Introduction
Gliomas are the most common types of tumors of the
central nervous system and are resistant to numerous treatments,
including radiation, chemotherapy or other adjuvant therapies.
Although considerable progress has been made in the treatment of
glioma, its prognosis remains poor (1). A new modality is urgently required in
order to combat gliomas. Sonodynamic therapy (SDT), a combination
of sonosensitizers and ultrasound irradiations, has been considered
to be an effective method owing to increasing
sonosensitizer-derived radicals. SDT is capable of effectively
enhancing the cytotoxicity of sonosensitizers, which preferentially
accumulate in the tumor site with minimal damage of peripheral
healthy tissues. Thus, this would be a valuable cancer therapy
modality (2,3).
The sonosensitizer is the key factor of SDT.
Photosensitizers have been used in SDT as well as in photodynamic
therapy (PDT). Certain drugs, including hematoporphyrin, photofrin
II, ATX-70 and ATX-S10, have been demonstrated to induce cell
killing when activated by ultrasound exposure. It has previously
been indicated that these chemicals, originally generated in PDT,
were therefore applicable as sonosensitizers for tumor treatment in
combination with ultrasound. In the present preliminary study, we
used a new domestic photosensitizer, di-sulfo-di-phthalimidomethyl
phthalolcyanine zinc (ZnPcS2P2) (4–6). The
new photosensitizer (wavelength 670 nm) has been studied in
choroidal neovascularization and bone marrow purging for PDT
(7).
ZnPcS2P2 has certain advantages over other
conventional photosensitizers, such as porphyrin derivatives
(8), for example, the exicited
triplet state of ZnPcS2P2 has a larger
quantum yield, longer lifetime and the amphipathic structure of
ZnPcS2P2 results in the increase of selective
uptake of sensitizer by tumor cells. Moreover, a repeated-dose
toxicity study of ZnPcS2P2-based PDT in
beagle dogs demonstrated it to be a safe and promising approach in
the clinic, as this drug is eliminated more quickly from the body
with the result of decreased skin phototoxicity from natural light.
It has been demonstrated that ZnPcS2P2 may be
a good sensitizer for use in SDT.
As we know, apoptosis is often initiated by an
extrinsic (activated caspase-8) or an intrinsic pathway (activated
caspase-9) (9). The extrinsic
pathway functions are capable of directly activating caspase-8 via
the death receptors on the cell surface; nevertheless, the
intrinsic pathway regulates the activation of caspase-9 and,
subsequently, the activation of caspase-3. In the present study, we
aimed to evaluate the effects of killing glioma cells and
indentified the intrinsic or extrinsic apoptosis pathways of the
newly established sonosensitizer by SDT in an in vitro
model.
Materials and methods
Sonosensitizer
ZnPcS2P2 was a gift from the
Department of Chemistry, Institute of Research on Functional
Materials, Fuzhou University (China). Its chemical structure is
shown in Fig. 1.
ZnPcS2P2 is an odorless, cyan liquid that is
insoluble in water. The liquid has been identified to have a
chemical purity of >95.0% via gas chromatography and infrared
spectroscopy. ZnPcS2P2 was dissolved in a
solution comprising Cremophor EL 2% (V/V), propylene glycol 20%
(V/V), NaCl 0.9% (W/W) and was stored in the dark at 4°C. These
dosing solutions were prepared immediately prior to use.
Cell culture
U251 human glioma cells were obtained from the
Shanghai institute of Cytobiology (Institute of Chinese Academic
Medical Scinence, China) and were continuously cultured in DMEM
(Gibco BRL; Carlsbad, CA, USA), supplemented with 10% fetal bovine
serum (FBS), 100 U/ml penicillin, 100 μg/ml streptomycin and 0.25
mmol/l L-glutamine in a humidified incubator at 37°C and 5%
CO2. Cells in the exponential growth phase were used for
all experiments.
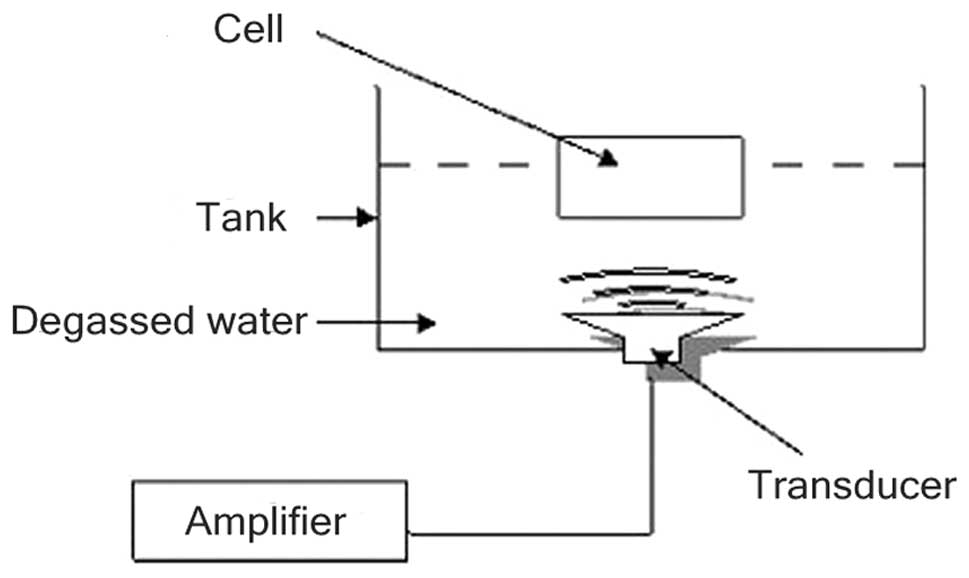
Setup of sonication experiment
A multi-function physical therapy ultrasound device
(Tianshi Technologies Ltd. Co., Beijing, China) was used to
generate ultrasound at 1 MHz. Ultrasonic intensities (0.5
W/cm2) were measured by a stainless-steel ball
radiometer (diameter 0.32 cm) (10). In this study, the ultrasound
transducer was placed in a 37°C water bath filled with degassed
water and stained with black ink (Fig.
