Non-coding RNAs (ncRNAs), which comprise long
non-coding RNAs (lncRNAs), short microRNAs (miRNA/miRs) and
circular RNAs (circRNAs), constitute the majority of total RNAs in
the eukaryotic transcriptome (1,2). CircRNAs
were first identified in viruses in 1970, and subsequently in
eukaryotic cells (3,4). CircRNAs are a relatively large group of
RNAs that form stable closed circles. A major proportion of ncRNAs
and circRNAs are involved in the regulation of transcriptional and
post-transcriptional gene expression (5). CircRNAs are produced from the
back-splicing of intronic and/or exonic RNA (2). They serve a significant role in cancer
development, metastasis and response to treatment (6). The specificity of circRNA in disease
states and the stability of circRNA in body fluids indicate that
they may be used molecular markers in the diagnosis of cancer
(6-9).
A high number of circRNAs have been identified following the
development of sequencing and bioinformatics analysis techniques
(10-12).
In particular, abnormal expressions of circRNAs have been
identified in leukemia. For instance, hsa_circ_0035381,
hsa_circ_0004136 and hsa_circ_0058058 are reportedly upregulated
while hsa_circ_0017446 and hsa_circ_0004277 are downregulated in
acute myeloid leukemia (6). In the
present review, the aim was to explain the role of defined
pathogenic circRNAs derived from leukemia fusion genes [mixed
lineage leukemia-ALL1 fused gene from chromosome 9 (MLL-AF9) and
promyelocytic leukemia-retinoic acid receptor α (PML-RARA)] and
hsa_circ_0004277 in leukemia cells.
CircRNAs are highly diverse, originating from any
region of the genomic subsequences (exon–intron circRNAs, intronic
circRNAs, exonic circRNAs) (2,12-16).
The majority of the circRNAs are generated from one or more excess
exons (17-20).
The exonic circRNAs predominantly reside in the cytoplasm (21,22). They
have a more stable structure than linear RNAs, with 5’-3’
polarities and no polyadenylation tails (10). CircRNAs can be degraded by RNA
exonuclease or ribonuclease (RNase) R (23). CircRNAs are enriched in exosomes
(24).
It is established that circRNAs are relatively
common in eukaryotic transcriptomics and are stable in
intracellularly in the cytoplasm and in the blood (7). CircRNAs can be secreted into body fluids
or developed in exosomes, and as such have emerged as major
biomarkers for cancer diagnosis (8,24,25). It has been demonstrated that the
expression of circRNA is specific, and that the molecules serve as
miRNA sponges to regulate gene expression (5,26-28).
CircRNAs may have biological activities by acting as miRNA sponges
and/or by binding with RNA-binding proteins (RBPs) and translation
peptides (5,24,29,30). In
the study of cancer, an ongoing aim is to identify potential
biomarkers that are differentially expressed between healthy and
cancerous tissues. In this regard, there are studies indicating
that circRNAs are associated with the initiation and development of
cancer (31-33).
CircRNAs are divided into three groups: Exonic
circRNAs, intronic circRNAs and exon-intron circRNAs. Throughout
the formation of all types of circRNAs, characteristic sequences
are preserved. Sequences from the downstream and upstream regions
enable the formation of mature circRNAs followingback-splicing by
covalently linking in the reverse direction (34). While the effect of back-splicing is
less significant for linear RNAs due to their durability and
half-life, circRNAs are present in excess in cells (35). The first back-splicing mechanism works
due to complementary intron matches that serve a role in the
formation of exonic circRNA (17).
CircRNA may interact with other RNA molecules and DNA, including
mRNA and lncRNA (36,37).
It has been shown that circRNAs can bind to miRNA as
RNA sponges, and by influencing miRNA activities, may increase gene
expression and contribute to the development of tumours (5,43-45).
In particular, it has been reported that circRNAs can increase or
suppress the development of cancer genes associated with migration,
differentiation, proliferation and carcinogenesis by suppressing
miRNA species (46). For instance,
among the first circRNAs to be identified, CiRS-7, contains an
miR-7 binding site and acts as an miR-7 sponge, thus reducing the
effect of miR-7 on its target mRNAs (47,48).
