Introduction
Non-small cell lung cancer (NSCLC) accounts for the
majority of lung cancers, and is the leading cause of
cancer-associated mortality worldwide (1). In NSCLC, the human epidermal growth
factor receptor (EGFR) is commonly amplified or mutated, leading to
constitutive activation of downstream signaling pathways, including
phosphoinositide 3-kinase (PI3K)/Akt and extracellular
signal-activated kinase signaling pathways. Furthermore EGFR
mutation is frequently encountered in women and non-smokers in East
Asia with NSCLC (2). The
application of EGFR tyrosine kinase inhibitors (TKIs), including
gefitinib, has marked efficacy with higher response rates and
longer progression-free survival (3,4).
However, patients with NSCLC who initially respond to EGFR-TKIs
often acquire resistance due to the secondary mutations of EGFR,
including T790M (5). Gene mutations
may be accelerated by environmental factors, including smoking, air
pollution and lifestyle. Environmental factors are as important as
genetic factors in provoking tumor occurrence and development.
Recently nutritional therapy has drawn increasing attention in
cancer treatment. Certain nutrients have been demonstrated to be
correlated with the progression of various diseases. For instance,
asparagine has been demonstrated to stimulate metastasis in a model
of breast cancer (6). Dietary fat
has been demonstrated to promote prostate cancer development,
suggesting that inhibition of fatty acid synthase may be useful for
controlling prostate cancer (7,8).
Hence, screening of functional dietary nutrients may provide an
approach for controlling lung cancer development and resistance to
therapies, including chemotherapy.
Heptadecanoic acid
(CH3(CH2)15COOH; C17:0), also
known as margaric acid, is an odd-chain saturated fatty acid
(OCS-FA). It exists in animal fat at a small concentration (1–5%),
while it hardly occurs in natural vegetable lipids. With the
development of analytical techniques, OCS-FAs were identified to be
rich in ruminant fats and fish oils. OCS-FAs are present at
insignificant plasma concentrations ranging from 0 to 1% in human
bodies. C17:0 and pentadecanoic acid
(CH3(CH2)13COOH; C15:0) are
commonly used as low cost internal standards in quantitative
analysis. Studies have indicated that C15:0 and C17:0 are
associated with health and several diseases, including the
incidence of coronary heart disease (9), prediabetes and type 2 diabetes
(10) as well as multiple sclerosis
(11). In addition, the tissue
levels of OCS-FAs were identified to be lower in Alzheimer's
disease compared with those in a control group (12). Certain studies have indicated that
odd branched-chain fatty acid (OBCFA) possesses a marked anti-tumor
activity (13,14). Similarly, C17:0 has been reported to
be effective against lymphoma cells (15). However, the efficacy of C17:0
against lung cancer cells and gefitinib-resistant NSCLC cells has
remained elusive. Studies on the association between OCS-FAs and
cancer would reveal more comprehensive functions of OCS-FAs and
provide a potential application of health nutritional therapy in
lung cancer prevention and treatment.
In the present study, the PC-9 NSCLC cell line and
PC-9 cells with acquired-gefitinib resistance (PC-9/GR) were used.
The effect of C17:0 on NSCLC was compared with that of other
dietary fatty acids contained in animal oils. In the experiments,
C17:0 was identified to be effective against PC-9 and PC-9/GR
cells. Furthermore, the effects of C17:0 on cell migration and
apoptosis, as well as alterations in the fatty acid composition and
associated signaling pathways were assessed.
Materials and methods
Cell culture
A normal human bronchial epithelioid (HBE) cell line
was purchased from the Cell Bank of Typical Culture Preservation
Committee of the Chinese Academy of Sciences (Shanghai, China), and
was maintained in KM medium (ScienCell Research Laboratories, Inc.,
San Diego, CA, USA). NSCLC cells (PC-9, sensitive to gefitinib;
PC-9/GR, resistant to gefitinib) were maintained in Dulbecco's
modified Eagle's medium (DMEM; Biological Industries, Kibbutz Beit
Haemek, Israel). The cell lines PC-9 and PC-9/GR were kindly
provided by Guangdong Lung Cancer Institute (Guangdong, China) and
were described in a previous study (16).
Cell proliferation and apoptosis
assay
Cells were equally seeded into 96-well plates at
37°C for incubation overnight. Various fatty acids (C14:0, C15:0,
C16:0, C17:0, C18:0, C18:1 or C20:0) (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) dissolved in anhydrous ethanol and filtered
(17) were added at the indicated
concentrations (10, 50, 90, 130, 170, 210 or 250 µM), followed by
incubation for 48 h. The cells were then incubated with MTT for 4 h
at room temperature and cell proliferation was determined using a
spectrophotometer (Molecular Devices, LLC, Sunnyvale, CA, USA) by
determining the optical density at OD 490 nm. For apoptosis
detection, an equal number of cells was washed with PBS and
re-suspended in binding buffer contained in the Annexin
V-fluorescein isothiocyanate (FITC) kit (TransGen Biotech Co.,
Ltd., Beijing, China). The cell suspension was then transferred to
a new reaction tube and stained by Annexin V-FITC and propidium
iodide (PI) according to the manufacturer's protocol. After
incubation in the dark for 15 min at 25°C, cells were re-suspended
in binding buffer and subjected to apoptosis measurement by flow
cytometry.
Cell migration assay
Cells were seeded in 6-well plates and cultured at
37°C with 5% CO2. until the concentration reached 95%.
Cells were scratched with a sterile pipette tip to generate a
cell-free path (width, ~1 mm) and cells were washed with PBS two
times, followed by culture in DMEM (Biological Industries)
containing the indicated fatty acids (C16:0, C17:0 or C18:0). After
48 h of incubation 37°C, images of the cells were captured through
a fluorescent microscope at low-magnification. The wound gaps were
measured and the mean healing ratios were quantified with ImageJ
software (National Institutes of Health, Bethesda, MA, USA).
Cell clone formation assay
PC-9 and PC-9/GR cells were seeded into a 6-well
plate at a density of 300 cells/well and incubated for 48 h. The
cells were refreshed with culture media with or without C17:0
(final concentraion, 50 µM) every 48 h. Approximately 12–14 days
later cells were fixed with 4% paraformaldehyde (Beijing Solarbio
Science & Technology Co., Ltd., Beijing, China) at 4°C for 30
min and stained with 0.1% crystal violet at 25°C for 10 min
(Beijing Solarbio Science & Technology Co., Ltd.) and cell
images were captured with a fluorescence microscope and the number
of cell colonies (>50 colonies) was counted.
Fatty acid analysis of the cells by
gas chromatography-mass spectrometry (GC-MS)
Equal amounts of PC-9 and PC-9/GR cells were seeded
for incubation overnight. Cells were incubated with or without
C17:0 (final concentration, 100 µM) for 48 h. The cells were then
collected and washed with PBS three times. Cells were counted and a
cell suspension was prepared with 3 ml HCl-methanol (v/v=5%),
followed by the addition of 10 µl of C15:0 internal standard (C15:0
in chloroform, the final concentration, 5 mg/ml). The tubes were
sealed and heated in an incubator at 90°C for 4 h. After natural
cooling to room temperature, 4 ml Na2CO3
(w/w=10%) was added. Subsequently 1 ml n-hexane was added, followed
by agitation for 5 min and centrifugation at 1,000 × g for 10 min.
A total of 400 µl of supernatant containing methyl esters of fatty
acids was transferred to an automatic injector vial and subjected
to a GC-MS assay according to the manufacturer's protocol. In
brief, the compositions of methyl esters of fatty acids in the
cells were analyzed by a 30-m polar capillary column (0.32 mm
internal diameter; SH-FameWax) on a Shimadzu gas chromatograph
containing a flame ionization detector (GCMS-QP2010 Ultra;
Shimadzu, Kyoto, Japan). Of the samples, 1 µl was injected
(injector temperature, 240°C). The column temperature was held at
130°C for 1 min, and increased at a rate of 3°C/min to 200°C, then
increased at a rate of 2°C/min to 220°C and maintained for 10 min.
