Introduction
Luteolin (3,4,5,7-tetrahydroxyflavone) is a natural
flavonoid abundant in various fruits and vegetables. Luteolin has
been reported to display a wide range of bioactive properties, such
as anti-virus (1,2), hepatoprotective (3), anti-inflammatory (4) and anti-diabetic effects (5). Recently, the antitumor effects of
luteolin have been extensively investigated in breast and gastric
cancer (6), melanoma (7), esophageal carcinoma (8), glioblastoma (9), hepatocellular carcinoma (10), placental choriocarcinoma (11) and lung adenocarcinoma (12). The role of luteolin in colorectal
cancer (CRC) has also been investigated. Luteolin has been found to
inhibit CRC cells proliferation, induce cells apoptosis and cell
cycle perturbation (13–15). Liu et al also reported that
luteolin was able to inhibit CRC cells epithelial to mesenchymal
transition by suppressing CREB1 expression (16). Luteolin was demonstrated to play the
protective roles in 1,2-dimethylhydrazine and azoxymethane
(AOM)-induced experimental colon carcinogenesis (17,18).
Moreover, luteolin increased the sensitivity of CRC cells to
chemotherapy (19,20).
MicroRNAs (miRNAs) are a category of small,
non-coding, single-stranded RNAs of 18–25 nucleotides in length
which bind to the 3′-untranslated regions (3′-UTRs) of their target
mRNAs. miRNAs regulate the expressions of target genes through
post-transcriptional silencing or induction of degradation
(21,22). Recent studies have revealed critical
functions of miRNAs in tumor biological processes, including cell
proliferation, differentiation, metastasis and chemosensitivity
(23,24). By regulating a variety of target
genes in cancer cells, miRNAs act as oncogenes or tumor
suppressors.
Several studies have reported the involvement of
miRNAs in the effects induced by luteolin. In prostate cancer
cells, luteolin treatment inhibited prostate cancer cells
proliferation and induced apoptosis through downregulation of
miR-301 by triggering DEDD2 expression (25). Zhou et al found that luteolin
upregulated miR-34 expression in gastric cancer cells and
upregulation of miR-34 enhanced the susceptibility of cells to
luteolin (6). Luteolin also
inhibited the tumorigenesis and induced the apoptosis of non-small
cell lung cancer cells by upregulation of miR-34a-5p by targeting
MDM4 (26). Thus, we hypothesized
that miRNAs play significant roles during luteolin treatment in CRC
cells and this has not yet been investigated.
In the present study, it was observed that luteolin
inhibited CRC cells migration and invasion both in vitro and
in vivo while had no effects on cells proliferation. miR-384
expression was markedly upregulated and inhibition of miR-384
partly reversed the inhibitory effects induced by luteolin.
However, the expression of pleiotrophin (PTN) was found to be
downregulated after luteolin treatment and was confirmed to be a
direct target of miR-384. By analyzing CRC tissues, we found that
miR-384 expression was downregulated while PTN expression was
upregulated compared with the paired-matched adjacent tissues. The
correlation analysis between PTN expression and clinical
characteristics revealed that PTN expression is positively related
to the cancer progression. Hence, the present study demonstrated
that luteolin exerts anticancer effects against CRC cells by
modulating PTN via miR-384 expression suggesting that PTN may serve
as a promising candidate for therapeutic applications in CRC
treatment.
Materials and methods
Cells and reagents
HT-29, SW480, SW620 and LoVo human CRC cell lines
were purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA). The normal human mucosal epithelial cell line
NCM460 was purchased from the National Institute of Cells
(Shanghai, China). All cells were maintained in RPMI-1640 medium
(HyClone; GE Healthcare, Chicago, IL, USA) supplemented with 10%
fetal bovine serum (FBS; Gibco™, Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) and 1% penicillin-streptomycin in a humidified
incubator containing 5% CO2 at 37°C. Luteolin was
purchased from Sigma-Aldrich/Merck (Shanghai, China) and dissolved
in dimethyl sulfoxide (DMSO).
CRC tissues and relative patient
clinical information
We collected a total of 50 cases of CRC and matched
normal tissues from patients who underwent surgery at Hangzhou
Hospital of Traditional Chinese Medicine from May 2017 to May 2018.
All postoperative pathology was determined to be colorectal
adenocarcinoma. The following patient characteristics were
recorded: age, sex, histologic differentiation, TNM stage and serum
levels of carcinoembryonic antigen (CEA) before surgery and all the
data are listed in Table I.
Approval of the present study was obtained from the Ethics
Committee of Hangzhou Hospital of Traditional Chinese Medicine and
informed consent was acquired from all the patients.
 | Table I.Clinicopathological characteristics
of PTN expression in the human CRC cases. |
Table I.
Clinicopathological characteristics
of PTN expression in the human CRC cases.
|
| Expression of
PTN |
|
|---|
|
|
|
|
|---|
|
Characteristics | Low (n=25) | High (n=25) | P-value |
|---|
| Age (years) |
|
| 0.7761 |
|
>60 | 15 | 13 |
|
|
≤60 | 10 | 12 |
|
| Sex |
|
| 0.3961 |
|
Male | 14 | 10 |
|
|
Female | 11 | 15 |
|
|
Differentiation |
|
| 0.1963 |
|
Well | 15 | 14 |
|
|
Moderate | 10 | 8 |
|
|
Poor | 0 | 3 |
|
| T factor |
|
| 0.0477a |
|
1+2 | 16 | 9 |
|
|
3+4 | 9 | 16 |
|
| N factor |
|
| 0.0449a |
| 0 | 18 | 11 |
|
|
1+2 | 7 | 14 |
|
| M factor |
|
| 0.0414a |
| 0 | 19 | 12 |
|
| 1 | 6 | 13 |
|
| CEA (ng/ml) |
|
| 0.3686 |
|
>5 | 13 | 9 |
|
| ≤5 | 12 | 16 |
|
Cell viability assay
Cell proliferation ability was detected by Cell
Counting Kit-8 (CCK-8) assay kit (Beyotime Institute of
Biotechnology, Shanghai, China). In brief, CRC cells were plated in
a 96-well plate at the initial density of 1,000 cells/well and
cultured in medium without or with luteolin at concentrations of 0,
10, 50 and 100 µM for 24, 48 and 72 h. CCK-8 solutions were added
into the cell cultures 2 h before harvesting. The optical density
at 450 nm was measured using an ultra-microplate reader (EMax;
Molecular Devices, Sunnyvale, CA, USA).
In vivo nude mouse study
Thirty-nine BALB/c nude mice (17–20 g) were utilized
to evaluate the effects of luteolin on tumor growth and tumor
metastasis in vivo. Five- to six-week-old female BABL/c nude
mice were purchased from Shanghai Slac Animal (Shanghai, China) and
maintained in the animal facility at the Zhejiang University
(Hangzhou, China). Mice were provided with water and food ad
libitum and kept under standard conditions (temperature 24±2°C,
humidity, 50–70 %, 12-h light/dark cycle). The study received
ethical approval from the Animal Care and Use Committee of Zhejiang
University, and experiments and animal care were performed
according to the approved protocols.
For the tumor growth assay, HT-29 cells
(1×106) were subcutaneously injected into the right
flanks of the nude mice. When the volumes of xenograft tumors
reached an average of 100 mm3, the mice were randomly
divided into two groups: the control group and the luteolin group
with 10 mice in each group (20 mice in total). Mice received
luteolin were intragastrically administered with luteolin at 100
mg/kg every 2 days for 30 days. The control group mice received the
same volume of phosphate-buffered saline (PBS). Tumor measurements
were performed with a caliper by measuring the largest diameter and
its perpendicular length every 3 days. According to the rules of
our ethics committee, humane endpoints set for the tumor
experiments were as follows: the tumor weight was >10% of the
original body weight or the average tumor diameter was >20 mm in
mice. Thus, when the tumor almost reached 20 mm, the experiment was
terminated. The mice were sacrificed by cervical dislocation, and
tumors were harvested and weighed on day 30. Tumor volume was
calculated using the following formula: V = ½ × (length ×
width2). To note, we repeated this experiment for the
same setting for three times (8 mice for the first experiment, and
(6 mice for the other two experiments).