2).
Reagents and antibodies
The monoclonal anti-β-actin antibody and
anti-caspase-3, -8 and -9 antibodies were obtained from
Sigma-Aldrich (St. Louis, MO, USA). Annexin-V-FITC Apoptosis
Detection Kit was obtained from BD Biosciences (Franklin Lakes, NJ,
USA). 2,7-dichlorodihydrofluorescein diacetate (DCFH-DA) was
obtained from Calbiochem (La Jolla, CA, USA). All other chemicals
and cell-culture reagents were purchased from Laohekou Jing hong
Chemical Co. Ltd. (China). All solvents used in chemical reactions
were anhydrous and obtained as such from commercial sources. All
other reagents were used as supplied, unless otherwise stated.
Selection of drug conditions for SDT
U251 human cells were transferred to 6 wells in the
centre of a 96-well plate at a density of 8×103
cells/well and in the same way, transferred to 36 sections of a
96-well plate. The following day, each of the 6 plates were changed
by DMEM with various final concentrations of
ZnPcS2P2 (0.625, 1.250, 2.500, 5.000, 10.000
and 20.000 μg/ml), cells were continuously cultured in a humidified
incubator at 37°C and 5% CO2 for 4 h, then subjected to
ultrasound treatment at 1.0 MHz and 0.5 W/cm2 for 2 min.
Cells were re-incubated for up to 4 h and subjected to MTT
assay.
Similarly, cells were transferred to 36 sections of
a 96-well plate, were re-incubated with a final concentration of
ZnPcS2P2 5.0 μg/ml for (1, 2, 3, 4, 5 and 6
h) and then underwent ultra-sound treatment. Cells were
subsequently evaluated by MTT assay following re-incubation for 4
h.
Cells of 36 sections of a 96-well plate (with a
final concentration of ZnPcS2P2 5.0 μg/ml)
were incubated for up to 4 h, followed by ultrasound treatment, and
subjected to MTT assay at various time-points post-SDT (3, 6, 12,
24, 48 and 96 h).
The growth inhibition rate was calculated as:
inhibition rate (%) = (1-OD treatment group/OD control group) x
100%.
Examination of the SDT effect on the
growth of human U251 glioma cells
In the control group, cells were neither treated
with ZnPcS2P2 nor with ultrasound. In the
ZnPcS2P2 group, cells were treated with
ZnPcS2P2 alone (final concentration 5.0
μg/ml, incubation time 4 h). In the ultrasound group, cells were
treated with ultrasound alone. In the SDT group, cells were treated
with ZnPcS2P2 (5.0 μg/ml, incubation time 4
h) and then subjected to ultrasound treatment at 1.0 MHz and 0.5
W/cm2 for 2 min. Cells were re-incubated for up to 6 h
and subjected to MTT assay following treatment.
Examination of SDT-induced apoptosis by
flow cytometry
Following treatment, cells were re-incubated for up
to 6 h, 4 groups of cells (control, ZnPcS2P2
alone, ultrasound alone and ZnPcS2P2 +
ultrasound) were re-suspended in a concentration of
5×106 cells/ml and washed twice with pre-cooling of the
phosphate-buffered saline (PBS). Following washing with Annexin-V
binding buffer, cells were stained with FITC-conjugated Annexin-V
and propidium iodide (PI) reagents for 15 min as per the
manufacturer’s instructions and further examined by flow cytometry
(FACScalibur; Becton-Dickinson). The percentages of dead cells and
those undergoing apoptosis were analyzed using Cell Quest
Software.
Transmission electron microscope
examination of subcellular structure
For the electron microscope examination, cells from
each group were re-incubated for up to 6 h and then washed twice
with PBS and fixed in a mixture of 2.5% glutaraldehyde and osmium
tetroxide. Cells were then dehydrated in progressive 10-min steps
for 3 times in ethanol/water (70, 90 and 96%, respectively),
followed by isoamyl acetate. Finally, cells were stained with
uranyl acetate (Guangzhou Chemical Reagent Factory, China) and lead
citrate (Guangzhou Chemical Reagent Factory). Ultra-thin sections
were examined under a transmission electron microscope (JEM-1220,
Japan).
