Natural killer (NK) cells and their
markers
NK cells are innate lymphoid immune cells comprising
5–20% of the peripheral blood, and serve an important role in the
immune control of cancer (1,2). They
are identified by the expression of the surface markers cluster of
differentiation (CD)56 and CD16, and the lack of CD3; the
CD3− subset therefore indicates T-cell depletion. The
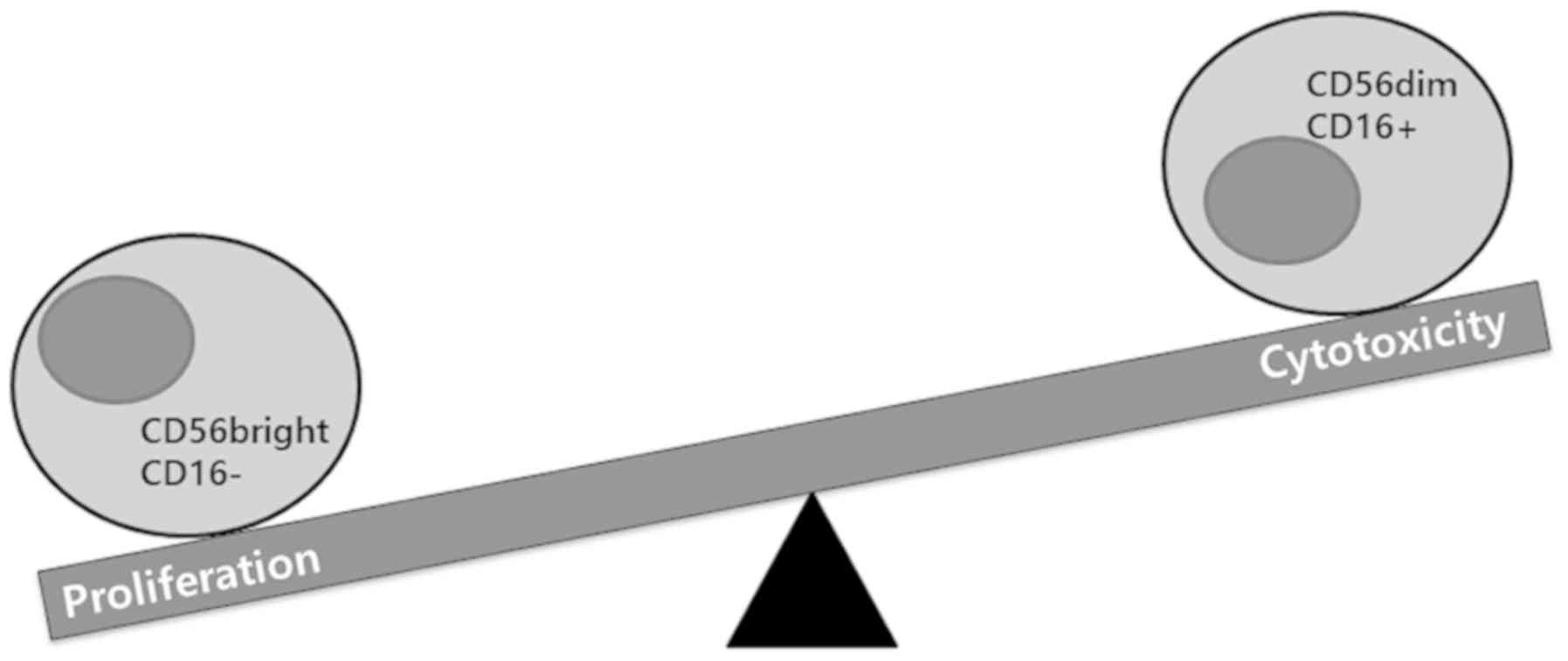
two major NK cell subset markers, CD56 and CD16, are present in
human cells, and induce different functions in NK cells. The
CD56brightCD16− subset has specialized
proliferative potential, whereas CD56dimCD16+
facilitates the cytolytic activity of NK cells. The higher density
of lytic granules in CD56dim cells and the capacity to
trigger cytokine production of CD16+ cells confers
cytotoxic activity on NK cells (1–3)
(Fig. 1).
Types of NK cell receptors
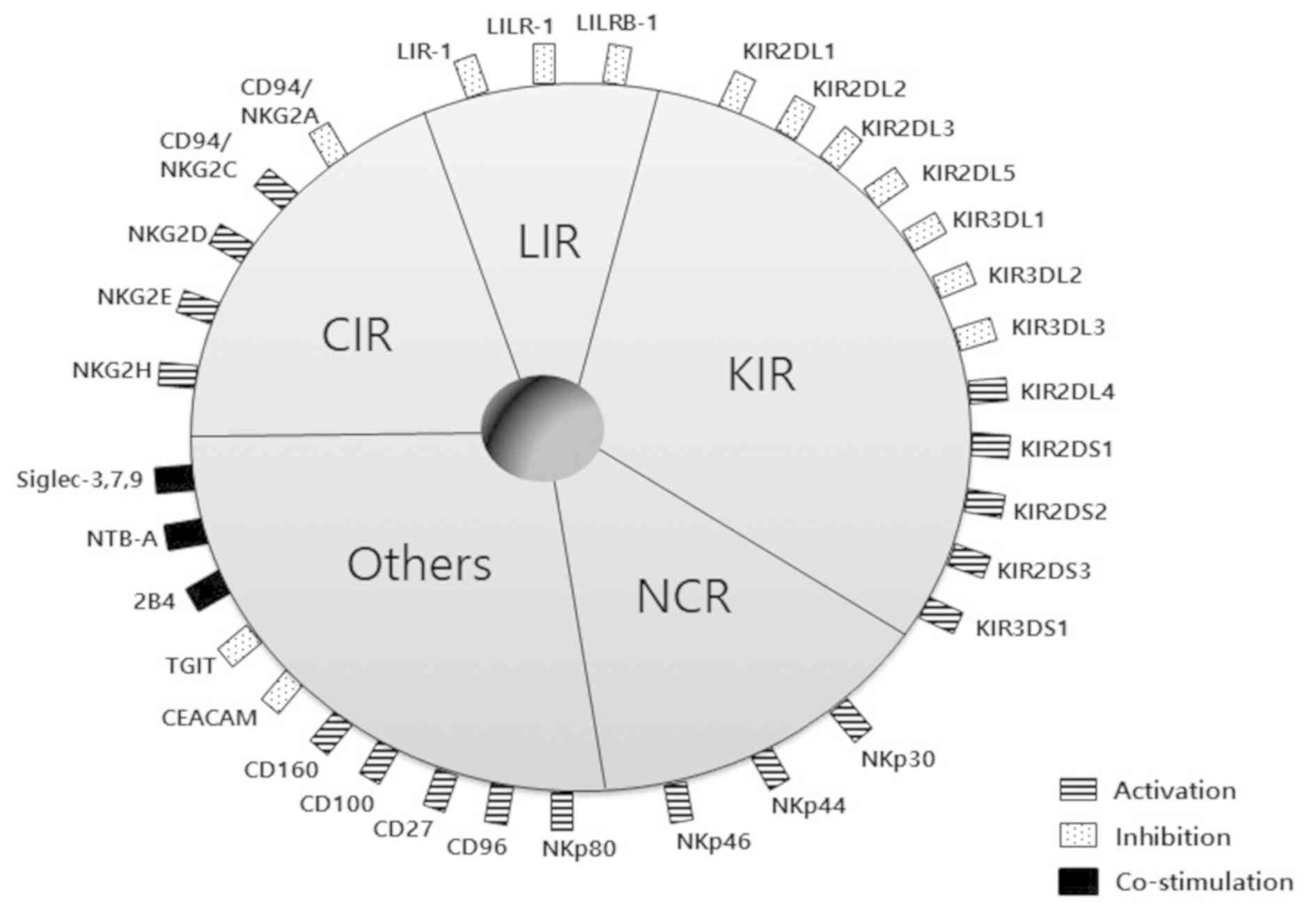
NK cell activity is adapted by a number of gene
families encoding inhibitory and activating receptors. The major
histocompatibility complex (MHC) class I-binding or non-binding
receptors include two principal types of receptors. The NK cell
receptor families belonging to MHC class I-binding receptors are
the c-type lectin receptors (CLRs), apart from NKG2-D type II
integral membrane protein (NKG2D), leukocyte immunoglobulin-like
receptors (LIRs) and killer cell immunoglobulin-like receptors
(KIRs). On the other hand, natural cytotoxicity receptors (NCRs)
and other NK cell receptors recognize tumor cells independently of
MHC class I molecules (1) (Fig. 2).