Fatty acids in cells were quantified by an internal standard method
using C15:0 methyl ester.
Western blotting
Cells were collected and incubated with RIPA lysis
buffer (Beyotime Institute of Biotechnology, Haimen, China),
followed by centrifugation at 10,000 × g, 4°C for 10 min.
Supernatants were harvested and total protein was quantified by a
BCA assay. Total protein (30–60 µg) was subjected to 10% SDS-PAGE
and transferred onto polyvinylidene fluoride (PVDF) membranes,
followed by blocking with TBST (10 mM Tris-HCl, pH 7.4, 150 mM NaCl
and 0.1% Tween-20) containing 5% dried skimmed milk for 2 h at room
temperature. Then the membranes were blotted with primary
antibodies against GLUT1 (dilution 1:5,000; cat. no. ab115730;
Abcam, Cambridge, UK), GLUT4 (dilution 1:2,000; cat. no. ab654;
Abcam), p-S6K (dilution 1:1,000; cat. no. sc-8416; Santa Cruz
Biotechnology, Dallas, TX, USA), S6K (dilution 1:1,000; cat. no.
sc-8418; Santa Cruz Biotechnology), AKT (dilution 1:5,000; cat. no.
ab179463; Abcam), p-AKT (dilution 1:5,000; cat. no. 81283; Abcam)
and GAPDH (dilution 1:5,000; cat. no. ab181602; Abcam) at 4°C, for
overnight. After washing with TBST for three times, membranes were
further incubated with secondary antibody goat anti-rabbit IgG-HRP
(dilution 1:5,000; cat. no. AS014; ABclonal) or goat anti-mouse
IgG-HRP (dilution 1:5,000; cat. no. AS003; ABclonal) for 1 h at
room temperature. At the end, the membranes were incubated with a
ECL kit (cwbiotech) and subjected to exposure with X-ray films in
dark room.
Statistical analysis
Differences between two groups were statistically
analyzed by one-way analysis of variance with a Tukey's post hoc
test using SPSS 19.0 software (IBM Corp., Armonk, NY, USA). All
experiments were performed at least three times. Results were
expressed as the mean ± standard deviation. P<0.05 was
considered to indicate a statistically significant difference.
Results
C17:0 inhibits PC-9 and PC-9/GR cell
proliferation
Dietary saturated fatty acids may exert antitumor
effects, and screening functional fatty acids may provide
candidates that may serve as adjuvants in cancer therapy. Beef
tallow and mutton tallow are common dietary animal oils. C16:0,
C18:0 and C18:1 are the three dominant fatty acids in beef and
mutton tallow. In a preliminary analysis, the content of C17:0 was
determined to be 2.09% in beef tallow and 2.05% in mutton tallow
(data not shown).
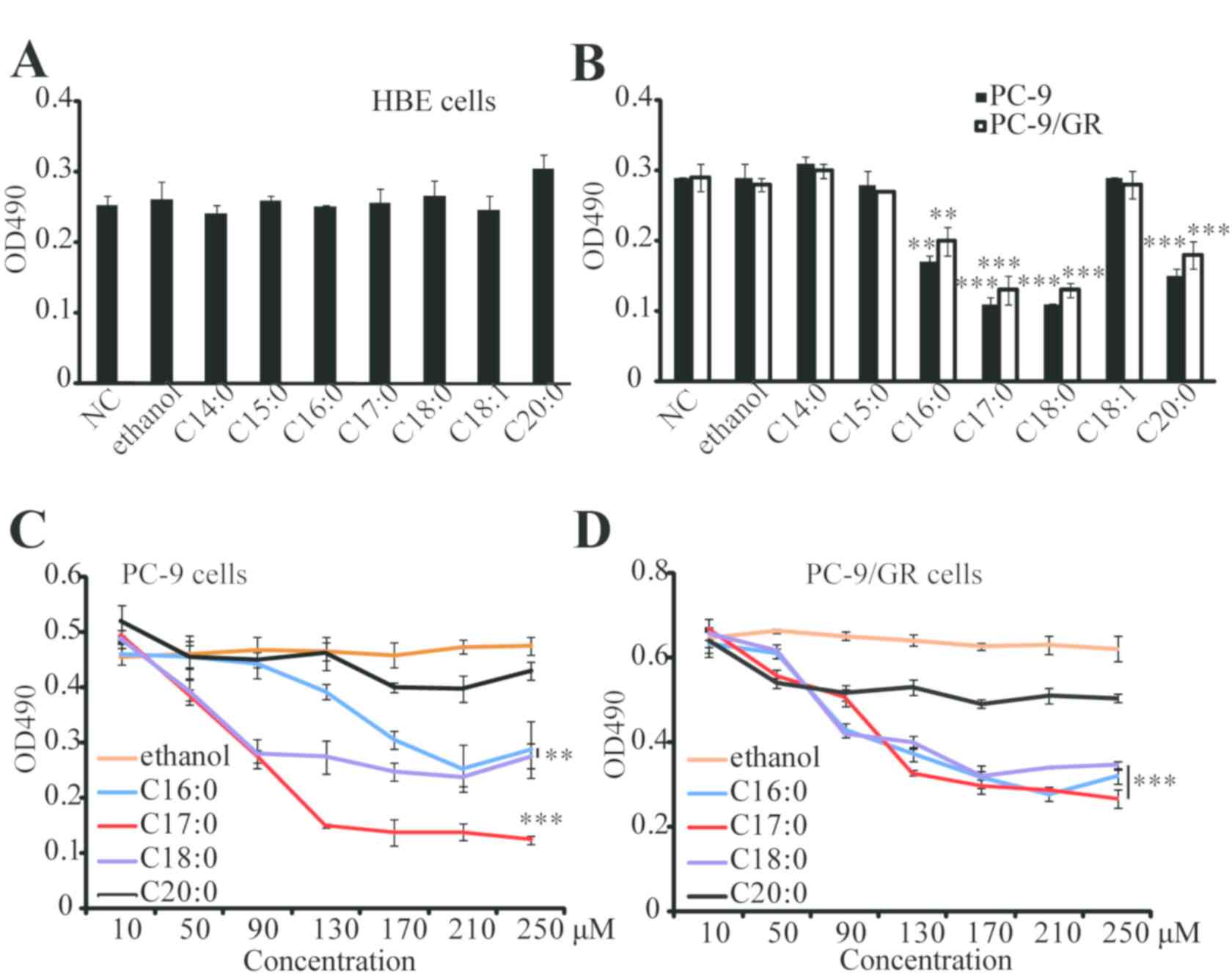
To investigate the impact of major dietary fatty
acids from ruminant animal oils on lung cancer cells, a cell
proliferation assay was performed. C14:0, C15:0, C16:0, C17:0,
C18:0, C18:1 or C20:0 were directly added into HBE, PC-9 and
PC-9/GR cells. As presented in Fig. 1A
and B, the results of the MTT assay indicated that these fatty
acids did not affect the proliferation of HBE cells, while 4 types
of fatty acids (C16:0, C17:0, C18:0 and C20:0) significantly
inhibited the proliferation of PC-9 and PC-9/GR cells. Furthermore,
C17:0 was identified to be the most effective fatty acid against
PC-9 and PC-9/GR cells, and its effects were dose-dependent
(Fig. 1C and D). The effect of
C17:0 in other lung cancer cells, including A549 and H1975, was
also examined. Similarly, the results revealed that C17:0 inhibited
cell proliferation in A549 and H1975 (data not shown). These
results indicated that C17:0 may be a fatty acid suitable for lung
cancer treatment.