Overexpression of ciRS-7 in colorectal cancer cells has been
reported to eliminate the tumor suppressor function of
miR-7(49).Circ-ZNF609 is also a
cancer-associated gene, being observed to inhibit proliferation in
colorectal cancer cells (50).
CircRNAs have been studied in hematopoietic
malignancies in primary patient samples. In bone marrow from
patients, Salzman et al (10)
determined that hyperdiploid B-linage acute lymphoblastic leukemia
(ALL) in childhood could rearrange chromosomal regions using an
RNA-seq method. Furthermore, they conducted research on the
rearrangement in specialized genomic exon regions subject to RNA
splicing. In the ALL patients, they identified hundreds of genes
that could produce circRNAs. Fusion genes encoded by abnormal
chromosomal translocations have been demonstrated to be associated
with malignant haematological diseases. Chromosomal rearrangements
in tumour cells affect ncRNA levels through the formation of
fusion-circRNAs (f-circRNAs) (51).
There are inherent chromosomal translocations in
various types of leukemia. A recurrent translocation is PML-RARA in
patients with acute promyelocytic leukemia (APL) (52). Recently, studies on circRNA expression
in AML have been conducted. Guarnerio et al (6) demonstrated that circRNAs are derived
from the transcription of fusion genes that occur from chromosomal
translocations. Indeed in various studies, circRNAs have been
associated with many cancers, including AML with MLL-AF9 and
promyelocytic leukemia with PML-RARA. Many are established as
f-circRNAs. Abnormal expression of circRNAs may be the result of
distant interrelated introns and introns of translational genes
that contain repeating sequences complementary to each other. Novel
back-splicing events are supposed to support their formation into
abnormal circRNAs (53).
A previous review noted that cancer-associated
fusion genes may cause the expression of f-circRNAs, including the
pro-proliferative, proto-oncogenic f-circPR and f-circM9_1(6). You and Conrad (54) determined differently expressed
f-circRNAs in APL and AML. Such is in accordance with f-circRNAs
being established to serve an active role in haematological events
(53). Li et al (53) reported that hsa_circ_0004277 in AML
patients was downregulated.
Circ-HIPK2 has been indicated to serve a role as a
transcription coactivator in nuclear bodies and is considered to
have important functions in the formation and development of AML
(55,56). Li et al (57) demonstrated that expression of
circ-HIPK2 affected ATRA-induced differentiation of APL cells. In
addition, the expression of circ-HIPK2 was lower in AML cells
compared with APL cells, and overexpression of circ-HIPK2 increased
differentiation in NB4 cells (APL cells with PML-RARA). Therefore,
HIPK2 appeared to be required for differentiation of ATRA-induced
APL cells. This suggests that circ-HIPK2 may be a biomarker in APL
cells.
There are many chromosomal translocations in several
types of leukemia. These chromosomal translocations may occur due
to the instability of the tumour cells’ genome. The abnormal
chromosomal translocations leading to the rearrangement of
non-homologous chromosomes brings two separate genes together,
giving rise to the production of a fusion genome. Chromosomal
translocations can encode such oncogenic fusion proteins in tumor
formation, these proteins may be a cause of cancer (58).