The CLRs of NK cell receptors structurally contain
the c-type lectin domain in their extracellular region. Lectins
contain a carbohydrate-binding protein domain, and the c-type form
requires calcium in order to bind with proteins. These lectins are
diversely functional in their immunopathological reactions in the
context of NK cell interactions.
The LIRs are a multigene family located on
chromosome 19, including the KIR gene family, containing
Ig-like extracellular domains in their cytoplasmic tails. Even
though LIRs bind to MHC class I molecules, they do not have an
important role in NK cell interactions since the binding of LIR to
UL18 has a higher affinity compared with the binding of LIR-1 to
MHC class I. The LIR uses the same binding site to interact with
MHC class I molecules or UL18, an example of which being the human
cytomegalovirus genes (1,4–6).
There are two types of KIRs: The inhibitory KIR
(iKIR) and activating KIR (aKIR). Structurally, KIRs have two or
three immunoglobulin-like domains that are transmembrane
glycoproteins. The cytoplasmic tail of iKIR contains the
immunoreceptor tyrosine-based inhibitory motif (ITIM), whereas the
short cytoplasmic tails of aKIR contain the immunoreceptor
tyrosine-based activation motif (ITAM) and have a positively
charged residue.
CD56dimCD16+ NK cells express
high levels of KIRs and CLRs compared with
CD56brightCD16− NK cells. Furthermore,
CD56dimCD16+ NK cells have a higher affinity
and recognize target cells bearing MHC class I ligands, thereby
exerting greater cytotoxicity compared with
CD56brightCD16− NK cells. Therefore, this
review focuses on the KIRs of NK cell receptors, due to their
increased recognition and interaction with MHC class I molecules on
target cells (4).
The NCRs and other NK cell receptors recognize tumor
cells irrespective of MHC class I. NCRs are of three types: NKp46,
NKp44 and NKp30. NKp30 have a crystal and dimeric structure, NKp44
comprises a typical v-type Ig-fold, whereas NKp46 consists of
labeled β strands. However, at present, the NCRs and their ligand
interactions are poorly understood (5,7).
Fundamentals of NK cell alloreactivity
(inhibitory receptors)
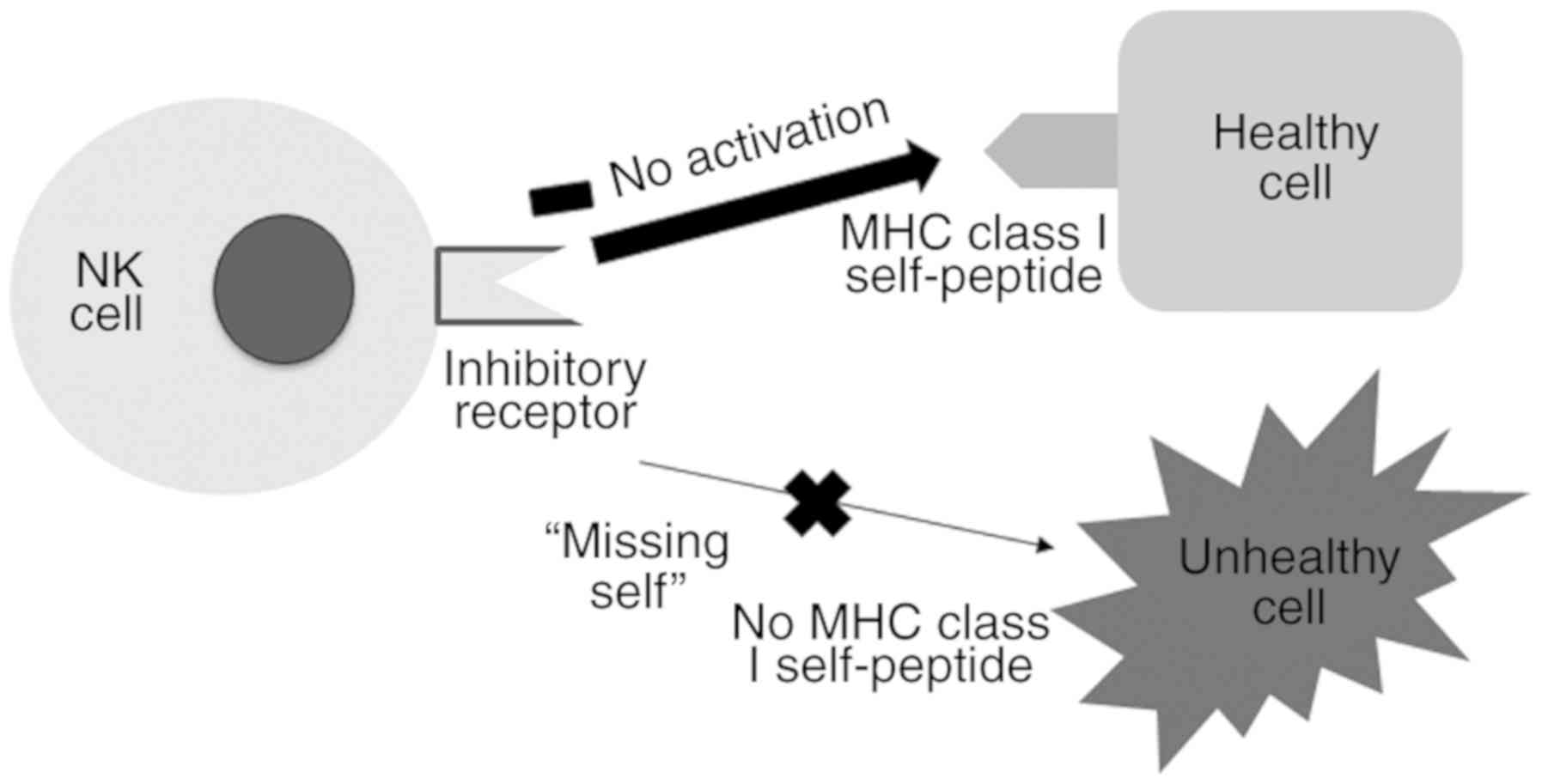
Since NK cells spontaneously circulate throughout
the human body via the peripheral blood, it is essential to have
inhibitory receptors that impede their attack on healthy tissues.
The majority of inhibitory receptors recognize the MHC class I
molecules present on the surface of cells. Since MHC class I
molecules are downregulated in tumor cells or virus-infected cells
(unhealthy cells), the NK cells are able to selectively eliminate
these unhealthy cells via a process termed ‘missing-self’ (8) (Fig.
3). CD94/NKG2-A/NKG2-B type II integral membrane protein, the
principal inhibitory receptor of NK cells, which binds to the MHC
class I molecule HLA-E, is present on target cells (9). The other inhibitory receptors are
primarily iKIRs, which bind to the MHC class I molecules HLA-A, B
and C (9,10). Since NK cell receptors are involved
in the binding of ligands, it is important to establish strategies
that kill tumor cells via NK cell cytotoxicity.
Inhibitory NK cell receptors exert their effect by
obstructing the activation of NK cells. The inhibitory NK cell
receptors generally express the ITIM, which recruits and
phosphorylates Src homology (SH)-containing tyrosine phosphatase
(SHP)-1 and SHP-2. Inhibition by ITIM-bearing receptors hampers the
tyrosine phosphorylation of essential signaling components,
including tyrosine phosphoprotein SLP-76 (SLP-76) and
proto-oncogene vav (Vav), to activate signaling in NK cells.
However, non-MHC-binding inhibitory receptor mechanisms have
various signaling pathways (11–14)
(Fig. 4). Specifically, 2B4 of NK
cell receptors bind to CD48, which exists on all human
hematopoietic cells. 2B4 comprises four tyrosine motifs (TxYxxI/V)
in the cytoplasmic tail, and the immunoreceptor tyrosine-based
switch motif interacts with SLAM-associated protein. The function
of 2B4 has been demonstrated to involve the recruitment of
signaling components of other activating receptors (12).
