Introduction
Serratia marcescens can be isolated from
soil, water, plants and air but rarely from insects (1). Periplaneta americana (a type of
American cockroach) is one of the most common household pests,
found around drainage basins, sewage ducts, garbage and wall slots.
However in China, it is a traditional Chinese medicine used in
anti-inflammatory, antiviral, antioxidant and anticancer treatments
(2). It has been reported that the
extractive Periplaneta americana exerts these effects on
cancer cell lines such as human cervical cancer cells (HeLa), human
hepatoma cells (Bel-7402) and human lung cancer cells (H125)
(3,4). Microorganisms in the gut of insects
interact with each other, as well as their hosts and their
environment. These interactions are thought to have the capacity to
degrade the protein and protect the host against diseases through
producing secondary metabolites. Serratia marcescens, a
Gram-negative bacterium, has been reported to produce a large
quantity of prodigiosin (5,6). Prodigiosin (2-methyl-3-pentyl-6-
methoxyprodiginine) is a red colored secondary metabolite that
belongs to a class of tripyrrole compounds. Prodigiosin has also
shown effective antimalarial, antifungal and immunosuppressive
activities, as well as anticancer activities (7–9).
In the present study, we successfully isolated a
strain of Serratia marcescens named WA12-1-18 from the gut
of Periplaneta americana, which was capable of producing
high levels of pigment reaching 2.77 g/l via solid fermentation,
which was identified as prodigiosin by ultraviolet (UV), high
performance liquid chromatography (HPLC), Fourier-transform
infrared spectroscopy (FTIR), LC-mass spectroscopy (LC-MS) and
nuclear magnetic resonance (NMR). The aim of the present study was
to analyze the antitumor effect of prodigiosin and investigate the
mechanisms underlying apoptosis in HeLa cells in vitro.
Materials and methods
Isolation of Serratia marcescens from
the gut of Periplaneta americana
All Periplaneta americana samples were
collected from the dormitories and the outdoor surroundings of
Guangdong Pharmaceutical University (Guangdong, China), which were
identified morphologically and molecularly. Periplaneta
americanas were fasted for 3 days and then sacrificed following
anesthesia with 100% CO2 for 2 h. They were then washed
with distilled water. Sterilization was conducted with 75% ethanol
and 3.5% sodium hypochlorite/water solution followed by 3 washes
with 1X sterile phosphate-buffered saline (PBS). The entire gut of
the Periplaneta americana was extracted from the abdomen and
then ground into a slurry. The bacterial fluid was diluted into
0.01, 0.005 and 0.00025 µg/ml by gradient, coated on the
agar-solidified Luria-Bertani (LB) medium (Amresco, LLC, Solon, OH,
USA), which contained of 75 µg/ml potassium dichromate, and then
incubated in the dark at 28°C for a week (10). The morphology and surface
characteristics of the bacterial colonies were examined using a
scanning electron microscope. Finally, the red-pigment-producing
bacterial strain was named, WA12-1-18.
Molecular identification of Serratia
marcescens
The strain WA12-1-18 was cultured in LB medium broth
for 2 days with shaking at 37°C and harvested by centrifugation
(12,000 × g at 20°C for 15 min). DNA was extracted and amplified by
polymerase chain reaction (PCR) using the genomic DNA as a template
and the bacterial universal primers, 27 forward
(5′-GAGTTTGATCACTGGCTCAG-3′) and 1492 reverse
(5′-TACGGCTACCTTGTTACGACTT-3′). The PCR mixture (25 µl) contained 1
µl of template DNA, 2.5 µl of 10X Taq DNA polymerase buffer, 1.5 µl
primers (each) and 18.5 µl ddH2O. The PCR mixture was
then incubated with the following thermocycling conditions: 94°C
for 3 min, followed by 35 cycles of 94°C for 0.5 min, 58°C for 0.5
min and 72°C for 1.5 min, followed by a final extension performed
at 72°C for 10 min (11). The
amplification products were purified using a DNA purification kit
(Qiagen, Inc. Valencia, CA, USA), and sequencing was performed by
Invitrogen (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Phylogenetic trees were then constructed from evolutionary
distances using the neighbor-joining method (12).
Extraction of prodigiosin
A single colony was inoculated in 10 ml of LB medium
and incubated for 12 h at 37°C, and then transferred into 200 ml LB
medium and incubated for a further 3 days at 37°C. Then, 250 µl
bacterial cultures were spread onto LB solid plates and incubated
in the dark at a low temperature (23°C) for 72 h (13). The lawn plates were harvested and
dissolved in ethanol and centrifuged at 10,000 × g for 20 min at
4°C. The upper layer was collected, dried and re-dissolved in
ethylacetate, the pigment was purified by silica gel column
chromatography (28×12 cm) with petroleum ether and
ethylacetate/n-hexane (40:1, v:v), as described previously
(14,15). At the end of these procedures, the
pigment fractions were pooled and dried.
Identification of prodigiosin
As described previously (16), prodigiosin was analyzed via HPLC at
the absorption wavelength 534 nm using Phenomenex Luna C-182
semipreparative column (250×10 mm; sample, 5 µl; Phenomenex,
Torrance, CA, USA) with the Bio-Rad HPLC system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The separation was
performed using water (solvent A) and acetonitrile/methanol (1:1;
solvent B) mobile phases, and a gradient elution program at 3
ml/min with the following parameters: 0–25 min 15–100% B (linear
gradient), 25–35 min 100% B and 35–40 min 15% B to re-equilibrate
the column. Fractions containing the targeted compounds were
combined and concentrated by solvent evaporation. UV absorption of
prodigiosin in methanol at pH 5 and pH 9 was scanned using a
spectrophotometer at a range of 200–800 nm. In the FTIR study, FTIR
spectra were made on a Nicolet Nexus 670 FT-IR infrared
spectrometer. All spectra were taken using the Attenuated Total
Reflection method (17). Briefly,
prodigiosin was mixed with potassium bromide in a clean glass
pestle and compressed to produce a pellet. Scanning was performed
following base line correction by setting a wave number range of
400–4,000/cm−1. In LC-MS analysis, the pigment was
dissolved in acetonitrile, and 10 µl was analyzed in API-4000 Q
trap LC-MS/MS. The operating conditions were as follows: 4,500
ionization voltage, 350°C ionization temperature, ionization mode
ESI positive, a MS C18 (50×4.6 mm) column was used, 90:10
(acetonitrile:water) mobile phase, and the injection volume was 10
µl with a flow rate of 0.5 ml/min; performed as described
previously (18). In addition,
prodigiosin dissolved in d-chloroform was analyzed by hydrogen
(1H)-NMR and carbon-13 (13C)-NMR to identify
the structure of the purified product. NMR characterization
(1H NMR or 13C NMR) was performed using the
pulse program zg30 with a spectral width of 8,278.146 Hz using
Bruker Advance 400 MHz NMR spectrophotometer (Bruker Corporation,
Ettlingen, Germany) with 5 mm probe in CDCl3
solvent.