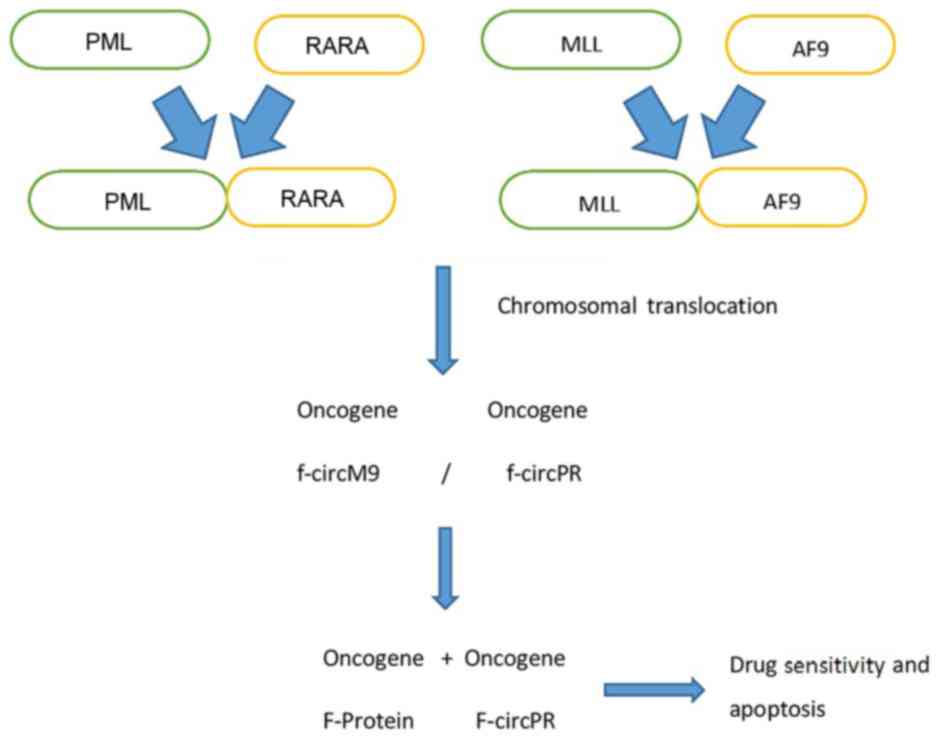
While fusion of circular RNA (f-circRNA) occurs,
gene fusions can also cause defects in mRNA. For example, PML-RARA
and MLL-AF9 genes are fused, and thus form f-circM9 and f-circPR.
Knockout of f-circM9 and f-circPR can increase apoptosis in cancer
cells and increase drug sensitivity to agents such as arsenic
(6). These data indicate that
f-circM9 and f-circPR have a role in haematological malignancies
(Fig. 1). Guarnerio et al (6) documented that expression of f-circRNAs
(f-circPR and f-circM9) in leukemia increased cell proliferation
and clonogenicity. They also showed that the phenotype of cells
with silenced f-circRNA was reversed. Thus, these f-circRNAs may
have pro-proliferative and proto-oncogenic properties in leukemia.
Indeed, it has been documented that f-circRNAs (produced from
PML-RARA and MLL-AF9 fusion genes) have a role in the survival of
leukemia cells (6).
The f-circRNAs expressed in human leukemia cells
have been associated with disease formation (59). However, the f-circRNA does not cause
leukemia alone. Combined expression of F-circM9 and MLL-AF9 fusion
protein have been reported to increase proliferation; notably only
cells expressing the fusion protein exhibited increased colonies.
The expression of f-circRNAs may provide protection for tumour
cells in standard leukemia therapy, including against arsenic
trioxide, and may also provide survival benefit to leukemic cells
during standard leukemia treatment with cytarabine (59).
Hsa_circ_0004277 is located in the region of
chromosome 10: 1125950-1126416. WDR37 is the gene symbol of
hsa_circ_0004277(60). WDR37, located
on the same chromosome, is the linear isoform of
Hsa_circ_0004277(53). This family is
associated with apoptosis, signalling pathways and cellular events
including the cell cycle (60). Wei
et al (53) identified
hsa_circ_0004277 as a target and biomarker in the treatment of AML.
When the circRNA-miRNA-mRNA interaction for hsa_circ_0004277 has
been examined by bioinformatics analysis, it has provided insight
into the underlying mechanism of its function in AML. For instance,
while hsa_circ_0035381, hsa_circ_0004136 and hsa_circ_0058058 are
upregulated, the hsa_circ_0017446 and hsa_circ_0004277 are
downregulated in AML (53). Newly
diagnosed cases of AML have been found to exhibit downregulated
hsa_circ_0004277, while, the expression of hsa_circ_0004277 was
increased in complete remission cases, and the level once again
downregulated in relapsed/refractory cases. These results indicate
that hsa_circ_0004277 may be used as a diagnostic marker and
therapeutic target in AML (53).
In the current review, we sought to summarize the
formation of circRNAs and their characteristics and functional
properties, and the roles of the defined pathogenic circRNAs
derived from the leukemia fusion genes and hsa_circ_0004277 in
leukemia cells. Studies on circRNAs for clinical diagnosis and
treatment are viewed as a guide for translational and clinical
medical developments. Considering the functions and mechanisms of
circRNAs, they may be considered among the main topics of cancer
research. Research on these molecules may provide insight into the
fundamental molecular events involved. The use of circRNAs as
diagnostic biomarkers in cases of cancer metastasis may be
clinically significant. Different cirRNA expression profiles
correlate with clinical features, including tumor stage and
recurrence of metastasis, supported by recent RNA-seq studies
(61,62). Additionally, circRNAs are established
as being markedly more stable than linear RNAs.