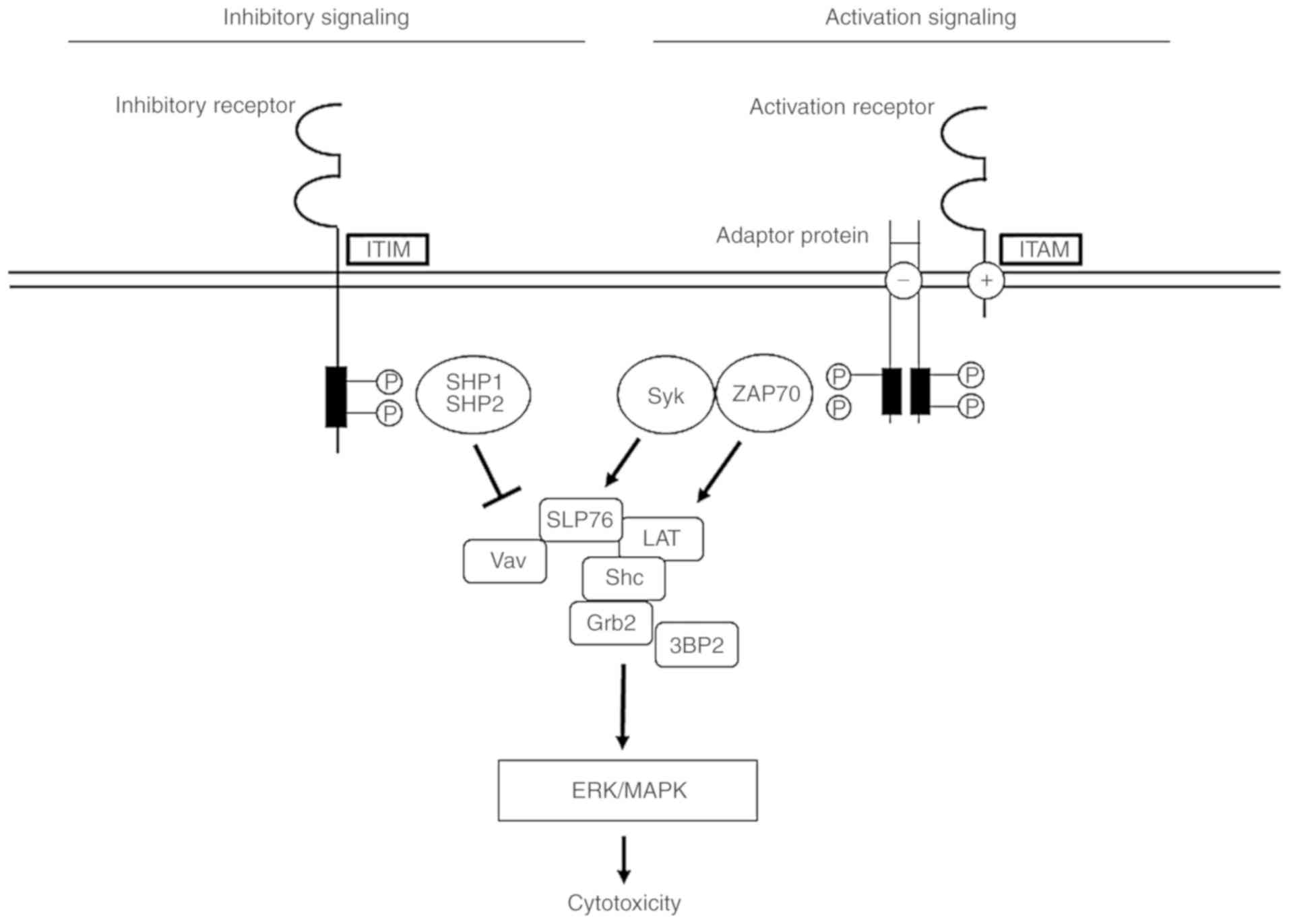 | Figure 4.NK cell receptor signaling pathways
associated with ITIM- or ITAM-containing adaptor proteins. In these
models, the phosphorylation of ITIM leads to the recruitment of
SHP-1 and SHP-2, which disturb SLP76 and Vav and are associated
with activating signals. On the other hand, the phosphorylation of
adaptor proteins with an ITAM triggers the recruitment of Sky and
ZAP-70, which activate various signaling molecules, including
SLP-76, LAT, Shc, Gb2 and 3BP2. Signals for cytotoxicity result in
the activation of downstream activating pathways, via the ERK/MARK
pathway. ITIM, immunoreceptor tyrosine-based inhibitory motif;
ITAM, immunoreceptor tyrosine-based activation motif; NK, natural
killer; SHP, Src homology-containing tyrosine phosphatase; SLP-76,
tyrosine phosphoprotein SLP-76; Vav, proto-oncogene vav; Syk,
tyrosine-protein kinase SYK; ZAP70, tyrosine-protein kinase ZAP-70;
LAT, linker for activation of T-cells family member 1; Shc,
SHC-transforming protein 1; Grb2, growth factor receptor-bound
protein 2; 3BP2, SH3 domain-binding protein 2; MAPK,
mitogen-activated protein kinase; ERK, extracellular
signal-regulated kinase. |
Fundamentals of NK cell alloreactivity
(activating receptors)
The activation of NK cells requires external signals
via their activating receptors that bind to the ligands on target
cells. Other important activating receptors include the NKG2
members (excluding A and B), NCRs and aKIRs. NKG2C, E and H bind to
the MHC class I molecule HLA-E, whereas NKG2D binds to MHC class I
chain-related genes A and B (MICA and MICB) and UL-16
protein-ligand family 1 and 6 (ULBP1 and ULBP6). aKIR binds to the
MHC class I molecules HLA-C or G, and another unknown receptor
(3,14–16).
The activating NK receptors possess small
transmembrane-anchored adaptor proteins that express ITAM in
addition to ITAM-bearing polypeptides (CD3ζ, FcRγ, DAP10 and DAP12)
in their intracellular membrane. ITAM-bearing receptor complexes
facilitate the recruitment of tyrosine-protein kinase SYK (Syk) and
tyrosine-protein kinase ZAP-70 (ZAP70). Moreover, since CD3ζ and
FcRγ facilitate CD16 signaling in NK cells, the ITAM-bearing
receptor complexes phosphorylate src family kinases. The
recruitment and phosphorylation of Syk and ZAP70 promote the
signals that trigger the activation of SLP-76, SHC-transforming
protein 1 (Shc), linker for activation of T-cells family member 1
(LAT), growth factor receptor-bound protein 2 and SH3
domain-binding protein 2, which are linkers for the activation of
proto-oncogene Vav and Ras-related C3 botulinum toxin substrate 1.
These signals produce downstream events, namely the activation of
the mitogen-activated protein kinase (MAPK) and extracellular
signal-regulated kinase (ERK) pathway (11–14)
(Fig. 4).
One example is the NCRs, comprising three major
proteins (NKp46, NKp30 and NKp44) that have an adaptor protein,
ITAM. Activation of the NCRs is associated with ITAM-bearing
receptor complexes that induce signaling pathways such as MAPK and
ERK. Other examples are the NKG2 members, which are associated with
TYRO protein tyrosine kinase-binding protein/DNAX-activation
protein (DAP) 10 or DAP12 adaptor proteins. In particular, NKG2D is
expressed on the NK cell surface and binds to its ligands on target
cells (MICA, MICB and ULBP) in humans, resulting in the expression
of DAP10 and DAP12. NKG2D is further classified as the short form
(NKG2D-S) or the long form (NKG2D-L), in line with the size of its
cytoplasmic tail. NKG2D-S binds to either DAP10 or DAP12, but NKG-L
is able to bind only to DAP10. In humans, NKG2D pairs only with
DAP10 subsequent to the stimulation of the p85 PI3
kinase-associated pathway. In addition, the NKG2D signaling pathway
has cross-linking signals that are associated with the activation
of the signal transducer and activator of transcription (STAT)5,
tyrosine-protein kinase JAK2 (JAK2), ERK, MAPK and
phosphatidylinositol 3-kinase/RAC-α serine/threonine-protein kinase
signal transduction pathways (11,13).
KIRs
Due to their individual diversity, KIRs serve a
pivotal role in human NK cells with respect to targeting cells. In
terms of gene content, KIR receptors are classified as iKIR (2DL1,
2DL2, 2DL3, 3DL1, 3DL2, 3DL3, and 2DL5A and B) and aKIR (2DL4,
2DS1, 2DS2, 2DS3, 2DS4, 2DS5 and 3DS1). The KIR gene content
defines the KIR haplotype of an individual. KIR haplotypes with
distinct gene content are distinguished as ‘A haplotype’ and ‘B
haplotype’. The A haplotype exists in all populations
conventionally, and has a fixed gene content of 9 genes (3DL3,
2DL3, 2DP1, 2DL1, 3DP1, 2DL4, 3DL1, 2DS4 and 3DL2). Different B
haplotypes exist among different people (2DS1, 2DS2, 2DS3, 2DS5,
2DL2, 2DL1 and 3DS1). It encodes more active KIRs compared with the
A haplotype (1,17,18).