Detection of the cell proliferation
assay of prodigiosin
The HeLa cell line (Experimental Animal Center of
Sun Yat-sen University, Guangzhou, China) was maintained in
Dulbecco's modified Eagle's medium. An MTT assay was used to
measure the levels of cell proliferation. A total of
5×104 cells were treated with different concentrations
(0, 0.5, 1.0 and 2.0 µg/ml) of prodigiosin for 24, 48 and 72 h at
37°C, followed by incubation in MTT (0.1 mg/ml) at 37°C for 4 h and
dissolution in dimethyl sulfoxide at room temperature for 10 min.
Microplate reader (at 570 nm) was used to measure the absorbance of
each well. Three independent experiments were performed (19–21).
DAPI staining and transmission
electron microscopy (TEM)
In order to view the nucleus, 5×104 HeLa
cells were fixed in methanol, incubated with DAPI for 10 min at
37°C (Nanjing KeyGen Biotech Co., Ltd., Nanjing, China; cat. no.
KGA215-10), and analyzed using a fluorescence microscope (22). HeLa cells (5×105) were
treated with different concentrations of prodigiosin (0, 0.5, 1.0
and 2.0 µg/ml) for 24 h in 6-well plate. The HeLa cells were then
harvested, washed with PBS and fixed with 2.5% glutaraldehyde at
4°C for 12 h. Following washing with PBS, HeLa cells were
dehydrated with ethanol and acetone, sequentially fixed with 1%
osmium tetroxide (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
for 2 h at 37°C, followed by embedding in epoxy resin for 12 h at
37°C. Ultra-thin slices (80 nm) were then produced, stained with
1.0% uranylacetate for 5 min at room temperature and viewed on a
transmission electron microscope (23,24).
Detection of apoptosis by flow
cytometry
Since the emission wavelength of prodigiosin was
similar to that of propidium iodide, the levels of apoptosis were
detected using Annexin V. HeLa cells (5×105) were
treated with prodigiosin in a concentration- (0, 0.5, 1.0 and 2.0
µg/ml) and time (24, 36 and 48 h)-dependent manner at 37°C.
Following the different exposure times, cells were washed with PBS
and resuspended in 150 ml of binding buffer, which contained 5 ml
of Annexin V-fluorescein isothiocyanate (Biolegend Inc., San Diego,
CA, USA; cat. no. 640912) and were incubated at room temperature
for 15 min. The cells were analyzed using a FACScan flow cytometer
and the associated software (Becton, Dickinson and Company,
Franklin Lakes, NJ, USA).
Detection of the apoptotic pathway
using western blotting and reverse transcription-quantitative PCR
(RT-qPCR)
Western blotting was performed as previously
described (25). The human
caspase-3 antibody (cat. no. 9662) was purchased from Bioworld
Technology, Inc. (St. Louis Park, MN, USA), and the human B-cell
lymphoma 2 (Bcl-2; cat. no. 2872), Bcl-2-associated X (Bax; cat.
no. 2774) antibodies and β-actin (cat. no. 8457; dilution of all
primary antibodies; 1:1,000) were purchased from Bosterbio
Biological Technology (Pleasanton, CA, USA). The membranes were
washed with Tris-buffered saline with 0.1% Tween-20 (TBST) 3 times
and incubated with secondary horseradish peroxidase-conjugated
antibodies (Anti-rabbit Immunoglobulin G; cat. no. 7074; Thermo
Fisher Scientific, Inc.; dilution, 1:5,000) at 25°C for 2 h.
Membranes were then washed with TBST 3 more times and immune
reactive protein bands were detected using an enhanced
chemiluminescence kit (Beijing Transgen Biotech Co., Ltd., Beijing,
China). Image Quant TL 7.0 software (GE Healthcare, Chicago, IL,
USA) was employed to quantify protein expression levels. For
RT-qPCR, following the supplier's protocol, TRIzol™ reagent
(Promega Corporation, Madison, WI, USA) was used to extract the
total RNA of HeLa cells that were treated with different
concentrations (0, 0.5, 1.0 and 2.0 µg/ml) of prodigiosin, as
aforementioned. RNA was quantitated by optical density measurement
using a spectrophotometer. The RT-for-PCR Kit (Promega Corporation)
was used for RT-PCR. Briefly, 1 µg total RNA with 1 µl 20 µM
oligo(dT)18 primers was heated at 70°C for 2 min, then quenched
rapidly on ice. The RT reaction was followed by the addition of 4
µl 5X Reaction buffer (250 mM Tris-HCl, pH 8.3, 375 mM KCl, 15 mM
MgCl2), 1 µl dNTP mixture (10 mM each dATP, dGTP, dCTP
and dTTP), 20 U recombinant RNase inhibitor and 200 U MMLV reverse
transcriptase in a 20 µl reaction volume. Samples were incubated at
42°C for 1 h in an air incubator, followed by inactivation of the
reverse transcriptase at 95°C for 5 min. Then, 80 µl nuclease-free
water was added to a final volume of 100 µl. RT-qPCR was performed
using primer pairs for Bcl-2, Bax and GAPDH (Table I) with the SYBR-Green PCR Master mix
(Promega Corporation) with the following thermocycling conditions:
1 cycle at 94°C for 4 min, followed by 20 cycles at 94°C for 30 sec
and 68°C for 1.5 min, and finally 1 cycle at 68°C for 5 min. The
ABI PRISM 7000 sequence detection system (Applied Biosystems;
Thermo Fisher Scientific, Inc.) and the 2−∆∆Cq (26) method were used for quantitative
analysis.
 | Table I.List of oligonucleotides used in the
present study. |
Table I.