Different expression profiles of circRNAs have been
identified in studies on tissues and blood. The expression of
circRNAs is different from that of miRNAs and lncRNAs. By combining
with various biomarkers, it is possible to model for prognosis and
increase the accuracy and specificity of diagnosis (63,64). The
consequences of molecular circuits that control cellular
differentiation in the hematopoietic system are a topic of study.
In haematopoiesis, differentiated cell states can be controlled by
transcriptional circuits linked to each other (65). Recent study of f-circRNAs has revealed
the oncogenic roles of abnormal circRNAs in leukemogenesis
(51). Studies have also shown that
circRNAs may be molecular markers in tumours and affect cell
death.
CircRNAs have potential in targeted cancer therapy,
in that they may be used as a sponge for binding to abnormally
expressed regulatory RNAs and proteins (e.g., RBPs), to thus reduce
oncogenic activities (66). For
instance, certain f-circRNAs have been found to be resistant to
chemotherapy in leukemia patients. The knocking out of f-circM9
expression in leukemia cells can lead to apoptosis. In another
example, while hsa_circ_0004277 showed low expression in newly
diagnosed AML patients, no difference was determined in patients
following treatment. In patients with recurrent disease,
hsa_circ_0004277 has been observed to be downregulated (6). For this reason, hsa_circ_0004277 may be
a diagnostic marker in AML. The overall stability of the circRNAs
may be useful in indicating diseases that can be identified by body
fluids.
In conclusion, considering the functions and
mechanisms of circRNAs, they may be among the main topics in cancer
research. In particular, circRNAs may be a diagnostic marker for
leukemia. An improved understanding of circRNA biology may provide
a guide for novel therapeutic targets.
Not applicable.
Not applicable.
Not applicable.
TBO and CG searched the related literature. TBO
wrote the manuscript. All authors discussed the content and
contributed to the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Warner JR: The economics of ribosome
biosynthesis in yeast. Trends Biochem Sci. 24:437–440.
1999.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Jeck WR, Sorrentino JA, Wang K, Slevin MK,
Burd CE, LiuJ , et al: Circular RNAs are abundant, conserved,
and associated with ALU repeats. RNA. 19:141–157. 2013.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Sanger HL, Klotz G, Riesner D, Gross HJ
and Kleinschmidt AK: Viroids are single-stranded covalently closed
circular RNA molecules existing as highly base-paired rod-like
structures. Proc Natl Acad Sci USA. 73:3852–3856. 1976.PubMed/NCBI
|
|
4
|
Hsu MT and Coca-Prados M: Electron
microscopic evidence for the circular form of RNA in the cytoplasm
of eukaryotic cells. Nature. 280:339–340. 1979.PubMed/NCBI
|
|
5
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Guarnerio J, Bezzi M, Jeong JC, Paffenholz
SV, Berry K, Naldini MM, Lo-Coco F, Tay Y, Beck AH and Pandolfi PP:
Oncogenic role of fusion-circRNAs derived from cancer-associated
chromosomal translocations. Cell:165:289–302. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Memczak S, Papavasileiou P, Peters O and
Rajewsky N: Identification and characterization of circular RNAs as
a new class of putative biomarkers in human blood. PLoS One.
10:(e0141214). 2015.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Bahn JH, Zhang Q, Li F, Chan TM, Lin X,
Kim Y, Wong DT and Xiao X: The landscape of microRNA,
Piwi-interacting RNA, and circular RNA in human saliva. Clin Chem.
61:221–230. 2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Szabo L, Morey R, Palpant NJ, Wang PL,
Afari N, Jiang C, Parast MM, Murry CE, Laurent LC and Salzman J:
Statistically based splicing detection reveals neural enrichment
and tissue-specific induction of circular RNA during human fetal
development. Genome Biol. 16(126)2015.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Salzman J, Gawad C, Wang PL, Lacayo N and
Brown PO: Circular RNAs are the predominant transcript isoform from
hundreds of human genes in diverse cell types. PLoS One.
