Introduction
Ischemic stroke, caused by disruption of the brain
blood supply, is the primary cause of death and injury worldwide
(1). Intervention requires the
restoration of blood flow, which can lead to reperfusion injury.
Thus, ischemic stroke represents a major problem from a public
health perspective, and clarifying the mechanism of ischemic
cerebral injury and thereby identifying appropriate molecular
targets for stroke prevention and treatment is a priority.
MicroRNAs (miRNAs) are small non-coding RNA
molecules that regulate gene expression by inhibiting translation
or cutting RNA transcripts in a sequence-specific manner (2). miRNAs play an important role in cell
development, differentiation and apoptosis, and regulate at least
one-third of all human genes (3).
It has been reported that >20% of miRNAs are altered in the
ischemic brain, implying that miRNAs are important mediators in
ischemic stroke pathogenesis (4,5–9).
Therefore, identification of these miRNAs may not only reveal the
underlying molecular mechanisms of ischemic stroke, but also
provide novel therapeutic targets for early diagnosis and
treatment.
The miR-29 family of microRNAs is highly conserved
among mammalian species, and its members modulate somatic cell fate
reprogramming of fibroblasts to induced pluripotent stem cells
(iPSCs) (10–12). miR-29 is also enriched in astrocytes
(13). The miR-29 family includes
three variants (miR-29a, miR-29b and miR-29c). miR-29a and
miR-29b-1 are situated on chromosome 7q32.3, while miR-29b-2 and
miR-29c are located on chromosome 1q32.2 in human cells (14). Numerous small molecule regulators of
gene expression have been implicated in ischemic brain injury. miRs
are altered in human plasma (15)
and the rodent brain (16) in
response to ischemic stroke injury, whereas circulating miRs remain
stable and consistent, highlighting their potential application as
biomarkers and therapeutic targets for stroke (17). Furthermore, miR-424 overexpression
was revealed to reduce ischemic brain injury by inhibiting the
activation of microglial cells, suggesting a novel stroke
intervention strategy based on miRs (18). Stary et al revealed that
inhibiting miR-200c levels and upregulating reelin expression in
the acutely injured brain could minimise the development of injury
and enhance recovery (19). miR-21
has an anti-apoptotic effect, however miR-24 has the opposite
effect in oxygen glucose deprivation (OGD)-induced N2A
neuroblastoma cells, indicating that these microRNAs may be
potential therapeutic tools in ischemic injury treatment (20). Amplification of cerebral ischemia
injury induced by miR-182 was aggravated by inhibition of members
of the apoptosis-stimulating proteins of p53 (ASPP) family (iASPP)
(21). miRNA-124 was revealed to
prevent brain ischemia via Usp14-dependent degradation of
RE1-silencing transcription factor (REST) (22).
Aquaporin 4 (AQP4) is the main water channel protein
in the central neuropil, and its expression is highly polarised
(23). It is primarily found on
astrocytes, particularly on the end of astrocytes surrounding
capillaries and the blood-cerebral barrier, and also in glia cells
to a more limited extent (24,25).
It has been reported that knockdown of AQP4 protects neurocytes
against toxic oedema caused by water intoxication and permanent
focal cerebral ischemia in mice (26). Evidence has revealed that AQP4 plays
a role in promoting brain ischemia (27), and a recent study demonstrated that
a miR-29b mimic reduced injury to the blood-brain barrier by
inhibiting the expression of AQP4 in ischemic stroke (28). However, the molecular mechanism
underlying AQP4 upregulation at the transcriptional and
post-transcriptional levels remains unknown.
In the present study, the effects of miR-29a were
investigated using an in vitro rat astrocyte OGD injury
model, and focused on the involvement of AQP4 in this process.
Materials and methods
Animals
Middle cerebral artery occlusion
(MCAO)/reperfusion model
Six-week-old C57BL/6J mice (20–25 g; male) were
purchased from the Experimental Animal Center of Zhejiang
University School of Medicine (SCXK2012-002) and and their use was
approved by The Medical Ethics Committee and the Medical Faculty
Ethics Committee of The First Affiliated Zhejiang Hospital,
Zhejiang University (Ref. no. 2018-811). All experiments were
performed in accordance with the guidelines and regulations of the
Guide for the Care and Use of Experimental Animals of the Zhejiang
Unveristy School of Medicine and housed in a controlled 12-h
light/dark cycle environment with access to chow and water ad
libitum. Middle Cerebral Artery Occlusion (MCAO) was
established using previously published methods (29,30). A
total of 18 mice were randomly divided into three groups (n=6): The
sham group, the MCAO 24-h group, and the MCAO 48-h group. In brief,
4% chloral hydrate (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
was used to anesthetise the mice and a silicone-coated 6-0
monofilament nylon suture (Doccol Corp., Redlands, CA, USA) was
inserted into the left common carotid artery to occlude the origin
of the MCA. After 1 h, the suture was withdrawn. In the sham
groups, filaments were prepared and inserted into the left common
carotid artery. To anaesthetize the mice, 40 mg/kg chloral hydrate
was used and decapitation was performed to obtain the brain.
Oxygen glucose
deprivation-reoxygenation (OGD-RX) and cell culture
Primary astrocyte cells were isolated from 24-h
postnatal neonatal Sprague-Dawley (SD) rat brains (31). The SD rats were purchased from the
Experimental Animal Center of Zhejiang University School of
Medicine (SCXK2012-002), and their use was approved by the Medical
Ethics Committee and the Medical Faculty Ethics Committee of The
First Affiliated Zhejiang Hospital, Zhejiang University (Ref. no.
2018-811). All rats were housed in a controlled 12-h light/dark
cycle environment with access to chow and water ad libitum.
OGD (95% N2 and 5% O2 at 37°C) was induced
for 1, 3, 6 or 12 h in a hypoxic chamber as previously described
(32,33). Briefly, the cells were then
subjected to oxygen restoration (5% CO2 and 95%
atmospheric air) and cultured at 37°C for 24 h. According to the
results of preliminary experiments, astrocyte cells could maintain
their basic shape and demonstrated relatively high cell viability
after 6 h OGD and 24 h restoration. Therefore, these conditions
were used for subsequent experiments. Astrocyte cells were
maintained in Dulbecco's modified Eagle's medium (DMEM)/F12
containing 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) and 1% penicillin/streptomycin
(Sigma-Aldrich; Merck KGaA). Cells were maintained in 5%
CO2 at 37°C for 24 h in a humidified incubator.
Following this period, the cells were collected and used in
subsequent experiments.
siRNA transfection
According to the manufacturer's instructions,
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.) was
used to transfect astrocytes with AQP4 siRNA (cat. nos. sc-29716
and sc-156007) or a negative control siRNA (cat. no. sc-37007)
(Santa Cruz Biotechnology, Inc., Dallas, TX, USA) according to the
manufacturer's instructions. After transfection, complete medium
was replaced with transfection medium (Opti-MEM; Gibco; Thermo
Fisher Scientific, Inc.), and cells were cultured for the 48 h.
Western blot analysis
Cells were washed with phosphate-buffered saline
(PBS), lysed with protein lysis solution (Cell Signaling
Technology, Inc., Danvers, MA, USA) containing protease inhibitors
(Sigma-Aldrich; Merck KGaA), and centrifuged at 12,000 × g for 5
min at 4°C. The supernatant was collected, and a BCA Protein Assay
Kit (Sigma-Aldrich; Merck KGaA) was used to assess the protein
concentration. Protein samples (40 µg/lane) were separated by 10%
SDS-PAGE, and then proteins were transferred to a polyvinylidene
difluoride (PVDF) membrane (EMD Millipore, Billerica, MA, USA). The
membranes were blocked with Tris-buffered saline (TBS) and 0.1%
Tween-20 (TBS/T) containing 5% bovine serum albumin (BSA) and
incubated with primary antibody against AQP4 (cat. no. ab46182;
Abcam, Cambridge, USA) diluted 1:1,000 in TBS/T overnight at 4°C.