Specifically, iKIRs have a long tail in their intracytoplasmic
region, denoted by the letter ‘L’; aKIRs have comparatively shorter
tails, indicated by the letter ‘S’. Also, KIR polymorphism is
characterized by a different structure, wherein the iKIR contains
the ITIM in the cytoplasmic tails, and aKIRs contain positively
charged amino acid residues in the transmembrane regions (19).
Although they recognize identical MHC class I
alleles, the inherent differences between iKIR and aKIR lead to
different outcomes, namely inhibitory or activating responses,
respectively. As previously described in this review, iKIR
signaling results from the existence of ITIMs that induce
inhibitory signaling in connection with SHP-1 and SHP-2.
Conversely, aKIR signaling involves ITAMs and may produce
ITAM-bearing receptor complexes that activate signaling molecules,
including Syk, ZAP70, SLP-76, Shc and LAT, resulting from the
activation of downstream signals, including the MAPK and ERK
pathway (4,19) (Fig.
4).
Models to determine NK cell KIR
alloreactivity for cell therapy
Functional capacity is induced by the interaction
between KIR receptors and specific KIR ligands. Inhibitory KIR
receptors predominantly bind to distinct MHC class I molecule HLA
subtypes, but the ligands for activating KIR receptors are unknown.
It has been identified that the activating KIR2DS1 recognizes MHC
class I molecule HLA-C subtype ligands (1,18).
Thus, the repertoire of NK cells with diverse combinations of KIRs
requires more detailed study to investigate KIR-KIR ligand matches
or mismatches due to their requirement for type specificity
(3,17) (Fig.
5). Notably, an A haplotype KIR-KIRL mismatched donor-recipient
has been described in allogenic hematopoietic transplantation for
the treatment of leukemia (20).
The process of ‘licensing’ of NK cells may be beneficial as a
graft-versus-host (GVH) effect. This means that the iKIR of
licensed NK cells is not recognized by KIR ligands, and the absence
of MHC class I molecule HLA subset expression on the leukemia cells
of the recipient is able to trigger NK cell alloreactivity. Donor
NK cell alloreactivity has a critical role in the GVH effect
compared with donor T cell allografts, since the alloreactivity of
T cells may negatively affect healthy recipient cells (18,21).
Mechanism of NK cell cytotoxicity in
cancer
NK cells release various cytokines and acquire
certain cytotoxic effects. NK cells recognize tumor cells by their
surface markers or receptors in order to activate NK functions via
either of the two tumor-recognizing models: ‘Missing-self
recognition’ and ‘stress-induced recognition’. The missing-self
recognition model encompasses a loss of the inhibitory effect of NK
cells resulting from the absence of MHC class I molecules on tumor
cells. In the stress-induced recognition model, damaged proteins on
tumors bind to the activating receptors of NK cells and stimulate
the cytotoxic functions of NK cells (22–24).
NK cell activation facilitates killing of tumor
cells either through direct or indirect means. In the first
mechanism, granules are released, including perforin and granzymes,
which induce apoptosis in tumor cells (22,25).
The second mechanism includes antibody-dependent cell-mediated
cytotoxicity, wherein the Fc region of the IgG antibody binds to
antigens on the tumor cells in addition to the CD16 receptor on NK
cells, forming cross-linking via CD16. The cross-linking of CD16
promotes the granule-dependent mechanism that kills the tumor cells
via the activated NK cells by apoptosis (22,26,27). A
third mechanism is death receptor-dependent apoptosis. This
mechanism involves the Fas ligand and tumor necrosis factor
(TNF)-related apoptosis-inducing ligand, which induce
caspase-dependent apoptosis in tumor cells (22,25,28).
An indirect model of activated NK cells involves the secretion of
cytokines and chemokines, including interferon (IFN)-α, TNF-α, C-C
motif chemokine (CCL)3, CCL4 and CCL5. A number of cytokines
enhance the proliferation and activation of immune cells, such as
monocytes, macrophages, dendritic cells (DCs), B-cells and
cytotoxic T lymphocytes (CTLs). These cells are cytotoxic towards
tumor cells, which die due to the immune response associated with
apoptosis and necrosis (22,25,26,29).
Further studies are required to focus on NK cell immunotherapy for
cancer, since NK cells serve an important role in the immune
response and are capable of killing cancer cells.
Types of leukemia and the therapeutic effect
of NK cells
Among hematological cancers, leukemia exhibits rapid
proliferation of abnormal white blood cells in the bone marrow.
Leukemias are classified into four major types: Acute lymphocytic
leukemia (ALL), acute myelogenous leukemia (AML), chronic
lymphocytic leukemia (CLL) and chronic myelogenous leukemia (CML).
It is important to understand the various leukemia types, since
each type requires a different treatment. In leukemia, NK cells
exert their potential therapeutic effect by eliminating abnormal
leukemic cells due to their cytotoxicity. Various therapeutic
options for leukemia have been associated with NK cell receptors
(16).
ALL, a cancer of the lymphoid cells characterized by
an increased number of lymphocytes, is the most common childhood
leukemia. Although ALL is resistant to NK cell cytotoxicity, recent
clinical studies have revealed the therapeutic effect of NK cells
in pediatric patients (1,17). CLL encompasses the B-cell subtype of
lymphocytes (B-CLL) in the bone marrow. Since CLL blasts do not
express the MICA, MICB and ULBP ligands, NK cells are capable of
eradicating these blasts (30–32).
CML involves the constant proliferation of myeloid cells,
particularly mature granulocytes, which are clonal bone marrow stem
cells. Thus, CML is referred to as a clonal disorder, since the
majority of patients with CML bear the distinctive feature of a
chromosomal translocation termed Philadelphia chromosome (Ph). Ph
is a mutual translocation between the 9th and 22nd chromosomes in
all hematopoietic cells; the Abelson (abl) gene on
chromosome 9 is transferred to the breakpoint cluster region
(bcr) gene from chromosome 22. This translocation results in
an abnormal ‘fusion’ termed the bcr/abl gene. The
bcr/abl gene develops a tyrosine kinase protein, since the
abl gene product adds phosphate groups to tyrosine residues
(33,34). As a result, CML is a clonal
myeloproliferative disease of abnormal translocation of the
bcr-abl oncogene, which produces the oncokinase protein
BCR-ABL, responsible for the pathogenesis of CML (34,35).
In the search for CML therapies, previous in
vitro experiments investigated NK-92MI cell line cytotoxicity
against the K562 leukemia cell line (36,37).
In addition, CML may be treated with tyrosine kinase inhibitors
(TKIs) due to the presence of high levels of MICA and low
expression of NKG2D (33,36). Therefore, for the effective
treatment of cancer using immunotherapy, such as NK cells, it is
important to understand the expression of ligands and receptors in
patients with hematological cancer, which are summarized in
Table I. This review particularly
focuses on anti-leukemic effects in CML. Notably, NK cells exert
their cytotoxic effect by eliminating CML leukemic cells through
lysis.
 | Table I.Expression of ligands and receptors
in patients with hematological cancer. |
Table I.
Expression of ligands and receptors
in patients with hematological cancer.
|
|
| Expression levels
in patients |
|
|---|
|
|
|
|
|
|---|
| Author, year | Type of cancer | Ligands | Receptors | (Refs.) |
|---|
| Nückel et
al, 2010 | CLL | MICA/B, absent;
ULBP, absent | − | (30) |
| Osman et al,
2017 | ALL | MICA/B,
downregulated; ULBP, downregulated | − | (32) |
| Osman et al,
2017 | AML | DNAM-1,
downregulated | NCR, downregulated;
NKG2D, downregulated; CD94/NKG2C, downregulated | (32) |
| Boissel et
al, 2006 | CML | MICA,
upregulated | NKG2D,
downregulated | (33) |
Therapeutic targets in CML
As previously discussed in this review, the
bcr-abl oncogene is a critical gene in CML (34,35).
Thus, novel strategies targeting CML use TKIs, including imatinib
mesylate, which provide improved long-term survival of patients
(38). In the majority of early
studies, researchers proposed that TKIs could be an important
therapeutic avenue for CML treatment, although there was limited
information about the underlying molecular mechanism. To be
specific, statistics indicate that imatinib mesylate is one of the
TKIs that induces a complete hematological and cytogenetic response
in ~83% of CML patients for 10 years (37,38).