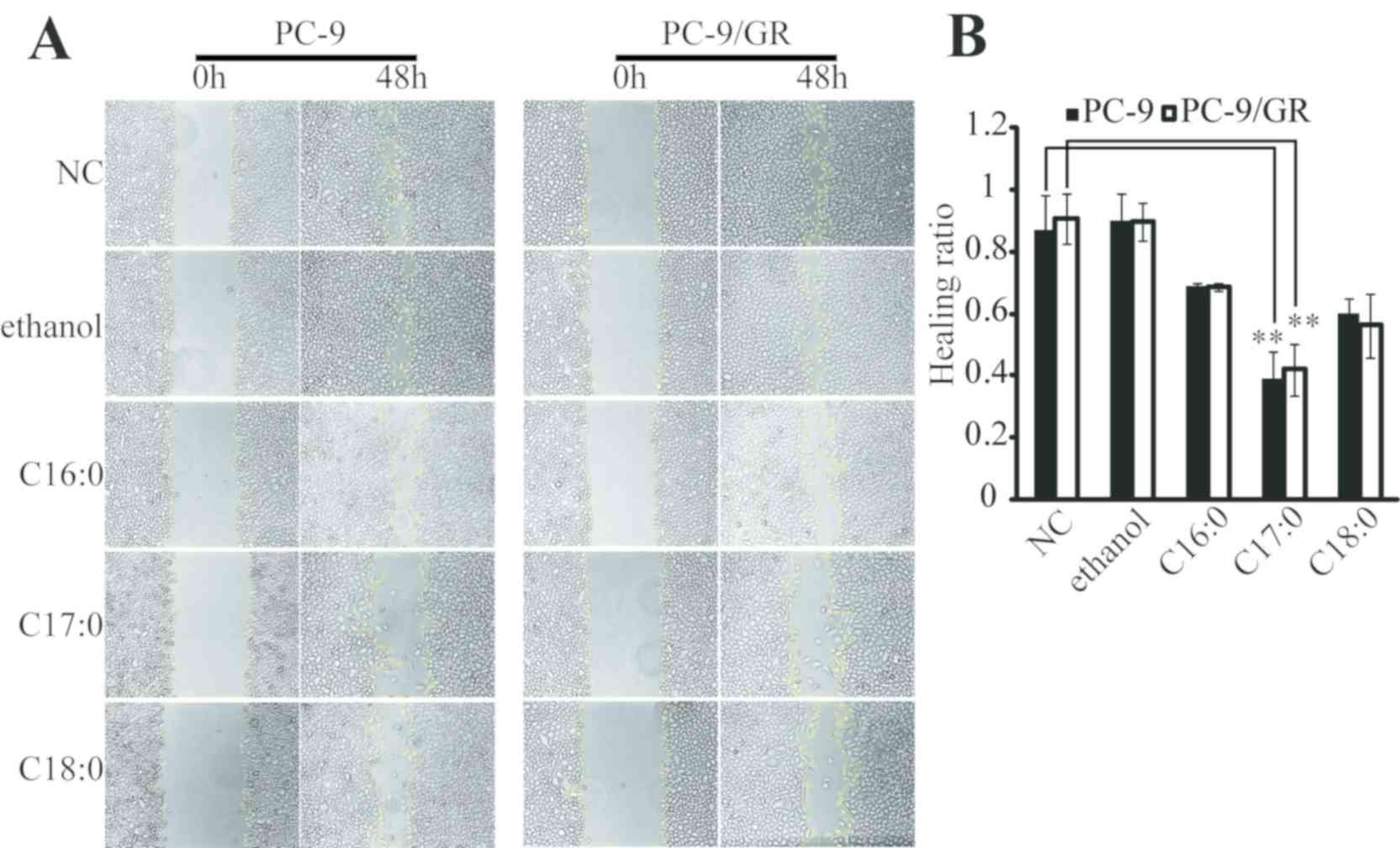
C17:0 inhibits cell migration, clone
formation and promotes apoptosis of PC-9 and PC-9/GR cells
The present study then focused on the functional
role of C17:0 in lung cancer cells. A wound healing assay indicated
that C16:0, C17:0 and C18:0 inhibited the wound closure ratio of
PC-9 and PC-9/GR cells (Fig. 2A and
B). Furthermore, the impact of C17:0 on cell migration was the
greatest among all fatty acids tested. The apoptosis ratio was also
determined by flow cytometry. As presented in Fig. 3A, C17:0 induced 16.62 and 29.63%
apoptosis in PC-9 cells and PC-9/GR cells respectively. C18:0 also
performed similarly in the two cell line variants. In addition, the
results of the cell clone formation assay indicated that C17:0
suppressed the proliferation of PC-9 and PC-9/GR cells (Fig. 3B). These results indicated that
C17:0 treatment in PC-9 and PC-9/GR cells was able to induce lung
cancer cell death.
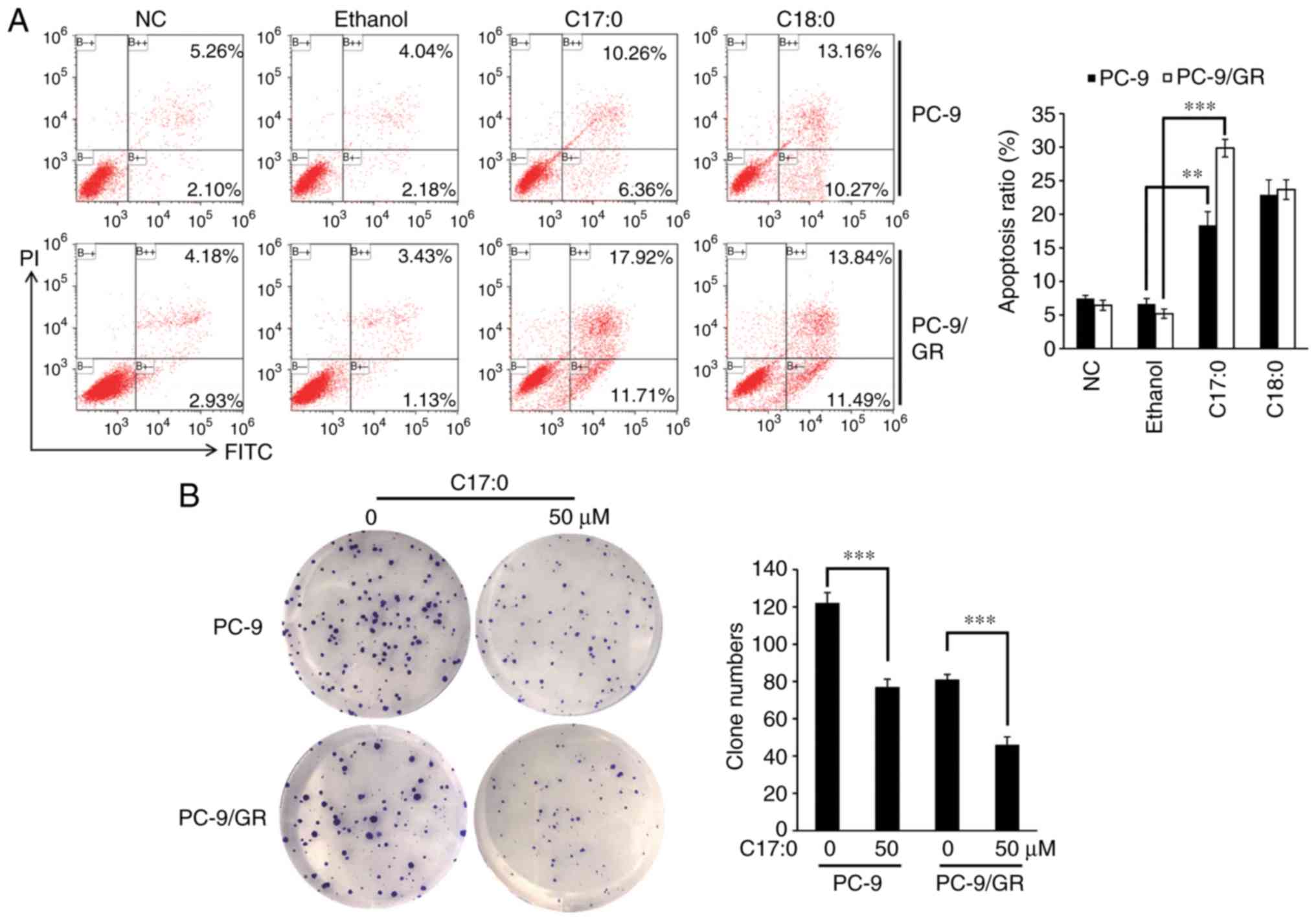 | Figure 3.C17:0 promotes apoptosis, while it
inhibits clone formation in PC-9 and PC-9/GR cells. (A) PC-9 and
PC-9/GR cells were seeded in 6-well plates and incubated overnight.