After washing three times, the membranes were incubated with a
horseradish peroxidase-labeled secondary antibody (cat. no. ab6721)
diluted 1:2,000 at room temperature for 2 h. Subsequently, the
protein bands were assessed by chemiluminescence (GE Healthcare,
Piscataway, NJ, USA). Bands were quantified by densitometry and
glyceraldehyde-3-phosphate dehydrogenase (GAPDH; product code:
EM1101; Hangzhou HuaAn Biotechnology Co., Ltd., Hangzhou, China)
was used as an internal control.
Quantitative reverse-transcription PCR
(qRT-PCR)
Total RNA was extracted using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. A 2-µg sample of total RNA was
reverse-transcribed into first-strand complementary DNA (cDNA)
using a PrimeScript RT Reagent Kit (Takara Biotechnology Co., Ltd.,
Dalian, China). After the RT reaction, 1 µl of cDNA was used for
subsequent qPCR with SYBR-Green dye (Takara Biotechnology Co.,
Ltd.) and a 7500 Real-Time PCR System (Applied Biosystems; Thermo
Fisher Scientific, Inc.). Reaction conditions were 95°C for 30 sec
followed by 40 cycles at 95°C for 5 sec and annealing at 60°C for
34 sec. DNA primers specific for AQP4 and β-actin were purchased
from GenePharma Co., Ltd. (Shanghai, China). U6 and β-actin were
used as internal controls, and the comparative threshold cycle
(2−ΔΔCt) (34) method
was employed to calculate the relative expression levels. All
reactions were performed in triplicate, and primers were as
follows: AQP4 forward, 5′-CCCGCAGUUAUCAUGGGAATT-3′ and reverse,
5′-UUCCCAUGAUAACUGCGGGTT-3′; miR-29a mimic forward,
5′-UAGCACCAUCUGAAAUCGGUUA-3′ and reverse,
5′-ACCGAUUUCAGAUGGUGCUAUU-3′; and miR-29a inhibitor,
5′-UAACCGAUUUCAGAUGGUGCUA-3′,
Immunofluorescence
Immunofluorescence was used to identify astrocytes.
Cells were washed with cold PBS, fixed in 4% paraformaldehyde for
15 min, blocked with 5% BSA at 37°C for 30 min, and incubated with
a 1:100 dilution of GFAP (cat. no. ab7260; Abcam) at 4°C overnight.
After washing with PBS three times, the cells were incubated with a
1:100 dilution of a secondary antibody (cat. no. ab6816; Abcam) at
37°C for 2 h. DAPI (Sigma-Aldrich; Merck KGaA) was used for nuclear
staining at 37°C for 2 min, and then cells were washed three times
with PBS, and observed using an inverted fluorescence microscope
(Olympus Corp., Tokyo, Japan).
Cell health assay
Cell health was assessed by calcein-AM/PI staining
(cat. no. 13837S; Cell Signaling Technology, Inc.). In brief, cells
were fixed with 70% ethanol for 15 min, stained with calcein-AM
solution (10 µmol/l) for 15 min, washed with PBS buffer and then
stained with PI (10 µmol/l) for 15 min. Excitation filters (490 and
535 nm) were used to observe healthy and dead cells,
respectively.
Flow cytometric analysis
Cells were digested with 0.25% trypsin solution
(Life Technologies; Thermo Fisher Scientific, Inc.) without EDTA
and diluted into a single-cell suspension (2×105
cells/ml). After washing three times with PBS, an Annexin V-FITC
Cell Apoptosis Detection kit (cat. no. 556547; BD Biosciences, San
Jose, CA, USA) was used to determine the proportion of apoptotic
cells. Analysis was carried out immediately (within 1 h) by flow
cytometry using a FACS instrument (BD Biosciences).
Enzyme-linked immunosorbent assay
(ELISA) analysis
For ELISA, the levels of lactate dehydrogenase (LDH)
were determined using an ELISA kit according to the manufacturer's
instructions (Cytotoxicity Detection KitPLUS; cat. no.
04744926001; Roche Diagnostics, Basel, Switzerland). The LDH
concentrations were calculated according to the absorbance of the
samples and the standard curve.
Luciferase reporter assay
The 3′-untranslated region (UTR) and mutated 3′-UTR
of the amplified AQP4 fragment was cloned into the pGL3 vector
containing the firefly luciferase reporter gene (Promega Corp.,
Madison, WI, USA). According to the manufacturer's protocol
supplied with the luciferase reporter assay, we co-transfected 200
ng of firefly luciferase construct with 4 ng of pRL-TK
Renilla luciferase plasmid and 50 nM of miR-29a-3p mimic
into rat astrocytes. At 48 h after transfection, dual-luciferase
reporter assays (Promega Corp.) were performed to assess the
activity of Renilla luciferase, and the results were
calculated as the relative luciferase activity (firefly
luciferase/Renilla luciferase).
Histomorphological analysis and oedema
formation
Brains were rapidly removed and immediately frozen
after animals were sacrificed at 1 or 3 days after MCAO. Brain
tissues were fixed in a 10% formalin solution (Sigma-Aldrich; Merck
KGaA) for 24 h, and were embedded in paraffin wax (Sigma-Aldrich;
Merck KGaA). After deparaffinization with xylene, embedded tissue
was cut into 7-µm sections. For histomorphological analysis,
sections were stained with hematoxylin and eosin (H&E;
Sigma-Aldrich; Merck KGaA) and examined using a light microscope
(BX40; Olympus). The oedema volume was assessed as described in a
previous study (35) using cresyl
violet staining. Brain oedema formation was calculated as [1 -
(total ipsilateral hemisphere-infarct)/total contralateral
hemisphere] × 100%.
Statistical analysis
All experimental data were expressed as the mean ±
SD. GraphPad Prism 5 (GraphPad Software Inc., San Diego, CA, USA)
was used for statistical analysis. Student's t-test
(parametric)/Mann-Whitney (non-parametric) test were used to
perform comparisons of two groups, and one-way analysis of variance
followed by Tukey's post hoc test was used to compare multiple
groups (P<0.05, P<0.01 and P<0.001 were considered
statistically significant). We used TargetScan (www.targetscan.org) to predicate the association
between miR-29a and AQP4.