However, certain patients with CML are resistant to the therapy due
to mutations in the ABL kinase domain; furthermore, the treatment
is associated with long-lasting side effects including nausea,
cramps and peripheral edema. In addition, TKI therapy involves a
high drug cost (39,40). Second-generation TKIs, including
dasatinib and nilotinib are under investigation for the treatment
of CML. To date, the number of patients with CML who have
benefitted from TKI treatment remains unknown (41–44).
Therefore, the development of new TKI agents and combination
therapies with other treatment strategies is urgently required for
TKI therapy. If certain patients with CML exhibit resistance to
TKIs, other approaches are required to improve the treatment of
CML. A number of patients with CML have exhibited a complete
cytogenetic response with no recurrence within 10 years following
IFN-α treatment (45). IFN-α
inhibits cell proliferation, regardless of the bcr-abl
oncogene. There remain certain problems associated with this
treatment, since certain CML patients receiving IFN-α therapy
experience side effects and develop tolerance to IFN-α (39,45).
Treatment with hydroxyurea, leukapheresis and stem
cell transplantation are other options in the treatment of CML.
Hydroxyurea is generally administered prior to treatment with
imatinib mesylate. Nevertheless, hydroxyurea has demonstrated
efficacy in patients with CML of up to 90%, but is not commonly
utilized in CML therapy for long-term administration due to its
inherent cytotoxicity (46–48). Leukapheresis is another treatment
applied to reduce the high levels of white blood cells in patients
with CML, and stem cell transplantation has also been considered as
another treatment option (49,50).
Considering all of the above, developing novel
effective treatment strategies for patients with CML is a clear
medical necessity. The understanding of tumor immunology has
allowed for considerable progress, which has been applied to the
immunotherapy of various tumors. Since immunosurveillance is
impaired in patients with CML, the lack of innate immune cells
including NK cells, B-cells, T-cells and dendritic cells results in
decreased quantitative and qualitative functions in the blood
(51).
Therapeutic targets in CML by
immunotherapy
Previous studies have included various
immunotherapeutic approaches for CML by targeting the underlying
pathways, such as the programmed cell death protein 1 (PD-1)/PD-1
ligand 1 (PDL-1), interleukin (IL)-1 and JAK/STAT pathways
(47–52) (Table
II).
| Table II.Immunotherapeutic approaches for
CML. |
Table II.
Immunotherapeutic approaches for
CML.
| Author, year | Pathway or
target | Treatment
strategy | (Refs.) |
|---|
| Cayssials and
Guilhot, 2017; Mahoney et al, 2015 | PD-1+ cytotoxic T
cells | Monoclonal
antibodies | (51,52) |
| Cayssials and
Guilhot, 2017; Mahoney et al, 2015; Mumprecht et al,
2009 | PD-L1+ myeloid
leukemic cells | PD-1/PD-L1
antagonists | (51–53) |
| Zhang et al,
2016; Stramucci and Perrotti, 2016; Ågerstam et al, 2016;
Arranz et al, 2017; Zhao et al, 2017 | IL-1 signaling in
CML stem cells | Monoclonal
antibodies CAR-T cells | (54–57) |
| Tarafdar et
al, 2017; Levescot et al, 2014; Rocca et al,
2018 | JAK/STAT
signaling | JAK/STAT inhibitors
(e.g. ruxolitinib) | (59–61) |
The PD-1/PDL-1 interaction may act as an immune
checkpoint inhibitor, having a marked effect on the anti-tumor
response. To facilitate signaling by PD-1, binding of PDL-1 is
essential. PD-1 signaling may be involved in the functional effect
of infiltrating T cells in tumors. In patients with CML, PD-1 is
highly expressed on CML-specific CTLs, whereas the CML cells
express PDL-1. Thus, PD-1/PDL-1 interaction leads to the exhaustion
and inhibition of CTLs in patients with CML (53). Therefore, blocking PD-1/PDL-1
binding as a treatment strategy represents a promising therapeutic
approach for CML (51–53). IL-1 is a regulator of inflammation
and the innate immune response, having numerous roles in
immunopathological functions. In hematological malignancies, drugs
that target IL-1 have an important therapeutic value. Recently,
there have been numerous clinical trials in patients focusing on
treatments targeting the IL-1 signaling pathway (54,55).
In CML, since IL-1 provides resistance to imatinib in CML, the
prevention of IL-1 signaling has the potential to increase the
efficacy of TKI treatment. Additionally, when using IFN-α treatment
in CML, IFN-α inhibits IL-1 due to its anti-inflammatory effect.
Hence, a combination therapy with TKIs and IFN-α may be a more
efficacious approach for CML patients. Meanwhile, IFN-α-resistant
CML patients have high levels of IL-1, which stimulates
IFN-α-sensitive CML cells. One way of the blocking IL-1 pathway is
using IL-1 receptor accessory (IL1RAP)-specific chimeric antigen
receptor-modified T (CAR-T) cells. IL1RAP-CAR-T cells represent an
important alternative therapy for patients with CML presenting with
TKI and IFN-α resistance (56–58).
The tyrosine kinase JNK has a critical role in
myelopoiesis by transducing cytokine signals, and binds to the
cytoplasmic domains of various cytokines and growth factor
receptors. Once the binding of ligands to their receptors in the
extracellular region occurs, JAK is phosphorylated in the
intracellular region. Phosphorylated JAK activates downstream
substrates such as the intracellular transcription factor STAT. The
JAK-STAT pathway is involved in facilitating myeloproliferation and
is abnormally activated in hematopoietic diseases (59,60).
Therefore, JAT-STAT inhibitors merit consideration for the
treatment of CML. Furthermore, patients with JAK-mutated CML may
benefit from TKI treatment, since the JAK-STAT pathway is not
normally activated. Thus, randomized trials are required to confirm
the potential advantages of the therapy in this cohort (61).
As a novel therapy for CML, NK cells have potential
for use in future therapeutic strategies. NK cells are cytotoxic
against malignant cells in leukemia, thereby mounting a defense
against tumor cells. In leukemic patients, NK cells decline as the
disease progresses from the leukemia blast crisis. Also, NK cells
isolated from patients with CML exhibit reduced cytotoxicity. Since
NK cells exert cellular cytotoxicity and produce an adaptive immune
response following the release of various cytokines, they serve an
important role in the antitumor immune response (36). Thus, the anti-leukemic function of
NK cells is a novel immunotherapeutic approach for CML.
Therapeutic targets in CML by NK cells
NK cells exert cytotoxic activity against tumor
cells and malignant cells in leukemia. Previous studies have
reported that the number and function of NK cells decreases in
leukemic patients. The anti-leukemic action of NK cells is
relatively low-level in patients with CML (62–64).
However, a previous study demonstrated that T cells and NK cells
are observed in patients as a consequence of receiving drug
treatment (65).
The NKG2D NK cell receptor is activated by
recognizing its ligands (NKG2DLs) on malignant cells, provoking the
cytotoxic action of NK cells. The majority of tumor cells express
NKG2DLs, including MICA, MICB, ULBP1 and ULBP2. MICA ligand is
expressed on all leukemia cells and is associated with the immune
response and anti-tumor activity. Thus, it is indicated that the
NKG2D/MICA interaction has an important role in NK cell
cytotoxicity against leukemia (62).
NK cells further influence the treatment of CML when
patients with CML are difficult to treat with TKIs. In
hematopoietic stem cell transplantation (HSCT), NK cell
alloreactivity diminishes the rate of leukemia relapse and protects
against an immunological GVH effect. This process is mediated by
the ‘missing-self’ recognition on NK cells, whereas this phenomenon
remains incomplete (66,67). Therefore, it is necessary to
investigate this process in order to develop an NK cell therapy for
CML in the future.
NK cell alloreactivity controls the GVH
effect
HSCT is also efficacious for myeloid malignancies,
which require high doses of chemotherapy (68,69).
HSCT exerts its influence by inducing an immunological GVH effect.
NK cells are thought to contribute to the GVH effect without
directly causing graft-versus-host disease (GVHD) (70). NK cell alloreactivity regulated by
their receptors is increased in the absence of HLA ligands. The
missing-self represents an inhibitory effect of NK cells, which
responds to activating signals to trigger the lysis of tumor
targets. In addition, NK cells work in combination with the
adaptive immune response to aggravate GVHD (70,71).