C17:0 or C18:0 was then added to the cells and after 48 h of
incubation, the cells were harvested and stained with FITC-Annexin
V and PI. Finally, cell apoptosis ratios in each group were
determined by flow cytometry. (B) PC-9 and PC-9/GR cells were
seeded in a 6-well plate and incubated for 48 h. The cells were
refreshed with media containing C17:0 (final concentration, 50 µM)
or without C17:0 every 48 h. After 12–14 days of culture, visible
cell colonies were fixed and stained. The number of cell colonies
was counted under a microscope. Values are expressed as the mean ±
standard deviation. **P<0.01, ***P<0.001,
experimental vs. the control group (ethanol group). C17:0,
heptadecanoic acid; C18:0, octadecanoic acid; PC-9/GR,
gefitinib-resistant PC-9 cell line; PI, propidium iodide; FITC,
fluorescein isothiocyanate. |
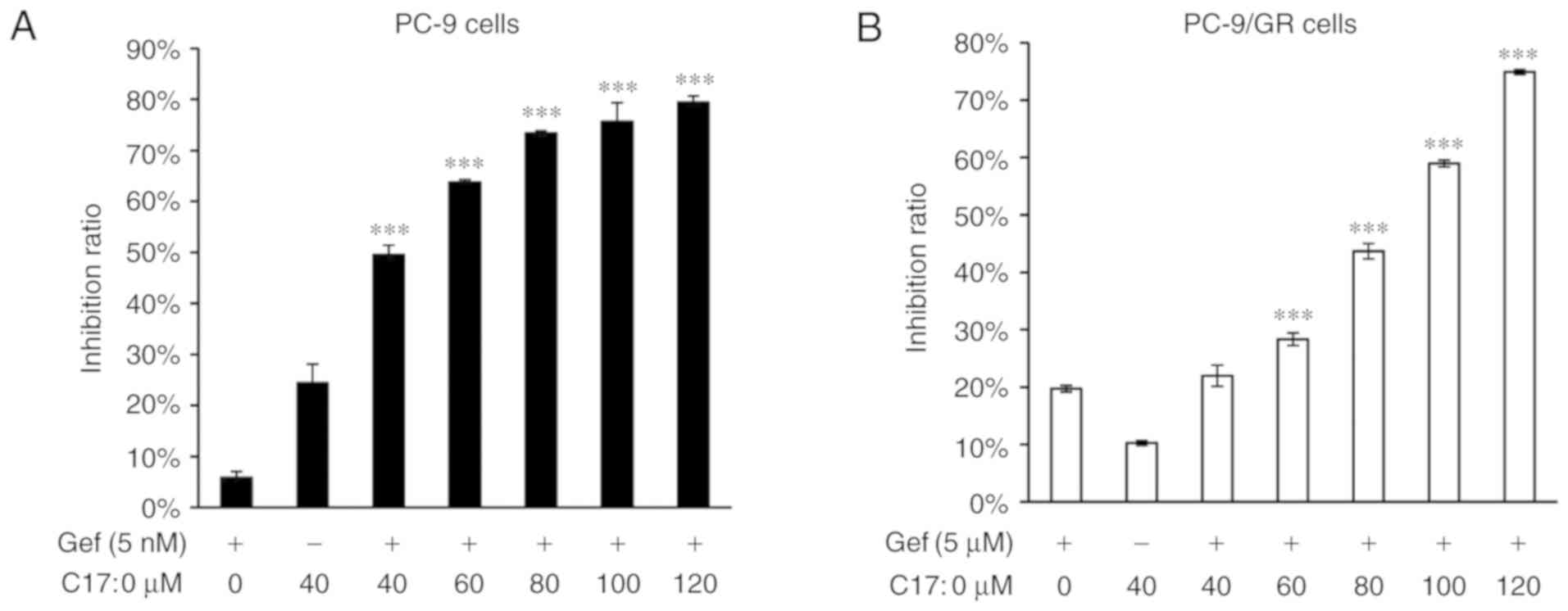
C17:0 enhances the cytotoxicity of
gefitinib in PC-9 and PC-9/GR cells
EGFR-TKIs are widely used in lung cancer therapy and
exhibit marked efficacy. To investigate the additive/enhancing
effect of C17:0 on EGFR-TKIs, an MTT assay was also performed on
PC-9 and PC-9/GR cells treated with gefitinib plus C17:0. As
presented in Fig. 4A and B, C17:0
promoted the inhibitory effect of gefitinib in PC-9 and PC-9/GR
cells in a dose-dependent manner. The greatest inhibition ratio in
the two cell variants reached ~80%. These results indicated that
C17:0 at increasing concentrations enhanced the effect of gefitinib
on NSCLC cells in vitro.
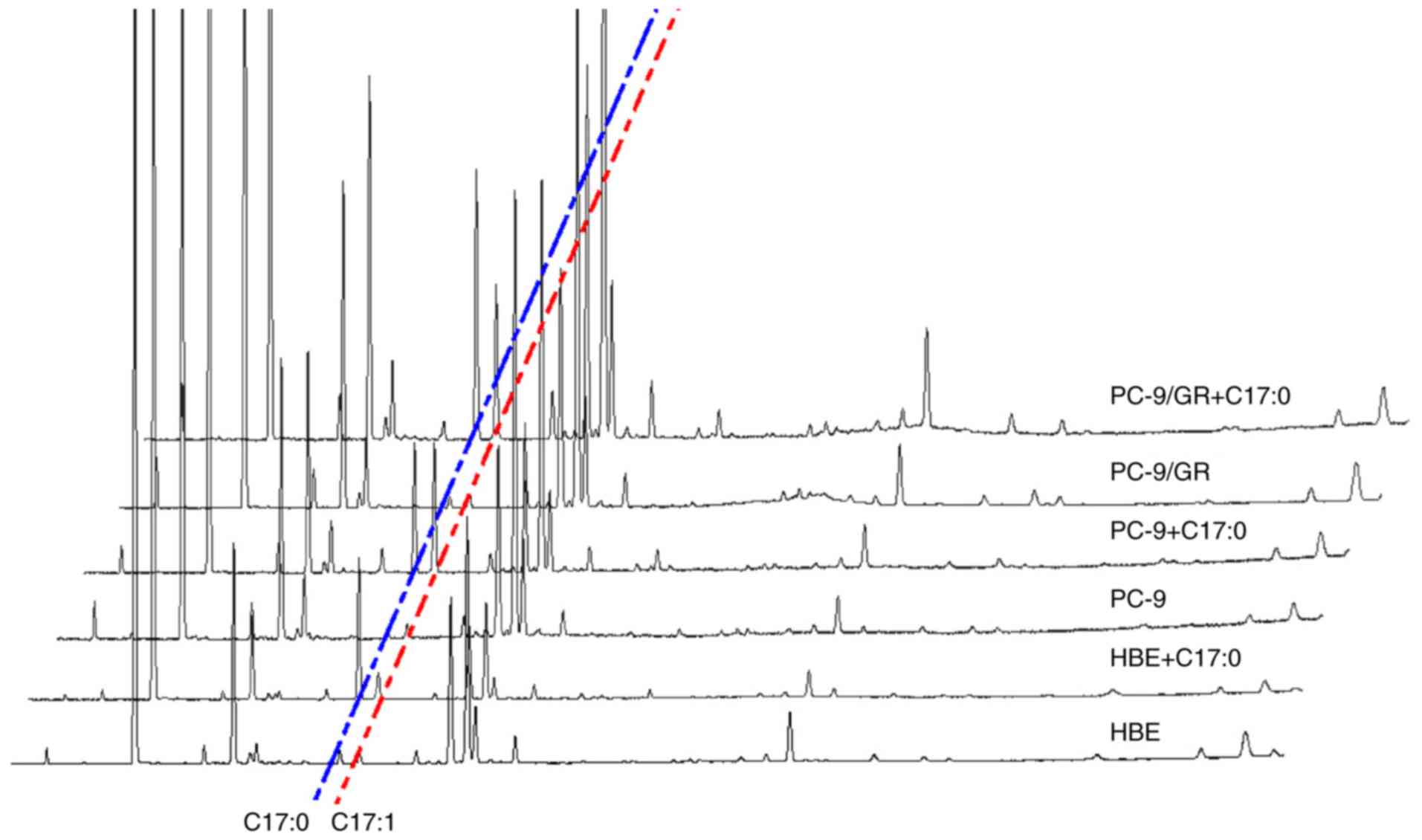
Effect of C17:0 on the fatty acid
composition of PC-9 and PC-9/GR cells
Changes in the fatty acid composition of PC-9 and
PC-9/GR cells in response to C17:0 treatment was assessed by GC-MS
and C15:0 was used as the internal standard. As presented in
Fig. 5 and Table I, the contents of various fatty
acids ranging from C14:0 to C24:0 were analyzed. In HBE, PC-9 and
PC-9/GR cells, the addition of C17:0 increased the content of
10-cis-heptadecenoic acid (C17:1). The concentration of
C17:1 in PC-9 and PC-9/GR cells was higher than that in HBE cells.