List of oligonucleotides used in the
present study.
| Primer name | Primer sequence
(5′-3′) |
|---|
| Bcl-2F |
CTGTGGATGACTGAGTACC |
| Bcl-2R |
CAGCCAGGAGAAATCAAAC |
| Bax-F |
CTGACATGTTTTCTGACGGCAA |
| Bax-R |
GAAGTCCAATGTCCAGCCCA |
| GAPDH-F |
CTCTGCTCCTGTTCGAC |
| GAPDH-R |
ACGACCAAATCCGTTGACTC |
Statistical analysis
SPSS 19.0 statistical software (IBM Corp., Armonk,
MA, USA) was used to evaluate the results. One-way analysis of
variance was used for comparisons between multiple groups. For post
hoc analyses, heterogeneity of variance was accepted if P>0.05,
and the Least Significant Difference method was used. Otherwise,
Dunnett's T3 method was used. Homogeneity of variance of the
samples to be compared was tested using a Levene's test. The
results were expressed as the mean ± standard deviation. P<0.05
was considered to indicate a statistically significant
difference.
Results
Identification of Serratia marcescens
in the gut of Periplaneta americana
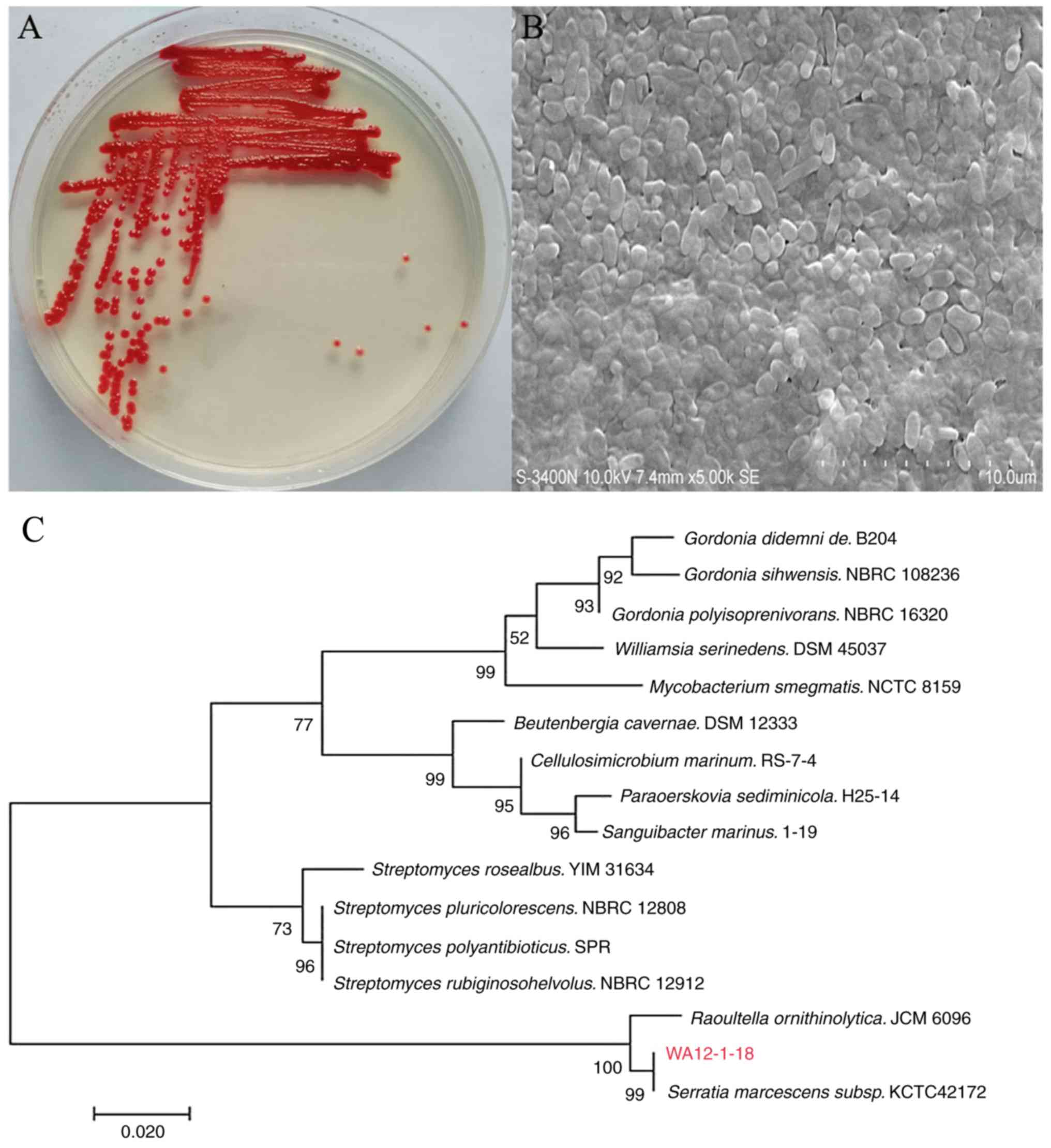
Observation of morphological characteristics
revealed that the colony of strain surface was smooth, convex and
shiny with a vivid red color (Fig.
1A). A straight pole shape with blunt ends and uneven surface
waves were observed via micromorphological analysis (Fig. 1B). The 16S r DNA sequences of the
red colony (comprising 1,562 nucleotides) were obtained and
submitted to EzBioCloud (www.ezbiocloud.net/identify). The sequence exhibited
the highest similarity (99.59%) to that of Serratia
marcescens. A phylogenetic tree was constructed based on the
16S r DNA PCR products as well as on the coding gene sequences of
the isolate and its nearest relatives (Fig. 1C). The strain was identified as
Serratia marcescens and named WA12-1-18.