Western blot analysis
To examine protein expression, cells of various
groups were separately washed, collected and homogenized in a lysis
buffer (10 mM Tris-HCl, pH 8.0, 0.32 mM sucrose, 5 mM EDTA, 2 mM
DTT, 1 mM phenylmethyl sulfonylfluoride and 1% Triton X-100),
followed by centrifugation. Proteins in various groups were
separately electrophoresed on sodium dodecyl sulfate (SDS)
polyacrylamide gel (12%), the gel-separated proteins were
transferred to nitropure nitrocellulose membranes (Santa Cruz
Biotechnology, CA, USA), and the membranes were probed overnight at
4°C with primary antibodies. Each of the targeted proteins was
immunostained by anti-β-actin and anti-caspases-3, -8 and -9 (each
1:1000) antibodies. Following probing, the membranes were washed 3
times and incubated for 1 h at room temperature with the respective
alkaline phosphatase-conjugated secondary antibodies (Sigma) prior
to visualization using a chemiluminescence detection kit
(Sigma).
Measurement production of intracellular
reactive oxygen species (ROS)
The production of intracellular ROS was assayed
spectrophotometrically with dichlorofluorescin diacetate (DCFH-DA)
as described previously (11).
Cells with or without treatment were harvested, washed and
re-suspended in 500 μl PBS containing DCFH-DA (final concentration,
5 μmol/l), and incubated at 37°C in the dark for 30 min. the
concentration of ROS was monitored spectrophotometrically (F-2000