Furthermore, the concentration of C18:1 in HBE cells was lower than
that in PC-9 and PC-9/GR cells. In response to the addition of
C17:0, C18:1 was identified to be downregulated in HBE cells, while
it was not altered in PC-9 cells and only slightly upregulated in
PC-9/GR cells. In HBE cells, the total fatty acid composition was
not altered during incubation with C17:0. However, the ratio of
total fatty acids in PC-9 and PC-9/GR cells was higher than that in
HBE cells, indicating that these lung cancer cells have a higher
lipid synthesis activity and accumulation (Fig. 6). The increase of C17:1 in PC-9/GR
cells may contribute to lung cancer cell death.
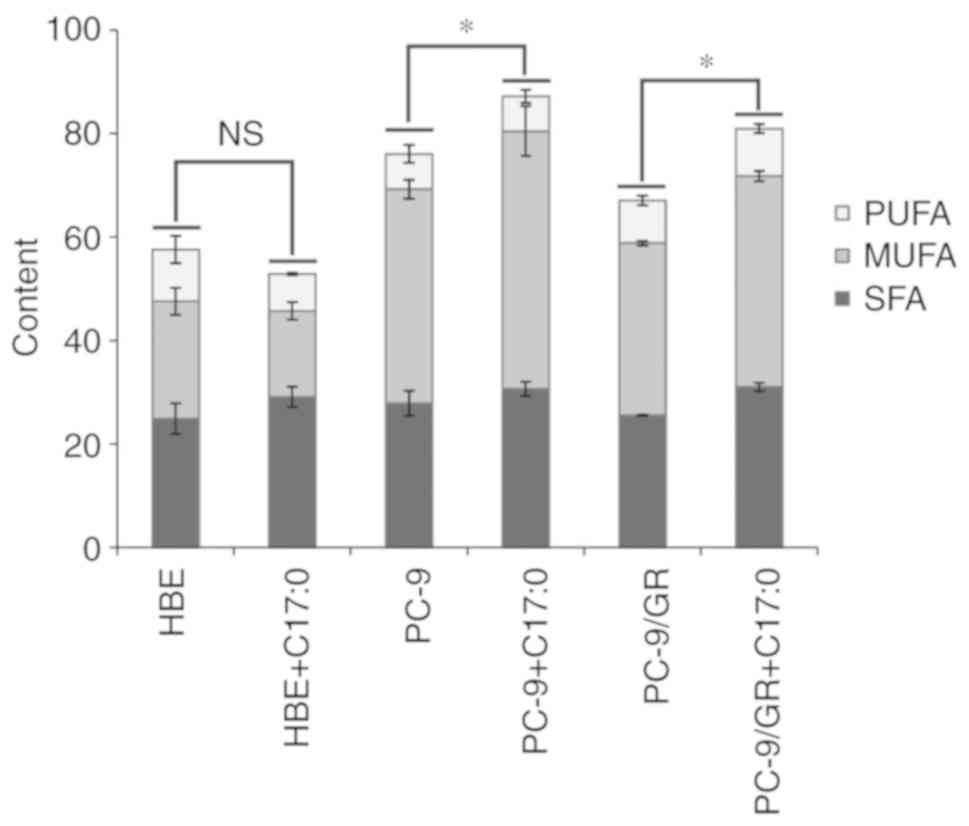 | Figure 6.Fatty acid composition assay by
GC-MS. Equal amounts of HBE, PC-9 and PC-9/GR cells were seeded
into plates and incubated overnight. C17:0 was then added and the
cells were incubated for 48 h, followed by collection and analysis
using the GC-MS assay. The ratios of SFA, MUFA and PUFA were
statistically analyzed. The ratios of total fatty acids in the
control group and C17:0-treated group were preceded to statistical
analysis. Values are expressed as the mean ± standard deviation.
*P<0.05, experimental vs. the control group. C17:0,
heptadecanoic acid; PC-9/GR, gefitinib-resistant PC-9 cell line;
HBE, human bronchial epithelioid cell line; GC-MS, gas
chromatography-mass spectrometry; SFA, saturated fatty acid; MUFA,
monounsaturated fatty acid; PUFA, polyunsaturated fatty acid. |
 | Table I.Fatty acid composition analysis. |
Table I.
Fatty acid composition analysis.