Extraction and identification of
prodigiosin
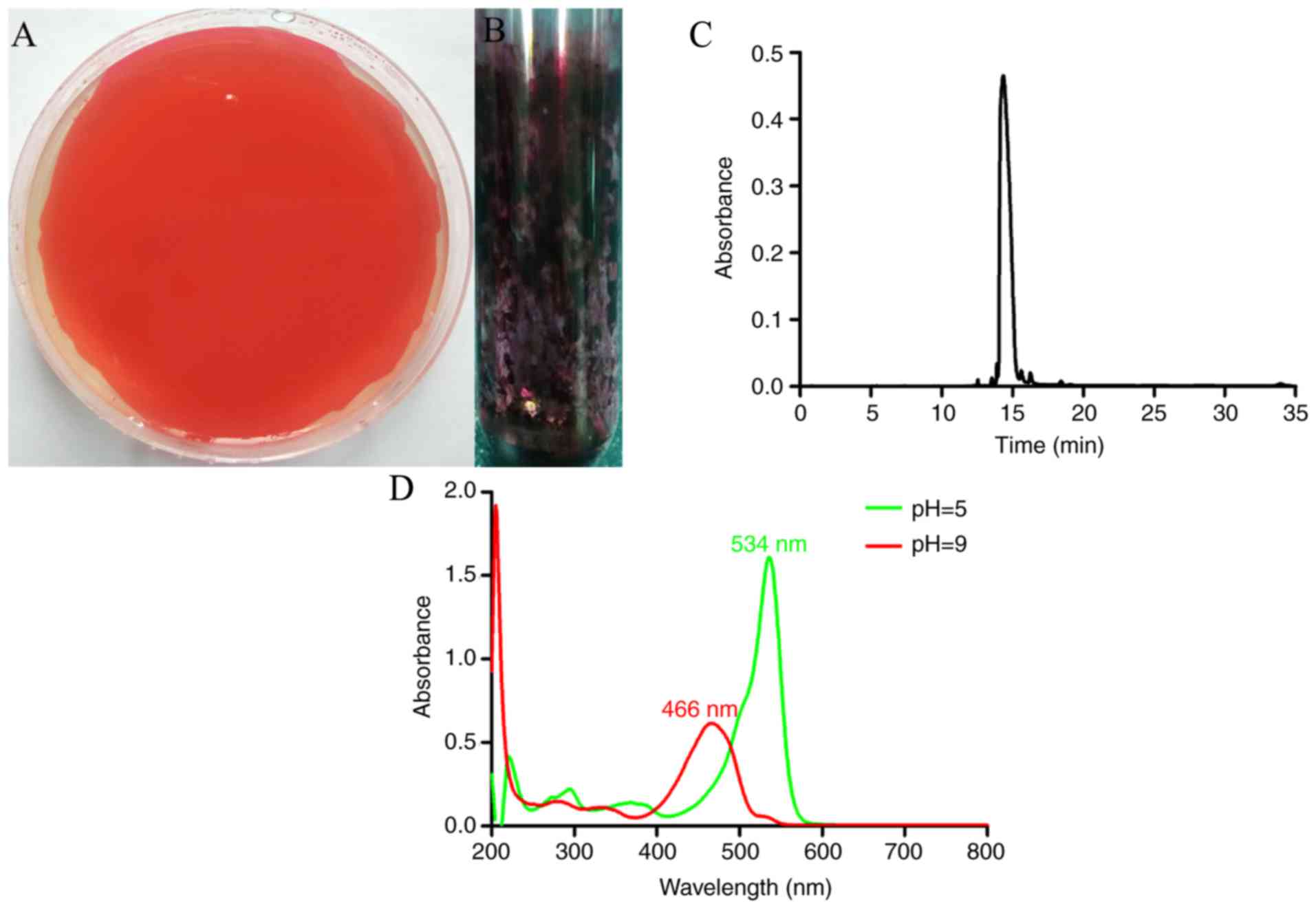
The red coating of the solid medium was collected
following culture for 72 h at 23°C. The solubility test
demonstrated that the dark-red powder had good solubility in polar
solvent (MeOH and EtOAc) but rarely dissolved in water (Fig. 2A and B). The yield of prodigiosin
was high, reaching 2.77 g/l, and the purity of this secondary
metabolite produced by Serratia marcescens was 98.25%
determined by HPLC (Fig. 2C). UV
spectrophotometry revealed that the maximum absorptions were at 534
nm (pH=5.0) and 466 nm (pH=9.0; Fig.
2D). The FTIR peak assignments showed that the characteristic
wave numbers (cm−1) were at 3,450 (N-H str aromatic),
2,924 and 2,853 (C-H2 str aliphatic), 1,729 (C-O str
ether), and 1,631 and 1,581 (C=C, C=N str aromatic; Fig. 2E). The molecular mass determined by
LC-MS was 324.207(m/z 324.207; Fig.
2F). The chemical shifts of prodigiosin were further confirmed
by NMR spectroscopy (Fig. 2G and
H): 1H-NMR (CDCl3, 300 MHz, ppm) δ 0.98 (3H, t,
H11″), 1.29 (2H, m,H9″), 1.45 (2H, m, H10″), 1.59 (2H, m, H8″),
2.38 (2H, t, H7″), 2.47 (3H, s, H6″), 3.31 (1H,m, H3), 4.29 (1H, d,
H3′), 4.88 (3H, s, OCH3), 5.10 (1H, brd, H3″), 5.34 (1H, m, H4),
6.37 (1H, brs, H6′),7.18 (1H, m, H2), 7.62 (1H, brs, H1), 7.71 (1H,
brs, H1′); 13C-NMR (CDCl3, 75 MHz, ppm) δ 12.03 (C6″),
14.59 (C11″), 23.78(C10″), 23.90 (C7″), 26.79 (C8″), 30.94 (C9″),
37.84 (C3′), 59.57 (OCH3), 66.83 (C3), 95.46 (C6′), 108.79 (C4),
112.46 (C5′), 117.04 (C5), 123.67 (C2″), 128.35 (C2), 130.03 (C3″),
131.02(C4″), 132.50(C5″), 154.30 (C2′), and 169.22 (C4′). All
characteristics corresponded to those of prodigiosin
(C20H25N3O).
Detection of the cell proliferation
assay of prodigiosin
Cell growth of prodigiosin-treated HeLa cells was
measured by the MTT assay at different time points over 72 h of the
treatment period. The results shown in Table II demonstrated a time- and
dose-dependent decline in number of viable cells within 72 h of
prodigiosin-treatment. Using the dose-response curves, the
half-maximal inhibitory concentration (IC50) values of
prodigiosin in HeLa were 2.1 µg/ml at 24 h, 1.2 µg/ml at 48 h and
0.5 µg/ml at 72 h, respectively.
| Table II.Inhibition effect of prodigiosin on
HeLa cells by MTT assay. |
Table II.