Hitachi; Hitachi Corp. Tokyo, Japan) at 534 nm following excitation
at 488 nm.
Statistical analysis
Statistical evaluation was performed using the least
significant difference t-test, Dunnett test or paired t-test using
the SPSS 11.0 Software system. Data are presented as the means ±
standard error (SE). P<0.05 was considered to indicate a
statistically significant difference.
Results
Selection of drug conditions and analysis
of growth inhibition rate in SDT by MTT
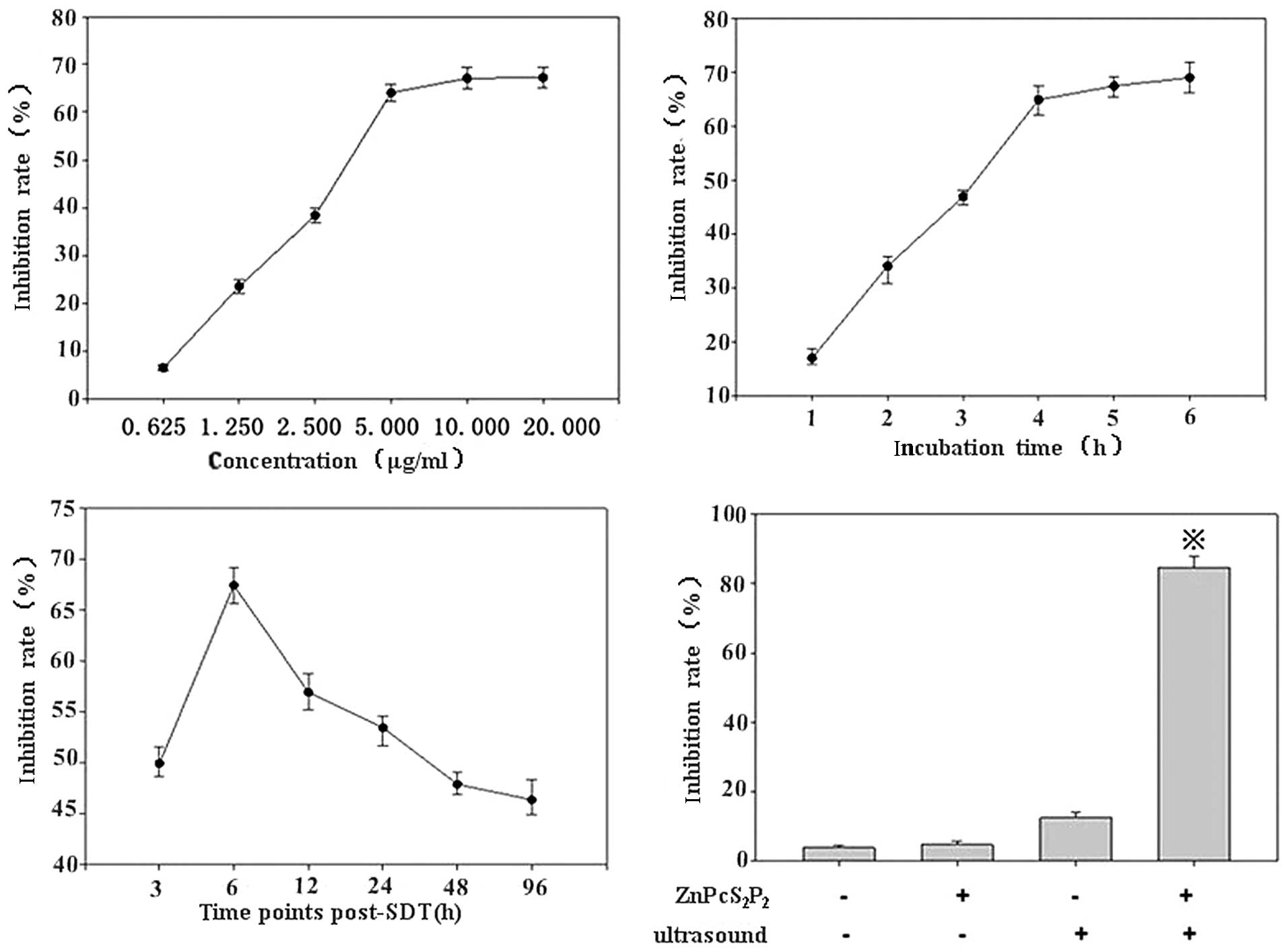
A MTT assay revealed that when the concentration of
ZnPcS2P2 was >5.0 μg/ml, the growth
inhibition rate was less concentration-dependent (Fig. 3A) and that when the incubation time
of ZnPcS2P2 was >4 h the growth inhibition
rate was less incubation time-dependent (Fig. 3B). Conversely, the MTT assay
revealed that growth inhibition occurred in a concentration- and
incubation time-dependent manner when the concentration of
ZnPcS2P2 was <5.0 μg/ml or the incubation
time was <4 h. In addition, the growth inhibition rate did not
significantly increase when re-incubation time was beyond 6 h
following SDT. Therefore, when the incubation time was 4 h, and the
concentration of ZnPcS2P2 was 5.0 μg/ ml, 6 h
following SDT, the inhibition rate was highly notable (Fig. 3A–C). Thus, we selected the 5.0
μg/ml concentration of ZnPcS2P2, the 4-h
incubation time and at 6 h post-SDT to study the
ZnPcS2P2-mediated SDT effects on U251 glioma
cells.
Examination of the SDT effect on the