|
| HBE | HBE+C17:0 | PC-9 | PC-9+C17:0 | PC-9/GR | PC-9/GR+C17:0 |
|---|
| C14:0 | 0.64±0.07 |
0.33±0.04b | 1.41±0.26 |
1.32±0.11b | 1.45±0.06 |
1.16±0.04b |
| C16:0 | 12.12±1.02 |
7.4±0.73b | 14.8±0.55 | 12.6±0.46 | 12.64±0.17 |
9.98±0.1c |
| C16:1 | 1.97±0.34 |
1.04±0.02b | 4.9±0.63 | 4.27±0.59 | 3.38±0.18 |
2.71±0.04b |
| C17:0 | 0.7±0.2 |
12.99±0.42c | 0.47±0.18 |
7.56±0.48c | 0.39±0.05 |
8.09±0.1c |
| C17:1 | 0.43±0.39 |
2.83±0.1c | 0.9±0.3 |
8.58±0.29c | 0.31±0.27 |
4.56±0.14c |
| C18:0 | 10.8±1.08 |
7.97±0.78b | 10.98±1.3 | 9.18±0.67 | 10.78±0.44 |
11.84±0.49a |
| C18:1 | 20.03±1.69 |
12.71±1.63b | 35.32±1.07 | 36.98±3.98 | 29.16±0.71 |
33.15±0.81b |
| C18:2 | 1.65±0.27 | 1.33±0.12 | 1.29±0.3 | 1.44±0.05 | 1.41±0.04 |
1.8±0.05b |
| C19:0 | 0±0 |
0.44±0.03c | 0±0 | 0±0 | 0±0 | 0±0 |
| C20:0 | 0±0 | 0±0 | 0±0 | 0±0 | 0.12±0.2 | 0±0 |
| C20:1 | 0.24±0.22 | 0±0 | 0.23±0.06 | 0±0b | 0.28±0.3 | 0.31±0.01 |
| C20:3 | 0.59±0.55 | 0.29±0.04 | 0.49±0.09 | 0.22±0.2 | 0.2±0.18 | 0.39±0.34 |
| C18:3 | 0±0 | 0±0 | 0±0 | 0±0 | 0.11±0.19 | 0.2±0.35 |
| C20:4 | 3.96±0.48 |
3.13±0.01b | 2.68±0.5 | 2.86±0.13 | 3.08±0.32 |
3.95±0.42a |
| C20:5 | 0.39±0.36 | 0.59±0.06 | 0.14±0.25 | 0±0 | 0.28±0.26 | 0.44±0.38 |
| C22:0 | 0.22±0.23 | 0±0 | 0.21±0.36 | 0±0 | 0.17±0.3 | 0±0 |
| C22:1 | 0±0 | 0±0 | 0±0 | 0±0 | 0.15±0.25 | 0±0 |
| C22:4 | 0.23±0.25 | 0±0 | 0.18±0.31 | 0±0 | 0.35±0.31 | 0.24±0.42 |
| C22:6 | 3.17±0.79 |
1.81±0.32b | 2.07±0.75 | 2.19±0.9 | 2.81±0.66 | 2.14±0.34 |
| C24:0 | 0.45±0.4 | 0±0 | 0±0 | 0±0 | 0±0 | 0±0 |
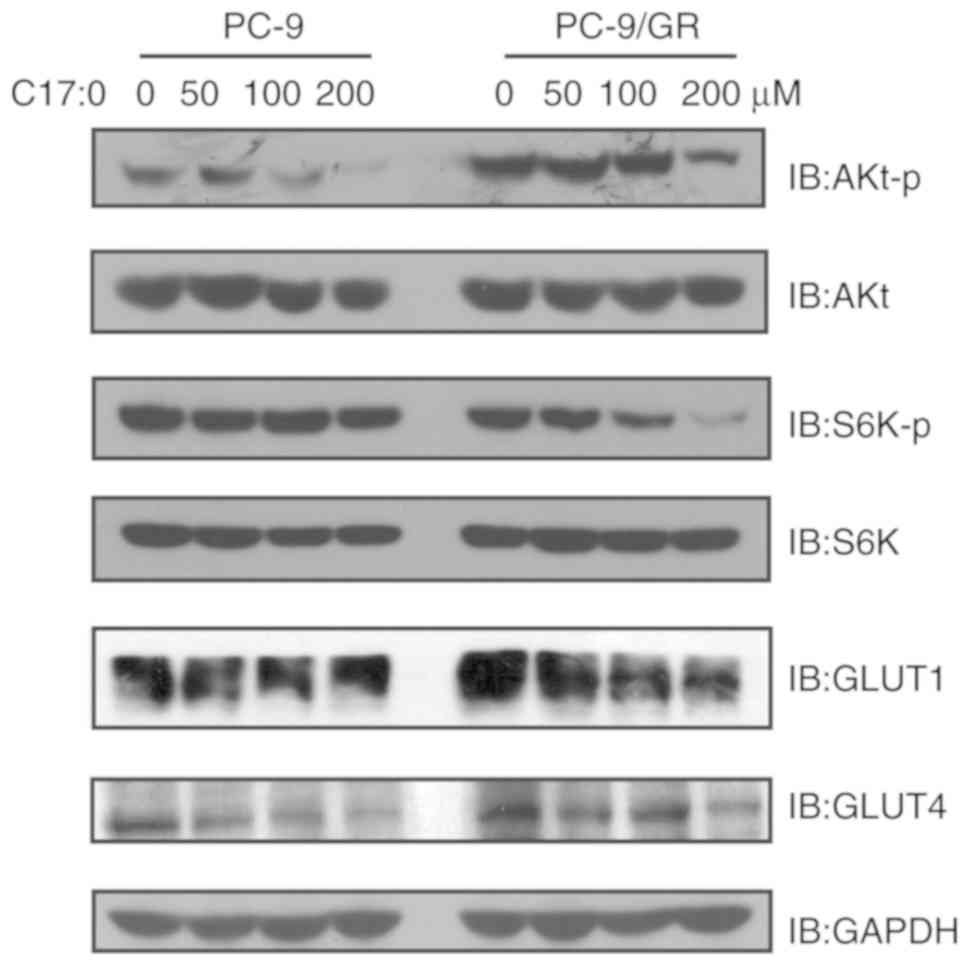
C17:0 inhibits the activation of
PI3K/Akt signaling in PC-9 and PC-9/GR cells
To further explore the molecular mechanisms
associated with the inhibition of C17:0 on PC-9 and PC-9/GR cells,
the activation of relevant signaling pathways was detected. The
PI3K/Akt signaling pathway is one pivotal pathway regulating cell
proliferation and tumor development. Therefore, its activation was
detected in PC-9 and PC-9/GR cells in response to treatment with
C17:0. As presented in Fig. 7, the
phosphorylation of Akt and S6K was inhibited. In particular, the
levels of p-S6K in PC-9/GR cells were markedly downregulated by
C17:0 in a dose-dependent manner. In general, cancer cells may
upregulate the expression or enhance the activation of glucose
transporters, including GLUT1 and GLUT4, to increase glucose import
from the extracellular environment into the cytoplasm to compensate
ATP production (18). Thus, the
expression of GLUT1 and GLUT4 in the two cell line variants in the
absence or presence of C17:0 was also detected. The results
indicated that treatment of PC-9 and PC-9/GR cells with C17:0
caused downregulation of GLUT1 and GLUT4. Collectively, the present
results indicated that addition of C17:0 to PC-9 and PC-9/GR cells
inhibited the activation of PI3K/Akt signaling and glucose
consumption.
Discussion
Recently, alteration of lipid metabolism in cancer
cells has drawn increasing attention. Accumulating studies have
reported that lipid metabolism is associated with lung cancer
development and resistance therapies, including chemotherapy
(19–23). Modulation of metabolic pathways has
been demonstrated to control cancer cell proliferation and death,
indicating that application of relative molecules or reagents
involved in this process may be beneficial for cancer therapy.
Fatty acids from dietary oils are the major substrates for lipid
synthase. The amounts and types of fatty acids ingested are
important for nutritional therapy in patients with cancer.
Polyunsaturated fatty acids (PUFAs) have also been demonstrated to
inhibit colon cancer cell growth (24). Palmitic acid has been demonstrated
to impair hepatocellular carcinoma development by suppressing cell
membrane fluidity and glucose intake (25). These studies indicate that
functional fatty acids may be beneficial for clinical cancer
therapy. Therefore investigation of the association between fatty
acid intake and lung cancer development may provide nutritional
suggestions for patients receiving lung cancer therapy. In the
present study, the effects of major dietary fatty acids derived
from ruminant animal oils in lung cancer cells were assessed using
the PC-9 cell line and its derivative with acquired
gefitinib-resistance PC-9/GR. The results indicated that C17:0
significantly inhibited the proliferation and migration of the two
cell line variants. Furthermore, treatment with C17:0 induced
greater apoptosis in PC-9 and PC-9/GR cells. The results of the MTT
assay also indicated that C17:0 enhanced the cytotoxic effect of
gefitinib. The present study demonstrated the role and function of
C17:0 in NSCLC and provided a potential application in clinical
treatment.
First, it was demonstrated that various
concentrations of C16:0, C17:0, C18:0 and C20:0 inhibit PC-9 and
PC-9/GR cells. Further study indicated that C17:0 had the greatest
cytotoxic effect in PC-9 cells, confirming that C17:0 is able to
inhibit NSCLC cells. This result indicated that certain saturated
fatty acids are able to inhibit tumor cell proliferation. Of note,
a previous study argued that saturated fatty acids are not the
primary cause for certain diseases, including arterial occlusion
(26). Therefore, saturated fatty
acids may have a potential function regarding health and diseases,
which requires further study and rational analysis. In addition,
C15:0 was not effective in PC-9 and PC-9/GR cells, while C17:0 was
particularly effective in NSCLC cells. The effect of OCS-FAs with
different lengths of carbon chain on NSCLC may also require further
study to provide more functional fatty acids for lung cancers.
Notably, C18:1 and cis−9,10-epoxy stearic acid, an oxidation
product of C18:1, have been previously reported to be effective in
decreasing the cell viability of HepG2 cells (27). However, the addition of C18:1 did
not affect the viability in PC-9 and PC-9/GR cells in the present
study, indicating that certain fatty acids may exert a different
impact on different tumor cell types.