7:(e30733). 2012.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Guo JU, Agarwal V, Guo H and Bartel DP:
Expanded identification and characterization of mammalian circular
RNAs. Genome Biol. 15(409)2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M and et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zhang Y, Zhang XO, Chen T, Xiang JF, Yin
QF, Xing YH, Zhu S, Yang L and Chen LL: Circular intronic long
noncoding RNAs. Mol Cell. 51:792–806. 2013.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Li Z, Huang C, Bao C, Chen L, Lin M, Wang
X, Zhong G, Yu B, Hu W, Dai L and et al: Exon-intron circular RNAs
regulate transcription in the nucleus. Nat Struct Mol Biol.
22:256–264. 2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Shen T, Han M, Wei G and Ni T: An
intriguing RNA species - perspectives of circularized RNA. Protein
Cell. 6:871–880. 2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Chen LL and Yang L: Regulation of circRNA
biogenesis. RNA Biol. 12:381–388. 2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zhang XO, Wang HB, Zhang Y, Lu X, Chen LL
and Yang L: Complementary sequence-mediated exon circularization.
Cell. 159:134–147. 2014.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Chen I, Chen CY and Chuang TJ: Biogenesis,
identification, and function of exonic circular RNAs. Wiley
Interdiscip Rev RNA. 6:563–579. 2015.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Starke S, Jost I, Rossbach O, Schneider T,
Schreiner S, Hung LH and Bindereif A: Exon circularization requires
canonical splice signals. Cell Reports. 10:103–111. 2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Suzuki H, Aoki Y, Kameyama T, Saito T,
Masuda S, Tanihata J, Nagata T, Mayeda A, Takeda S and Tsukahara T:
Endogenous multiple exon skipping and back-splicing at the DMD
mutation hotspot. Int J Mol Sci. 17(E1722)2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Salzman J, Chen RE, Olsen MN, Wang PL and
Brown PO: Cell-type specific features of circular RNA expression.
PLoS Genet. 9(e1003777)2013.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Westholm JO, Miura P, Olson S, Shenker S,
Joseph B, Sanfilippo P, Celniker SE, Graveley BR and Lai EC:
Genome-wide analysis of drosophila circular RNAs reveals their
structural and sequence properties and age-dependent neural
accumulation. Cell Reports. 9:1966–1980. 2014.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Suzuki H, Zuo Y, Wang J, Zhang MQ,
Malhotra A and Mayeda A: Characterization of RNase R-digested
cellular RNA source that consists of lariat and circular RNAs from
pre-mRNA splicing. Nucleic Acids Res. 34(e63)2006.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Li Y, Zheng Q, Bao C, Li S, Guo W, Zhao J,
Chen D, Gu J, He X and Huang S: Circular RNA is enriched and stable
in exosomes: A promising biomarker for cancer diagnosis. Cell Res.
25:981–984. 2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Galasso M, Costantino G, Pasquali L,
Minotti L, Baldassari F, Corrà F, Agnoletto C and Volinia S:
Profiling of the predicted circular RNAs in ductal in situ and
invasive breast cancer: A pilot study. Int J Genomics.
2016(4503840)2016.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Wang PL, Bao Y, Yee MC, Barrett SP, Hogan
GJ, Olsen MN, Dinneny JR, Brown PO and Salzman J: Circular RNA is
expressed across the eukaryotic tree of life. PLoS One.
9(e90859)2014.(CrossRef) (PubMed). PubMed/NCBI View Article : Google Scholar
|
|
27
|
Rybak-Wolf A, Stottmeister C, Glažar P,
Jens M, Pino N, Giusti S, Hanan M, Behm M, Bartok O, Ashwal-Fluss R
and et al: Circular RNAs in the mammalian brain are highly
abundant, conserved, and dynamically expressed. Mol Cell.
58:870–885. 2015.(CrossRef) (PubMed). PubMed/NCBI View Article : Google Scholar
|
|
28
|
Zheng Q, Bao C, Guo W, Li S, Chen J, Chen
B, Luo Y, Lyu D, Li Y, Shi G and et al: Circular RNA profiling
reveals an abundant circHIPK3 that regulates cell growth by
sponging multiple miRNAs. Nat Commun. 7(11215)2016.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Granados-Riveron JT and Aquino-Jarquin G:
The complexity of the translation ability of circRNAs. Biochim
Biophys Acta. 1859:1245–1251. 2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Wang Y and Wang Z: Efficient backsplicing
produces translatable circular mRNAs. RNA. 21:172–179.