For liver metastasis assay, mice (n=19) were
anesthetized with 75 mg/kg pentobarbital by intraperitoneal
injection. Mice were fixed and then a small left abdominal flank
incision (1 cm) was made and the spleen was exteriorized for the
intrasplenic injection. HT-29 cells (5×106) suspended in
100 µl ice-cold PBS were injected into the spleen with a 27-gauge
needle. A sterile cotton was held over the site of injection for 30
sec to prevent tumor cell leakage and bleeding. The injected spleen
was returned to the abdomen and the wound was sutured with 5-0
black silk. Mice receiving luteolin were intragastrically
administered with luteolin at 100 mg/kg every 2 days for 30 days.
The control mice received the equal volume of PBS. After 3 weeks,
the mice were sacrificed by cervical dislocation, spleen and liver
were removed and observed. The liver metastatic tumors on the
surface were calculated and H&E staining was performed. H&E
staining was performed at Servicebio Co., Ltd. (Wuhan, China). To
note, we also repeated this experiment three times (6 mice for the
first two experiments and 7 mice for the third trial).
Wound healing assay
HT-29 cells were seeded in 24-well plate until
reaching 80% confluency in the complete medium. The monolayer was
wounded using a sterile 200-µl micropipette tip. The cells were
washed with PBS three times to rinse off the detached cells. Cells
were incubated for 48 h in 2% FBS medium containing the indicated
concentrations of luteolin. Images of the wound morphology were
acquired under a light microscopy at ×100 magnification.
Transwell migration and invasion
assays
Cell migration and invasion potential were assessed
using 8-µm Transwell chambers (Corning Costar, Corning, NY, USA).
Briefly, 8×104 CRC cells suspended in 200 µl serum-free
medium treated with luteolin or not were seeded in the upper
chambers with the lower chamber filled with 600 µl of medium
supplemented with 20% FBS. For the invasion assay, the upper
surfaces of the membranes were coated with 50 µl Matrigel (BD
Biosciences, Franklin Lakes, NJ, USA) 6 h before cells were seeded.
Cells were allowed to migrate for 36 h before being fixed with
ice-cold methanol and stained with 0.1% crystal violet for 30 min.
Cells that passed through the membrane were photographed and
counted with an Olympus CKX41 light microscope (Olympus, Shanghai,
China; magnification, ×100).
Western blotting
Cells or CRC tissues for protein extraction were
lysed with RIPA buffer (Beyotime Institute of Biotechnology).
Protein concentration was determined by BCA assay (Cwbiotech,
Beijing, China). Total proteins (30 µg) were separated by 10%
SDS-PAGE gels and transferred to polyvinylidene difluoride (PVDF)
membranes. The membranes were incubated with primary antibodies
against human MMP-16 (cat. no. E-AB-32127), MMP-2 (cat. no.
10373-2-AP), MMP-3 (cat. no. 17873-1-AP), MMP-9 (cat. no.
10375-2-AP), PTN (cat. no. 27117-1-AP) (all diluted 1:1,000) and
GAPDH (cat. no. 60004-1-Ig) (dilution 1:5,000) at 4°C overnight.
Subsequently, the membranes were incubated with the peroxidase
(HRP)-conjugated goat anti-rabbit (dilution 1:5,000; cat. no.
SA00001-2) or goat anti-mouse (dilution 1:5,000; cat. no.
SA00001-1) secondary antibody at room temperature for 2 h. MMP-16
was purchased from Elabscience (Shanghai, China). All the other
antibodies were purchased from Proteintech Group Inc. (Chicago, IL,
USA). The protein bands were detected with FluorChem E System
(ProteinSimple, Santa Clara, CA, USA).
RNA isolation and quantitative
real-time PCR (qPCR)
Total RNA was extracted from the cells or the
surgically resected CRC and adjacent non-tumor tissues using TRIzol
reagent (Thermo Fisher Scientific, Inc.). Then, 1 µg of total RNA
was reverse transcribed using the HiFiScript cDNA Synthesis kit or
miRNA cDNA Synthesis kit (Cwbiotech) according to the
manufacturer's instructions. The PTN expression levels were
determined with UltraSYBR Mixture (Cwbiotech) and the miR-384
levels were determined with miRNA qPCR Assay Kit (Cwbiotech) on
CFX96 Touch Real-Time PCR Detection system (Bio-Rad Laboratories,
Hercules, CA, USA). The miR-384 and U6 primers were synthesized by
Shanghai GenePharma Co., Ltd. (Shanghai, China), and the PTN and
β-actin primers were synthesized by Thermo Fisher Scientific, Inc.
The primers were as follows: miR-384 forward,
5′-TGTTAAATCAGGAATTTTAA-3′ and reverse, 5′-TGTTACAGGCATTATGAA-3′;
U6 forward, 5′-CTCGTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′; PTN forward, 5′-ACCAGTGAGTCATCCGTCCA-3′
and reverse, 5′-TGCAAATTTTCGACGCTGCT-3′; β-actin forward,
5′-GTATCCTGACCCTGAAGTACC-3′ and reverse,
5′-TGAAGGTCTCAAACATGATCT-3′. The thermocycling conditions were as
follows: initial denaturation at 95°C for 10 min, followed by 40
cycles of 15 sec at 95°C and 1 min at 60°C. Relative fold-change in
expression of miR-384 and PTN was normalized to U6 and β-actin
expression, respectively. The relative expression was analyzed
using the 2−∆∆Cq (27)
method.
Plasmid, miR-384 mimic, miR-384
inhibitor and transfection
miR-384 mimic, miR-384 inhibitor and its relative
control miRNAs were synthesized by Shanghai GenePharma Co., Ltd.
The sequences are as follows (28):
miR-384 mimic: 5′-AUUCCUAGAAAUUGUUCAUA-3′, miR-384 inhibitor:
5′-UAUGAACAAUUUCUAGGAAU-3′, miR-384 mimic NC:
5′-UUCUCCGAACGUGUCACGU-3′, miR-384 inhibitor NC:
5′-CAGUACUUUUGUGUAGUACAA-3′. Full length clone DNA of human
pleiotrophin was obtained from Sino Biological Inc. (Shanghai,
China). Transfections were performed using Lipofectamine 2000
transfection reagent (Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions.
Mapping of prediction algorithm
data
Target genes for miR-384 were predicted using miRDB
(http://www.mirdb.org) and TargetScan v.5.1
(http://www.targetscan.org/). From the
TargetScan, we found that PTN ranked the top 2 genes that was the
targets of miR-384. We then confirmed it in the miRDR database and
found that PTN ranked 86. Thus, we chose PTN as one candidate of
miR-384 target genes.
Luciferase reporter assay
Wild-type (WT) or the mutated type (MUT) of
PTN-3′-UTR containing miR-384 binding sites was synthesized by
Shanghai GenePharma Co., Ltd., and cloned downstream of the
luciferase reporter gene. HT-29 cells were seeded into 24-well
plates (5×104 cells/well) one day before and then
co-transfected with the WT PTN-3′-UTR or the MUT PTN-3′-UTR vectors
(500 ng) and the miR-384 mimics or negative controls with
Lipofectamine 2000 transfection reagent (Thermo Fisher Scientific,
Inc.). After 48 h of transfection, cells were harvested, and the
luciferase activity was detected with the Dual-Luciferase Reporter
Assay system (Promega Corporation, Madison, WI, USA).
