NSCLC were collected and are documented in Table I. Ten milliliters of peripheral
blood was centrifuged at 1,000 × g for 10 min at 4°C, and serum was
collected and saved at −80°C.
 | Table I.Characteristics of the NSCLC
patients. |
Table I.
Characteristics of the NSCLC
patients.
| Variables | NSCLC cases
(n=6) | Healthy subjects
(n=6) |
|---|
| Age (years) |
| ≤55 | 4 | 3 |
|
>55 | 2 | 3 |
| Sex |
|
Female | 0 | 0 |
| Male | 6 | 6 |
| Tumor size (cm) |
| ≤3.0 | 2 | 0 |
|
>3.0 | 4 | 0 |
| Edmondson grade |
| I | 0 | 0 |
| II | 2 | 0 |
| III | 4 | 0 |
Cell culture and transfection
Human NSCLC cell line A549 was purchased from the
American Type Culture Collection (ATCC) (Manassas, VA, USA) and was
cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented
with 10% fetal bovine serum (FBS) (both from Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) at 37°C in a 5% CO2
incubator. Anti-miRNA-223, miRNA-223 and control negative mimics
were designed and obtained from GenePharma (Shanghai, Beijing,
China). A549 cells were transfected with anti-miRNA-223, miRNA-223
and the control negative mimics using Invitrogen™ Lipofectamine
2000 (Thermo Fisher Scientific, Inc.). AG1478, an EGFR special
inhibitor and LY294002, a PI3K special inhibitor were purchased
from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
qRT-PCR analysis
Total RNA was extracted from prepared cells or serum
using TRizol reagent (Invitrogen; Thermo Fisher Scientific, Inc.).
cDNA was synthesized with the PrimiScript RT reagent kit (Takara,
Shiga, Japan). qRT-PCR analysis was performed using SYBR Green PCR
Master Mix (PE Applied Biosystems; Thermo Fisher Scientific, Inc.)
in the Applied Biosystems 7900HT real-time PCR machine (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The probes for
miRNA-223 were: 5′-GCGTGTATTTGACAAGCTGAGTT-3′ and
5′-GTGTCAGTTTGTCAAATACCCCA-3′ and U6: 5′-CCGCCCGCCGCCAGGCCCC-3′ and
5′-ATATGGAACGCTTCACGAATT-3′. The reaction conditions were as
follows: hot start at 94°C for 5 min; 40 cycles of 30 sec at 95°C,
30 sec at 60°C and 30 sec at 72°C, and 10 min at 72°C. The relative
expression of miRNAs was calculated with the 2−∆∆Ct
method.
Microarray analysis
RNA (200 ng) was amplified using Cy3 using the Low
Input Quick-Amp Labeling kit (both from Agilent Technologies, Santa
Clara, CA, USA). The cRNAs were subjected to hybridization into
Agilent SurePrint G3 Mouse GE 8X 60 K Microarray Chips (Agilent
Technologies). Data were quantified by Feature Extraction 10.5.1.1
image analysis software (Agilent Technologies).
Cell proliferation assay and lactate
dehydrogenase (LDH)
At 24, 48, 72 h after transfection, cell
proliferation was measured using the MTT assay and incubation was
carried out for 4 h at 37 °C. DMSO was added into the cells and
then incubated for 20 min. Optical density (OD) value was read
using a microplate reader (Bio-Rad Laboratories, Hercules, CA, USA)
at 490 nm.
LDH activity was measured using LDH activity kits
(Beyotime Biotechnology, Co., Ltd., Shanghai, China). Optical
density (OD) value was read using a microplate reader (Bio-Rad
Laboratories) at 450 nm.
Cell apoptosis assay and caspase-3/9
activity assay
Seventy-two hours after transfection, apoptotic
cells were stained with an Annexin V-FITC/PI double staining
apoptosis detection kit (BD Biosciences, Franklin Lakes, NJ, USA).
Apoptotic cells were assessed by flow cytometry (BD FACSCalibur, BD
Biosciences).
At 72 h after transfection, cells were lysed in RIPA
buffer (Beyotime Biotechnology, Co., Ltd.) and protein content was
measured using BCA assay (Beyotime Biotechnology, Co., Ltd.). Equal
amounts of protein were used to determine caspase-3/9 activity
using a caspase-3/9 activity kit (Beyotime Biotechnology, Co.,
Ltd.). Optical density (OD) value was read using a microplate
reader (Bio-Rad Laboratories) at 405 nm.
Dual luciferase reporter gene
assay
EGFR and miRNA-223 plasmids were co-transfected into
A549 cells using Lipofectamine 2000. Reporter gene assays were
performed using the Dual Luciferase Reporter Assay kit (Promega,
Madison, WI, USA) for 24 h.
Cell migration assay
A549 cells (5×105 cell/ml) were seeded on
24-well plates and were added to the upper chamber of each
migration well, and 500 µl of DMEM with 10% FBS was added to the
lower part for 16 h. Cells that migrated to the lower side were
fixed with 75% ice-alcohol for 30 min and stained with 1% crystal
violet solution for 1 h. A549 cells were fixed and counted under a
fluorescence microscope (Axio version II, Carl Zeiss Inc.,
Oberkochen, Germany).
Western blot analysis
Seventy-two hours after transfection, cells were
lysed in RIPA buffer (Beyotime Biotechnology, Co., Ltd.) and
protein content was measured using the BCA assay (Beyotime
Biotechnology, Co., Ltd.). Equal amounts of protein (40 µg) were
subjected to 8–12% SDS gel electrophoresis and transferred to a
Amersham polyvinylidene difluoride membrane (GE Healthcare Life
Sciences, Little Chalfont, UK). Western blot analysis was performed
with anti-Bax (sc-6236, 1:500), anti-EGFR (sc-365829, 1:500),
anti-PI3K (sc-7174, 1:500), anti-p-Akt (sc-135651, 1:500) and
anti-GAPDH (sc-25778, 1:2000, Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) antibodies at 4°C overnight. The membranes were
incubated with appropriate horseradish peroxidase-conjugated
secondary antibodies (1:2000, Santa Cruz Biotechnology, Inc.) for 1
h at room temperature and visualized with the chemiluminescent
detection method using the SuperSignal West Pico Substrate (Pierce;
Thermo Fisher Scientific, Inc.).
Statistical analyses
Data are expressed as mean ± SD. Data among groups
were analyzed using one-way analysis of variance followed by a
Tukey's post hoc test. P<0.05 was considered to indicate a
statistically significant result.