Inhibition effect of prodigiosin on
HeLa cells by MTT assay.
|
| 24 h | 48 h | 72 h |
|---|
|
|
|
|
|
|---|
| Groups | Inhibition rate
(%) | P-value | Inhibition rate
(%) | P-value | Inhibition rate
(%) | P-value |
|---|
| Control | 0 | – | 0 | – | 0 | – |
| 0.5 µg/ml |
14.3±2.7a | 0.040 |
25.5±1.2a | 0.007 |
41.5±1.2b | 0.005 |
| 1.0 µg/ml |
26.3±3.1a | 0.039 |
40.7±2.3b | 0.006 |
74.7±2.3b | 0.003 |
| 2.0 µg/m |
50.3±1.6b | 0.006 |
63.5±1.0c | <0.0001 |
86.5±1.0c | 0.002 |
| 4.0 µg/ml |
57.8±2.9b | 0.026 |
84.4±0.1c | <0.0001 |
90.4±1.1c | <0.0001 |
| 8.0 µg/ml |
88.1±2.3c | <0.0001 |
92.3±2.2c | <0.0001 |
95.3±2.2c | <0.0001 |
|
IC50 | 2.1 µg/ml |
| 1.2 µg/ml |
| 0.5 µg/ml |
|
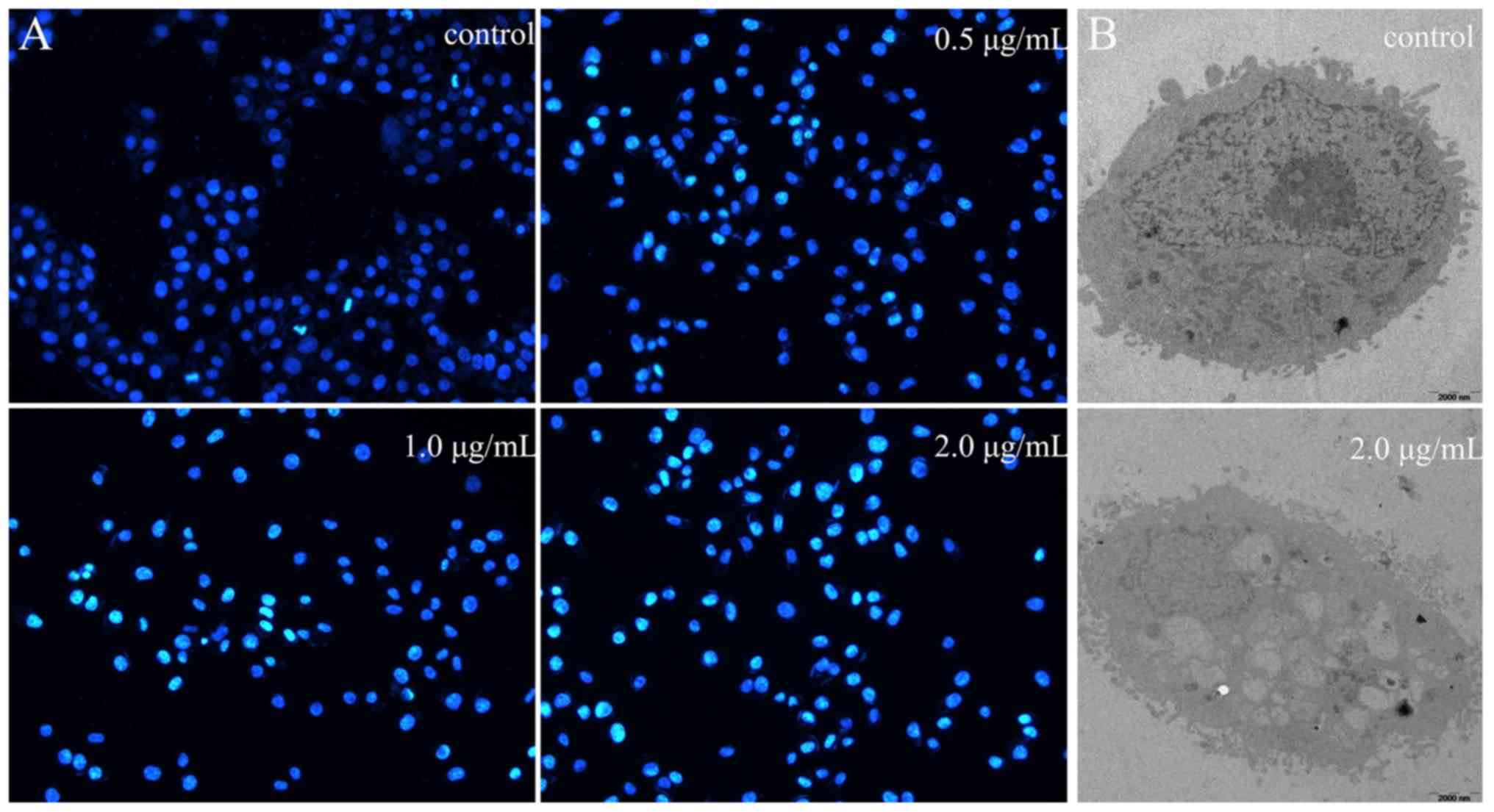
DAPI staining and TEM
The results of the DAPI staining assay revealed that
the number of apoptotic cells increased with the prodigiosin
concentration following 24 h (Fig.
3A). The morphology of control HeLa cells at 0 µg/ml
prodigiosin was used as the normal morphology (Fig. 3B). TEM revealed rich cell surface
microvilli, normal nucleus and abundant cytoplasm; the endoplasmic
reticulum can also be seen clearly. However, apoptosis was induced
when HeLa cells were treated with 2 µg/ml prodigiosin (Fig. 3B), presenting no cell surface
microvilli, folded nuclear membrane, increased nuclear
heterochromatin and condensed chromatin. These phenomena suggested
that the apoptotic process was induced by prodigiosin
treatment.
Detection of apoptosis using flow
cytometry
To gain further understanding of the mechanism
underlying the inhibition of proliferation, the present study
investigated the effect on apoptosis by flow cytometry. HeLa cells
were treated with 0, 0.5, 1.0 and 2.0 µg/ml of prodigiosin for 24,
36 and 48 h, respectively. Annexin V stained cells were tested by
flow cytometry for quantitation. There were Annexin V-negative and
Annexin V-positive on the dot-plots, as well as the stages of
apoptosis located on the upper quadrant of the dot-plot (data not
shown). The results shown in Table
III demonstrated that there were dose- and time-associated
increases in the apoptosis of HeLa cells following prodigiosin
treatment.
| Table III.Effect of prodigiosin on the
apoptosis of HeLa cell as determined by flow cytometry. |
Table III.