Results
miR-29a is associated to OGD/RX
astrocyte cell injury in vitro and MCAO in vivo
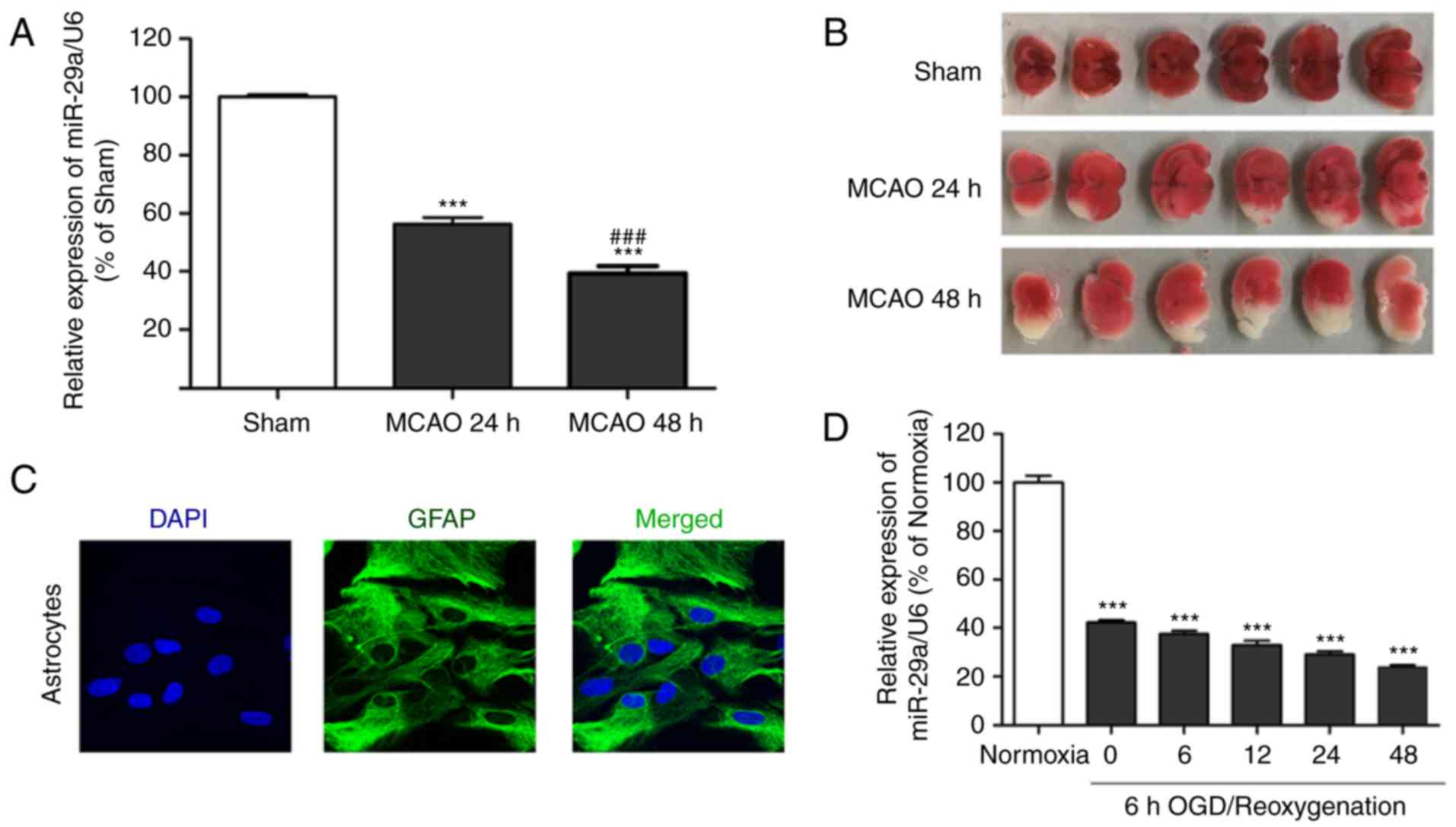
To investigate the involvement of miR-29a in OGD/RX
astrocyte cell injury, we assessed miR-29a expression by qRT-PCR.
The results demonstrated that miR-29a expression was significantly
decreased in MCAO mice at 24 and 48 h compared with the sham group
(Fig. 1A). The in vivo study
confirmed that MCAO resulted in increased infarct size (Fig. 1B). Astrocyte cells from rat brains
were isolated and the cell type was confirmed by immunofluorescence
assay with GFAP antibody and DAPI staining (Fig. 1C). It was also revealed that miR-29a
was significantly decreased in the OGD/RX group compared with the
normoxia group (Fig. 1D).
Collectively, these results demonstrated that miR-29a was
associated to OGD-induced astrocyte cell injury and MCAO.
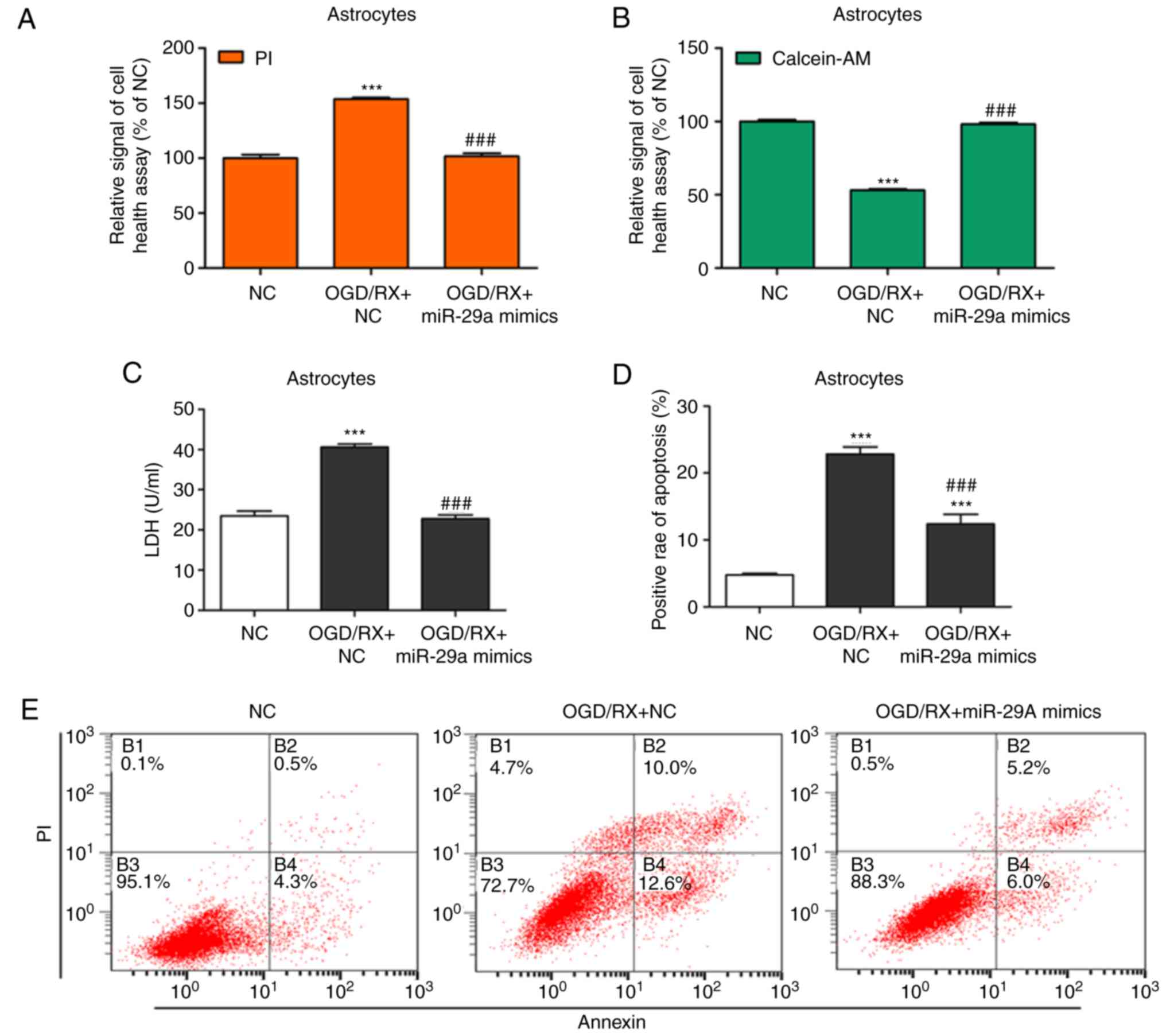
miR-29a protects against
OGD/RX-induced astrocyte cell injury
To study the protective effects of miR-29a in
OGD-induced astrocyte cell injury, cell supernatant LDH levels,
cell health, and cell apoptosis were examined in primary cultured
astrocytes in the NC (negative control), OGD/RX, OGD/RX+miR-29a
mimic, and OGD/RX+miR-29a inhibitor groups. Overexpression of
miR-29a significantly decreased apoptosis and promoted astrocyte
cell health (Fig. 2A, B, D and E),
indicating a protective role in astrocyte ischemia injury. In
addition, miR-29a inhibition reversed these effects (Fig. 2F, G, I and J). As revealed as in
Fig. 2C, transfection with the
miR-29a mimic significantly decreased LDH expression compared with
the NC group, while the miR-29a inhibitor significantly increased
the level of LDH (Fig. 2H). Western
blotting revealed that the expression of Bax was increased and
Bcl-2 was decreased in the OGD/RX group compared with the control,
while cells transfected with the miR-29 mimic displayed
downregulated Bax and upregulated Bcl-2 compared with the OGD/RX
group, and the miR-29a inhibitor had the opposite effect (Fig. 2K and L). These findings revealed
that overexpression of miR-29a protected astrocytes against OGD
injury.