Oncomine database
Oncomine database (http://www.oncomine.org) was selected to search the
PTN expression levels between the CRC and normal groups.
Statistical analysis
Data are expressed as means ± SEM. Statistical
significance was determined by Student's t-test or one-way analysis
of variance (ANOVA) followed by Dunnett's test. The relationship
between the PTN expression and the CRC clinical features was
analyzed with the Chi-square test. P-value <0.05 was considered
as indicative of statistical significance.
Results
Luteolin inhibits CRC cells migration
and invasion both in vitro and in vivo
In order to explore the functions of luteolin in
CRC, we first detected HT-29 and SW480 cells proliferation after
luteolin treatment by CCK-8 assay. The results showed that luteolin
had no effects on CRC cells proliferation (Fig. 1A). We then examined cells migration
and invasion by Transwell and wound healing assays. As shown in
Fig. 1B, we observed that
luteolin-treated HT-29 and SW620 cells displayed significantly
reduced cells migration and invasion. Consistently, luteolin
treatment induced a slower closing of scratch wounds in the HT-29
cells (Fig. 1C), indicating that
luteolin inhibited cells migratory ability. We also detected the
expressions of several matrix metalloproteinases (MMPs), which are
strongly related with tumor migration and invasion and
overexpressed in human CRCs. As shown in Fig. 1D, luteolin obviously downregulated
MMP-2, MMP-3, MM-9 and MMP-16 expressions.
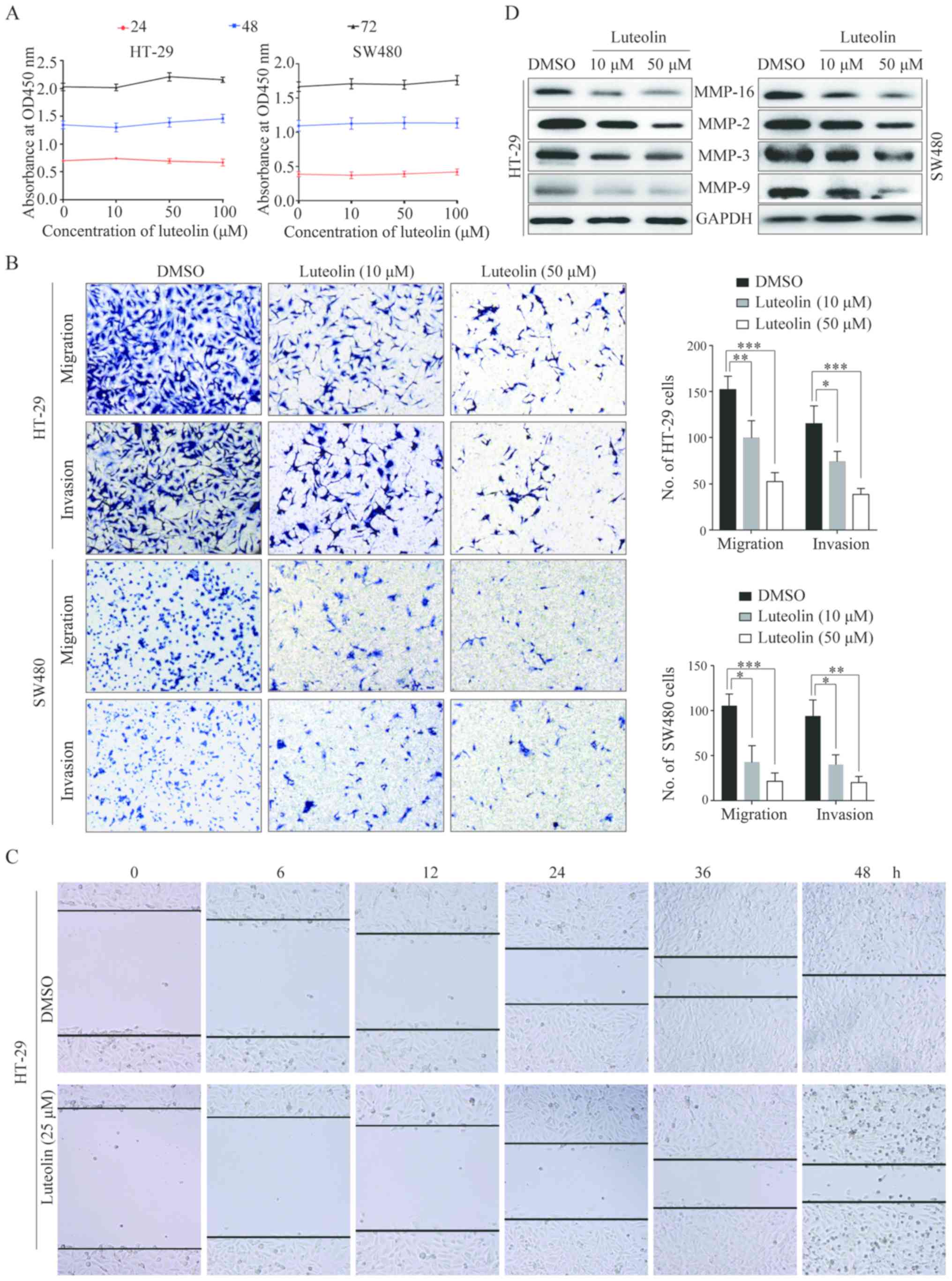 | Figure 1.Luteolin inhibits CRC cells migration
and invasion in vitro and in vivo. (A) HT-29 and
SW480 cells were treated with luteolin at the concentrations of 10,
50 and 100 µM. CCK-8 assay was performed after culturing for 24, 48
and 72 h. (B) Migration and invasion capacity of HT-29 and SW480
cells after treated with luteolin were determined and
representative images were taken. Original magnification, ×100. (C)
Monolayers of HT-29 cells were wounded by a tip and treated with
the indicated concentrations of luteolin. The representative images
of the wound healing were taken at 0, 6, 12 and 24 h after the
scratch was created. Original magnification, ×100. (D) Expressions
of MMP-2, MMP-3, MMP-9 and MMP-16 in HT-29 and SW480 cells were