Certain OBCFAs have been demonstrated to be
effective against certain types of tumor cells, e.g. breast cancer
(28), indicating that straight
odd-chain or branched-chain fatty acids may exert similar effects
on tumor cells. The present results indicated that C17:0
significantly suppressed wound healing in PC-9 and PC-9/GR cells.
Furthermore, C17:0 produced a greater increase in FITC-Annexin V
and PI-stained cells, suggesting that C17:0 triggers cell
apoptosis. Similarly, omega-3 PUFAs have been demonstrated to
inhibit A549 cell proliferation by promoting apoptosis and
inhibiting the PI3K/Akt signaling pathway (29). In the present study, the impact of
C17:0 on apoptosis and relevant signaling pathways was also
evaluated. The results were consistent with those of the
aforementioned study, indicating that C17:0 and omega-3
PUFA/docosahexenoic acid inhibit lung cancer cells via a similar
mechanism. Furthermore, potential changes in the fatty acid
profiles in the cell lines upon treatment with C17:0 were detected.
An increase of C17:1 was observed in two cell lines after
supplementation with C17:0. The function of C17:1 in lung cancer
cells remains elusive. However, stearoyl-CoA desaturase-1, a key
rate-limiting enzyme for monounsaturated fatty acid (MUFA)
synthesis, has been revealed to be involved in maintaining rapid
cell proliferation, apoptosis evasion, cancer cell development and
transformation in a panel of cancer types, including lung cancer
(30,31). The alteration of MUFAs is correlated
with cancer development, which may be contrary to the present
results, as no alteration of C17:1 was detected in these studies.
The results of the present study indicated that C17:1 may inhibit
lung cancer cell proliferation. Furthermore, supplementation of
C17:0 increased the ratios of C18:2 and C20:4, which may contribute
to enhanced oxidative stress and cell death. The function of C17:0
and C17:1 in lung cancer cells requires a more comprehensive
analysis.
To evaluate the effect of EGFR-TKIs plus C17:0 on
NSCLC, PC-9 and PC-9/GR cells treated with a combination of C17:0
and gefitinib were subjected to MTT assays. C17:0 was indicated to
enhance the inhibitory effect of gefitinib on PC-9 as well as
PC-9/GR cells. This combination effect was greater in PC-9 cells
than that in PC-9/GR cells, consistent with the effects of C17:0 in
the two cell line variants alone. These results indicated that
C17:0 may be a suitable candidate for nutritional therapy for lung
cancer. In fact, there are some references concerning the
physiological concentration of C17:0 in humans. The OCS-FAs are one
class of fatty acids with insignificant plasma concentrations,
which account for <0.5% total plasma fatty acid concentration
and their varying range in blood plasma is 0–1% (32,33).
Therefore, the physiological concentration of C17:0 is low.
However, studies have reported that C17:0 is a biomarker or
potential protective fatty acid against a series diseases, such as
metabolic syndrome and type 2 diabetes (34–36).
In addition, C17:0 concentration in plasma could be elevated
successfully by increasing people's dietary intake of C17:0-rich
dairy food (37,38). For example, a study reported that
the intake of conventional dairy products containing 1% milk, 1.5%
yogurt and 34% cheese could increase the level of cardioprotective
fatty acid C15:0 and C17:0 and have a minor effect on the lipid
profile (38). C17:0 is high in
milk and whole fat yogurt. Thus, we propose that C17:0 may be a
potential contributor for clinical treatment of lung cancer.
Supplementation of n-3 PUFAs has been demonstrated to be effective
to increase body weight and control the inflammatory status of
patients with lung cancer clinically (39,40).
Therefore, further study of EGFR-TKIs plus C17:0 in vivo is
required to validate the efficacy of C17:0 in lung cancer
treatment.
Acknowledgements
We would like to thank all the staff at the Public
Experiment Platform of the Institute of Health Sciences, Anhui
University for their help in GC-MS and flow cytometric
experiments.
Funding
The present study was supported, in part by the
National Natural Science Foundation of China (grant no. 31600749),
the Key Research and Development Projects in Anhui Province
(1704a07020075), the College Students' Innovation and
Entrepreneurship Training Project in Anhui Province (201810357296),
and the Initial Foundation of Doctoral Scientific Research in Anhui
University (Y01001487).
Availability of data and materials
The datasets/materials used in our present study are
available from the corresponding author through reasonable
request.
Authors' contributions
CX, PW, JG and LZ performed the main experiments and
collected the data. TM and BM conducted the GC-MS assays, collected
and analyzed the data. SY, GS and YY conducted the flow cytometric
assays, collected and analyzed the data. CX, XH, XY and BZ designed
the study, analyzed and drafted the manuscript. All authors read
and approved the manuscript and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pao W and Chmielecki J: Rational,
biologically based treatment of EGFR-mutant non-small-cell lung
cancer. Nat Rev Cancer. 10:760–774. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Maemondo M, Inoue A, Kobayashi K, Sugawara
S, Oizumi S, Isobe H, Gemma A, Harada M, Yoshizawa H, Kinoshita I,
et al: Gefitinib or chemotherapy for non-small-cell lung cancer
with mutated EGFR. N Engl J Med. 362:2380–2388. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sequist LV, Yang JC, Yamamoto N, O'Byrne
K, Hirsh V, Mok T, Geater SL, Orlov S, Tsai CM, Boyer M, et al:
Phase III study of afatinib or cisplatin plus pemetrexed in
patients with metastatic lung adenocarcinoma with EGFR mutations. J
Clin Oncol. 31:3327–3334. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yun CH, Mengwasser KE, Toms AV, Woo MS,
Greulich H, Wong KK, Meyerson M and Eck MJ: The T790M mutation in
EGFR kinase causes drug resistance by increasing the affinity for
ATP. Proc Natl Acad Sci USA. 105:2070–2075. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Knott SRV, Wagenblast E, Khan S, Kim SY,
Soto M, Wagner M, Turgeon MO, Fish L, Erard N, Gable AL, et al:
Asparagine bioavailability governs metastasis in a model of breast
cancer. Nature. 554:378–381. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen M, Wan L, Zhang J, Zhang J, Mendez L,
Clohessy JG, Berry K, Victor J, Yin Q, Zhu Y, et al: Deregulated
PP1alpha phosphatase activity towards MAPK activation is
antagonized by a tumor suppressive failsafe mechanism. Nature
communications. 9:1592018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen M, Zhang J, Sampieri K, Clohessy JG,
Mendez L, Gonzalez-Billalabeitia E, Liu XS, Lee YR, Fung J, Katon
JM, et al: An aberrant SREBP-dependent lipogenic program promotes
metastatic prostate cancer. Nat Genet. 50:206–218. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Khaw KT, Friesen MD, Riboli E, Luben R and
Wareham N: Plasma phospholipid fatty acid concentration and
incident coronary heart disease in men and women: The EPIC-Norfolk
prospective study. PLoS Med. 9:e10012552012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Meikle PJ, Wong G, Barlow CK, Weir JM,
Greeve MA, MacIntosh GL, Almasy L, Comuzzie AG, Mahaney MC,
Kowalczyk A, et al: Plasma lipid profiling shows similar
associations with prediabetes and type 2 diabetes. PLoS One.
8:e743412013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Holman RT, Johnson SB and Kokmen E:
Deficiencies of polyunsaturated fatty acids and replacement by
nonessential fatty acids in plasma lipids in multiple sclerosis.
Proc Natl Acad Sci USA. 86:4720–4724. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fonteh AN, Cipolla M, Chiang J, Arakaki X
and Harrington MG: Human cerebrospinal fluid fatty acid levels
differ between supernatant fluid and brain-derived nanoparticle
fractions, and are altered in Alzheimer's disease. PLoS One.