Adoptive NK cell therapy in
hematopoiesis
In hematopoietic transplants, donors having a
KIR-ligand mismatch may be selected, thereby creating an
environment in which recipient tumor cells act against alloreactive
donor NK cells due to the missing KIR ligand (67,70).
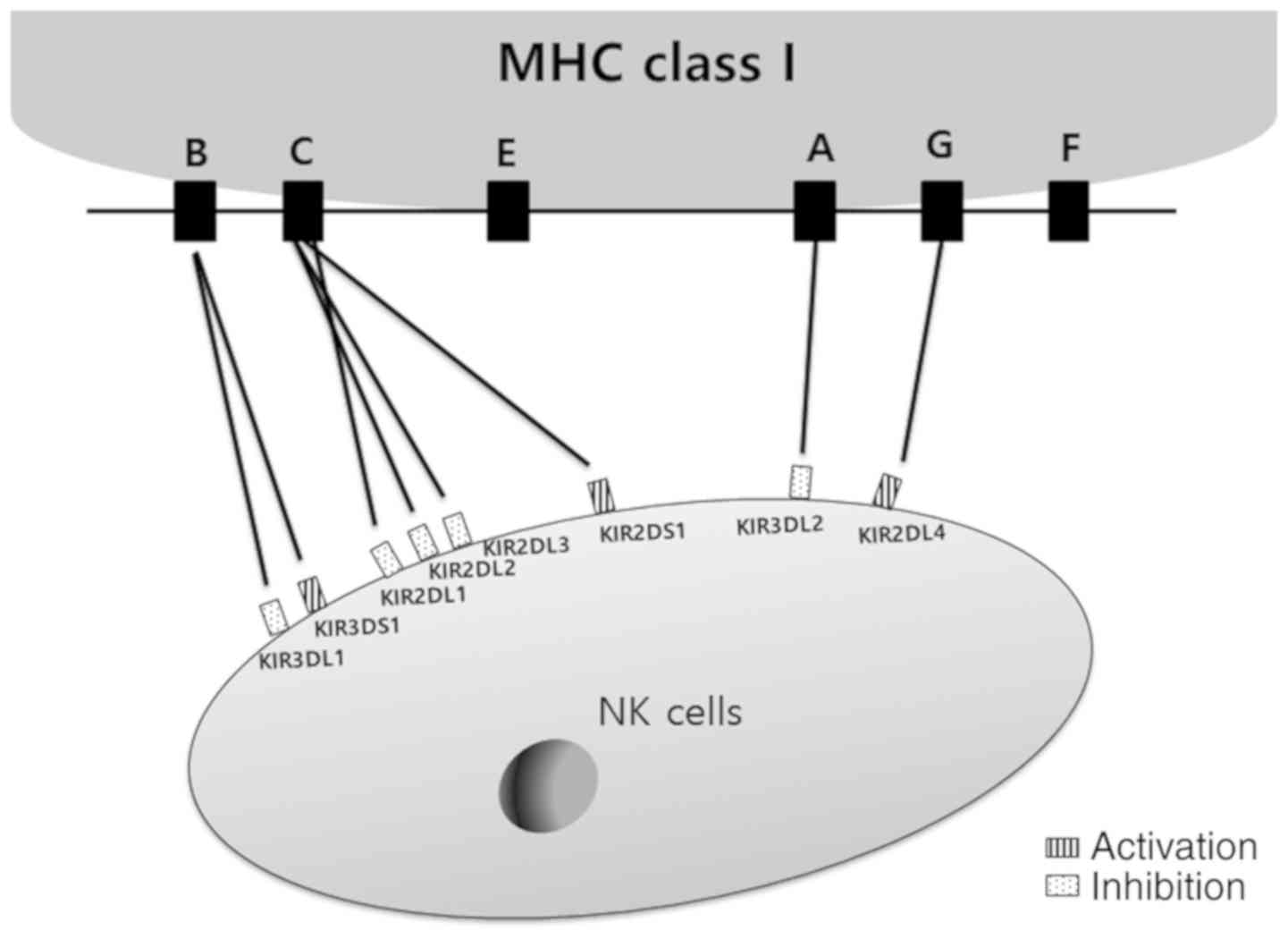
The alloreactivity of NK cells relies on the expression of a KIR
and its MHC-I ligand in the donor, and the absence of the MHC-I
ligand in the recipient. KIR and MHC-I ligand matching displays are
summarized in Fig. 5. There are two
principal donor-recipient mismatch models. The first mismatch model
comprises donor and recipient KIR ligands (ligand-ligand model),
and the second model includes recipients having no expression of
KIR ligands for donor-inhibitory KIR receptors (missing self-model)
(70). In alloreactive NK-cell
responses, KIR2DS1 expression offers a striking advantage since it
enables the efficient killing of C2/C2 or C1/C2 myelomonocytic
dendritic cells and T-cell blasts. The interaction between KIR3DS1
and human leukocyte antigen (HLA)-Bw4 leads to a prognosis of slow
disease progression following autologous stem cell transplantation
(SCT) in patients with multiple myeloma (71–74).
Thus, KIR2DS1 and KIR3DS1 expression in donor NK cells considerably
increases the efficacy of the alloreactive NK-cell subset. A
previous study reported a higher risk of developing CML due to the
prevalence of tolerogenic NK cells, resulting from an increase in
inhibitory signals generated by binding of iKIRs to ligands in the
homozygous state. Other studies have reported a notable response to
first or second generation TKI therapy to increase NK cell
activity, mediated at least in part by aKIRs or a mismatch between
iKIRs and their respective ligands (74,75).
The reason why the presence of active NK cells is paradoxically
associated with a disadvantageous therapeutic response in CML
remain poorly understood. One study undertook to further
investigate the role of KIR receptors and their ligands at the gene
level of a homogeneous group of CML patients and controls (76,77).
It was reported that KIR-HLA mismatches had a negative impact on
the disease course of CML, and that the KIR-ligand combination
KIR2DS2/KIR2DL2-absent/HLA-C1-present was markedly reduced in
patients with CML (76,77).
KIR genes in patients with leukemia
Another study analyzed the KIR genotype in patients
with hematological diseases (35).
Sequence specific primer-polymerase chain reaction (PCR-SSP)
analysis was performed for the amplification of six inhibitory
KIR genes (KIR2DL1-2DL4 and 3DL1-3DL2) and six activating
KIR genes (KIR2DS1-S5 and 3DS1). The PCR-SSP analysis
determined the KIR genotypes of 54 patients with leukemia,
including patients with AML, ALL, CML myelodysplastic syndrome and
acute myeloid-lymphoblast leukemia. The results of the present
study demonstrated that the frequency of activation of the
KIR genes 2DS1, 3DS1 and 2DS3 was increased in standard-risk
patients with acute AML compared with high-risk patients with acute
AML, but there was no association with CML (35).
Conclusions
This review has determined that understanding of NK
cell functions and signals may improve the treatment of tumors,
particularly in CML. Previous and current CML therapeutic
strategies were summarized, from TKI treatment to
immunotherapy.
Future clinical trials ought to consider that
matching or mismatching of KIRs and KIR-ligands (HLAs) may be
important for the treatment of CML using immunotherapy. Further
research is required to investigate the cellular functions of
specific KIR genes in patients with CML, and these studies
may be applied to the improvement of therapeutic responses in
CML.
Acknowledgements
The authors would like to thank the members of
Professor Kwang-Hyun Baek's laboratory for their critical comments
on the manuscript.
Funding
This study was supported by a grant from the Brain
Korea 21 (BK21) PLUS project in Korea (grant no.
22A20130012771).
Availability of data and materials
Not applicable.
Authors' contributions
HRL and KHB performed the literature search and
wrote the paper. KHB approved the final version of the manuscript
to be published. All authors read and approved the manuscript and
agree to be accountable for all aspects of the research in ensuring
that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Handgretinger R, Lang P and André MC:
Exploitation of natural killer cells for the treatment of acute
leukemia. Blood. 127:3341–3349. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Moretta L: Dissecting CD56dim
human NK cells. Blood. 116:3689–3691. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kannan GS, Aquino-Lopez A and Lee DA:
Natural killer cells in malignant hematology: A primer for the
non-immunologist. Blood Rev. 31:1–10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Farag SS, Fehniger TA, Ruggeri L, Velardi
A and Caligiuri MA: Natural killer cell receptors: New biology and
insights into the graft-versus-leukemia effect. Blood.