detected by western blotting after stimulation with luteolin for 24
h. Results were obtained from 3 independent experiments and are
expressed as the means ± SEM. Statistical significance was
determined by one-way analysis of variance (ANOVA) followed by
Dunnett's test. *P<0.05, **P<0.01, ***P<0.001. CRC,
colorectal cancer; CCK-8, Cell Counting Kit-8; DMSO, dimethyl
sulfoxide. |
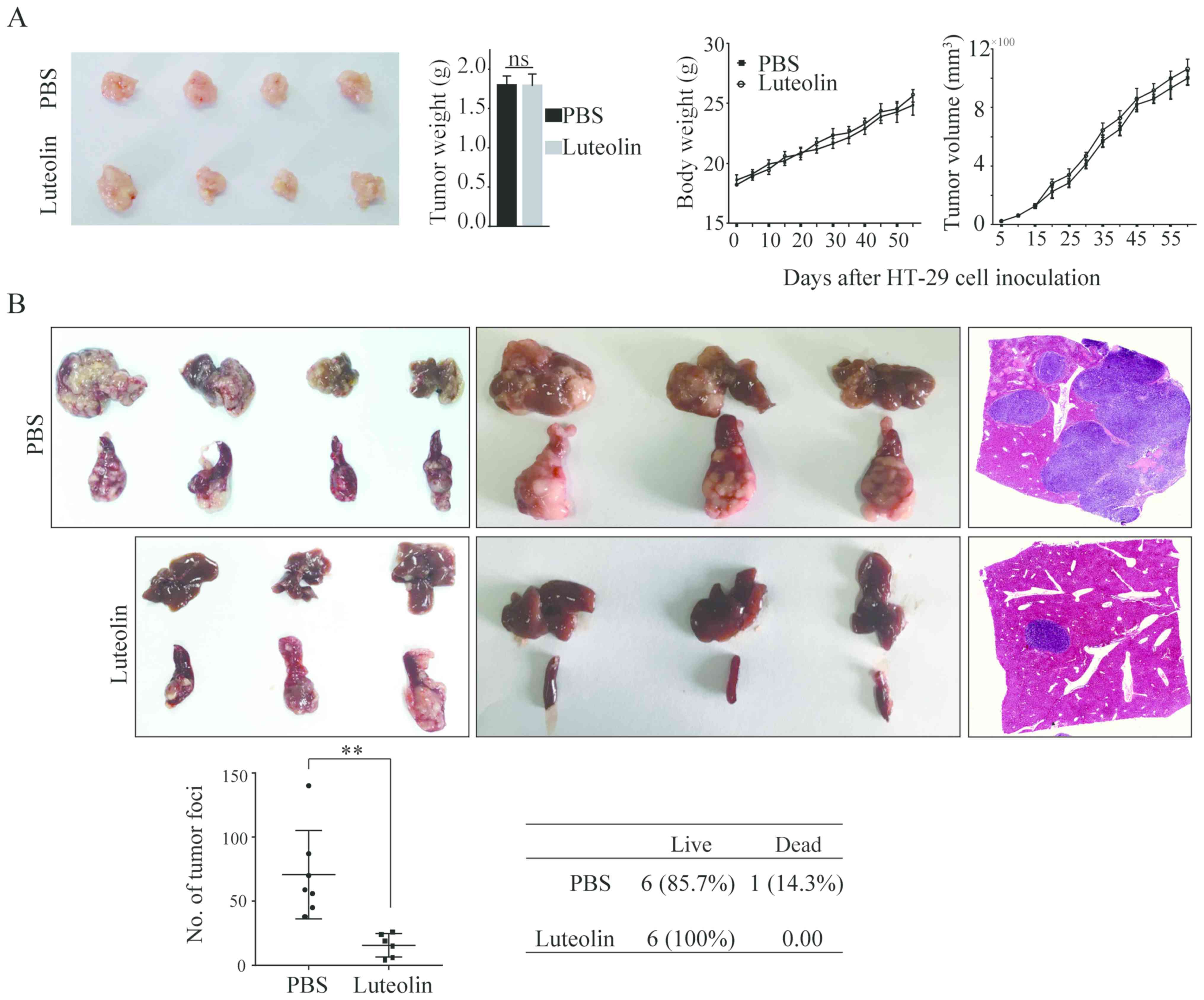
We then performed the in vivo assay to
observe the tumor growth and tumor metastasis by injecting HT-29
cells into the nude mice subcutaneously and into the spleen,
respectively. In accordance with the in vitro results,
intake of luteolin did not impact the tumor growth as evidenced by
no differences in tumor volume and tumor weight (Fig. 2A). However, luteolin significantly
inhibited HT-29 cells metastasis from the spleen to the liver
(Fig. 2B). The number of metastatic
nodules in the liver was significantly decreased. Taken together,
all these data suggest that luteolin plays a significant role in
suppressing the migration and invasion of CRC cells both in
vitro and in vivo while has no effects on cells
proliferation.
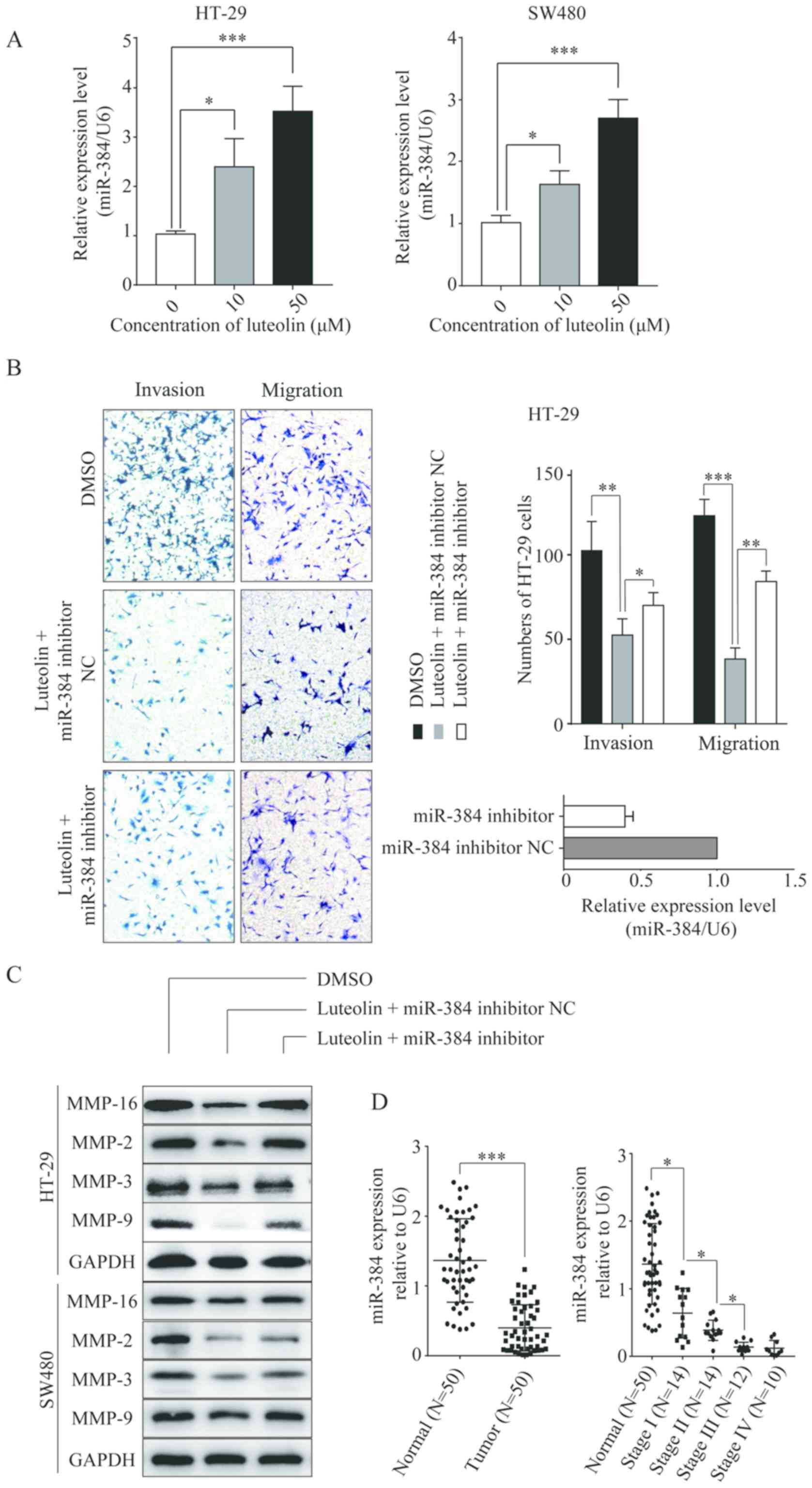
Luteolin inhibits CRC cells migration
and invasion by upregulating miR-384
To identify the specific miRNA involved in the
luteolin-induced inhibition of CRC cells migration and invasion,
the expression level of miR-384 was determined in CRC cells treated
with luteolin. It was found that the expression of miR-384 was
significantly elevated in the CRC cell lines following treatment
with luteolin in a dose-dependent manner compared with no-luteolin
stimulation (Fig. 3A). Then, we
constructed the miR-384 inhibitor and transfected into the CRC
cells before luteolin treatment. As shown in Fig. 3B, the miR-384 inhibitor partially
reversed the inhibition of cells migration and invasion induced by
luteolin. Simultaneously, the expression levels of MMP-2, MMP-3,
MMP-9 and MMP-16 which were decreased by luteolin treatment showed
an increase compared to the miR-384 inhibitor NC group (Fig. 3C). All these results imply that
miR-384 is involved in the anticancer effects of luteolin. We then
detected the miR-384 expression level in CRC tissues.