growth of U251 human glioma cells
SDT will be most effective when the
ZnPcS2P2 concentration in the glioma cells is
at its maximum. As shown in Fig.
3D, when the concentration of ZnPcS2P2
was 5.0 μg/ml and the incubation time was 4 h, the synergistic
effect between ZnPcS2P2 and ultrasound
exposure on cell growth inhibition was evident. The growth
inhibition rate of the 4 groups of the cells were determined by MTT
assay post-treatment, the inhibition rate in the SDT group was
significantly higher than in the other 3 groups (p<0.05), the
inhibition rate was at its peak (87.86±2.45%) 6 h following SDT
(Fig. 3D).
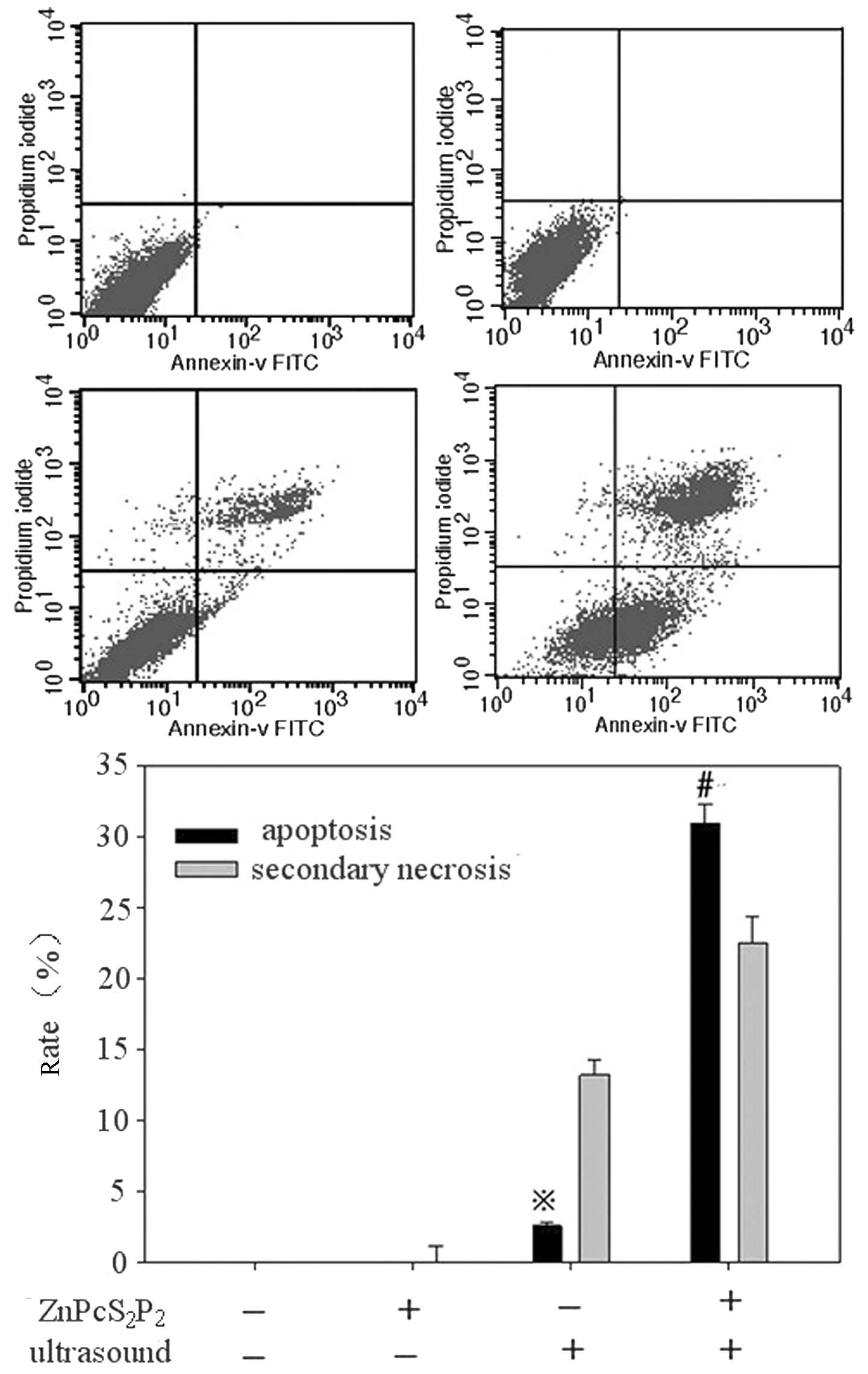
Apoptosis assessment by flow
cytometry
The percentage of apoptotic cells was calculated by
flow cytometry using the Annexin-V/PI double-staining assay
(Fig. 4A–D).
ZnPcS2P2 treatment alone showed no
significant apoptotic effect (0.03±0.01 vs. 0.15±0.01%, p>0.05),
ultrasound treatment displayed a slight apoptotic effect (4.61±0.24
vs. 0.03±0.01%, p<0.05) and SDT treatment displayed a clear
apoptotic effect (30.93±1.34 vs. 0.03±0.01%, p<0.01).
Fine structure changes
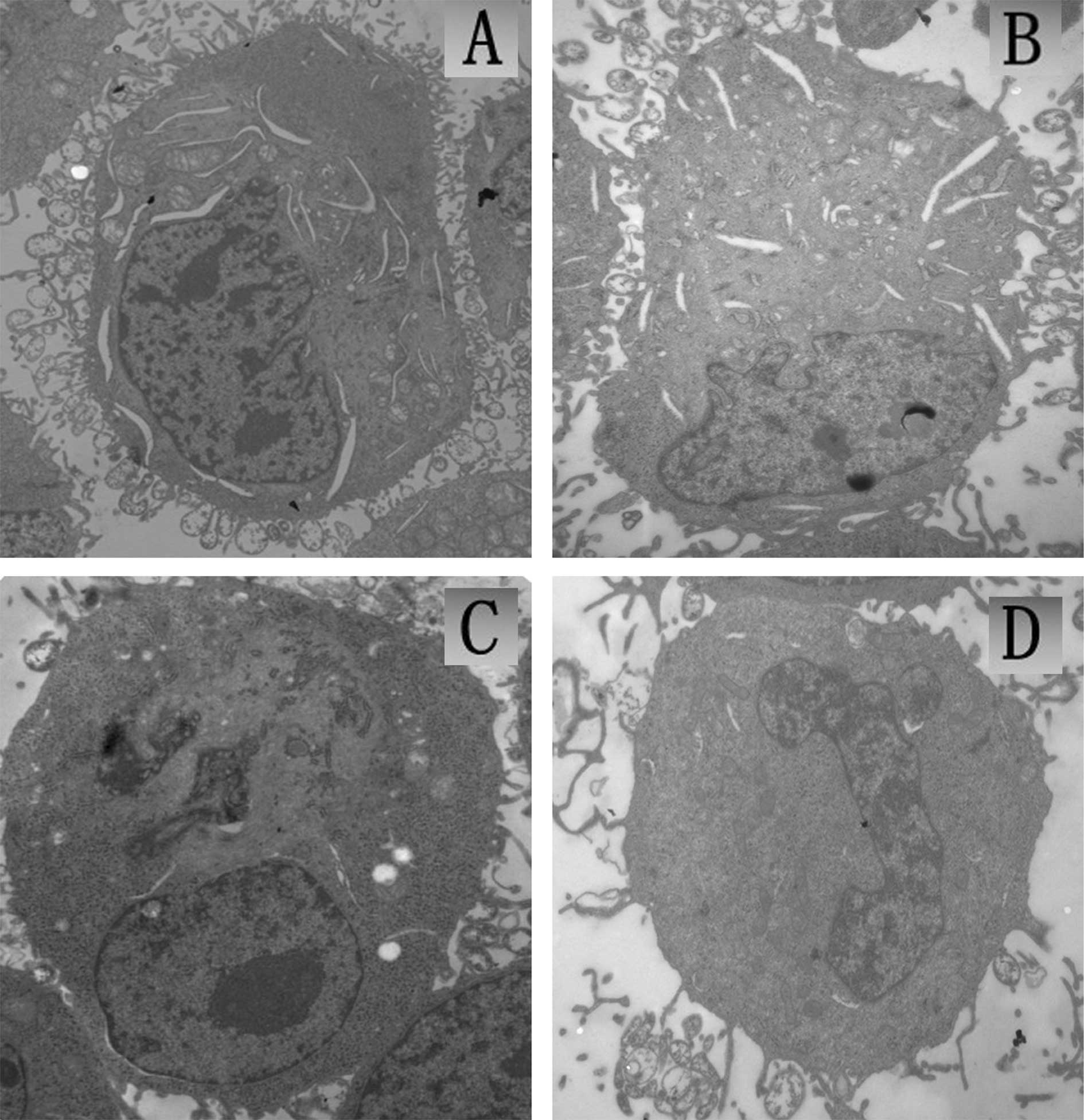
The TEM observation of U251 tumor cells immediately
following SDT treatment is shown in Fig. 5. The cells in the control group
were intact with a rich cytoplasm, mitochondria were integrated,
cell membranes and the nuclear envelope were intact and nuclear
materials were dense (Fig. 5A).