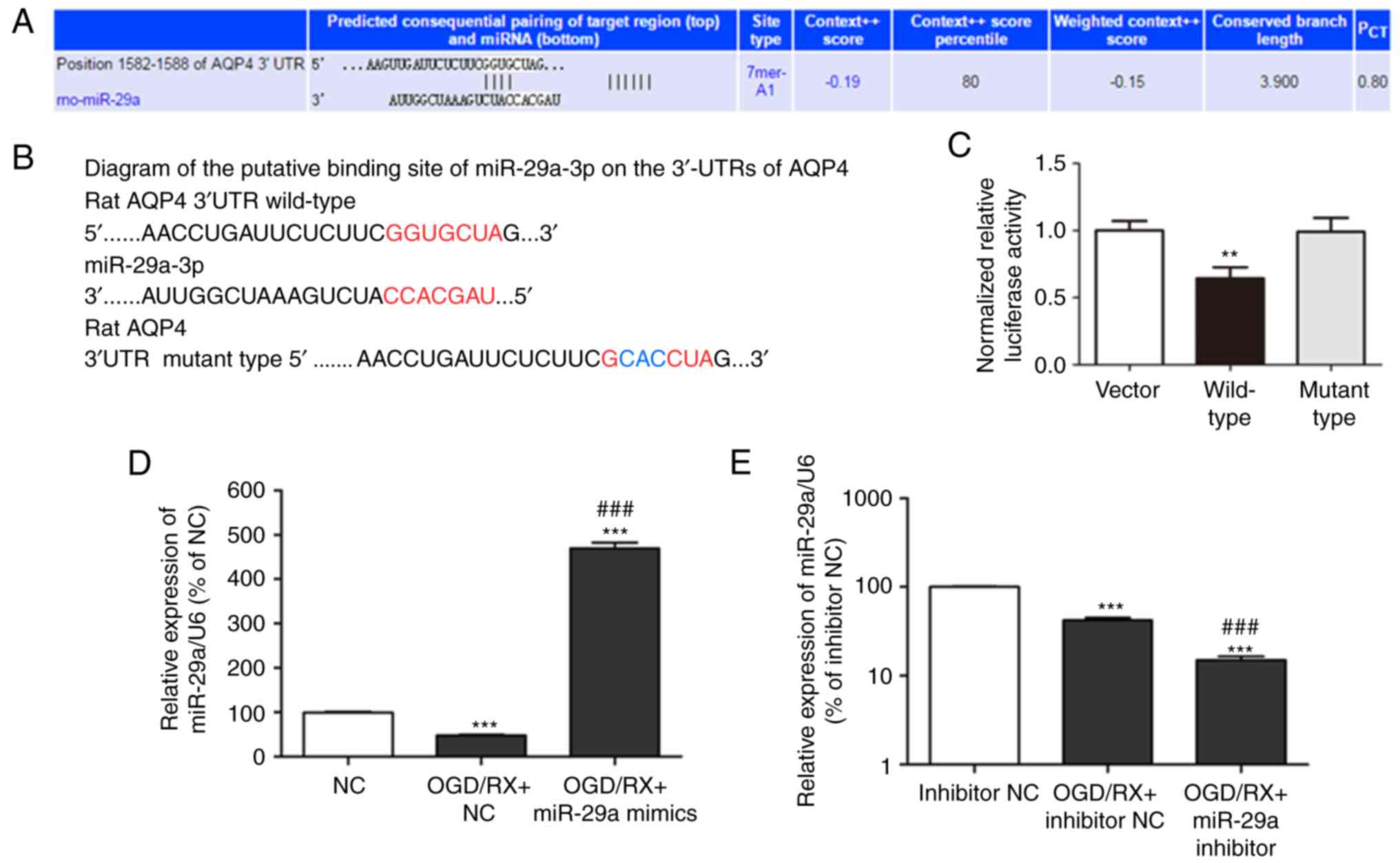
miR-29a regulates AQP4 expression
It was hypothesised that AQP4 is regulated by
miR-29a, and the association between miR-29a and AQP4 was predicted
using TargetScan (www.targetscan.org; Fig.
3A). In order to confirm whether AQP4 was regulated by
miR-29a-3p, native and mutated fragments of the AQP4 mRNA 3′-UTR
were used which included two miR-29a-3p binding sites upstream of
the luciferase coding sequence. Luciferase reporter assays were
then performed after co-transfection of rat astrocytes with
miR-29a-3p mimics (Fig. 3B).
Luciferase activity was decreased after co-transfection with
miR-29a-3p mimics and the AQP4 mRNA 3′-UTR fragment, but not with
the co-transfection of miR-29a-3p mimics and the mutant 3′-UTR
fragment (Fig. 3C). These findings
revealed that AQP4 was a direct target of miR-29a-3p. AQP4 protein
and miR-29a expression levels were also assessed in cells
transfected with the miR-29a mimic or the miR-29a inhibitor, and
compared with negative controls (NC). qRT-PCR analysis revealed
that miR-29a was decreased in OGD/RX cells compared with the NC
group, however treatment with the miR-29a mimic upregulated miR-29a
expression, and the miR-29a inhibitor had the opposite effect
(Fig. 3D and E). Western blot
analysis indicated that the expression of the AQP4 protein was
increased in the OGD/RX group compared to the NC group, while AQP4
protein levels were decreased in the OGD+miR-29a mimic group
compared with the OGD/RX and NC groups, while it was increased in
the OGD+miR-29a inhibitor group (Fig.
3F and G). These results indicated that the miR-29a mimic
downregulated OGD-mediated AQP4 expression to protect against
astrocyte cell injury.
AQP4 promotes OGD/RX-induced astrocyte
cell injury and MCAO
The expression of AQP4 at the protein and mRNA
levels was then assessed by western blotting and qRT-PCR,
respectively. The OGD group revealed higher AQP4 expression than
the normoxia group (Fig. 4A and B),
and MCAO had the same effect (Fig. 4I
and J). AQP4 and Bax expression was increased, while Bcl-2
expression was decreased in the OGD/RX group compared with the NC
group, but knockdown of AQP4 could reverse this effect (Fig. 4C). Cell health experiments revealed
that knockdown of AQP4 could protect against OGD-induced astrocyte
cell injury (Fig. 4D and E). The
ELISA experiment demonstrated that the level of LDH in astrocytes
of the OGD/RX+AQP4 siRNA group was significantly decreased compared
with that in cells of the OGD/RX group (Fig. 4F). In addition, the rates of
apoptotic astrocytes were examined by flow cytometry (Fig. 4G and H). The results revealed a
significant decrease in the rate of apoptotic cells in the
OGD/RX+AQP4 siRNA group compared with the OGD/RX group.