Downregulation of miR-384 expression was noted in the CRC tissues
compared with that observed in the non-tumor tissues (P<0.001;
Fig. 3D). Moreover, with the
progression of the tumor, the expression was decreased
significantly. The expression of miR-384 was lowest in patients
with TNM stage IV.
PTN is a direct target of miR-384
miRNAs exert their function by regulating the
expression of their target genes. Thus, we took advantage of the
prediction algorithms (miRDB and TargetScan v.5.1) to identify the
potential targets of miR-384. Among the potential target genes,
pleiotrophin (PTN) caught our interest. Thus, we detected the PTN
expression after luteolin stimulation. The results showed that PTN
expression was downregulated in a dose-dependent manner (Fig. 4A). Overexpression of PTN was able to
significantly reverse the luteolin-mediated inhibition of cell
migration and invasion as well as MMP expressions (Fig. 4B-D). This finding suggests that
luteolin inhibited CRC cells migration and invasion by
downregulating PTN expression. We next detected the relationship
between PTN and miR-384. The expression of PTN was decreased when
cells transfected with miR-384 mimic (Fig. 4E). Then miR-384 mimic or miR-384
mimic NC and PTN were co-transfected into CRC cells and the
luciferase activity was detected. Compared with the control, a
decrease in relative luciferase activity was observed when the WT
PTN 3′-UTR was co-transfected with miR-384. However, the miR-384
mimic did not affect luciferase activity in the mutant construct
(Fig. 4F). These data indicate that
miR-384 directly modulates PTN expression by binding to its 3′-UTR.
We also detected PTN expression after cells transfected with the
miR-384 inhibitor followed by treatment with luteolin. Results
showed that miR-384-inhibitor rescued the PTN downregulation
induced by luteolin (Fig. 4G),
confirming that luteolin-induced suppression of PTN expression was
mediated by miR-384.
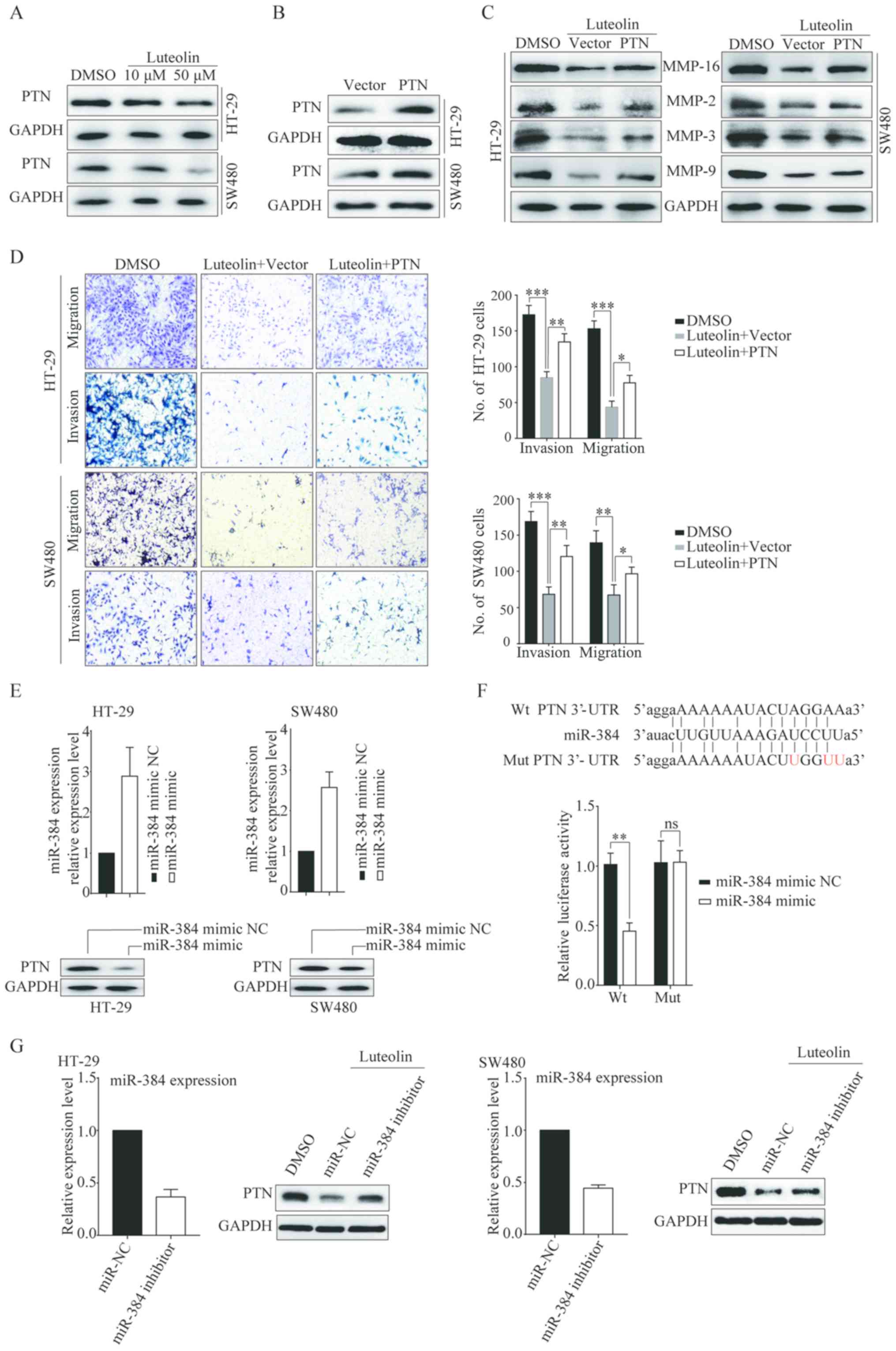 | Figure 4.PTN is a direct target of miR-384.
(A) The protein level of PTN in HT-29, and SW480 cells treated with
luteolin after 24 h was determined by western blotting. (B) HT-29
and SW480 cells were transfected with the vector or PTN
overexpression plasmid for 24 h, and the transfection efficiency
was determined by western blotting. (C) Transfected cells were
treated with luteolin for 24 h before protein was extracted for
western blotting to determine MMP-2, MMP-3, MMP-9 and MMP-16
expression. (D) Transfected cells were treated with luteolin for 24
h and cell migration and invasion abilities were determined by
Transwell assay. Original magnification, ×100. (E) HT-29 and SW480
cells were transfected with miR-384 mimic NC or miR-384 mimic for
24 h before proteins were extracted for western blotting to
determine PTN expression. (F) Putative Wt and Mut miR-384 binding
sites in the 3′-UTR of PTN. Relative luciferase activities were
analyzed in HT-29 and SW48T0 cells co-transfected with Wt or Mut
reporter plasmids and miR-384 mimic or miR-384 mimic NC. (G) HT-29
and SW480 cells were transfected with miR-384 inhibitor NC or
miR-384 inhibitor for 24 h before cells were treated with luteolin
for 24 h and then protein was extracted for western blotting to
determine PTN expression. Results are representative of 3
independent experiments and are expressed as mean ± SEM.