2015.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Dong Y, He D, Peng Z, Peng W, Shi W, Wang
J, Li B, Zhang C and Duan C: Circular RNAs in cancer: An emerging
key player. J Hematol Oncol. 10(2)2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Qu S, Zhong Y, Shang R, Zhang X, Song W,
Kjems J and Li H: The emerging landscape of circular RNA in life
processes. RNA Biol. 14:992–999. 2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Xin Z, Ma Q, Ren S, Wang G and Li F: The
understanding of circular RNAs as special triggers in
carcinogenesis. Brief Funct Genomics. 16:80–86. 2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Vicens Q and Westhof E: Biogenesis of
Circular RNAs. Cell. 159:13–14. 2014.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Qu S, Yang X, Li X, Wang J, Gao Y, Shang
R, Sun W, Dou K and Li H: Circular RNA: A new star of noncoding
RNAs. Cancer Lett. 365:141–148. 2015.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Chen CY and Sarnow P: Initiation of
protein synthesis by the eukaryotic translational apparatus on
circular RNAs. Science. 268:415–417. 1995.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Ashwal-Fluss R, Meyer M, Pamudurti NR,
Ivanov A, Bartok O, Hanan M, Evantal N, Memczak S, Rajewsky N and
Kadener S: circRNA biogenesis competes with pre-mRNA splicing. Mol
Cell. 56:55–66. 2014.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Rinn JL and Chang HY: Genome regulation by
long noncoding RNAs. Annu Rev Biochem. 81:145–166. 2012.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009.PubMed/NCBI View
Article : Google Scholar
|
|
40
|
Perriman R and Ares M Jr: Circular mRNA
can direct translation of extremely long repeating-sequence
proteins in vivo. RNA. 4:1047–1054. 1998.PubMed/NCBI
|
|
41
|
Zhang XO, Dong R, Zhang Y, Zhang JL, Luo
Z, Zhang J, Chen LL and Yang L: Diverse alternative back-splicing
and alternative splicing landscape of circular RNAs. Genome Res.
26:1277–1287. 2016.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Hentze MW and Preiss T: Circular RNAs:
Splicing's enigma variations. EMBO J. 32:923–925. 2013.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Ebert MS and Sharp PA: MicroRNA sponges:
Progress and possibilities. RNA. 16:2043–2050. 2010.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Ebert MS and Sharp PA: Emerging roles for
natural microRNA sponges. Curr Biol. 20:R858–R861. 2010.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Chen LL: The biogenesis and emerging roles
of circular RNAs. Nat Rev Mol Cell Biol. 17:205–211.
2016.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Conn SJ, Pillman KA, Toubia J, Conn VM,
Salmanidis M, Phillips CA, Roslan S, Schreiber AW, Gregory PA and
Goodall GJ: The RNA binding protein quaking regulates formation of
circRNAs. Cell. 160:1125–1134. 2015.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Hansen TB, Kjems J and Damgaard CK:
Circular RNA and miR-7 in cancer. Cancer Res. 73:5609–5612.
2013.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Junn E, Lee KW, Jeong BS, Chan TW, Im JY
and Mouradian MM: Repression of alpha-synuclein expression and
toxicity by microRNA-7. Proc Natl Acad Sci USA. 106:13052–13057.
2009.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Weng W, Wei Q, Toden S, Yoshida K,
Nagasaka T, Fujiwara T, Cai S and et al: Circular RNA ciRS-7-A
Promising Prognostic Biomarker and a Potential Therapeutic Target
in Colorectal Cancer. Clin Cancer Res. 23:3918–3928.
2017.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Wu L, Xia J, Yang J, Shi Y, Xia H, Xiang X
and et al: Circ-ZNF609 promotes migration of colorectal cancer by
inhibiting Gli1 expression via microRNA-150. J BUON. 23:1343–1349.