Collectively, these results demonstrated that inhibition of AQP4
could protect astrocytes against OGD-induced cell injury.
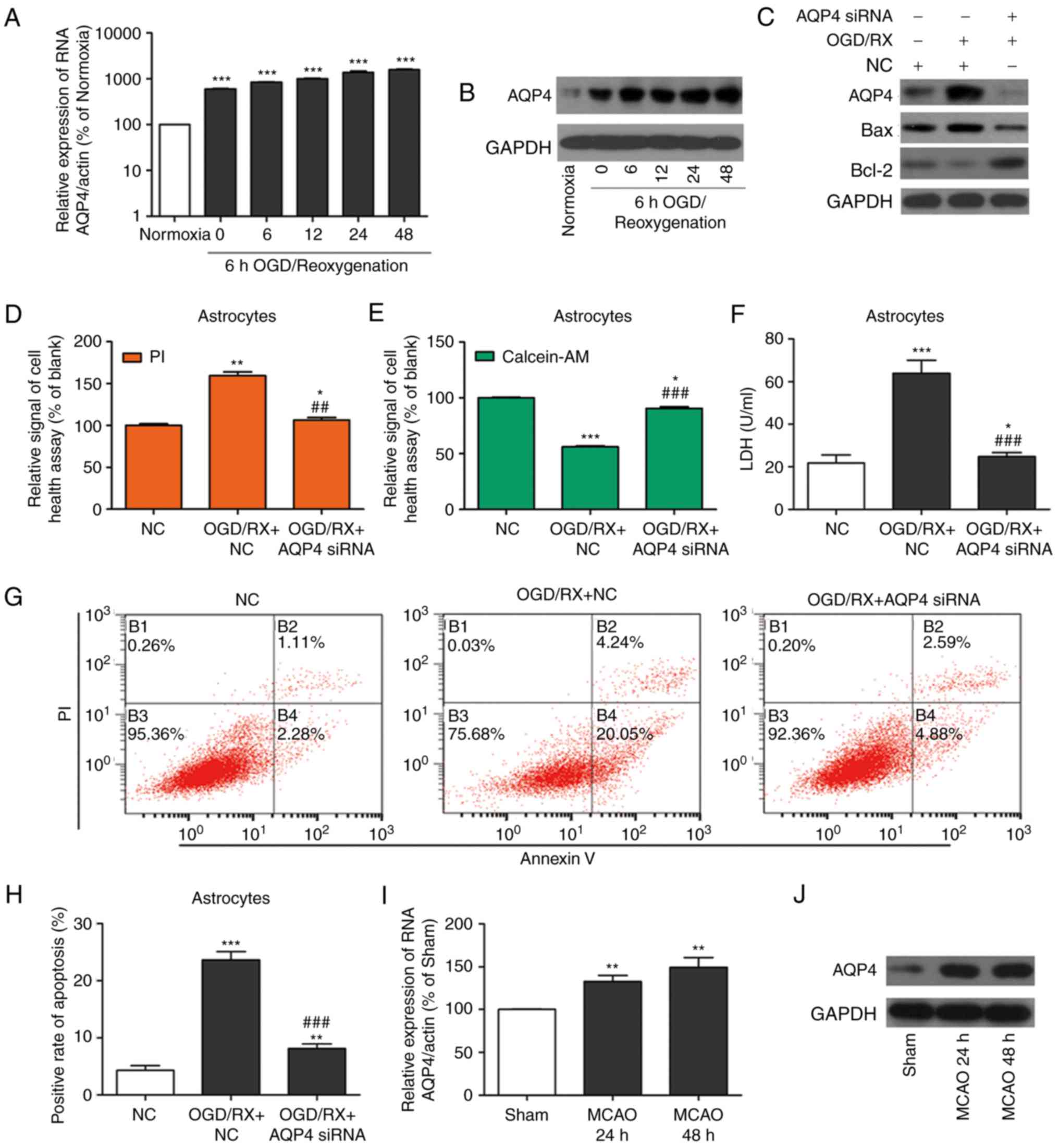 | Figure 4.Inhibition of AQP4 ameliorates
OGD/RX-induced astrocyte cell injury and MCAO. (A) qRT-PCR analysis
of the expression of AQP4 mRNA. ***P<0.001 vs. normoxia (n=3 for
each experiment). (B) Western blotting of AQP4 protein expression
in astrocyte cells treated with OGD for 6 h and following different
reoxygenation times (0, 6 12, 24 or 48 h) or normoxia. All
experiments were repeated in triplicate. Bands have been cropped
from different parts of different gels, and respective western blot
images have been cropped for Figure implementation. (C) Expression
of AQP4, Bax and Bcl-2 in OGD-treated astrocytes transfected with
miR-29a siRNA or NC examined by western blotting. All experiments
were repeated in triplicate. Bands have been cropped from different
parts of different gels, and respective western blot images have
been cropped for Figure implementation. (D and E) Cell health assay
of OGD-treated primary cultured astrocytes transfected with AQP4
siRNA or NC following PI/calcein staining. *P<0.05, **P<0.01,
***P<0.001 vs. NC; ##P<0.01,
###P<0.001 vs. OGD/RX+NC (n=3 for each experiment).
(F) Expression of LDH determined by ELISA. *P<0.05,
***P<0.001 vs. NC; ###P<0.001 vs. OGD/RX+NC (n=3
for each experiment). (G and H) Flow cytometric estimation of
apoptosis in astrocytes transfected with AQP4 siRNA or NC after OGD
injury. **P<0.001, ***P<0.001 vs. NC;
###P<0.001 vs. OGD/RX+NC (n=3 for each experiment).
(I) qRT-PCR analysis of the expression of AQP4 mRNA at 24 and 48 h
after MCAO. **P<0.01 vs. sham. All experiments were repeated in
triplicate. (J) Expression of AQP4 protein analysed by western
blotting at 24 and 48 h after MCAO (n=3 for each experiment). AQP4,
aquaporin 4; OGD/RX, oxygen glucose deprivation-reoxygenation; NC,
negative control; PI, propidium iodide; MCAO, middle cerebral
artery occlusion. |
Protective effect of miR-29a on
OGD/RX-induced astrocyte cell injury is mediated via AQP4
To determine whether miR-29a achieved OGD protection
by regulating AQP4 expression, OGD-astrocyte cells were
co-transfected with AQP4 siRNA, miR-29a inhibitor, and OGD/RX.
qRT-PCR revealed that miR-29a was downregulated after transfection
with miR-29a inhibitor compared with OGD/RX, however AQP4 siRNA
could block the effect of the miR-29a inhibitor (Fig. 5A and B). Western blotting revealed
that miR-29a inhibitor could increase AQP4 expression, however
co-transfection with AQP4 siRNA abrogated the effect of the miR-29
inhibitor (Fig. 5A and B).
Furthermore, the LDH levels in the cell supernatant as well as cell
health and cell apoptosis were examined, and the results revealed
that the miR-29a inhibitor could significantly enhance LDH
expression and apoptosis, and impair astrocyte cell health,
compared to OGD/RX, however, knockdown of AQP4 could prevent the
effects of the miR-29a inhibitor LDH levels, cell health and cell
apoptosis (Fig. 5C-G). H&E
analysis and oedema formation revealed that under OGD/RX
conditions, the oedema area was increased, and injury was enhanced
after transfection with the miR-29a inhibitor compared to the
controls. However, transfection with AQP4 siRNA and co-transfection
with AQP4 siRNA and the miR-29a inhibitor reduced the oedema area
compared with the controls (Fig. 5H and
I). The relative rate of apoptosis is represented as a
histogram. These findings ascertained that the protective role of
miR-29a was mediated via AQP4 inhibition.