The morphology of cells in the ZnPcS2P2 group
was similar to that in the control group, cell membranes remained
intact with a rich cytoplasm (Fig.
5B). In the ultrasound group, a small number of cells displayed
limited microvilli, cavitation bubbles, slightly condensed
chromatin and presented characteristics of early stage apoptosis
(Fig. 5C). In the SDT group, a
portion of cell microvilli had vanished, the volume of the cell
nucleus had decreased and chromatin was gathered densely, thus
presenting characteristics of apoptosis (Fig. 5D).
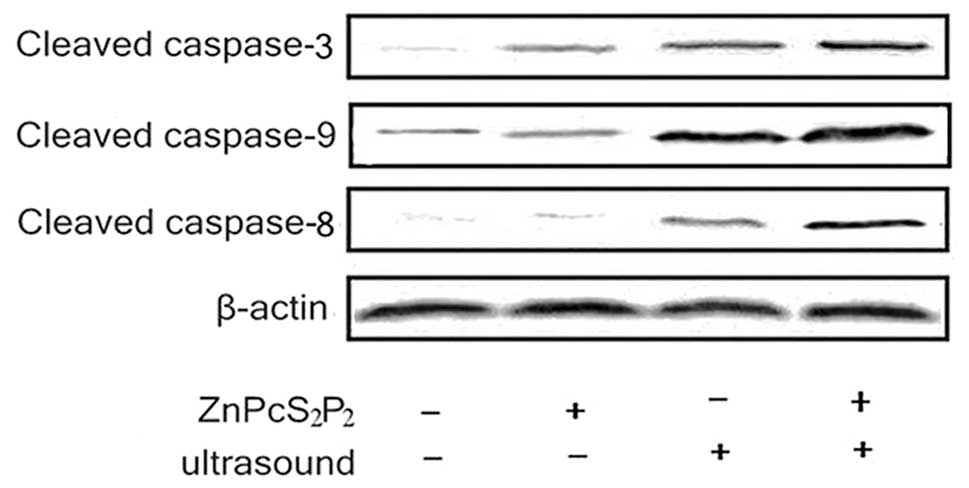
Analysis of caspase expressions
The SDT-induced apoptotic pathway was investigated
by Western blot analysis of caspase expressions. Caspases-8, -9 and
-3 are key factors in the extrinsic and intrinsic apoptosis
pathway, thus, we measured the activities of these caspases. As
shown in Fig. 7, caspases-3, -8
and -9 were significantly activated following ultrasound treatment
with ZnPcS2P2. The expression of activated
caspases-8, -9 and -3 significantly increased in the SDT group
(Fig. 6)
Intracellular ROS
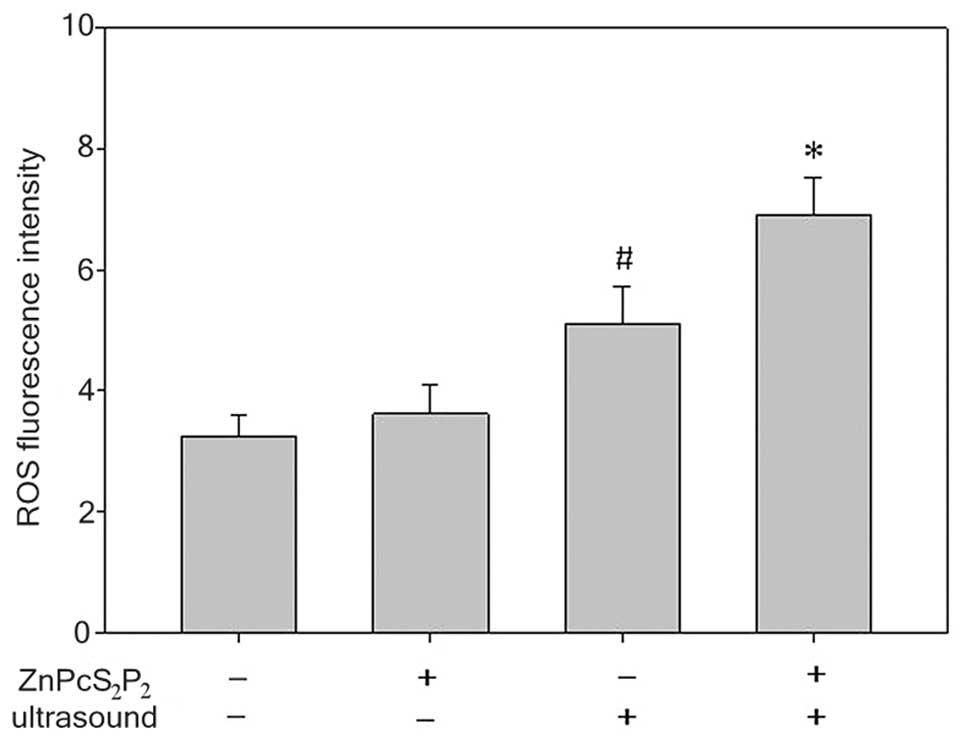
The production of ROS was determined by a
fluorescence spectrophotometer with DCFH-DA (Fig. 7). The production of ROS in the SDT
group significantly increased compared to the other 3 groups
(p<0.05). In the ultrasound group, the production of ROS
slightly increased compared to the ZnPcS2P2
group and the control group (p<0.05). However, there was no
significant difference in ROS production between the
ZnPcS2P2 and the control groups (p>0.05).