2018.PubMed/NCBI
|
|
51
|
Bonizzato A, Gaffo E and Te Kronnie G and
Bortoluzzi S: CircRNAs in hematopoiesis and hematological
malignancies. Blood Cancer J. 6(e483)2016.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Dos Santos GA, Kats L and Pandolfi PP:
Synergy against PML-RARa: Targeting transcription, proteolysis,
differentiation, and self-renewal in acute promyelocytic leukemia.
J Exp Med. 210:2793–2802. 2013.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Li W, Zhong C, Jiao J, Li P, Cui B, Ji C
and Ma D: Characterization of hsa_circ_0004277 as a new biomarker
for acute myeloid leukemia via circular RNA profile and
bioinformatics analysis. Int J Mol Sci. 18(597)2017.PubMed/NCBI View Article : Google Scholar
|
|
54
|
You X and Conrad TO: Acfs: Accurate
circRNA identification and quantification from RNA-Seq data. Sci
Rep. 6(38820)2016.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Li XL, Arai Y, Harada H, Shima Y, Yoshida
H, Rokudai S, Aikawa Y, Kimura A and Kitabayashi I: Mutations of
the HIPK2 gene in acute myeloid leukemia and myelodysplastic
syndrome impair AML1- and p53-mediated transcription. Oncogene.
26:7231–7239. 2007.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Shima Y, Honma Y and Kitabayashi I:
PML-RARα and its phosphorylation regulate pml oligomerization and
HIPK2 stability. Cancer Res. 73:4278–4288. 2013.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Li S, Ma Y, Tan Y, Ma X, Zhao M, Chen B,
Zhang R, Chen Z and Wang K: Profiling and functional analysis of
circular RNAs in acute promyelocytic leukemia and their dynamic
regulation during all-trans retinoic acid treatment. Cell Death
Dis. 9(651)2018.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Greuber EK, Smith-Pearson P, Wang J and
Pendergast AM: Role of ABL family kinases in cancer: From leukaemia
to solid tumours. Nat Rev Cancer. 13:559–571. 2013.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Greene J, Baird AM, Brady L, Lim M, Gray
SG, McDermott R and Finn SP: Circular RNAs: Biogenesis, Function
and Role in Human Diseases. Front Mol Biosci. 4(38)2017.PubMed/NCBI View Article : Google Scholar
|
|
60
|
http://www.circbase.org/cgi-bin/singlerecord.cgi?id=hsa_circ_0004277.
PubMed/NCBI View Article : Google Scholar
|
|
61
|
Xu L, Zhang M, Zheng X, Yi P, Lan C and Xu
M: The circular RNA ciRS-7 (Cdr1as) acts as a risk factor of
hepatic microvascular invasion in hepatocellular carcinoma. J
Cancer Res Clin Oncol. 143:17–27. 2017.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Zhang Y, Li J, Yu J, Liu H, Shen Z, Ye G,
Mou T, Qi X and Li G: Circular RNAs signature predicts the early
recurrence of stage III gastric cancer after radical surgery.
Oncotarget. 8:22936–22943. 2017.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Gong Z, Zhang S, Zhang W, Huang H, Li Q,
Deng H, Ma J, Zhou M, Xiang J, Wu M and et al: Long non-coding RNAs
in cancer. Sci China Life Sci. 55:1120–1124. 2012.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Huang M, Zhong Z, Lv M, Shu J, Tian Q and
Chen J: Comprehensive analysis of differentially expressed profiles
of lncRNAs and circRNAs with associated co-expression and ceRNA
networks in bladder carcinoma. Oncotarget. 7:47186–47200.
2016.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Novershtern N, Subramanian A, Lawton LN,
Mak RH, Haining WN, McConkey ME, Habib N, Yosef N, Chang CY, Shay T
and et al: Densely interconnected transcriptional circuits control
cell states in human hematopoiesis. Cell. 144:296–309.
2011.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Bachmayr-Heyda A, Reiner AT, Auer K,
Sukhbaatar N, Aust S, Bachleitner-Hofmann T, Mesteri I, Grunt TW,
Zeillinger R and Pils D: Correlation of circular RNA abundance with
proliferation - exemplified with colorectal and ovarian cancer,
idiopathic lung fibrosis, and normal human tissues. Sci Rep.
5(8057)2015.PubMed/NCBI View Article : Google Scholar
|