9:e1005192014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Adamska A and Rutkowska J: Odd- and
branched-chain fatty acids in milk fat-characteristic and health
properties. Postepy Hig Med Dosw (Online). 68:998–1007. 2014.(In
Polish). View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang Z, Liu S, Chen X, Chen H, Huang M and
Zheng J: Induction of apoptotic cell death and in vivo growth
inhibition of human cancer cells by a saturated branched-chain
fatty acid, 13-methyltetradecanoic acid. Cancer Res. 60:505–509.
2000.PubMed/NCBI
|
|
15
|
Fukuzawa M, Yamaguchi R, Hide I, Chen Z,
Hirai Y, Sugimoto A, Yasuhara T and Nakata Y: Possible involvement
of long chain fatty acids in the spores of Ganoderma lucidum
(Reishi Houshi) to its anti-tumor activity. Biol Pharm Bull.
31:1933–1937. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fei SJ, Zhang XC, Dong S, Cheng H, Zhang
YF, Huang L, Zhou HY, Xie Z, Chen ZH and Wu YL: Targeting mTOR to
overcome epidermal growth factor receptor tyrosine kinase inhibitor
resistance in non-small cell lung cancer cells. PLoS One.
8:e691042013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Meng H, Shen Y, Shen J, Zhou F, Shen S and
Das UN: Effect of n-3 and n-6 unsaturated fatty acids on prostate
cancer (PC-3) and prostate epithelial (RWPE-1) cells in vitro.
Lipids Health Dis. 12:1602013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gatenby RA and Gillies RJ: Why do cancers
have high aerobic glycolysis? Nat Rev Cancer. 4:891–899. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhan N, Li B, Xu X, Xu J and Hu S:
Inhibition of FASN expression enhances radiosensitivity in human
non-small cell lung cancer. Oncol Lett. 15:4578–4584.
2018.PubMed/NCBI
|
|
20
|
Ali A, Levantini E, Teo JT, Goggi J,
Clohessy JG, Wu CS, Chen L, Yang H, Krishnan I, Kocher O, et al:
Fatty acid synthase mediates EGFR palmitoylation in EGFR mutated
non-small cell lung cancer. EMBO Mol Med. 10:e83132018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shen M, Tsai Y, Zhu R, Keng PC, Chen Y,
Chen Y and Lee SO: FASN-TGF-beta1-PD-L1 axis contributes to the
development of resistance to NK cell cytotoxicity of
cisplatin-resistant lung cancer cells. Biochim Biophys Acta Mol
Cell Biol Lipids. 1863:313–322. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gouw AM, Eberlin LS, Margulis K, Sullivan
DK, Toal GG, Tong L, Zare RN and Felsher DW: Oncogene KRAS
activates fatty acid synthase, resulting in specific ERK and lipid
signatures associated with lung adenocarcinoma. Proc Natl Acad Sci
USA. 114:4300–4305. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li G, Li M, Hu J, Lei R, Xiong H, Ji H,
Yin H, Wei Q and Hu G: The microRNA-182-PDK4 axis regulates lung
tumorigenesis by modulating pyruvate dehydrogenase and lipogenesis.
Oncogene. 36:989–998. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang C, Yu H, Ni X, Shen S and Das UN:
Growth inhibitory effect of polyunsaturated fatty acids (PUFAs) on
colon cancer cells via their growth inhibitory metabolites and
fatty acid composition changes. PLoS One. 10:e01232562015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lin L, Ding Y, Wang Y, Wang Z, Yin X, Yan
G, Zhang L, Yang P and Shen H: Functional lipidomics: Palmitic acid
impairs hepatocellular carcinoma development by modulating membrane
fluidity and glucose metabolism. Hepatology. 66:432–448. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Malhotra A, Redberg RF and Meier P:
Saturated fat does not clog the arteries: Coronary heart disease is
a chronic inflammatory condition, the risk of which can be
effectively reduced from healthy lifestyle interventions. Br J
Sports Med. 51:1111–1112. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu Y, Cheng Y, Li J, Wang Y and Liu Y:
Epoxy stearic acid, an oxidative product derived from oleic acid,
induces cytotoxicity, oxidative stress, and apoptosis in HepG2
cells. J Agric Food Chem. 66:5237–5246. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wongtangtintharn S, Oku H, Iwasaki H and
Toda T: Effect of branched-chain fatty acids on fatty acid
biosynthesis of human breast cancer cells. J Nutr Sci Vitaminol
(Tokyo). 50:137–143. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yin Y, Sui C, Meng F, Ma P and Jiang Y:
The omega-3 polyunsaturated fatty acid docosahexaenoic acid
inhibits proliferation and progression of non-small cell lung
cancer cells through the reactive oxygen species-mediated
inactivation of the PI3K/Akt pathway. Lipids Health Dis. 16:872017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Igal RA: Roles of stearoylcoa desaturase-1
in the regulation of cancer cell growth, survival and
tumorigenesis. Cancers (Basel). 3:2462–2477. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Noto A, Raffa S, De Vitis C, Roscilli G,
Malpicci D, Coluccia P, Di Napoli A, Ricci A, Giovagnoli MR,
Aurisicchio L, et al: Stearoyl-CoA desaturase-1 is a key factor for
lung cancer-initiating cells. Cell Death Dis. 4:e9472013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ferrannini E, Barrett EJ, Bevilacqua S and
DeFronzo RA: Effect of fatty acids on glucose production and
utilization in man. J Clin Invest. 72:1737–1747. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nestel PJ, Straznicky N, Mellett NA, Wong
G, De Souza DP, Tull DL, Barlow CK, Grima MT and Meikle PJ:
Specific plasma lipid classes and phospholipid fatty acids
indicative of dairy food consumption associate with insulin
sensitivity. Am J Clin Nutr. 99:46–53. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Krachler B, Norberg M, Eriksson JW,
Hallmans G, Johansson I, Vessby B, Weinehall L and Lindahl B: Fatty
acid profile of the erythrocyte membrane preceding development of
type 2 diabetes mellitus. Nutr Metab Cardiovasc Dis. 18:503–510.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Maruyama C, Yoneyama M, Suyama N, Yoshimi
K, Teramoto A, Sakaki Y, Suto Y, Takahashi K, Araki R, Ishizaka Y,
et al: Differences in serum phospholipid fatty acid compositions
and estimated desaturase activities between Japanese men with and
without metabolic syndrome. J Atheroscler Thromb. 15:306–313. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Magnusdottir OK, Landberg R, Gunnarsdottir
I, Cloetens L, Akesson B, Landin-Olsson M, Rosqvist F, Iggman D,
Schwab U, Herzig KH, et al: Plasma alkylresorcinols C17:0/C21:0
ratio, a biomarker of relative whole-grain rye intake, is
associated to insulin sensitivity: A randomized study. Eur J Clin
Nutr. 68:453–458. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Forouhi NG, Koulman A, Sharp SJ, Imamura
F, Kröger J, Schulze MB, Crowe FL, Huerta JM, Guevara M, Beulens
JW, et al: Differences in the prospective association between
individual plasma phospholipid saturated fatty acids and incident
type 2 diabetes: The EPIC-InterAct case-cohort study. Lancet
Diabetes Endocrinol. 2:810–818. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Abdullah MM, Cyr A, Lépine MC, Labonté MÈ,
Couture P, Jones PJ and Lamarche B: Recommended dairy product
intake modulates circulating fatty acid profile in healthy adults:
A multi-centre cross-over study. Br J Nutr. 113:435–444. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Finocchiaro C, Segre O, Fadda M, Monge T,
Scigliano M, Schena M, Tinivella M, Tiozzo E, Catalano MG, Pugliese
M, et al: Effect of n-3 fatty acids on patients with advanced lung
cancer: A double-blind, placebo-controlled study. Br J Nutr.
108:327–333. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
van der Meij BS, Langius JA, Spreeuwenberg
MD, Slootmaker SM, Paul MA, Smit EF and van Leeuwen PA: Oral
nutritional supplements containing n-3 polyunsaturated fatty acids
affect quality of life and functional status in lung cancer
patients during multimodality treatment: An RCT. Eur J Clin Nutr.
66:399–404. 2012. View Article : Google Scholar : PubMed/NCBI
|