The results indicate that the synergistic effect of ultrasound and
ZnPcS2P2 clearly enhances ROS production.
Discussion
Sonodynamic therapy (SDT) is the foundation on which
ultrasound is capable of activating the sonosensitizer in order to
generate cytotoxic substances and further kill tumor cells. Owing
to the fact that the sonosensitizer has high accumulation in tumor
tissue, SDT can not only maximally kill tumor cells, but also
protect normal brain tissue from impairment (12).
Until now, the majority of sonosensitizers in SDT
are not chemosynthetic compounds. In the present experiment,
ZnPcS2P2, a porphyrin compound created by
artificial chemosynthesis that has stable physical and chemical
properties, was used. In a previous study,
ZnPcS2P2-mediated PDT improved the effects of
killing leukemia HL60 and K562 cells (7). In this study, we aimed to observe the
effects and mechanism of SDT with
ZnPcS2P2.
In order to evaluate the effects of
ZnPcS2P2-mediated SDT, we first aimed to
pinpoint optimal concentrations and incubation times of
ZnPcS2P2 in order to ensure the best result
of ZnPcS2P2-mediated, SDT-induced U251 glioma
cell death by MTT assay. Our experimental results indicated that
the inhibition rate of tumor cells increased gradually with the
elevation of drug concentration to some extent. When the
concentration of the drug was ≥5.0 μg/ml, the inhibition rate of
the tumor did not significantly increase (Fig. 3A). A similar phenomenon presented
in the screening of the incubation time. The inhibation rate of the
tumor had a close correlation with the incubation time to some
extent. However, when the incubation time was ≥4 h, the inhibition
rate of the tumor also did not significantly increase (Fig. 3B). In addition, the growth
inhibition rate did not significantly increase when re-incubation
time exceeded 6 h post-SDT. This phenomenon may be explained by the
fact that ZnPcS2P2 was slowly absorbed by the
U251 glioma cells, and when the glioma cells achieved maximum
saturation, the inhibition rate of the glioma cells plateaued.
(Fig. 3A and B). We observed
various inhibition rates of the tumor cells in the 4 groups
(control, ZnPcS2P2 alone, ultrasound alone
and SDT). The results revealed that the inhibition rate of the
tumor cells in the SDT group was significantly higher than in the
other 3 groups at 6 h. This shows that the ultrasound activation of
ZnPcS2P2 can have a greater killing effect on
tumor cells compared with sonosentizer or ultrasound therapy alone.
Therefore, we considered ZnPcS2P2 to be a
good sonosensitizer for use in SDT.
Apoptosis is one of the major modes of death of
neurospongioma. Li et al first discovered that SDT with
hematoporphyrin monomethyl ether was capable of inducing apoptosis
of C6 tumor cells through the mitochondrial pathway (13). The apoptotic rate in the 4 groups
was evaluated by flow cytometry. The early apoptotic rate was
30.93%, significantly higher than the other 3 groups. These results
indicate that ZnPcS2P2, similar to 5-AlA or
porphyrins, could be used in SDT. It is suggested that the
ZnPcS2P2-mediated SDT killing of U251 cells
involves the induction of apoptosis.
To further clarify the apoptosis phenomena by
morphological change, we observed the subcellular structure in U251
cells following SDT. In the SDT group, it was revealed that a
section of the cell microvilli had vanished, the volume of the
cellular nucleus became small and chromatin gathered densely, thus
displaying the characteristics of apoptosis or necrosis (13). The result of TEM examination also
suggested that ZnPcS2P2-mediated SDT
destroyed not only the biological membrane but also the formation
of the genetic system of tumor cells (Fig. 5). The damage of membranous
organelles greatly influenced the stabilization of the
microsurroundings and inward, as well as outward, energy exchange.