Results
Serum levels of miRNA-223 in NSCLC
patients
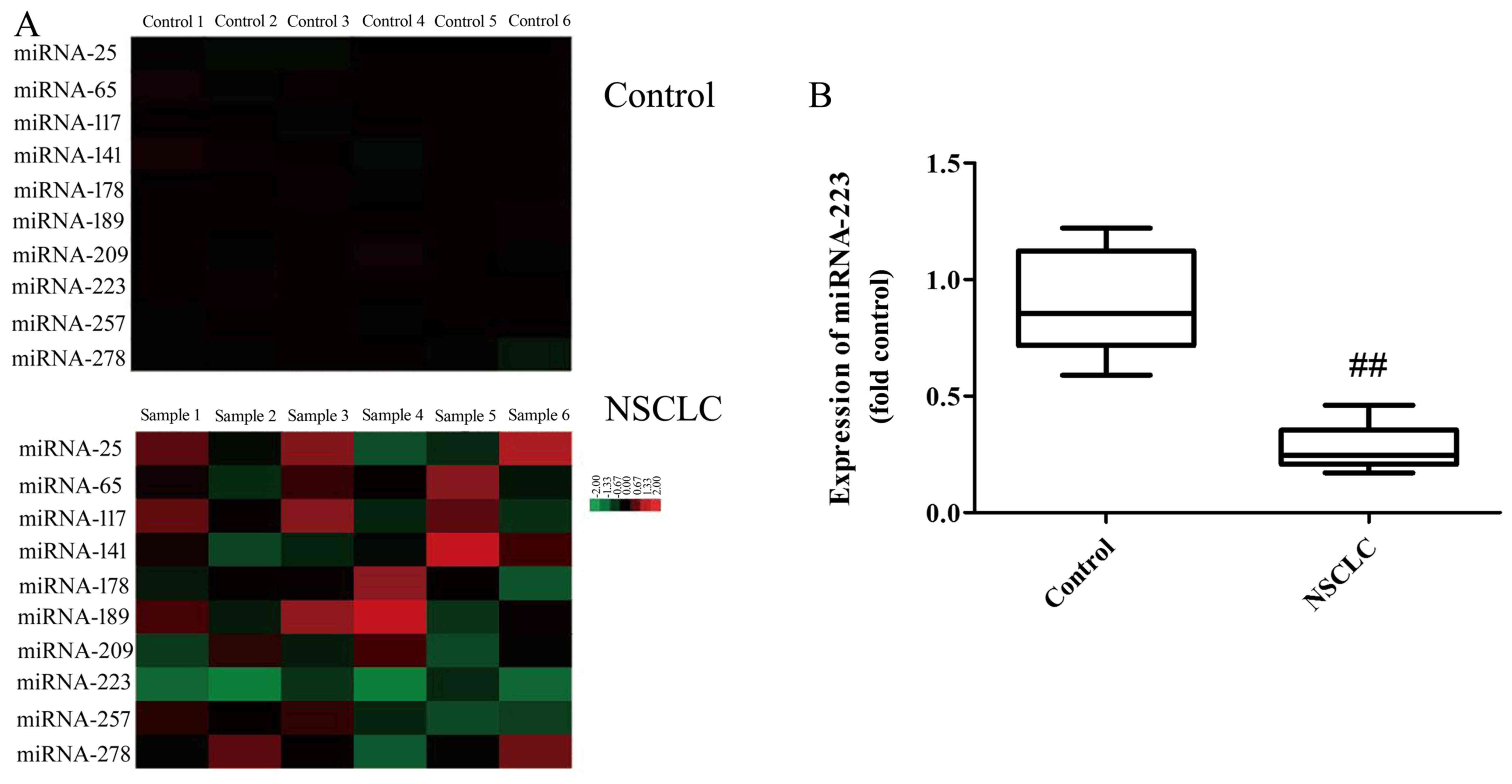
Gene chip shows that the serum levels of miRNA-223
in NSCLC patients were downregulated compared with the normal group
(Fig. 1A). Fig. 1B shows that the serum levels of
miRNA-223 in NSCLC patients were downregulated compared with the
normal group. Taken together, miRNA-223 may be an important
strategy for the anticancer effect in NSCLC.
miRNA-223 affects the proliferation
and migration of A549 cells
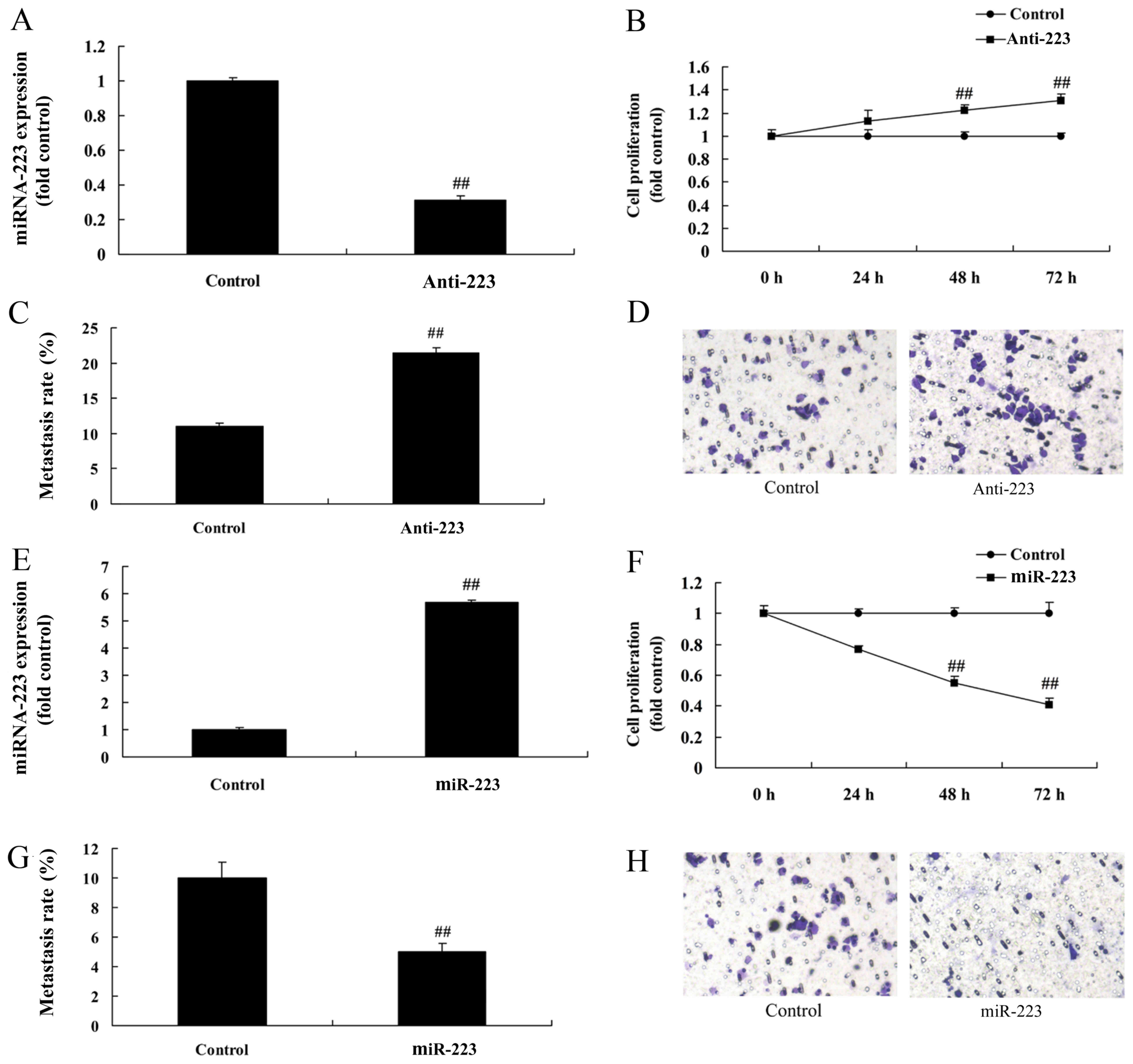
To confirm the function of miRNA-223, miRNA-223
expression was downregulated or upregulated using anti-miRNA-223
mimics or miRNA-223 mimics. As shown in Fig. 2A, miRNA-223 expression in A549 cells
was significantly inhibited following transfection with the
anti-miRNA-223 mimics compared with the control group.