Statistical significance in D was determined by one-way analysis of
variance (ANOVA) followed by Dunnett's test and statistical
significance in F was determined by Student's t-test. ns, no
significant; *P<0.05, **P<0.01, ***P<0.001; ns, not
significant. PTN, pleiotrophin; DMSO, dimethyl sulfoxide; Wt,
wild-type; Mut, mutant. |
Expression of PTN in CRC tissues and
its relationship with clinical characteristics
Data from Oncomine database were analyzed to examine
the differential expression levels of PTN DNA between CRC and
normal tissues. As shown in Fig.
5A, PTN was upregulated in tumor tissues (P<0.001). We also
collected 50 cases of human CRC tissues and the paired adjacent
normal tissues. The relative expression level of PTN was measured
by qPCR (Fig. 5B) and western
blotting (Fig. 5C). Results showed
that PTN was highly expressed in CRC tissues. Furthermore, as the
tumor progressed, the expression level of PTN increased as well. We
also measured the PTN expression level in four CRC cell lines
(SW480, SW620, LoVo and HT-29) and one normal colon epithelial cell
(NCM460). The expression level of PTN was markedly higher in CRC
cell lines than that noted in the NCM460 cells (Fig. 5D). Then, we analyzed the expression
levels of miR-384 and PTN in tumor tissues, and we found that the
Pearson's correlation coefficient was −0.58, demonstrating a
negative correlation between miR-384 and PTN (Fig. 5E).
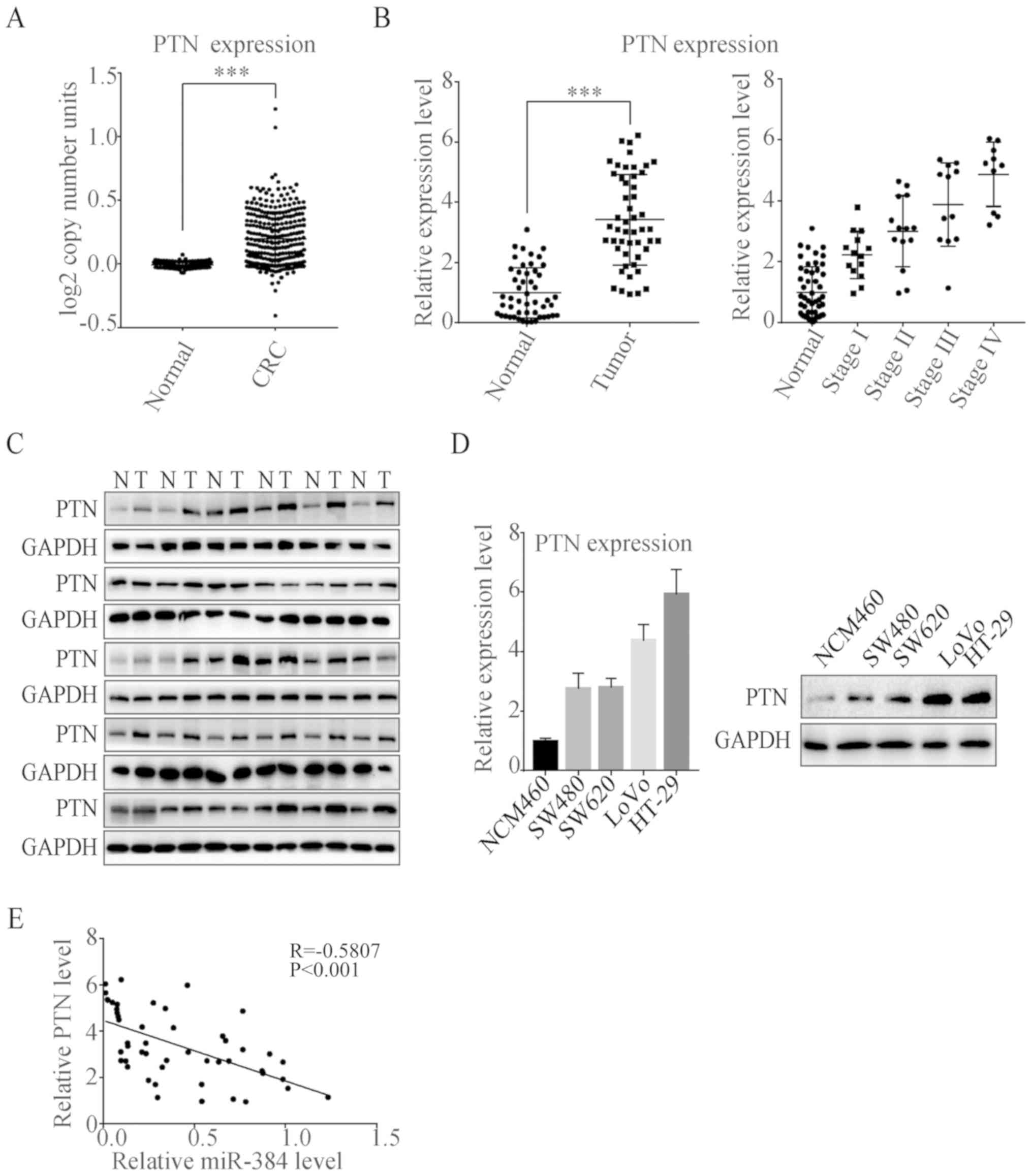 | Figure 5.PTN expression is downregulated in
CRC tissues and cell lines. (A) Comparison of DNA expression of PTN
in CRC and normal tissues from Oncomine database. (Normal, n=445;
CRC, n=302) (B) The PTN mRNA expression in 50 pairs of CRC tumor
tissues and the corresponding adjacent normal tissues were
determined by qPCR (Normal, n=50; Stage I, n=14; Stage II, n=14;
Stage III, n=12; Stage IV, n=10). (C) The PTN protein expression in
30 pairs of CRC tissues (T) and the corresponding adjacent normal
tissues (N) were determined by western blotting. (D) The mRNA and
protein level of PTN in four CRC cell lines (SW480, SW620, LoVo and
HT-29) and one normal human colon epithelium cell line NCM460 were
determined by qPCR and western blotting, respectively. Results are
representative of 3 independent experiments. (E) Pearson's
correlation analysis of the negative correlation between the
expression of miR-384 and PTN in colorectal cancer tissues. Results
are expressed as mean ± SEM. ***P<0.001. PTN, pleiotrophin; CRC,
colorectal cancer. |
We next divided the patients into two groups
according to the PTN expression level (25 patients in each group).
The relationship between PTN expression and the clinical
characteristics including age, sex, tumor differentiation, TNM
stage and serum CEA level was determined. The results are shown in
Table I. The differential
expression of PTN was significantly associated with tumor size, N
stage and M stage of CRC patients (all P<0.05).
Discussion
Currently, the prognosis or therapy for patients
with advanced stage of colorectal cancer (CRC) is poor. Treatment
includes chemotherapy, radiotherapy and surgery. Recently,
traditional Chinese medicine has attracted much attention in the
treatment of cancer due to their extensive tumor-suppressive
activity, safety and inexpensiveness. One such reagent is luteolin.
In the present study, luteolin was found to inhibit CRC cells
migration and invasion both in vitro and in vivo, but
it did not affect the proliferation of CRC cells. Further
experiments revealed the important role of the miR-384/PTN axis in
the luteolin-induced effects.
Many reports have reported that luteolin can induce
cell cycle arrest and apoptosis in CRC cells. For example, Krifa
et al reported that luteolin induced cytotoxicity in human
CRC cell line BE and the IC50 value was 90.13 and 48.32
µM for 24 and 48 h, respectively (13). Kang et al showed that
luteolin inhibited HT-29 cells proliferation in a dose-dependent
manner from 5 to 100 µg/ml (29).