Effect of prodigiosin on the
apoptosis of HeLa cell as determined by flow cytometry.
| Groups | 24 h Apoptosis rate
(%) | P-value | 36 h Apoptosis rate
(%) | P-value | 48 h Apoptosis rate
(%) | P-value |
|---|
| Control | 1.2±0.5 | – | 2.8±1.1 | – | 2.6±0.6 | – |
| 0.5 µg/ml | 3.8±0.7 | 0.068 |
8.5±1.2a | 0.043 |
19.7±1.4b | 0.006 |
| 1.0 µg/ml |
6.2±1.2a | 0.046 |
10.9±1.9b | 0.008 |
23.7±2.4b | 0.003 |
| 2.0 µg/m |
7.6±1.1a | 0.039 |
14.2±1.8b | 0.007 |
26.2±2.3b | 0.001 |
Detection of the apoptotic pathway
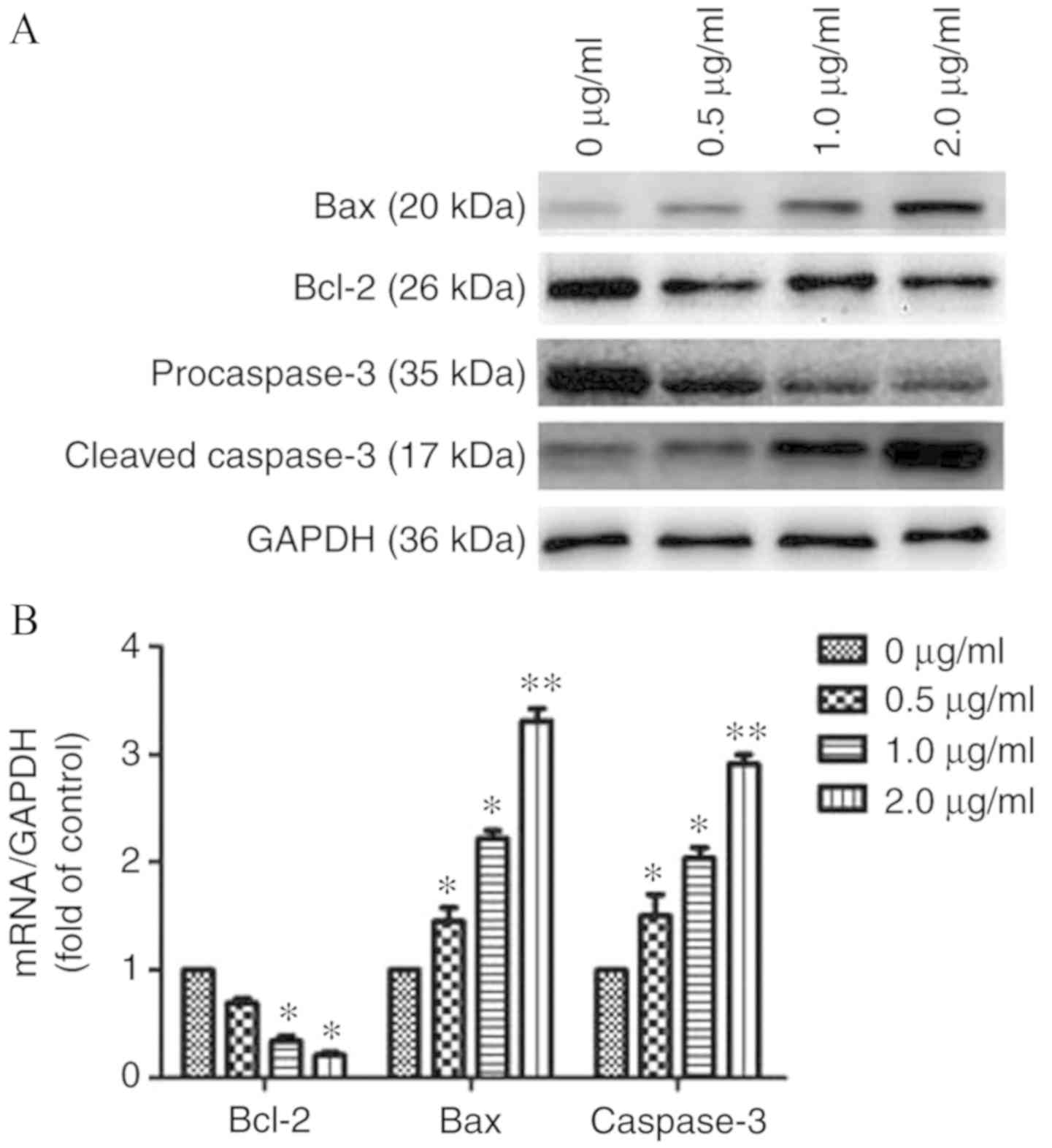
using western blotting and RT-qPCR
To further elucidate the mechanism of apoptosis
induced by prodigiosin, the present study evaluated the protein and
mRNA levels of key regulators via western blotting and RT-qPCR. The
results demonstrated that prodigiosin treatment could downregulate
Bcl-2 levels, and concomitantly upregulate Bax and caspase-3 levels
when compared with the control (0 µg/ml; Fig. 4A and B).
Discussion
The intestinal microbiota of insects has become a
key area of interest in recent years (27). Periplaneta americanas are
insects of the order Blattodea, some of which are associated
with the human environment and carrier numerous microorganisms
(28). There are few studies
focusing on the endophytic bacteria in Periplaneta
americana, especially Serratia marcescens. The microbial
community in the gut is responsible for digestion, but how they
participate and carry out this activity is unknown. The results of
the present study therefore confirm the idea that the
Periplaneta americana gut may be a specialized microhabitat
of enhanced microbial activities (29). In the present study, the 16S r DNA
gene based phylogenetic tree analysis showed the highest similarity
(99.59%), confirming that Serratia marcescens was
successfully isolated from the gut of Periplaneta americana.
The authors hope that more novel microorganisms can be isolated in
the group's future research. The Gram-negative bacterium
Serratia marcescens is known for its ability to produce
numerous secondary metabolites (30). Prodigiosin is one of the bioactivity
secondary metabolites produced by Serratia marcescens. Low
productivity of prodigiosin was reported under liquid fermentation
(7). Therefore, in the present
study, prodigiosin was obtained by solid fermentation. Compared
with submerged fermentation, solid fermentation is simpler and more
efficient in terms of the preparation of prodigiosin (31). Furthermore, the yield of prodigiosin
from the gut of the cockroaches was 2.77 g/l, which was higher than
that obtained from the body of a grasshopper and the common
Serratia marcescens (32,33).