Downregulation of miRNA-223 expression significantly promoted the
proliferation and migration of A549 cells compared with the
negative group (Fig. 2B-D). In
contrast, miRNA-223 mimics significantly upregulated miRNA-223
expression in A549 cells, which significantly reduced cell
proliferation and migration when compared with the negative group
(Fig. 2E-H).
miRNA-223 affects the apoptosis and
LDH activity in A549 cells
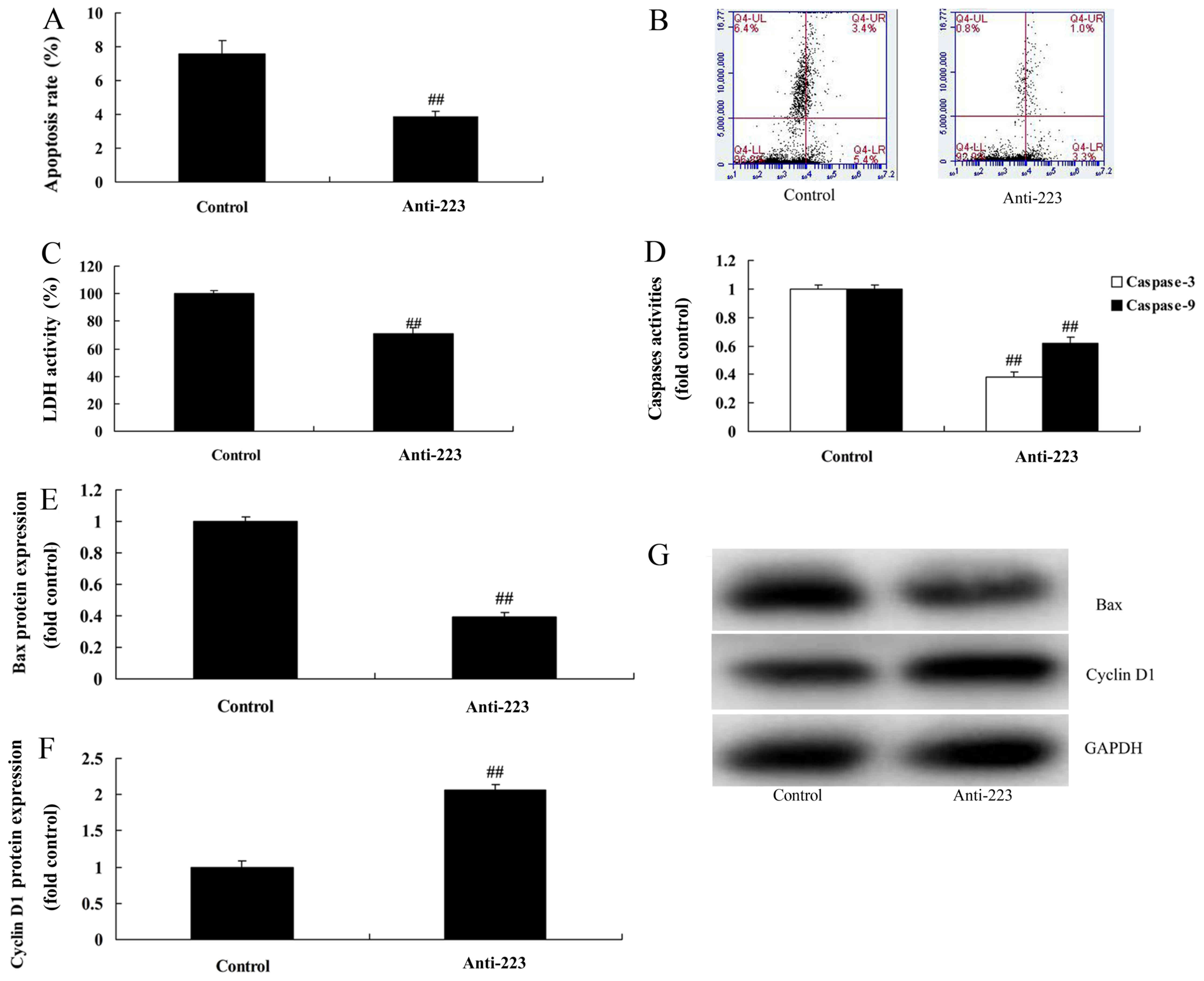
We subsequently found that miRNA-223 downregulation
evidently reduced the apoptosis rate, LDH activity and caspase-3/9
activity in A549 cells compared with the control group (Fig. 3A-D). Compared with the control
group, the protein expression of Bax and cyclin D1 was markedly
suppressed and induced by downregulation of miRNA-223 expression in
A549 cells, respectively (Fig.
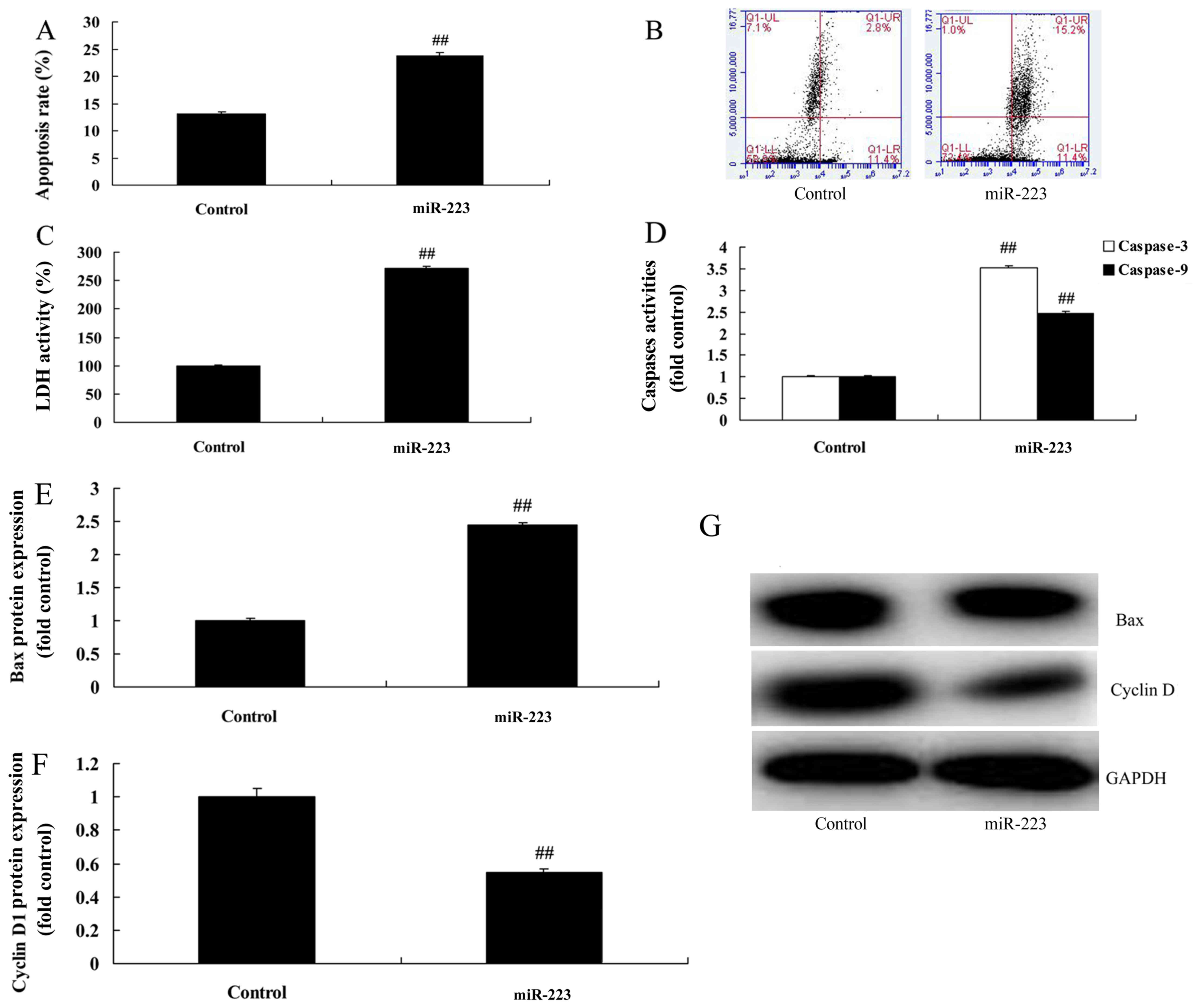
3E-G). However, upregulation of miRNA-223 evidently increased
the apoptosis rate, LDH activity and caspase-3/9 activity in the
A549 cells compared with the control group (Fig. 4A-D). Compared with the control
group, miRNA-223 upregulation significantly induced Bax protein
expression and suppressed cyclin D1 protein expression in the A549
cells, respectively (Fig.