100:1935–1947. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sun PD: Structure and function of
natural-killer-cell receptors. Immunol Res. 27:539–548. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Willcox BE, Thomas LM and Bjorkman PJ:
Crystal structure of HLA-A2 bound to LIR-1, a host and viral major
histocompatibility complex receptor. Nat Immunol. 4:913–919. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Joyce MG, Tran P, Zhuravleva MA, Jaw J,
Colonna M and Sun PD: Crystal structure of human natural
cytotoxicity receptor NKp30 and identification of its ligand
binding site. Proc Natl Acad Sci USA. 108:6223–6228. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Suck G, Linn YC and Tonn T: Natural killer
cells for therapy of leukemia. Transfus Med Hemother. 43:89–95.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Parham P and Moffett A: Variable NK cell
receptors and their MHC class I ligands in immunity, reproduction
and human evolution. Nat Rev Immunol. 13:133–144. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Muntasel A, Ochoa MC, Cordeiro L,
Berraondo P, López-Díaz de Cerio A, Cabo M, López-Botet M and
Melero I: Targeting NK-cell checkpoints for cancer immunotherapy.
Curr Opin Immunol. 45:73–81. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lanier LL: Natural killer cell receptor
signaling. Curr Opin Immunol. 15:308–314. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sentman CL, Barber MA, Barber A and Zhang
T: NK cell receptors as tools in cancer immunotherapy. Adv Cancer
Res. 95:249–292. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vivier E, Nune JA and Vély F: Natural
killer cell signaling pathways. Science. 306:1517–1519. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Linnartz B, Wang Y and Neumann H:
Microglial immunoreceptor tyrosine-based activation and inhibition
motif signaling in neuroinflammation. Int J Alzheimers Dis.
2010:5874632010.PubMed/NCBI
|
|
15
|
Pugh JL, Nemat-Gorgani N, Norman PJ,
Guethlein LA and Parham P: Human NK cells downregulate Zap70 and
Syk in response to prolonged activation or DNA damage. J Immunol.
200:1146–1158. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Teresa Rios-Paredes: Use of NK cells in
haematological cancer therapy. Faculty of Medicine University of
Oslo. 2014.
|
|
17
|
Verneris MR and Miller JS: KIR B or not to
be?…that is the question for ALL. Blood. 124:2623–2624. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gaafar A, Sheereen A, Almohareb F, Eldali
A, Chaudhri N, Mohamed SY, Hanbali A, Shaheen M, Alfraih F, El
Fakih R, et al: Prognostic role of KIR genes and HLA-C after
hematopoietic stem cell transplantation in a patient cohort with
acute myeloid leukemia from a consanguineous community. Bone Marrow
Transplant. 58:1170–1179. 2018. View Article : Google Scholar
|
|
19
|
Long EO, Barber DF, Burshtyn DN, Faure M,
Peterson M, Rajagopalan S, Renard V, Sandusky M, Stebbins CC,
Wagtmann N, et al: Inhibition of natural killer cell activation
signals by killer cell immunoglobulin-like receptors (CD158).
Immunol Rev. 181:223–233. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ruggeri L, Capanni M, Urbani E, Perruccio
K, Shlomchik WD, Tosti A, Posati S, Rogaia D, Frassoni F, Aversa F,
et al: Effectiveness of donor natural killer cell alloreactivity in
mismatched hematopoietic transplants. Science. 295:2097–2100. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rajalingam R: Human diversity of killer
cell immunoglobulin-like receptors and disease. Korean J Hematol.
46:216–228. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fang F, Xiao W and Tian Z: NK cell-based
immunotherapy for cancer. Semin Immunol. 31:37–54. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ljunggren HG and Kärre K: In search of the
missing self-MHC molecules and NK cell recognition. Immunol Today.
11:237–244. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bottino C, Dondero A, Bellora F, Moretta
L, Locatelli F, Pistoia V, Moretta A and Castriconi R: Natural
killer cells and neuroblastoma: tumor recognition, escape
mechanisms, and possible novel immunotherapeutic approaches. Front
Immunol. 5:562014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Carotta S: Targeting NK cells for
anticancer immunotherapy: Clinical and preclinical approaches.
Front Immunol. 7:1522016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Martinet L and Smyth MJ: Balancing natural
killer cell activation through paired receptors. Nat Rev Immunol.
15:243–254. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shimasaki N, Coustan-Smith E, Kamiya T and
Campana D: Expanded and armed natural killer cells for cancer
treatment. Cytotherapy. 18:1422–1434. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu Y, Huang B and Shi J: Fas ligand and
lytic granule differentially control cytotoxic dynamics of natural
killer cell against cancer target. Oncotarget. 7:47163–47172.
2016.PubMed/NCBI
|
|
29
|
Childs RW and Carlsten M: Therapeutic
approaches to enhance natural killer cell cytotoxicity against
cancer: The force awakens. Nat Rev Drug Discov. 14:487–498. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nückel H, Switala M, Sellmann L, Horn PA,
Dürig J, Dührsen U, Küppers R, Grosse-Wilde H and Rebmann V: The
prognostic significance of soluble NKG2D ligands in B-cell chronic
lymphocytic leukemia. Leukemia. 24:1152–1159. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Burger JA and Gribben JG: The
microenvironment in chronic lymphocytic leukemia (CLL) and other B
cell malignancies: Insight into disease biology and new targeted
therapies. Semin Cancer Biol. 24:71–81. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Osman AE, AlJuryyan A, Alharthi H and
Almoshary M: Association between the killer cell
immunoglobulin-like receptor a haplotype and childhood acute
lymphoblastic leukemia. Hum Immunol. 78:510–514. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Boissel N, Rea D, Tieng V, Dulphy N, Brun
M, Cayuela JM, Rousselot P, Tamouza R, Le Bouteiller P, Mahon FX,
et al: BCR/ABL oncogene directly controls MHC class I chain-related
molecule A expression in chronic myelogenous leukemia. J Immunol.
176:5108–5116. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Maru Y: Molecular biology of chronic
myeloid leukemia. Cancer Sci. 103:1601–1610. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
https://www.ncbi.nlm.nih.gov/books/NBK65916/
|
|
36
|
Danier AC, de Melo RP, Napimoga MH and
Laguna-Abreu MT: The role of natural killer cells in chronic
myeloid leukemia. Rev Bras Hematol Hemoter. 33:216–220. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Huang CH, Liao YJ, Fan TH, Chiou TJ, Lin
YH and Twu YC: A Developed NK-92MI cell line with
Siglec-7neg phenotype exhibits high and sustainable
cytotoxicity against leukemia cells. Int J Mol Sci. 19:E10732018.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
O'Brien SG, Guilhot F, Larson RA, Gathmann
I, Baccarani M, Cervantes F, Cornelissen JJ, Fischer T, Hochhaus A,
Hughes T, et al: Imatinib compared with interferon and low-dose
cytarabine for newly diagnosed chronic-phase chronic myeloid
leukemia. N Engl J Med. 348:994–1004. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rossignol A, Levescot A, Jacomet F, Robin
A, Basbous S, Giraud C, Roy L, Guilhot F, Turhan AG, Barra A, et
al: Evidence for BCR-ABL-dependent dysfunctions of iNKT cells from
chronic myeloid leukemia patients. Eur J Immunol. 42:1870–1875.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Heaney NB and Holyoake TL: Therapeutic
targets in chronic myeloid leukaemia. Hematol Oncol. 25:66–75.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pophali PA and Patnaik MM: The role of new
tyrosine kinase inhibitors in chronic myeloid leukemia. Cancer J.
22:40–50. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Shapira T, Pereg D and Lishner M: How I
treat acute and chronic leukemia in pregnancy. Blood Rev.
22:247–259. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Goldman JM: How I treat chronic myeloid
leukemia in the imatinib era. Blood. 110:2828–2837. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Garderet L, Santacruz R, Barbu V, van den
Akker J, Carbonne B and Gorin NC: Two successful pregnancies in
chronic myeloid leukemia patient treated with imatinib.
Haematologica. 92:e9–e10. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Roth MS and Foon KA: Alpha interferon in
the treatment of hematologic malignancies. Am J Med. 81:871–882.