It is possible that the intestinal bacteria may have the capacity
to protect the host against diseases by producing secondary
metabolites. The purity of prodigiosin was as high as 98.25%, as
determined by HPLC, which was consistent with a previous study
(34).
The structure of prodigiosin is similar to that of
obatoclax, which is a Bcl-2 inhibitor (35). In the present study, prodigiosin
treatment in a HeLa cell line was investigated and the results
revealed that prodigiosin inhibited the proliferation of HeLa cells
in a time- and dose-dependent manner, and the IC50
values were 0.5–2.1 µg/ml over 24, 48 and 72 h. In addition,
another study showed that the A549 cell line was very sensitive to
the prodigiosin analogue, with an IC50 value of ~2.2
mg/l (36). The results of the DAPI
staining assay, flow cytometry and TEM demonstrated that
prodigiosin levels increased in the apoptotic cell population in a
dose- and time-dependent manner. The possible anticancer mechanisms
of prodigiosin reviewed by Sruthy et al (29) is attributed as pH modulators, cell
cycle inhibitors, DNA cleavage agents and mitogen activated protein
kinase regulators. The present results suggested that
prodigiosin-induced apoptosis may be due to the down regulation of
Bcl-2 levels and the concomitant upregulation of Bax and caspase-3
levels in HeLa cells through the mitochondria pathway; these
results were different from those of a previous study (37). Some studies have shown that
prodigiosin from the supernatant of Serratia marcescens
could induce apoptosis as an immunosuppressor in hematopoietic
cancer cell lines (38). In
addition, there is also evidence that prodigiosin-induced apoptosis
could be associated with Bcl-2 and survivin inhibition in HT-29
cells (14). The most notable
finding was that prodigiosin had nearly identical cytotoxic effects
on the resistant cells when compared with their parental lines
(14). An in vitro study
also revealed that p53 was not induced by prodigiosin in Jurkat and
HL-60 cells (39). These results
suggest that prodigiosin induces apoptosis independently of p53 and
DNA damage.
In conclusion, to the best of the our knowledge, the
present study is the first to demonstrate that prodigiosin produced
by Serratia marcescens in the Periplaneta americana
gut has functions associated with inhibiting HeLa cell
proliferation and promoting apoptosis. Therefore, prodigiosin
isolated from Serratia marcescens from Periplaneta
americana may represent a novel class of anticancer agent.
However, due to the specific mechanism of apoptosis and complex
regulation, the role of prodigiosin in apoptosis mechanisms
requires further research to provide a theoretical and practical
foundation for the future use of prodigiosin in cancer
treatment.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from the
Public Welfare Research and Capacity Building Project of Guangdong
Province (grant no. 2016A030303059).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XBJ designed the experiments. PBL, JS, PYO, LYL and
ZYC conducted the experiments. FJC and JW analyzed the experimental
results. XBJ and PBL wrote the manuscript. All authors have read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Elkenawy NM, Yassin AS, Elhifnawy HN and
Amin MA: Optimization of prodigiosin production by Serratia
marcescens using crude glycerol and enhancing production using
gamma radiation. Biotechnol Rep. 14:47–53. 2017. View Article : Google Scholar
|
|
2
|
Zhang HC, Geng FN and Shen YM: The insect
gut microbiota plays crucial roles in modulating the interactions
between the host and intestinal pathogens. Chin J Ethnomed
Ethnopharmacy. 26:57–60. 2016.
|
|
3
|
Wang J: Effect of Periplaneta
americana extract on H125 lung cancer cells. Chin J Public
Health. 30:1400–1402. 2014.
|
|
4
|
Wang J and Xin L: Effects of
Periplaneta americana L. Extract on human hepatoma cells
Bel-7402. Chin J Modern Appl Pharm. 29:876–880. 2012.
|
|
5
|
Oh DC, Kauffman CA, Jensen PR and Fenical
W: Induced production of emericellamides A and B from the
marine-derived fungus emericella sp. in competing co-culture. J Nat
Prod. 70:515–520. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rastogi S, Zhang D and Davis JT: The
natural product prodigiosin binds G-quadruplex DNA. Supramolecular
Chem. 28:1–12. 2016. View Article : Google Scholar
|
|
7
|
Xia Y, Wang G, Lin X, Song X and Ai L:
Solid-state fermentation with Serratia marcescens, Xd-1
enhanced production of prodigiosin by using bagasse as an inertia
matrix. Annal Microbiol. 66:1239–1247. 2016. View Article : Google Scholar
|
|
8
|
Nakashima T, Tamura T, Kurachi M,
Yamaguchi K and Oda T: Apoptosis mediated cytotoxicity of
prodigiosin-like red pigment produced by gamma proteobacterium and
its multiple bioactivities. Biol Pharm Bull. 28:2289–2295. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sumathi C, MohanaPriya D, Swarnalatha S,
Dinesh MG and Sekaran G: Production of prodigiosin using tannery
fleshing and evaluating its pharmacological effects.
ScientificWorldJournal. 2014:290–327. 2014. View Article : Google Scholar
|
|
10
|
Osei-Poku J, Mbogo CM, Palmer WJ and
Jiggins FM: Deep sequencing reveals extensive variation in the gut
microbiota of wild mosquitoes from Kenya. Mol Ecol. 21:5138–5150.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Suzuki N, Ohtaguro N, Yoshida Y, Hirai M,
Matsuo H, Yamada YL, Imamura N and Tsuchiya T: A compound inhibits
biofilm formation of staphylococcus aureus from streptomyces. Biol
Pharm Bull. 38:889–892. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Douglas AE: Multiorganismal insects:
Diversity and function of resident microorganisms. Annu Rev
Entomol. 60:17–34. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Someya N, Nakajima M, Hamamoto H,
Yamaguchi I and Akutsu K: Effects of light conditions on
prodigiosin stability in the biocontrol bacterium Serratia
marcescens, strain b2. J General Plant Pathol. 70:367–370.
2004. View Article : Google Scholar
|
|
14
|
Hassankhani R, Sam MR, Esmaeilou M and
Ahangar P: Prodigiosin isolated from cell wall of Serratia
marcescens alters expression of apoptosis-related genes and
increases apoptosis in colorectal cancer cells. Med Oncol.