4E-G).
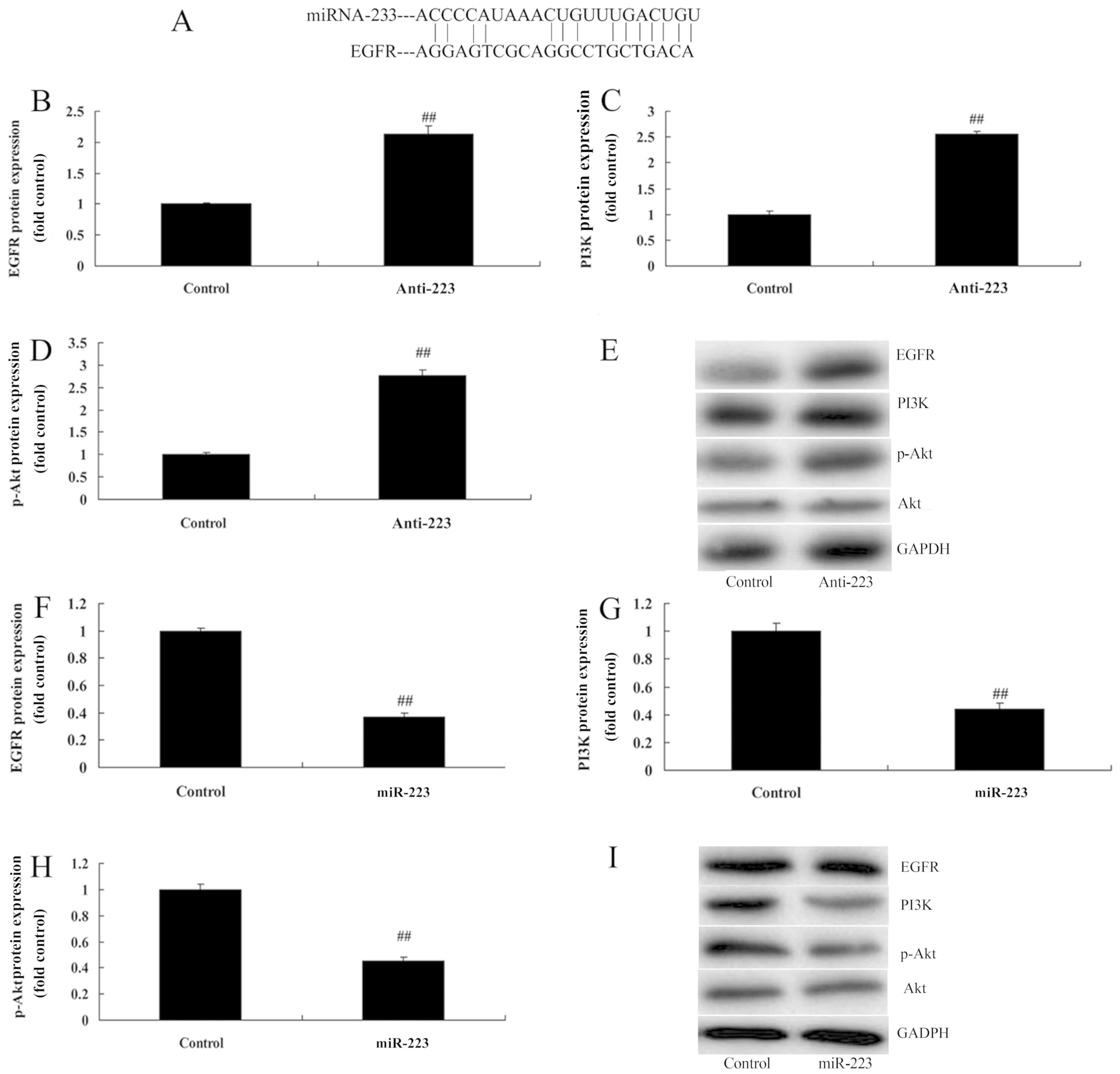
miRNA-223 affects EGFR, PI3K and p-Akt
protein expression in A549 cells
We determined whether miRNA-223 achieved its effects
on NSCLC through the EGFR/PI3K/AKT pathway. Schematic
representation of putative miRNA-223-binding sites in EGFR mRNA was
found (Fig. 5A). However,
downregulation of the expression of miRNA-223 dramatically induced
EGFR, PI3K and p-Akt protein expression in A549 cells compared with
the control group (Fig. 5B-E). In
comparison, upregulation of miRNA-223 significantly suppressed
EGFR, PI3K and p-Akt protein expression in A549 cells compared with
the control group (Fig. 5F-I).
There results indicate that miRNA-223 may be suppressed by the
EGFR/PI3K/AKT pathway in NSCLC for its anticancer effects.
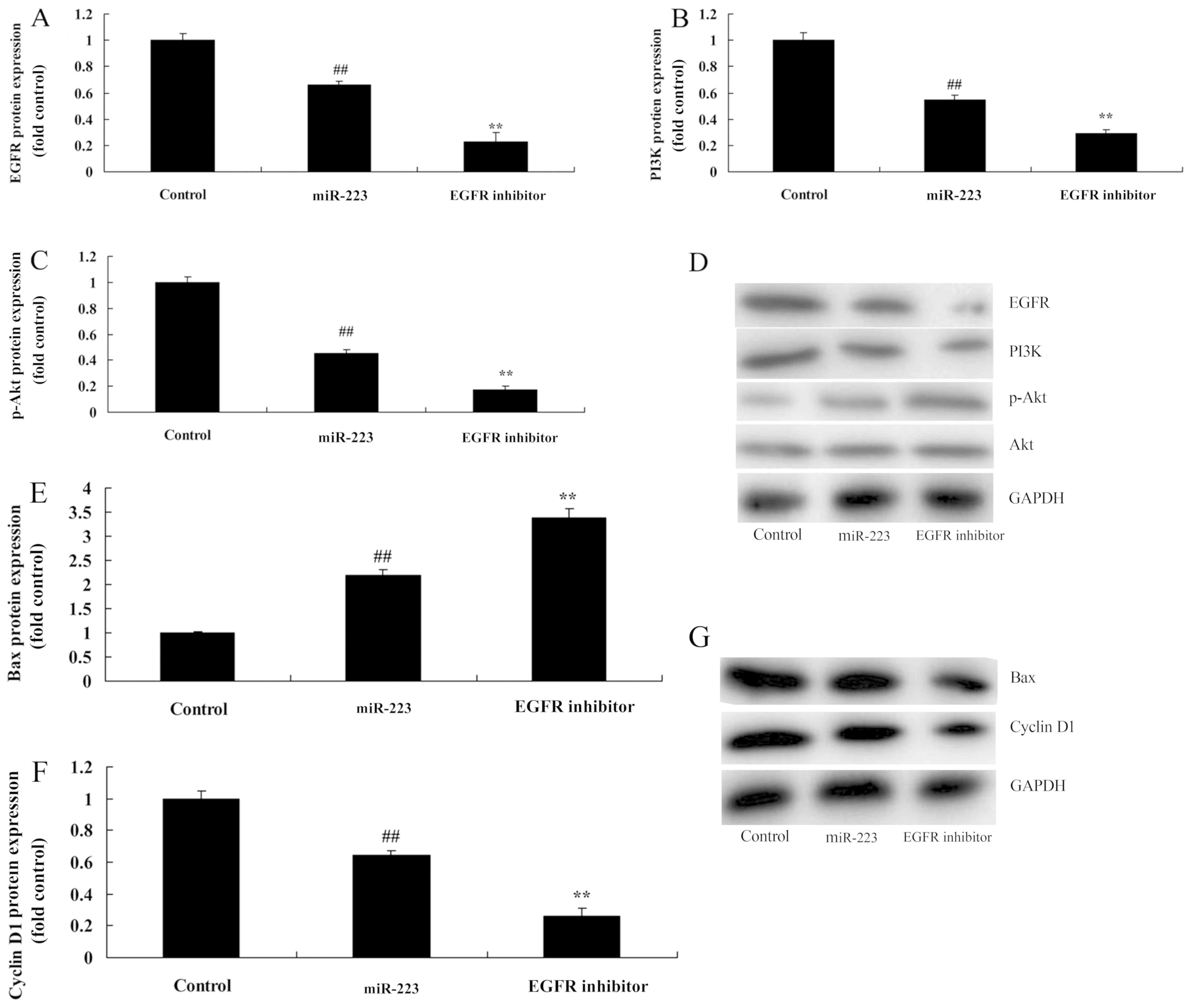
EGFR inhibition increases the
anticancer effects of miRNA-223 on EGFR protein expression in A549
cells
We next examined the impact of EGFR on the
anticancer effects of miRNA-223 on the apoptosis of A549 cells.