1986. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Giallongo C, La Cava P, Tibullo D,
Parrinello N, Barbagallo I, Del Fabro V, Stagno F, Conticello C,
Romano A, Chiarenza A, et al: Imatinib increases cytotoxicity of
melphalan and their combination allows an efficient killing of
chronic myeloid leukemia cells. Eur J Haematol. 86:216–225. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Millicovsky G, DeSesso JM, Kleinman LI and
Clark KE: Effects of hydroxyurea on hemodynamics of pregnant
rabbits: A maternally mediated mechanism of embryotoxicity. Am J
Obstet Gynecol. 140:747–752. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Thauvin-Robinet C, Maingueneau C, Robert
E, Elefant E, Guy H, Caillot D, Casasnovas RO, Douvier S and
Nivelon-Chevallier A: Exposure to hydroxyurea during pregnancy: A
case series. Leukemia. 15:1309–1311. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yellu M, Pinkard S, Ghose A and Medlin S:
CML in pregnancy: A case report using leukapheresis and literature
review. Transfus Apher Sci. 53:289–292. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ali R, Ozkalemkaş F, Ozkocaman V, Ozçelik
T, Ozan U, Kimya Y and Tunali A: Successful pregnancy and delivery
in a patient with chronic myelogenous leukemia (CML), and
management of CML with leukapheresis during pregnancy: A case
report and review of the literature. Jpn J Clin Oncol. 34:215–217.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Cayssials E and Guilhot F: Chronic myeloid
leukemia: Immunobiology and novel immunotherapeutic approaches.
BioDrugs. 31:143–149. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Mahoney KM, Rennert PD and Freeman GJ:
Combination cancer immunotherapy and new immunomodulatory targets.
Nat Rev Drug Discov. 14:561–584. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Mumprecht S, Schürch C, Schwaller J,
Solenthaler M and Ochsenbein AF: Programmed death 1 signaling on
chronic myeloid leukemia-specific T cells results in T-cell
exhaustion and disease progression. Blood. 114:1528–1536. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhang B, Chu S, Agarwal P, Campbell VL,
Hopcroft L, Jørgensen HG, Lin A, Gaal K, Holyoake TL and Bhatia R:
Inhibition of interleukin-1 signaling enhances elimination of
tyrosine kinase inhibitor-treated CML stem cells. Blood.
128:2671–2682. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ågerstam H, Hansen N, von Palffy S, Sandén
C, Reckzeh K, Karlsson C, Lilljebjörn H, Landberg N, Askmyr M,
Högberg C, et al: IL1RAP antibodies block IL-1-induced expansion of
candidate CML stem cells and mediate cell killing in xenograft
models. Blood. 128:2683–2693. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Arranz L, Arriero MD and Villatoro A:
Interleukin-1β as emerging therapeutic target in hematological
malignancies and potentially in their complications. Blood Rev.
31:306–317. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhao K, Yuan S, Yin L, Xia J and Xu K:
Potential efficacy of human IL-1RAP specific CAR-T cell in
eliminating leukemic stem cells of chronic myeloid leukemia. J
Leukemia. 5:2322017. View Article : Google Scholar
|
|
58
|
Stramucci L and Perrotti D: Twisting IL-1
signaling to kill CML stem cells. Blood. 128:2592–2593. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Tarafdar A, Hopcroft LE, Gallipoli P,
Pellicano F, Cassels J, Hair A, Korfi K, Jørgensen HG, Vetrie D,
Holyoake TL, et al: CML cells actively evade host immune
surveillance through cytokine-mediated downregulation of MHCII
expression. Blood. 129:199–208. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Levescot A, Flamant S, Basbous S, Jacomet
F, Féraud O, Anne Bourgeois E, Bonnet ML, Giraud C, Roy L, Barra A,
et al: BCR-ABL-induced deregulation of the IL-33/ST2 pathway in
CD34+ progenitors from chronic myeloid leukemia
patients. Cancer Res. 74:2669–2676. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Rocca S, Carrà G, Poggio P, Morotti A and
Brancaccio M: Targeting few to help hundreds: JAK, MAPK and ROCK
pathways as druggable targets in atypical chronic myeloid leukemia.
Mol Cancer. 17:402018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Pierson BA and Miller JS: The role of
autologous natural killer cells in chronic myelogenous leukemia.
Leuk Lymphoma. 27:387–399. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Verfaillie C, Kay N, Miller W and McGlave
P: Diminished A-LAK cytotoxicity and proliferation accompany
disease progression in chronic myelogenous leukemia. Blood.
76:401–408. 1990.PubMed/NCBI
|
|
64
|
Pawelec G, Schneider E, Ehninger G,
Rehbein A and Schmidt H: Partial correction of defective generation
of lymphokine-activated killer cells in patients with chronic
myelogenous leukaemia after in vivo treatment with interferon-alpha
(Wellferon). Cancer Immunol Immunother. 29:63–66. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Mustjoki S, Ekblom M, Arstila TP, Dybedal
I, Epling-Burnette PK, Guilhot F, Hjorth-Hansen H, Höglund M,
Kovanen P, Laurinolli T, et al: Clonal expansion of T/NK-cells
during tyrosine kinase inhibitor dasatinib therapy. Leukemia.
23:1398–1405. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kijima M, Gardiol N and Held W: Natural
killer cell mediated missing-self recognition can protect mice from
primary chronic myeloid leukemia in vivo. PLoS One. 6:e276392011.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zhao XY, Chang YJ, Xu LP, Zhang XH, Liu
KY, Li D and Huang XJ: HLA and KIR genotyping correlates with
relapse after T-cell-replete haploidentical transplantation in
chronic myeloid leukaemia patients. Br J Cancer. 111:1080–1088.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Necchi A, Lanza F, Rosti G, Martino M,
Farè E, Pedrazzoli P, European Society for Blood, Marrow
Transplantation and Solid Tumors Working Party; (EBMT-STWP) the
Italian Germ Cell Cancer Group (IGG), : High-dose chemotherapy for
germ cell tumors: Do we have a model? Expert Opin Biol Ther.
15:33–44. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Zahid U, Akbar F, Amaraneni A, Husnain M,
Chan O, Riaz IB, McBride A, Iftikhar A and Anwer F: A review of
autologous stem cell transplantation in lymphoma. Curr Hematol
Malig Rep. 12:217–226. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Kim S, Poursine-Laurent J, Truscott SM,
Lybarger L, Song YJ, Yang L, French AR, Sunwoo JB, Lemieux S,
Hansen TH, et al: Licensing of natural killer cells by host major
histocompatibility complex class I molecules. Nature. 436:709–713.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Sivori S, Carlomagno S, Falco M, Romeo E,
Moretta L and Moretta A: Natural killer cells expressing the
KIR2DS1-activating receptor efficiently kill T-cell blasts and
dendritic cells: Implications in haploidentical HSCT. Blood.
117:4284–4292. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Gabriel IH, Sergeant R, Szydlo R, Apperley
JF, DeLavallade H, Alsuliman A, Khoder A, Marin D, Kanfer E, Cooper
N, et al: Interaction between KIR3DS1 and HLA-Bw4
predicts for progression-free survival after autologous stem cell
transplantation in patients with multiple myeloma. Blood.
116:2033–2039. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Middleton D, Diler AS, Meenagh A, Sleator
C and Gourraud PA: Killer immunoglobulin-like receptors (KIR2DL2
and/or KIR2DS2) in presence of their ligand (HLA-C1 group) protect
against chronic myeloid leukaemia. Tissue Antigens. 73:553–560.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Marin D, Gabriel IH, Ahmad S, Foroni L, de
Lavallade H, Clark R, O'Brien S, Sergeant R, Hedgley C, Milojkovic
D, et al: KIR2DS1 genotype predicts for complete cytogenetic
response and survival in newly diagnosed chronic myeloid leukemia
patients treated with imatinib. Leukemia. 26:296–302. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Kreutzman A, Jaatinen T, Greco D, Vakkila
E, Richter J, Ekblom M, Hjorth-Hansen H, Stenke L, Melo T, Paquette
R, et al: Killer-cell immunoglobulin-like receptor gene profile
predicts good molecular response to dasatinib therapy in chronic
myeloid leukemia. Exp Hematol. 40:906–913. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
La Nasa G, Caocci G, Littera R, Atzeni S,
Vacca A, Mulas O, Langiu M, Greco M, Orrù S, Orrù N, et al:
Homozygosity for killer immunoglobin-like receptor haplotype A
predicts complete molecular response to treatment with tyrosine
kinase inhibitors in chronic myeloid leukemia patients. Exp
Hematol. 41:424–431. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Zhao XY, Chang YJ and Huang XJ:
Differential expression levels of killer immunoglobin-like receptor
genotype in patients with hematological malignancies between
high-risk and standard-risk groups. Zhongguo Shi Yan Xue Ye Xue Za
Zhi. 16:746–749. 2008.(In Chinese). PubMed/NCBI
|