Luteolin also decreased the cell viability of human colon carcinoma
cell line Caco-2 (30). In the
present study, we found that luteolin had no effects on HT-29 and
SW480 cells proliferation. We hypothesized that the CRC cells used
were different and thus the sensitivity to luteolin may be varied.
Thus, the concentrations of 0–100 µM of luteolin in this study had
no effects on cells proliferation.
Pleiotrophin (PTN) is a small heparin-binding
cytokine which can be induced during tumorigenesis (31). Levels of PTN have been found to be
increased in several cancer cell lines and primary tumors,
including pancreatic cancer (32),
hepatocellular carcinoma (33),
breast (34) and papillary thyroid
cancer (35). PTN promotes tumor
progression either through direct effects on tumor cells or through
stimulation of angiogenesis and remodeling of the tumor
microenvironment (36–38). In CRC, Yamakawa et al first
reported that the PTN expression level was decreased in CRC
compared with those in normal adjacent mucosae (39). Kong et al reported that PTN
was highly expressed in CRC tissues and its expression was related
to CRC differentiation and TNM staging, and a high level of PTN is
a predictor of poor prognosis (40). In this study, by analyzing the data
from the Oncomine database and clinical samples, we also found that
PTN expression was much higher in CRC tissues than that in the
normal tissues and its expression was related to TNM staging while
PTN had no relationship with tumor differentiation (Table I). We speculate that the probable
reason is that the sample size in the present study was too small.
Further research with an increased number of samples is needed.
Growing evidence suggests that miR-384 is a potent
tumor-suppressor molecule. The functions of miR-384 have been
studied in various types of cancers including papillary thyroid
(41), pancreatic (42) and non-small cell lung cancer
(43). Astrocyte elevated gene-1
(43,44), PTN (33), piwi-like RNA-mediated gene silencing
4 (PIWIL4) (45), KRAS and CDC42
(28) have been identified as
target genes of miR-384. Meanwhile, miR-384 was found to be
negatively regulated by long non-coding RNA CRNDE (45,46).
In CRC, miR-384 was downregulated and closely related with the
progression. Inhibition of miR-384 promoted the invasive and
metastatic abilities of CRC cells (28). Sun et al reported that in
papillary thyroid cancer cells PTN was found to be a downstream
target of miR-384 (41). We also
confirmed by luciferase reporter assay that in CRC cells PTN is a
direct target of miR-384.
In summary, our results demonstrated the antitumor
role of luteolin in CRC and the mechanism of which is partly
mediated via the miR-384/PTN axis. Thus, miR-384 or PTN can be
developed as a therapeutic target in the treatment of CRC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
YY, CR and SW participated in the conception and
design of the study. YY, CR and GZ performed the statistical
analysis and were involved in the preparation of the figures. YY
and SW reviewed the results and participated in the discussion of
the data. YY, CR and SW prepared the manuscript and revised it. All
authors read and approved the manuscript and agreed to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Approval of present study was obtained from the
Ethics Committee of the Hangzhou Hospital of Traditional Chinese
Medicine and informed consent was acquired from all the patients.
The animal study received ethical approval from the Animal Care and
Use Committee of Zhejiang University, and experiments and animal
care were performed according to the approved protocols.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Peng M, Watanabe S, Chan KW, He Q, Zhao Y,
Zhang Z, Lai X, Luo D, Vasudevan SG and Li G: Luteolin restricts
dengue virus replication through inhibition of the proprotein
convertase furin. Antiviral Res. 143:176–185. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Peng M, Swarbrick CMD, Chan KW, Luo D,
Zhang W, Lai X, Li G and Vasudevan SG: Luteolin escape mutants of
dengue virus map to prM and NS2B and reveal viral plasticity during
maturation. Antiviral Res. 154:87–96. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang H, Tan X, Yang D, Lu J, Liu B,
Baiyun R and Zhang Z: Dietary luteolin attenuates chronic liver
injury induced by mercuric chloride via the Nrf2/NF-κB/P53
signaling pathway in rats. Oncotarget. 8:40982–40993.
2017.PubMed/NCBI
|
|
4
|
Guo YF, Xu NN, Sun W, Zhao Y, Li CY and
Guo MY: Luteolin reduces inflammation in Staphylococcus
aureus-induced mastitis by inhibiting NF-κB activation and MMPs
expression. Oncotarget. 8:28481–28493. 2017.PubMed/NCBI
|
|
5
|
Zang Y, Igarashi K and Li Y: Anti-diabetic
effects of luteolin and luteolin-7-O-glucoside on
KK-Ay mice. Biosci Biotechnol Biochem.
80:1580–1586. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou Y, Ding BZ, Lin YP and Wang HB:
MiR-34a, as a suppressor, enhance the susceptibility of gastric
cancer cell to luteolin by directly targeting HK1. Gene. 644:56–65.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Byun EB, Song HY, Mushtaq S, Kim HM, Kang
JA, Yang MS, Sung NY, Jang BS and Byun EH: Gamma-irradiated
luteolin inhibits 3-isobutyl-1-methylxanthine-induced melanogenesis
through the regulation of CREB/MITF, PI3K/Akt, and ERK pathways in
B16BL6 melanoma cells. J Med Food. 20:812–819. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen P, Zhang JY, Sha BB, Ma YE, Hu T, Ma
YC, Sun H, Shi JX, Dong ZM and Li P: Luteolin inhibits cell
proliferation and induces cell apoptosis via down-regulation of
mitochondrial membrane potential in esophageal carcinoma cells EC1
and KYSE450. Oncotarget. 8:27471–27480. 2017.PubMed/NCBI
|
|
9
|
Wang Q, Wang H, Jia Y, Pan H and Ding H:
Luteolin induces apoptosis by ROS/ER stress and mitochondrial
dysfunction in gliomablastoma. Cancer Chemother Pharmacol.
79:1031–1041. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cao Z, Zhang H, Cai X, Fang W, Chai D, Wen
Y, Chen H, Chu F and Zhang Y: Luteolin promotes cell apoptosis by
inducing autophagy in hepatocellular carcinoma. Cell Physiol
Biochem. 43:1803–1812. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lim W, Yang C, Bazer FW and Song G:
Luteolin inhibits proliferation and induces apoptosis of human
placental choriocarcinoma cells by blocking the PI3K/AKT pathway
and regulating sterol regulatory element binding protein activity.
Biol Reprod. 95:822016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shen XF, Teng Y, Sha KH, Wang XY, Yang XL,
Guo XJ, Ren LB, Wang XY, Li J and Huang N: Dietary flavonoid
luteolin attenuates uropathogenic Escherichia. Coli invasion of the
urinary bladder. Biofactors. 42:674–685. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Krifa M, Leloup L, Ghedira K, Mousli M and
Chekir-Ghedira L: Luteolin induces apoptosis in BE colorectal
cancer cells by downregulating calpain, UHRF1, and DNMT1
expressions. Nutr Cancer. 66:1220–1227. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xavier CP, Lima CF, Preto A, Seruca R,
Fernandes-Ferreira M and Pereira-Wilson C: Luteolin, quercetin and
ursolic acid are potent inhibitors of proliferation and inducers of
apoptosis in both KRAS and BRAF mutated human colorectal cancer
cells. Cancer Lett. 281:162–170. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pandurangan AK, Dharmalingam P, Sadagopan
SK, Ramar M, Munusamy A and Ganapasam S: Luteolin induces growth
arrest in colon cancer cells through involvement of
Wnt/β-catenin/GSK-3β signaling. J Environ Pathol Toxicol Oncol.