Treatment with 300 nM of AG1478 (EGFR inhibitor) for 48 h notably
suppressed EGFR, PI3K and p-Akt protein expression in the A549
cells following miRNA-223 transfection, compared with the miRNA-223
group (Fig. 6A-D). Compared with
the miRNA-223 group, EGFR inhibition evidently increased the
anticancer effects of miRNA-223 on the induction of Bax protein
expression and the suppression of cyclin D1 protein expression in
A549 cells (Fig. 6E-G).
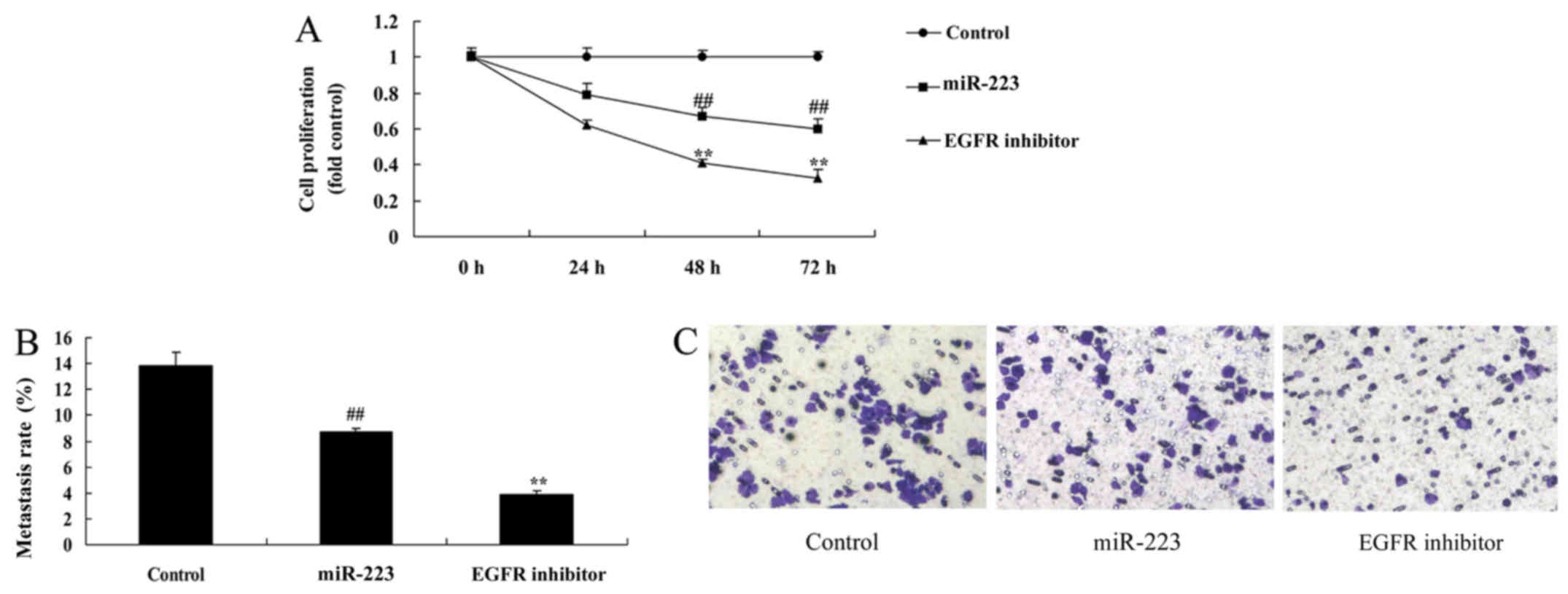
EGFR inhibition increases the
anticancer effects of miRNA-223 in A549 cells
It was found that EGFR inhibition dramatically
enhanced the anticancer effects of miRNA-223 in regards to
inhibition of cell proliferation and migration (Fig. 7A-C), while promoting apoptosis, LDH
activity and caspase-3/9 activity in A549 cells (Fig. 7D-G), compared with the miRNA-223
group.
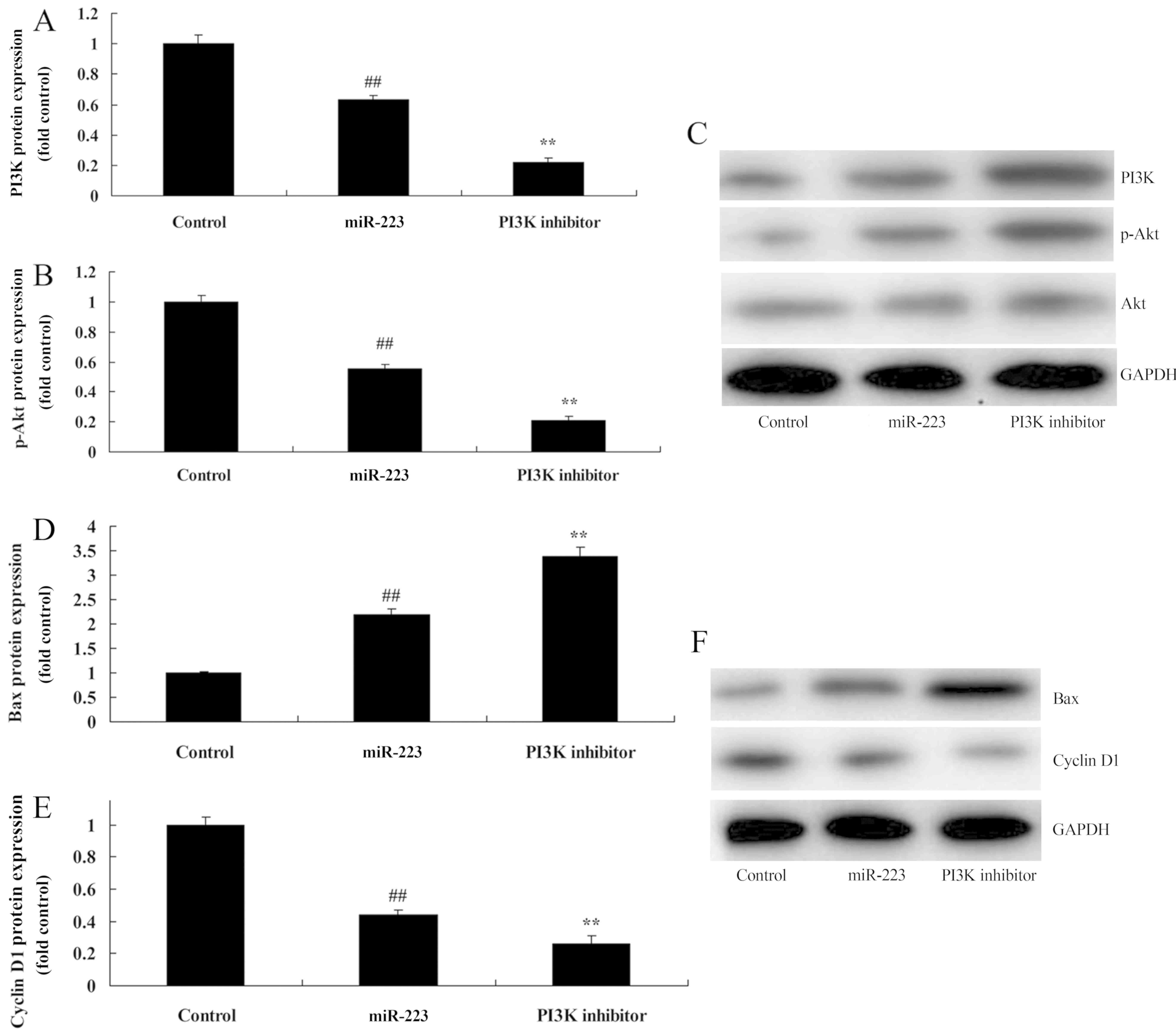
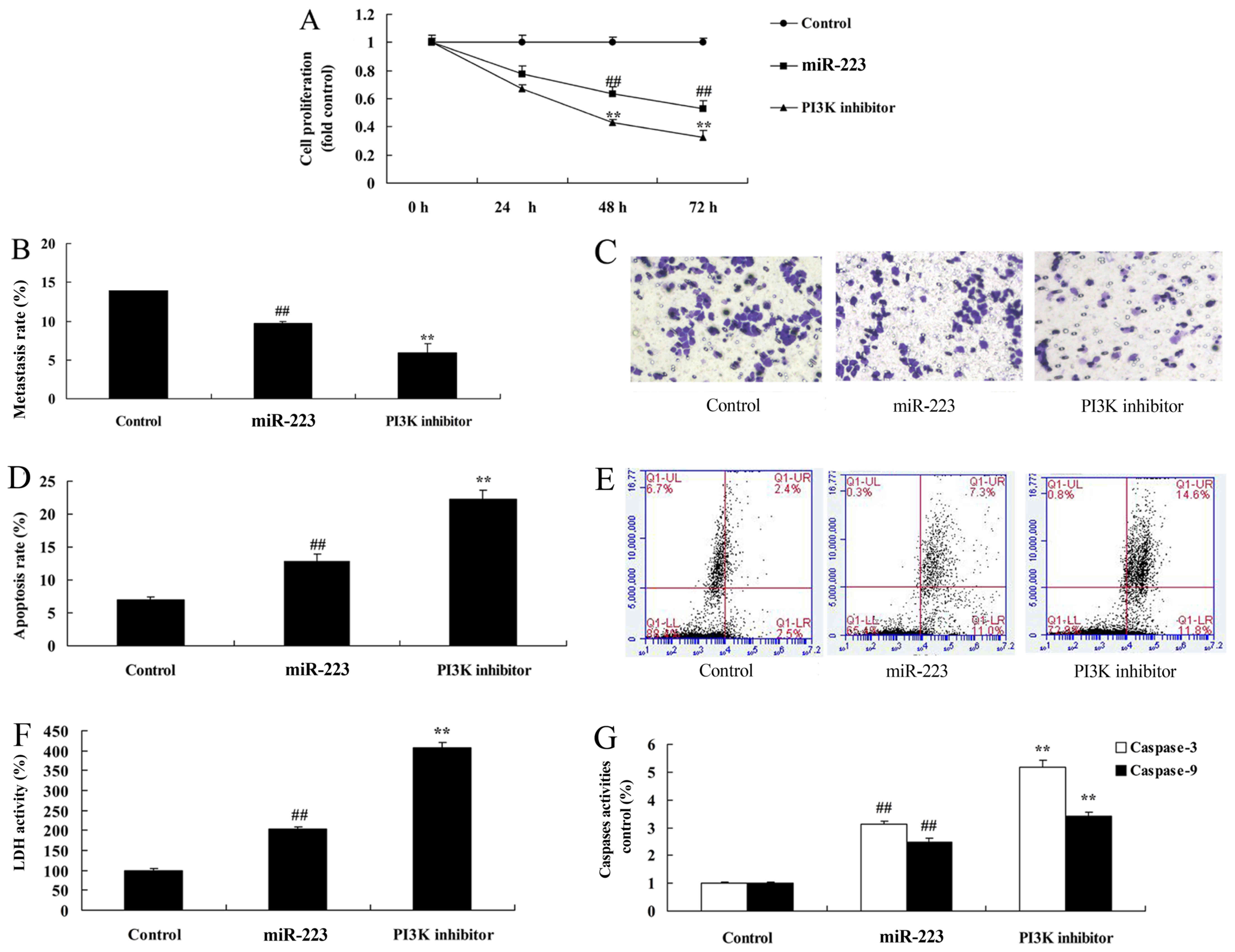
PI3K inhibition increases the
anticancer effects of miRNA-223 on PI3K protein expression in A549
cells
To further identify the potential role of PI3K in
the anticancer effects of miRNA-223 on the apoptosis of NSCLC, we
comparatively analyzed PI3K protein expression in A549 cells. As
shown in Fig. 8A-C, treatment of
200 nM of LY294002 (PI3K inhibitor) for 48 h evidently suppressed
PI3K and p-Akt protein expression in A549 cells following miRNA-223
transfection, compared with the miRNA-223 group. Compared with the
miR-223 group, PI3K inhibition significantly promoted the
anticancer effects of miRNA-223 on the induction of Bax protein
expression and the suppression of cyclin D1 protein expression in
A549 cells (Fig. 8D-F).
PI3K inhibition enhances the
anticancer effects of miRNA-223 in A549 cells