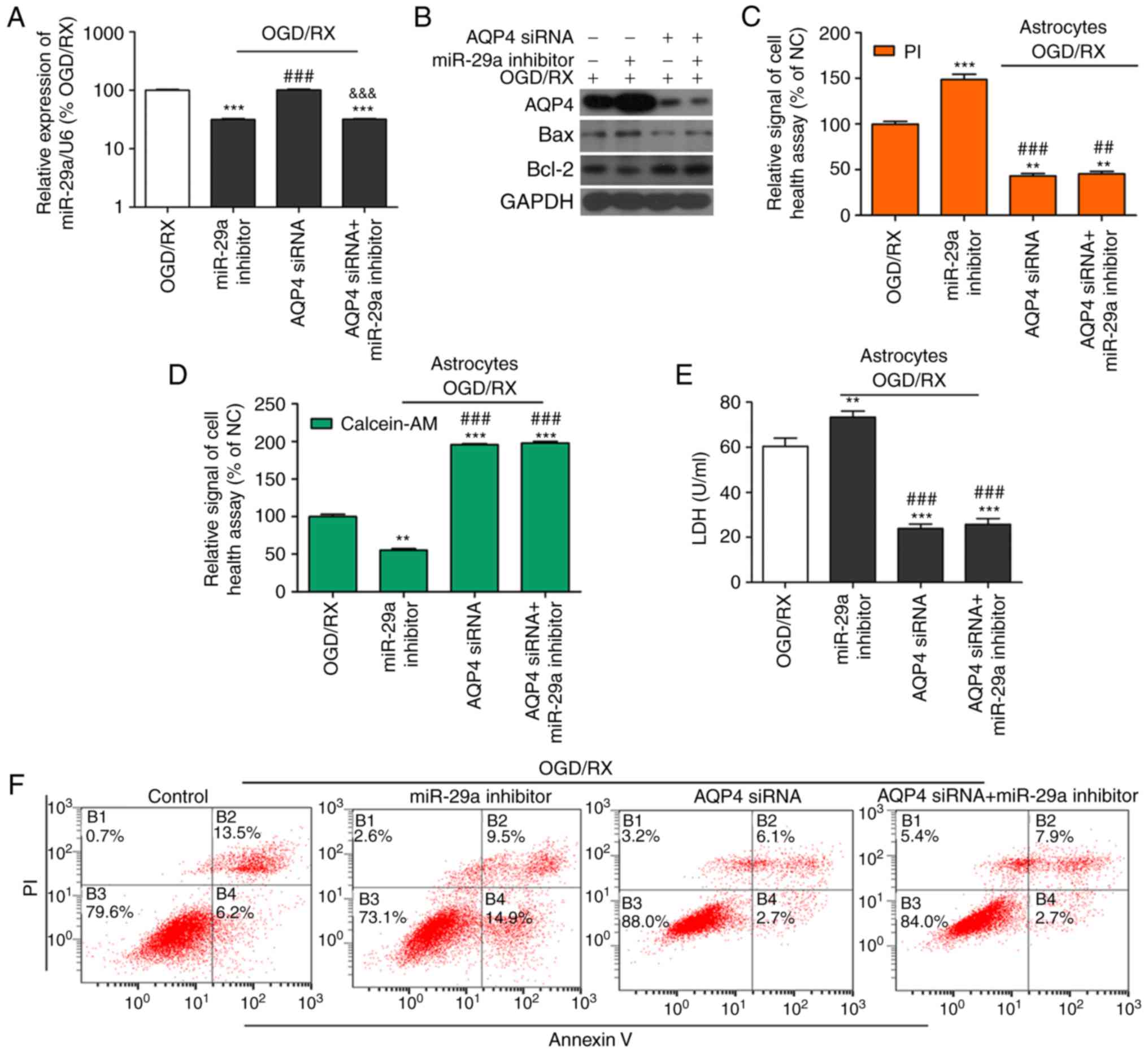 | Figure 5.AQP4 mediates the protective effects
of miR-29a in ischemia-induced astrocyte cell injury. (A) qRT-PCR
determination of the expression of miR-29a in OGD/RX-induced
astrocytes or NC. ***P<0.001 vs. NC; ###P<0.001
vs. miR-29 inhibitor, &&&P<0.001 vs. AQP4
siRNA. (n=3 for each experiment). (B) Western blot analysis of the
expression of miR-29a in astrocytes treated with OGD for 6 h
following different groups. All experiments were repeated in
triplicate. Bands have been cropped from different parts of
different gels, and respective western blot images have been
cropped for Figure implementation. (C and D) Cell health assay of
OGD-treated primary cultured astrocytes co-transfected with or
without AQP4 siRNA and miR-29a inhibitor following PI/calcein
staining. **P<0.01, ***P<0.001 vs. OGD/RX;
##P<0.01, ###P<0.001 vs. OGD/RX+miR-29a
inhibitor (n=3 for each experiment). (E) Expression of LDH assessed
by ELISA. **P<0.01, ***P<0.001 vs. OGD/RX;
###P<0.001 vs. OGD/RX+miR-29a inhibitor (n=3). (F and
G) Apoptotic cells among OGD-induced astrocytes co-transfected with
or without AQP4 siRNA and miR-29a inhibitor estimated by flow
cytometry (n=3 for each experiment). **P<0.01, ***P<0.001 vs.
OGD/RX; ###P<0.001 vs. OGD/RX+miR-29a inhibitor. (H)
H&E detection of cell damage. (I) Bar graph of calculated brain
oedema volume. *P<0.05, **P<0.01, ***P<0.001 vs. OGD/RX;
##P<0.01 vs. OGD/RX+miR-29a inhibitor. AQP4,
aquaporin 4; OGD/RX, oxygen glucose deprivation-reoxygenation; NC,
negative control; PI, propidium iodide. |
Discussion
Ischemic stroke results in a high rate of
disability, morbidity and mortality, and is a leading cause of
brain disease (36). MicroRNAs
(miRNAs) are a family of small, genome-encoded endogenous RNAs that
are transcribed but not translated into proteins (37). They act as important regulators in
many types of human diseases. Accumulating evidence suggests that
the expression of aberrant miRNAs plays a vital role in ischemic
stroke and other pathologies (37,38).
In the present study, the effects of miR-29a on OGD/RX-treated
primary astrocytes and potential targeting of AQP4 were
researched.
In humans and rodents, miR-29 members are highly
conserved and include three subtypes; miR-29a, miR-29b and miR-29c
(38). It has been reported that
the focal ischemic cortex is downregulated (39), and increased in the forebrain
ischemic hippocampus (40). miR-29a
inhibitors enhance cell injury, while overexpression of miR-29a
protects against ischemia-reperfusion injury (41). miR-29a mimics enhance cell survival
in astrocytes by inhibiting the expression of VDAC1 (42). The present study indicated that
miR-29a overexpression could protect primary astrocytes from
ischemic injury. miR-29a mimics significantly decreased apoptosis
and promoted astrocyte cell health compared with OGD/RX-treated
primary astrocytes. In addition, miR-29a inhibition reversed these
effects. Quantitative RT-PCR demonstrated that the expression level
of miR-29a decreased in OGD/RX-treated primary astrocytes compared
with NC, and the miR-29a level was negatively associated with AQP4
in OGD/RX-treated primary astrocytes. Furthermore, miR-29a could
downregulate OGD/RX-mediated AQP4 expression, and miR-29a
downregulated AQP4 expression after transfection with the miR-29
mimic, while the miR-29a inhibitor upregulated AQP4 expression in
primary astrocytes under normoxic and OGD/RX conditions compared
with OGD/RX. However, these effects were no longer present after
knockdown of AQP4. After ischemic stroke, miR-455 was revealed to
inhibit the survival of neuronal cells by inhibiting TRAF3
expression (43), and the miR-29b
mimic could reduce cell injury by decreasing AQP4 expression after
ischemic stroke (28). Recent
studies demonstrated a role for miR138 in the treatment of cerebral
ischemia reperfusion injury (44).