32:131–139. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Y, Lang T, Jin B, Chen F, Zhang Y,
Beuerman RW, Zhou L and Zhang Z: Luteolin inhibits colorectal
cancer cell epithelial-to-mesenchymal transition by suppressing
CREB1 expression revealed by comparative proteomics study. J
Proteomics. 161:1–10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Manju V and Nalini N: Protective role of
luteolin in 1,2-dimethylhydrazine induced experimental colon
carcinogenesis. Cell Biochem Funct. 25:189–194. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ashokkumar P and Sudhandiran G: Protective
role of luteolin on the status of lipid peroxidation and
antioxidant defense against azoxymethane-induced experimental colon
carcinogenesis. Biomed Pharmacother. 62:590–597. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chian S, Li YY, Wang XJ and Tang XW:
Luteolin sensitizes two oxaliplatin-resistant colorectal cancer
cell lines to chemotherapeutic drugs via inhibition of the Nrf2
pathway. Asian Pac J Cancer Prev. 15:2911–2916. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qu Q, Qu J, Guo Y, Zhou BT and Zhou HH:
Luteolin potentiates the sensitivity of colorectal cancer cell
lines to oxaliplatin through the PPARγ/OCTN2 pathway. Anticancer
Drugs. 25:1016–1027. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lin S and Gregory RI: MicroRNA biogenesis
pathways in cancer. Nat Rev Cancer. 15:321–333. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bracken CP, Scott HS and Goodall GJ: A
network-biology perspective of microRNA function and dysfunction in
cancer. Nat Rev Genet. 17:719–732. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Eichmuller SB, Osen W, Mandelboim O and
Seliger B: Immune modulatory microRNAs involved in tumor attack and
tumor immune escape. J Natl Cancer Inst. 109:2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rupaimoole R and Slack FJ: MicroRNA
therapeutics: Towards a new era for the management of cancer and
other diseases. Nat Rev Drug Discov. 16:203–222. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Han K, Meng W, Zhang JJ, Zhou Y, Wang YL,
Su Y, Lin SC, Gan ZH, Sun YN and Min DL: Luteolin inhibited
proliferation and induced apoptosis of prostate cancer cells
through miR-301. Onco Targets Ther. 9:3085–3094. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jiang ZQ, Li MH, Qin YM, Jiang HY, Zhang X
and Wu MH: Luteolin inhibits tumorigenesis and induces apoptosis of
non-small cell lung cancer cells via regulation of microRNA-34a-5p.
Int J Mol Sci. 19:E4472018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang YX, Chen YR, Liu SS, Ye YP, Jiao HL,
Wang SY, Xiao ZY, Wei WT, Qiu JF, Liang L, et al: MiR-384 inhibits
human colorectal cancer metastasis by targeting KRAS and CDC42.
Oncotarget. 7:84826–84838. 2016.PubMed/NCBI
|
|
29
|
Kang KA, Piao MJ, Ryu YS, Hyun YJ, Park
JE, Shilnikova K, Zhen AX, Kang HK, Koh YS, Jeong YJ, et al:
Luteolin induces apoptotic cell death via antioxidant activity in
human colon cancer cells. Int J Oncol. 51:1169–1178. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Abdel Hadi L, Di Vito C, Marfia G,
Ferraretto A, Tringali C, Viani P and Riboni L: Sphingosine kinase
2 and ceramide transport as key targets of the natural flavonoid
luteolin to induce apoptosis in colon cancer cells. PLoS One.
10:e01433842015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Muramatsu T: Midkine and pleiotrophin: Two
related proteins involved in development, survival, inflammation
and tumorigenesis. J Biochem. 132:359–371. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yao J, Zhang LL, Huang XM, Li WY and Gao
SG: Pleiotrophin and N-syndecan promote perineural invasion and
tumor progression in an orthotopic mouse model of pancreatic
cancer. World J Gastroenterol. 23:3907–3914. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bai PS, Xia N, Sun H and Kong Y:
Pleiotrophin, a target of miR-384, promotes proliferation,
metastasis and lipogenesis in HBV-related hepatocellular carcinoma.
J Cell Mol Med. 21:3023–3043. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ma J, Kong Y, Nan H, Qu S, Fu X, Jiang L,
Wang W, Guo H, Zhao S, He J and Nan K: Pleiotrophin as a potential
biomarker in breast cancer patients. Clin Chim Acta. 466:6–12.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jee YH, Sadowski SM, Celi FS, Xi L,
Raffeld M, Sacks DB, Remaley AT, Wellstein A, Kebebew E and Baron
J: Increased pleiotrophin concentrations in papillary thyroid
cancer. PLoS One. 11:e01493832016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chang Y, Zuka M, Perez-Pinera P, Astudillo
A, Mortimer J, Berenson JR and Deuel TF: Secretion of pleiotrophin
stimulates breast cancer progression through remodeling of the
tumor microenvironment. Proc Natl Acad Sci USA. 104:10888–10893.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lampropoulou E, Logoviti I, Koutsioumpa M,
Hatziapostolou M, Polytarchou C, Skandalis SS, Hellman U, Fousteris
M, Nikolaropoulos S, Choleva E, et al: Cyclin-dependent kinase 5
mediates pleiotrophin-induced endothelial cell migration. Sci Rep.
8:58932018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shi Y, Ping YF, Zhou W, He ZC, Chen C,
Bian BS, Zhang L, Chen L, Lan X, Zhang XC, et al: Tumour-associated
macrophages secrete pleiotrophin to promote PTPRZ1 signalling in
glioblastoma stem cells for tumour growth. Nat Commun. 8:150802017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yamakawa T, Kurosawa N, Kadomatsu K,
Matsui T, Itoh K, Maeda N, Noda M and Muramatsu T: Levels of
expression of pleiotrophin and protein tyrosine phosphatase zeta
are decreased in human colorectal cancers. Cancer Lett. 135:91–96.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kong Y, Bai PS, Nan KJ, Sun H, Chen NZ and
Qi XG: Pleiotrophin is a potential colorectal cancer prognostic
factor that promotes VEGF expression and induces angiogenesis in
colorectal cancer. Int J Colorectal Dis. 27:287–298. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sun H, He L, Ma L, Lu T, Wei J, Xie K and
Wang X: LncRNA CRNDE promotes cell proliferation, invasion and
migration by competitively binding miR-384 in papillary thyroid
cancer. Oncotarget. 8:110552–110565. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang G, Pan J, Zhang L, Wei Y and Wang C:
Long non-coding RNA CRNDE sponges miR-384 to promote proliferation
and metastasis of pancreatic cancer cells through upregulating
IRS1. Cell Prolif. 50:2017. View Article : Google Scholar :
|
|
43
|
Fan N, Zhang J, Cheng C, Zhang X, Feng J
and Kong R: MicroRNA-384 represses the growth and invasion of
non-small-cell lung cancer by targeting astrocyte elevated
gene-1/Wnt signaling. Biomed Pharmacother. 95:1331–1337. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Song H, Rao Y, Zhang G and Kong X:
MicroRNA-384 inhibits the growth and invasion of renal cell
carcinoma cells by targeting astrocyte elevated gene 1. Oncol Res.
26:457–466. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zheng J, Liu X, Wang P, Xue Y, Ma J, Qu C
and Liu Y: CRNDE promotes malignant progression of glioma by
attenuating miR-384/PIWIL4/STAT3 axis. Mol Ther. 24:1199–1215.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chen Z, Yu C, Zhan L, Pan Y, Chen L and
Sun C: LncRNA CRNDE promotes hepatic carcinoma cell proliferation,
migration and invasion by suppressing miR-384. Am J Cancer Res.
6:2299–2309. 2016.PubMed/NCBI
|