Compared with the miRNA-223 group, PI3K inhibition
significantly increased the anticancer effects of miRNA-223 in
regards to the inhibition of cell proliferation and migration
(Fig. 9A-C) while promoting
apoptosis, LDH and caspase-3/9 activities (Fig. 9D-G) in A549 cells.
Discussion
Lung cancer is associated with high morbidity and
poor patient prognosis (12). It
has become the leading major malignant tumor threatening the life
and health of individuals worldwide (12). Treatments for lung cancer include
surgery, radiotherapy, chemotherapy and molecular-targeted therapy.
Of these, surgical treatment exhibits the best efficacy on lung
cancer (12). Moreover, the overall
postoperative 5-year survival rate has reached 46.4% (13). The 5-year survival rates for stage
I, II, III and IV patients are 58.6, 25.9, 16.8 and 3.9%,
respectively. Surgical treatment has provided the greatest chance
of healing for stage I and II patients (13). However, 80% of Chinese patients with
lung cancer are in the advanced stage at diagnosis. As a result,
they are not suitable for surgical treatment. At present,
chemotherapy-oriented comprehensive treatment has become an
important therapeutic means for advanced NSCLC (14).
In recent years, it has been discovered that miRNAs
are factors involved in cancer (15). They have biological regulatory
function. They are involved in biological processes such as cell
division, proliferation, differentiation and development. Moreover,
they exhibit functions that are similar to oncogenes and
tumor-suppressor genes (16). Thus,
they play a vital role in tumor genesis and development (16). Approximately 1/3 of human genes are
regulated by miRNAs (16). miRNAs
display complicated biological functions during the genesis and
development of lung cancer (17).