Our present results indicated that the protective role of miR-29a
in OGD/RX-treated primary astrocytes may be mediated by inhibiting
the expression of AQP4, and that miR-29a may be an effective
therapeutic target for cerebral ischemic stroke. AQP4 may be the
specific gene regulated by some underlying miRNAs.
AQP4 is expressed in glial cells, capillary
endothelial cells in the brain, and particularly the cerebrospinal
fluid, which contains numerous membrane proteins (45). AQP4 regulates water metabolism, its
expression is altered following brain injury, and this protein is
associated with changes in pathological status in the damaged side
of the brain (27). A previous
study revealed that MSC treatment maintained blood-brain barrier
integrity by inhibiting the expression of AQP4 after cerebral
ischemia (46). Research
demonstrated that the level of AQP4 was significantly decreased by
OGD/RX injury, but increased after reoxygenation (47). In rat ischemic stroke models, AQP4
aggravated brain ischemia by enhancing cerebral oedema (48–50).
In the present study, TargetScan was used to reveal that AQP4 is a
potential target for the regulation of miR-29a at the
post-transcriptional level; miR-29a could bind to the 3′-UTR of
AQP4 mRNA. In addition, it was revealed that AQP4 expression was
increased in OGD/RX-treated primary astrocytes compared with
NC.
It has been indicated that knockdown of AQP4 could
improve patient outcome and neurological function, reduce
infarction volume, increase neuronal survival, and reduce apoptosis
and the inflammatory response following cerebral ischemia, in
accordance with brain oedema reduction (51). In the present study, it was
demonstrated that knockdown of AQP4 decreased apoptosis, altered
the expression of apoptotic-related proteins, and promoted
astrocyte cell health compared with OGD/RX-treated primary
astrocytes. These findings indicated that AQP4 may regulate cell
death in OGD/RX-treated primary astrocytes. Furthermore, it was
confirmed that miR-29a overexpression could protect primary
astrocytes from ischemic injury, and miR-29a inhibitors could
damage primary astrocytes from ischemic injury, possibly by
regulating the expression of AQP4. However, the effects of miR-29a
transfection and the miR-29a inhibitor were reversed after
knockdown of AQP4 in primary astrocytes.
In conclusion, our findings demonstrated that
miR-29a overexpression reduced OGD/RX-induced injury in primary
cultured astrocytes via AQP4 mediation. Therefore, miR-29a could be
used as a novel biomarker in stroke, and may be an effective
therapeutic agent for the treatment of cerebral ischemia.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81271274, 81771194
and 81471171), the Zhejiang Medical Technology and Education
(2014KYA088 and 2017KY323), and the Project of Experimental Animal
Science and Technology Plan of Zhejiang Province (no.
2018C37116).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
SZ and YZ conceived and designed the study. YZ, LX
and CP performed the experiments. YZ, MC, AP and SZ analyzed the
data. YZ and AP wrote the manuscript. All authors read and approved
the manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
The present study concerning animal experiments was
approved by the Medical Ethics Committee and the Medical Faculty
Ethics Committee of The First Affiliated Zhejiang Hospital,
Zhejiang University (Ref. no. 2018-811).
Patient consent for publication
Not applicable.
Competing interests
All authors declare that they have no competing
interests.
References
|
1
|
Liu P, Zhao H, Wang R, Wang P, Tao Z, Gao
L, Yan F, Liu X, Yu S, Ji X, et al: MicroRNA-424 protects against
focal cerebral ischemia and reperfusion injury in mice by
suppressing oxidative stress. Stroke. 46:513–519. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yin KJ, Deng Z, Huang H, Hamblin M, Xie C,
Zhang J and Chen YE: miR-497 regulates neuronal death in mouse
brain after transient focal cerebral ischemia. Neurobiol Dis.
38:17–26. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tan JR, Tan KS, Koo YX, Yong FL, Wang CW,
Armugam A and Jeyaseelan K: Blood microRNAs in low or no risk
ischemic stroke patients. Int J Mol Sci. 14:2072–2084. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Saito Y and Saito H: MicroRNAs in cancers
and neurodegenerative disorders. Front Genet. 3:1942012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rink C and Khanna S: MicroRNA in ischemic
stroke etiology and pathology. Physiol Genomics. 43:521–528. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jickling GC, Ander BP, Zhan X, Noblett D,
Stamova B and Liu D: microRNA expression in peripheral blood cells
following acute ischemic stroke and their predicted gene targets.
PLoS One. 9:e992832014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bhalala OG, Srikanth M and Kessler JA: The
emerging roles of microRNAs in CNS injuries. Nat Rev Neurol.
9:328–339. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sun Y, Gui H, Li Q, Luo ZM, Zheng MJ, Duan
JL and Liu X: MicroRNA-124 protects neurons against apoptosis in
cerebral ischemic stroke. CNS Neurosci Ther. 19:813–819.
2013.PubMed/NCBI
|
|
10
|
Guo X, Liu Q, Wang G, Zhu S, Gao L, Hong
W, Chen Y, Wu M, Liu H, Jiang C, et al: microRNA-29b is a novel
mediator of Sox2 function in the regulation of somatic cell
reprogramming. Cell Res. 23:142–156. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pfaff N, Fiedler J, Holzmann A, Schambach
A, Moritz T, Cantz T and Thum T: miRNA screening reveals a new
miRNA family stimulating iPS cell generation via regulation of
Meox2. EMBO Rep. 12:1153–1159. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yang CS, Li Z and Rana TM: microRNAs
modulate iPS cell generation. RNA. 17:1451–1460. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Smirnova L, Gräfe A, Seiler A, Schumacher
S, Nitsch R and Wulczyn FG: Regulation of miRNA expression during
neural cell specification. Eur J Neurosci. 21:1469–1477. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mott JL, Kurita S, Cazanave SC, Bronk SF,
Werneburg NW and Fernandez-Zapico ME: Transcriptional suppression
of mir-29b-1/mir-29a promoter by c-Myc, hedgehog, and NF-kappaB. J
Cell Biochem. 110:1155–1164. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tan KS, Armugam A, Sepramaniam S, Lim KY,
Setyowati KD, Wang CW and Jeyaseelan K: Expression profile of
MicroRNAs in young stroke patients. PLoS One. 4:e76892009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu DZ, Tian Y, Ander BP, Xu H, Stamova
BS, Zhan X, Turner RJ, Jickling G and Sharp FR: Brain and blood
microRNA expression profiling of ischemic stroke, intracerebral
hemorrhage, and kainate seizures. J Cereb Blood Flow Metab.
30:92–101. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hutchison ER, Okun E and Mattson MP: The
therapeutic potential of microRNAs in nervous system damage,
degeneration, and repair. Neuromolecular Med. 11:153–161. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao H, Wang J, Gao L, Wang R, Liu X, Gao
Z, Tao Z, Xu C, Song J, Ji X, et al: MiRNA-424 protects against
permanent focal cerebral ischemia injury in mice involving
suppressing microglia activation. Stroke. 44:1706–1713. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Stary CM, Xu L, Sun X, Ouyang YB, White
RE, Leong J, Li J, Xiong X and Giffard RG: MicroRNA-200c
contributes to injury from transient focal cerebral ischemia by
targeting Reelin. Stroke. 46:551–556. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou J and Zhang J: Identification of
miRNA-21 and miRNA-24 in plasma as potential early stage markers of
acute cerebral infarction. Mol Med Rep. 10:971–976. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yi H, Huang Y, Yang F, Liu W, He S and Hu
X: MicroRNA-182 aggravates cerebral ischemia injury by targeting
inhibitory member of the ASPP family (iASPP). Arch Biochem Biophys.