32:3662015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Miao L, Wang X, Jiang W, Yang S, Zhou H,
Zhai Y, Zhou X and Dong K: Optimization of the culture condition
for an antitumor bacterium serratia proteamacula 657 and
identification of the active compounds. World J Microbiol
Biotechnol. 29:855–863. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Patil CD, Patil SV, Salunke BK and
Salunkhe RB: Prodigiosin produced by Serratia marcescens
NMCC46 as a mosquito larvicidal agent against Aedes aegypti
and Anopheles stephensi. Parasitol Res. 109:1179–1187. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Marsac P, Pierard N, Porot L, Wim Van den
bergh, Grenfell J, Mouillet V, Pouget S, Besamusca J, Farcas F,
Gabet T, et al: Potential and limits of FTIR methods for reclaimed
asphalt characterization. Materials Structures. 47:1273–1286. 2014.
View Article : Google Scholar
|
|
18
|
Pradeep S, Sarath Josh MK, Balachandran S,
Sudha Devi R, Sadasivam R, Thirugnanam PE, Doble M, Anderson RC and
Benjamin S: Achromobacter denitrificans, SP1 produces
pharmaceutically active 25C prodigiosin upon utilizing hazardous
di(2-ethylhexyl)phthalate. Bioresour Technol. 171:482–486. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Danevčič T, Borić Vezjak M, Tabor M, Zorec
M and Stopar D: Prodigiosin induces autolysins in actively grown
bacillus subtilis cells. Front Microbiol. 7:272016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kimyon Ö, Das T, Ibugo AI, Kutty SK, Ho
KK, Tebben J, Kumar N and Manefield M: Serratia secondary
metabolite prodigiosin inhibits Pseudomonas aeruginosa
biofilm development by producing reactive oxygen species that
damage biological molecules. Front Microbiol. 7:9722016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Darshan N and Manonmani HK: Prodigiosin
inhibits motility and activates bacterial cell death revealing
molecular biomarkers of programmed cell death. AMB Express.
6:502016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee DJ, Lee JB, Jang HA, Ferrandon D and
Lee BL: An antimicrobial protein of the riptortus pedestris,
salivary gland was cleaved by a virulence factor of Serratia
marcescens. Dev Comp Immunol. 67:427–443. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen W, Liu Y, Zhang L, Gu X, Liu G,
Shahid M, Gao J, Ali T and Han B: Nocardia cyriacigeogica
from bovine mastitis induced in vitro apoptosis of bovine mammary
epithelial cells via activation of mitochondrial-caspase pathway.
Front Cell Infect Microbiol. 7:1942017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang J, Liu H, Sun Z, Xie J, Zhong G and
Yi X: Azadirachtin induced apoptosis in the prothoracic gland in
Bombyx mori and a pronounced Ca2+ release effect
in Sf9 cells. Int J Biol Sci. 13:1532–1539. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zheng B, Zhou Y, Zhang H, Yang G, Hong Z,
Han D, Wang Q, He Z, Liu Y, Wu F, et al: Dl-3-n-butylphthalide
prevents the disruption of blood-spinal cord barrier via inhibiting
endoplasmic reticulum stress following spinal cord injury. Int J
Biol Sci. 13:1520–1532. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Demain AL and Fang A: The natural
functions of secondary metabolites. Adv Biochem Eng Biotechnol.
69:1–39. 2000.PubMed/NCBI
|
|
28
|
Kan SC, Zang CZ, Yeh CW, Chang WF, Lin CC,
Hung TH, Shieh CJ and Liu YC: Enhanced bioconversion rate and
released substrate inhibition in (R)-phenylephrine
whole-cell bioconversion via partial acetone treatment. Enzyme
Microb Technol. 86:34–38. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sruthy PB, Anjana JC, Rathinamala J and
Jayashree S: The role of red pigment prodigiosin from bacteria of
earthworm gut as an anticancer agent. J Microbiol Biotechnol Food
Sci. 4:246–251. 2015. View Article : Google Scholar
|
|
30
|
Lapenda JC, Silva PA, Vicalvi MC, Sena KX
and Nascimento SC: Antimicrobial activity of prodigiosin isolated
from Serratia marcescens UFPEDA 398. World J Microbiol
Biotechnol. 31:399–406. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mienda BS, Idi A and Umar A:
Microbiological features of solid state fermentation and its
applications - An overview. Res Biotechnol. 2:21–26. 2011.
|
|
32
|
Kurbanoglu EB, Ozdal M, Ozdal OG and Algur
OF: Enhanced production of prodigiosin by Serratia
marcescens mo-1 using ram horn peptone. Braz J Microbial.
46:631–637. 2015. View Article : Google Scholar
|
|
33
|
Wei YH and Chen WC: Enhanced production of
prodigiosin-like pigment from Serratia marcescens, SMR by
medium improvement and oil-supplementation strategies. J Biosci
Bioeng. 99:616–622. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Suryavanshi MV, Waghmode SR, Bharti N,
Choudhari PB, Hingamire TB and Shouche YS: Isolation and virtual
screening of antimicrobial prodigiosin pigment from oxalotrophic
Serratia marcescens OX_R strain. J Appl Pharm Sci. 6:52–58.
2016. View Article : Google Scholar
|
|
35
|
Su JC, Chang JH, Huang JW, Chen PP, Chen
KF, Tseng PH and Shiau CW: Copper-obatoclax derivative complexes
mediate DNA cleavage and exhibit anti-cancer effects in
hepatocellular carcinoma. Chem Biol Interact. 228:108–113. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhou W, Jin ZX and Wan YJ: Apoptosis of
human lung adenocarcinoma A549 cells induced by prodigiosin
analogue obtained from an entomopathogenic bacterium Serratia
marcescens. Appl Microbiol Biotechnol. 88:1269–1275. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pérez-Tomás R and Vinas M: New insights on
the antitumoral properties of prodiginines. Curr Med Chem.
17:2222–2231. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Montaner B, Navarro S, Piqué M, Vilaseca
M, Martinell M, Giralt E, Gil J and Pérez-Tomás R: Prodigiosin from
the supernatant of Serratia marcescens induces apoptosis in
haematopoietic cancer cell lines. Br J Pharmacol. 131:585–593.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Elahian F, Moghimi B, Dinmohammadi F,
Ghamghami M, Hamidi M and Mirzaei SA: The anticancer agent
prodigiosin is not a multidrug resistance protein substrate. DNA
Cell Biol. 32:90–97. 2013. View Article : Google Scholar : PubMed/NCBI
|