We first demonstrated that serum levels of miRNA-223 in NSCLC
patients were downregulated; the downregulation of the expression
of miRNA-223 increased cell proliferation and migration of A549
cells.
EGFR plays a regulatory role in tumor cell
proliferation, differentiation and anti-apoptosis (6). Its abnormal expression can promote
tumor cell proliferation, adhesion, invasion and metastasis
(6). In addition, it induces tumor
angiogenesis. Research indicates that expression of phosphorylated
EGFR in NSCLC tissue is higher than that in para-carcinoma tissue
(7). This has revealed that high
EGFR expression is closely related to the genesis and development
of NSCLC (18). Binding of EGFR
with related ligands can activate the tyrosine signaling pathway
and PI3K/AKT signaling pathway. Meanwhile, it can upregulate the
expression of vascular endothelial growth factor (VEGF) and
epidermal growth factor (EGF). Consequently, excessive activation
of such pathways may enhance tumor cell proliferation, invasion and
metastasis. Meanwhile, it inhibits cell apoptosis (18). EGFR may activate the PI3K/AKT
signaling pathway (19). Our study
reported that overexpression of miRNA-223 suppressed EGFR protein
expression in A549 cells. It was demonstrated that miRNA-223
suppressed cervical cancer cell growth by targeting the
EGFR/AKT2/CCND1 pathway (11).
These results were in keeping with our results, and miRNA-223
suppressed EGFR protein expression to regulate the cell apoptosis
of NSCLC.
Theoretically, any inhibitor for blocking expression
of EGFR and PI3K/AKT signal pathway-related proteins can be used to
treat NSCLC. The application of EGFR tyrosine inhibitor in NSCLC
has ushered in a new horizon of targeted drug therapy aimed at the
activated signaling pathway (20).
However, the expected efficacy cannot be achieved in NSCLC patients
after applying EGFR tyrosine kinase inhibitor. PI3K inhibitor at
the current stage mainly targets catalytic subunit P110 (20). As reported in the literature,
abnormality in the PI3K/AKT signaling pathway is related to tumor
growth, maintenance and chemoresistance (20). Therefore, inhibiting such a
signaling pathway can reverse chemoresistance and poor prognosis
induced by activation of this pathway (21). At the same time, the combined
application of a PI3K/AKT signaling pathway inhibitor and a
traditional chemotherapeutic agent can enhance the chemotherapeutic
effect and enhance radiotherapy sensitivity (22). Our study showed that overexpression
of miRNA-223 suppressed PI3K and p-Akt protein expression in A549
cells. It was previously found that miRNA-223 suppressed cervical
cancer cell growth via targeting the EGFR/AKT2/CCND1 pathway
(11). Downregulation of miR-223
was also found to promote degranulation via the PI3K/Akt pathway by
targeting IGF-1R in mast cells (23), which demonstrated that the
miRNA-223/EGFR/PI3K/Akt pathway regulates the cell growth of
NSCLC.
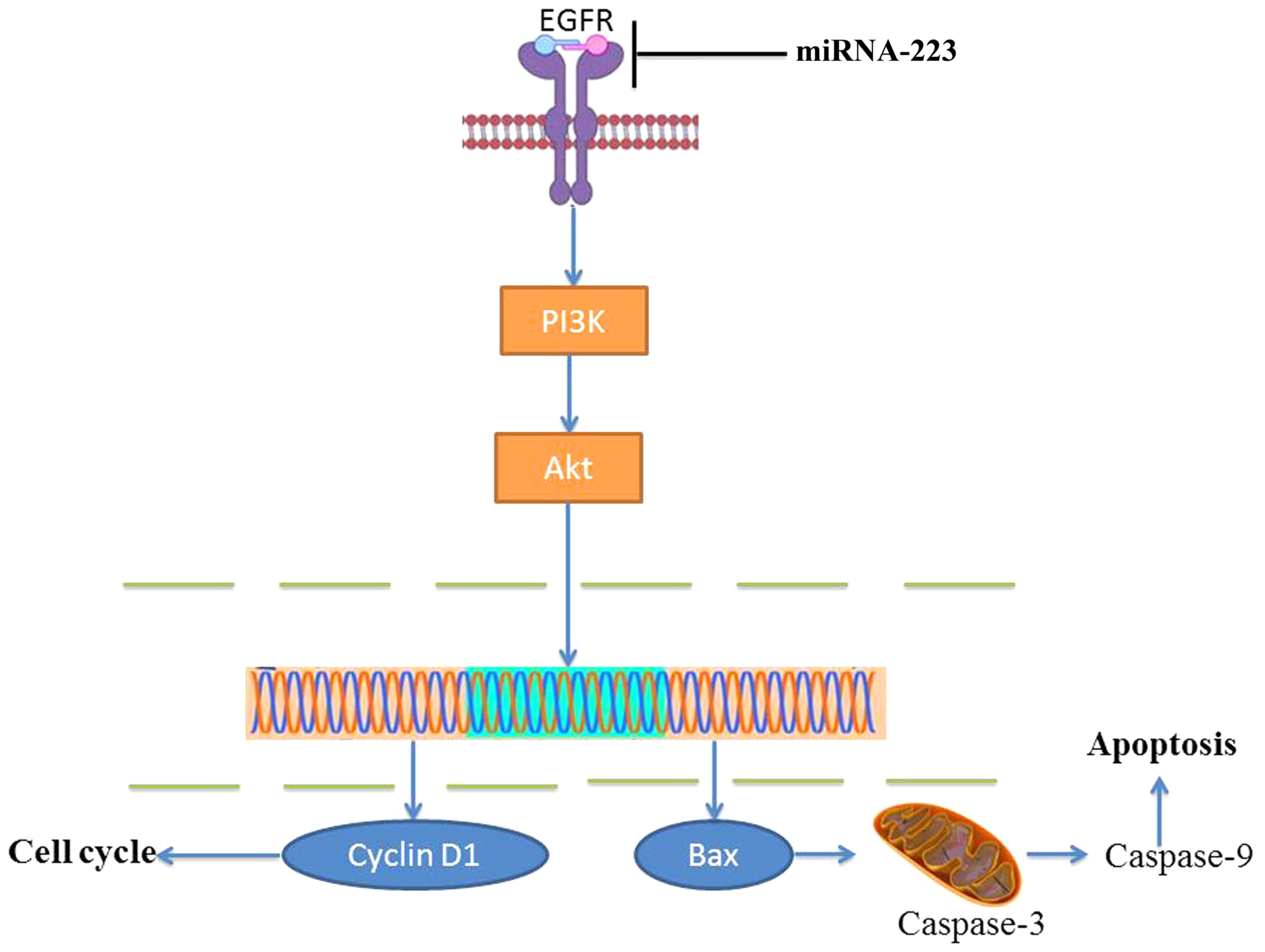
In conclusion, for the first time, we revealed that
overexpression of miRNA-223 increased the apoptosis of non-small
cell lung cancer cells through the PI3K/AKT pathway by targeting
EGFR (Fig. 10). This study
provides promising results supporting miRNA-223 as a novel
antitumor candidate target for non-small cell lung cancer and has
potential for further development as a single method for anticancer
therapy.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author upon reasonable
request.
Authors' contributions
XYL designed the experiment. HPM, WXK, WL, YZ and YW
performed the experiment. HPM and XYL analyzed the data. XYL wrote
the manuscript. All authors read and approved the final version of
the manuscript.