620:52–58. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Doeppner TR, Doehring M, Bretschneider E,
Zechariah A, Kaltwasser B, Müller B, Koch JC, Bähr M, Hermann DM
and Michel U: MicroRNA-124 protects against focal cerebral ischemia
via mechanisms involving Usp14-dependent REST degradation. Acta
Neuropathol. 126:251–265. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nielsen S, Nagelhus EA, Amiry-Moghaddam M,
Bourque C, Agre P and Ottersen OP: Specialized membrane domains for
water transport in glial cells: High-resolution immunogold
cytochemistry of aquaporin-4 in rat brain. J Neurosci. 17:171–180.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nagelhus EA, Mathiisen TM and Ottersen OP:
Aquaporin-4 in the central nervous system: Cellular and subcellular
distribution and coexpression with KIR4.1. Neuroscience.
129:905–913. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Costa C, Tortosa R, Domènech A, Vidal E,
Pumarola M and Bassols A: Mapping of aggrecan, hyaluronic acid,
heparan sulphate proteoglycans and aquaporin 4 in the central
nervous system of the mouse. J Chem Neuroanat. 33:111–123. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Manley GT, Fujimura M, Ma T, Noshita N,
Filiz F, Bollen AW, Chan P and Verkman AS: Aquaporin-4 deletion in
mice reduces brain edema after acute water intoxication and
ischemic stroke. Nat Med. 6:159–163. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang C, Chen J and Lu H: Expression of
aquaporin-4 and pathological characteristics of brain injury in a
rat model of traumatic brain injury. Mol Med Rep. 12:7351–7357.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang Y, Huang J, Ma Y, Tang G, Liu Y, Chen
X, Zhang Z, Zeng L, Wang Y, Ouyang YB and Yang GY: MicroRNA-29b is
a therapeutic target in cerebral ischemia associated with aquaporin
4. J Cereb Blood Flow Metab. 35:1977–1984. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Oh TW, Park KH, Jung HW and Park YK:
Neuroprotective effect of the hairy root extract of Angelica
gigas NAKAI on transient focal cerebral ischemia in rats
through the regulation of angiogenesis. BMC Complement Altern Med.
15:1012015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li Q, He Q, Baral S, Mao L, Li Y, Jin H,
Chen S, An T, Xia Y and Hu B: MicroRNA-493 regulates angiogenesis
in a rat model of ischemic stroke by targeting MIF. FEBS J.
283:1720–1733. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Iwata-Ichikawa E, Kondo Y, Miyazaki I,
Asanuma M and Ogawa N: Glial cells protect neurons against
oxidative stress via transcriptional up-regulation of the
glutathione synthesis. J Neurochem. 72:2334–2344. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lin SP, Ye S, Long Y, Fan Y, Mao HF, Chen
MT and Ma QJ: Circular RNA expression alterations are involved in
OGD/R-induced neuron injury. Biochem Biophys Res Commun. 471:52–56.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zheng L, Cheng W, Wang X, Yang Z, Zhou X
and Pan C: Overexpression of MicroRNA-145 ameliorates astrocyte
injury by targeting aquaporin 4 in cerebral ischemic stroke. Biomed
Res Int 2017. 95309512017.
|
|
34
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang J, Li Y, Tang Y, Tang G, Yang GY and
Wang Y: CXCR4 antagonist AMD3100 protects blood-brain barrier
integrity and reduces inflammatory response after focal ischemia in
mice. Stroke. 44:190–197. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang P, Zhang N, Liang J and Li J, Han S
and Li J: Micro-RNA-30a regulates ischemia-induced cell death by
targeting heat shock protein HSPA5 in primary cultured cortical
neurons and mouse brain after stroke. J Neurosci Res. 93:1756–1768.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Martinez B and Peplow PV: Blood microRNAs
as potential diagnostic and prognostic markers in cerebral ischemic
injury. Neural Regen Res. 11:1375–1378. 2016.PubMed/NCBI
|
|
38
|
Kriegel AJ, Liu Y, Fang Y, Ding X and
Liang M: The miR-29 family: Genomics, cell biology, and relevance
to renal and cardiovascular injury. Physiol Genomics. 44:237–244.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jeyaseelan K, Lim KY and Armugam A:
MicroRNA expression in the blood and brain of rats subjected to
transient focal ischemia by middle cerebral artery occlusion.
Stroke. 39:959–966. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu P, Sun J, Zhao J, Liu X, Gu X, Li J,
Xiao T and Xu LX: Microvascular imaging using synchrotron
radiation. J Synchrotron Radiat. 17:517–521. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ouyang YB, Xu L, Lu Y, Sun X, Yue S, Xiong
XX and Giffard RG: Astrocyte-enriched miR-29a targets PUMA and
reduces neuronal vulnerability to forebrain ischemia. Glia.
61:1784–1794. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Stary CM, Sun X, Ouyang Y, Li L and
Giffard RG: miR-29a differentially regulates cell survival in
astrocytes from cornu ammonis 1 and dentate gyrus by targeting
VDAC1. Mitochondrion. 30:248–254. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yao S, Tang B, Li G, Fan R and Cao F:
miR-455 inhibits neuronal cell death by targeting TRAF3 in cerebral
ischemic stroke. Neuropsychiatr Dis Treat. 12:3083–3092. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tang XJ, Yang MH, Cao G, Lu JT, Luo J, Dai
LJ, Huang KM and Zhang LI: Protective effect of microRNA-138
against cerebral ischemia/reperfusion injury in rats. Exp Ther Med.
11:1045–1050. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xu J, Qiu GP, Huang J, Zhang B, Sun SQ,
Gan SW, Lu WT, Wang KJ, Huang SQ and Zhu SJ: Internalization of
aquaporin-4 after collagenase-induced intracerebral hemorrhage.
Anat Rec. 298:554–561. 2015. View Article : Google Scholar
|
|
46
|
Tang G, Liu Y, Zhang Z, Lu Y, Wang Y,
Huang J, Li Y, Chen X, Gu X, Wang Y, et al: Mesenchymal stem cells
maintain blood-brain barrier integrity by inhibiting aquaporin-4
upregulation after cerebral ischemia. Stem Cells. 32:3150–3162.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Nito C, Kamada H, Endo H, Narasimhan P,
Lee YS and Chan PH: Involvement of mitogen-activated protein kinase
pathways in expression of the water channel protein aquaporin-4
after ischemia in rat cortical astrocytes. J Neurotrauma.
29:2404–2412. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Fukuda AM and Badaut J: Aquaporin 4: A
player in cerebral edema and neuroinflammation. J
Neuroinflammation. 9:2792012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Thrane AS, Rappold PM, Fujita T, Torres A,
Bekar LK, Takano T, Peng W, Wang F, Rangroo Thrane V, Enger R, et
al: Critical role of aquaporin-4 (AQP4) in astrocytic
Ca2+ signaling events elicited by cerebral edema. Proc
Natl Acad Sci USA. 108:846–851. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zeng HK, Wang QS, Deng YY, Fang M, Chen
CB, Fu YH, Jiang WQ and Jiang X: Hypertonic saline ameliorates
cerebral edema through downregulation of aquaporin-4 expression in
the astrocytes. Neuroscience. 166:878–885. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Tang G and Yang GY: Aquaporin-4: A
potential therapeutic target for cerebral edema. Int J Mol Sci.
17:E14132016. View Article : Google Scholar : PubMed/NCBI
|