Ethics approval and consent to
participate
The study was approved by Jining First People's
Hospital, Jining, China.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Price TJ, Peeters M, Kim TW, Li J, Cascinu
S, Ruff P, Suresh AS, Thomas A, Tjulandin S, Zhang K, et al:
Panitumumab versus cetuximab in patients with
chemotherapy-refractory wild-type KRAS exon 2 metastatic colorectal
cancer (ASPECCT): A randomised, multicentre, open-label,
non-inferiority phase 3 study. Lancet Oncol. 15:569–579. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Denis F, Lethrosne C, Pourel N, Molinier
O, Pointreau Y, Domont J, Bourgeois H, Senellart H, Trémolières P,
Lizée T, et al: Randomized Trial Comparing a Web-Mediated Follow-up
With Routine Surveillance in Lung Cancer Patients. J Natl Cancer
Inst. 109:1092017. View Article : Google Scholar
|
|
3
|
Guo Y, Sun W, Gong T, Chai Y, Wang J, Hui
B, Li Y, Song L and Gao Y: miR-30a radiosensitizes non-small cell
lung cancer by targeting ATF1 that is involved in the
phosphorylation of ATM. Oncol Rep. 37:1980–1988. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li H, Zhou H, Luo J and Huang J:
MicroRNA-17-5p inhibits proliferation and triggers apoptosis in
non-small cell lung cancer by targeting transforming growth factor
β receptor 2. Exp Ther Med. 13:2715–2722. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ju L, Han M, Li X and Zhao C: MicroRNA
signature of lung adenocarcinoma with EGFR exon 19 deletion. J
Cancer. 8:1311–1318. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li X, Liu Y, Shi W, Xu H, Hu H, Dong Z,
Zhu G, Sun Y, Liu B, Gao H, et al: Droplet digital PCR improved the
EGFR mutation diagnosis with pleural fluid samples in
non-small-cell lung cancer patients. Clin Chim Acta. 471:177–184.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ni J, Weng L, Liu Y, Sun Z, Bai C and Wang
Y: Dynamic monitoring of EGFR mutations in circulating cell-free
DNA for EGFR-mutant metastatic patients with lung cancer: Early
detection of drug resistance and prognostic significance. Oncol
Lett. 13:4549–4557. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gao JW, Zhan P, Qiu XY, Jin JJ, Lv TF and
Song Y: Erlotinib-based doublet targeted therapy versus erlotinib
alone in previously treated advanced non-small-cell lung cancer: A
meta-analysis from 24 randomized controlled trials. Oncotarget.
8:73258–73270. 2017.PubMed/NCBI
|
|
9
|
Zhou G, Zhang F, Guo Y, Huang J, Xie Y,
Yue S, Chen M, Jiang H and Li M: miR-200c enhances sensitivity of
drug-resistant non-small cell lung cancer to gefitinib by
suppression of PI3K/Akt signaling pathway and inhibites cell
migration via targeting ZEB1. Biomed Pharmacother. 85:113–119.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu XL, Zhang XT, Meng J, Zhang HF, Zhao
Y, Li C, Sun Y, Mei QB, Zhang F and Zhang T: ING5 knockdown
enhances migration and invasion of lung cancer cells by inducing
EMT via EGFR/PI3K/Akt and IL-6/STAT3 signaling pathways.
Oncotarget. 8:54265–54276. 2017.PubMed/NCBI
|
|
11
|
Tang Y, Wang Y, Chen Q, Qiu N, Zhao Y and
You X: MiR-223 inhibited cell metastasis of human cervical cancer
by modulating epithelial-mesenchymal transition. Int J Clin Exp
Pathol. 8:11224–11229. 2015.PubMed/NCBI
|
|
12
|
Soria JC, Adjei AA, Bahleda R, Besse B,
Ferte C, Planchard D, Zhou J, Ware J, Morrissey K, Shankar G, et
al: A phase IB dose-escalation study of the safety and
pharmacokinetics of pictilisib in combination with either
paclitaxel and carboplatin (with or without bevacizumab) or
pemetrexed and cisplatin (with or without bevacizumab) in patients
with advanced non-small cell lung cancer. Eur J Cancer. 86:186–196.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wakelee HA, Dahlberg SE, Keller SM, Tester
WJ, Gandara DR, Graziano SL, Adjei AA, Leighl NB, Aisner SC,
Rothman JM, et al: ECOG-ACRIN: Adjuvant chemotherapy with or
without bevacizumab in patients with resected non-small-cell lung
cancer (E1505): An open-label, multicentre, randomised, phase 3
trial. Lancet Oncol. 18:1610–1623. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Melichar B, Adenis A, Lockhart AC,
Bennouna J, Dees EC, Kayaleh O, Obermannova R, DeMichele A,
Zatloukal P, Zhang B, et al: Safety and activity of alisertib, an
investigational aurora kinase A inhibitor, in patients with breast
cancer, small-cell lung cancer, non-small-cell lung cancer, head
and neck squamous-cell carcinoma, and gastro-oesophageal
adenocarcinoma: A five-arm phase 2 study. Lancet Oncol. 16:395–405.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Schofield P, Ugalde A, Gough K, Reece J,
Krishnasamy M, Carey M, Ball D and Aranda S: A tailored, supportive
care intervention using systematic assessment designed for people
with inoperable lung cancer: A randomised controlled trial.
Psychooncology. 22:2445–2453. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schuette W, Behringer D, Stoehlmacher J,
Kollmeier J, Schmager S, Fischer von Weikersthal L, Schumann C and
Buchmann J: CHAMP: A phase II study of panitumumab with pemetrexed
and cisplatin versus pemetrexed and cisplatin in the treatment of
patients with advanced-stage primary nonsquamous non-small-cell
lung cancer with particular regard to the KRAS status. Clin Lung
Cancer. 16:447–456. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou W, Ye XL, Xu J, Cao MG, Fang ZY, Li
LY, Guan GH, Liu Q, Qian YH and Xie D: The lncRNA H19 mediates
breast cancer cell plasticity during EMT and MET plasticity by
differentially sponging miR-200b/c and let-7b. Sci Signal.
10:102017. View Article : Google Scholar
|
|
18
|
Daly C, Castanaro C, Zhang W, Zhang Q, Wei
Y, Ni M, Young TM, Zhang L, Burova E and Thurston G: FGFR3-TACC3
fusion proteins act as naturally occurring drivers of tumor
resistance by functionally substituting for EGFR/ERK signaling.
Oncogene. 36:471–481. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim JY, Welsh EA, Fang B, Bai Y, Kinose F,
Eschrich SA, Koomen JM and Haura EB: Phosphoproteomics reveals MAPK
inhibitors enhance MET- and EGFR-driven AKT signaling in
KRAS-mutant lung cancer. Mol Cancer Res. 14:1019–1029. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pan H, Jiang T, Cheng N, Wang Q, Ren S, Li
X, Zhao C, Zhang L, Cai W and Zhou C: Long non-coding RNA BC087858
induces non-T790M mutation acquired resistance to EGFR-TKIs by
activating PI3K/AKT and MEK/ERK pathways and EMT in non-small-cell
lung cancer. Oncotarget. 7:49948–49960. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Torres AF, Nogueira C, Magalhaes J, Costa
IS, Aragao A, Gomes Neto A, Martins F and Tavora F: Expression of
EGFR and molecules downstream to PI3K/Akt, Raf-1-MEK-1-MAP
(Erk1/2), and JAK (STAT3) pathways in invasive lung adenocarcinomas
resected at a single institution. Anal Cell Pathol (Amst) 2014.
3529252014.
|
|
22
|
Li H, Schmid-Bindert G, Wang D, Zhao Y,
Yang X, Su B and Zhou C: Blocking the PI3K/AKT and MEK/ERK
signaling pathways can overcome gefitinib-resistance in non-small
cell lung cancer cell lines. Adv Med Sci. 56:275–284. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Q, Zhao DY, Xu H, Zhou H, Yang QY,
Liu F and Zhou GP: Down-regulation of microRNA-223 promotes
degranulation via the PI3K/Akt pathway by targeting IGF-1R in mast
cells. PLoS One. 10:e01235752015. View Article : Google Scholar : PubMed/NCBI
|