The condensation of nuclear chromatin would result in an
interruption of gene expression and regulation. These changes could
destroy metabolism, affect functions and eventually lead to cell
death (14).
Apoptosis is characterized by a number of
well-defined features, including phosphatidyserine exposure,
membrane blabbing, activation of caspase, chromatin condensation
and DNA fragmentation (15).
Classical apoptosis may proceed by the intrinsic and/or extrinsic
pathway (16–18). In the intrinsic pathway,
mitochondria are induced to release a number of factors, leading to
the formation of the apoptosome, which is comprised of the adapter
protein Apaf-1, cytochrome c and caspase-9 (19,20).
The extrinsic pathway, is believed to be based on death
receptor-dependent recruitment of the adaptor protein,
Fas-associated death domain (FADD), which, in turn, promotes
dimerization and subsequent activation of caspase-8 (21). Liu et al first discovered
that SDT was capable of inducing apoptosis through a mitochondrial
pathway (22,23). Two apoptotic pathways activate
certain members of the caspase family, respectively, but the
activation of caspase-3 finally commences apoptosis. Some
investigators report that the death acceptor pathway is not
included in HMME-meditated SDT induction of C6 cells apoptois
(24), which indicates that the
ultrasound activation of various sonosensitizers to kill different
cell lines involves various apoptotic pathways. Western blot
analysis in the 4 groups revealed that caspases-3, -8 and -9 were
significantly activated following ultrasound treatment with
ZnPcS2P2. It was indicated that both the
death acceptor and mitochondrial pathways occured in process of
ultrasound-activing ZnPcS2P2 to kill U251
cells.
To further study the mechanisms of the ultrasound
activationof ZnPcS2P2 to kill U251 cells, we
detected the level of cellular ROS. ROS, which include hydroxyl
radicals, superoxide anions, singlet oxygens and hydrogen
peroxides, are by-products of cellular metabolism. Excessive
intracellular ROS is capable of damaging critical biomolecules and
eventually results in several biological effect disorders,
including alterations in signal transduction and gene expression
for mitogenesis, mutagenesis and cell death (25,26).
Analysis of data revealed that the level of ROS was markedly
elevated in SDT compared with the other groups. This indicates that
ROS plays a crucial role in the
ZnPcS2P2-mediated SDT killing of U251
cells.
In conclusion, ZnPcS2P2-SDT is
capable of inducing cell apoptosis and necrosis in U251 cells.
Ultrasound therapy, combined with ZnPcS2P2,
has satisfying antitumor effects. ROS generation in U251 cells
plays a crucial role in ZnPcS2P2-SDT-induced
cell death and initiates subsequent apoptosis through mitochondrial
and death receptor pathways following
ZnPcS2P2-SDT.
Acknowledgements
This study was supported by the
National Natural Science Foundation (30970834; 81072079),
Technological Key Research Projects of Heilongjiang province
(GC10C304-1).
References
|
1.
|
G PerilongoConsiderations on the role of
chemotherapy and modern radiotherapy in the treatment of childhood
low grade gliomaJ
Neurooncol75301307200510.1007/s11060-005-6754-816195800
|
|
2.
|
N YumitaR NishigakiK UmemuraS
UmemuraHematoporphyrin as a sensitizer of cell damaging effect of
ultrasoundJpn J Cancer
Res80219222198910.1111/j.1349-7006.1989.tb02295.x2470713
|
|
3.
|
D KesselJ LoR JeffersJB FowlkesC CainModes
of photodynamic vs. sonodynamic cytotoxicityJ Photochem Photobiol
B28219221199510.1016/1011-1344(94)07111-Z7623187
|
|
4.
|
JL HuangNS ChenJD HuangMetal
phthalocyanine as photosensitizer for photodynamic therapy(PDT) -
preparation, characterization and anticancer activities of an
amphiphilic phthalocyanine ZnPcS2P2Sci China
Ser B441131222001
|
|
5.
|
W LiuN ChenH JinIntravenous repeated-dose
toxicity study of ZnPcS2P2-based-photodynamic
therapy in beagle dogsRegul Toxicol
Pharmacol47221231200710.1016/j.yrtph.2006.06.00617367655
|
|
6.
|
HF HuangYZ ChenY WuP ChenPurging of murine
erythroblastic leukemia by
ZnPcS2P2-based-photodynamic therapyBone
Marrow Transplant37213217200610.1038/sj.bmt.170521616284611
|
|
7.
|
HF HuangYZ ChenY WuExperimental studies of
the effects of ZnPcS2P2-based-photodynamic
therapy on bone marrow purgingChin Med J (Engl)1181051102005
|
|
8.
|
HF HuangYZ ChenY WuMitochondria-dependent
apoptosis induced by a novel amphipathic photochemotherapeutic
agent ZnPcS2P2 in HL60 cellsActa Pharmacol
Sin2611381144200510.1111/j.1745-7254.2005.00160.x16115383
|
|
9.
|
I ChowdhuryB TharakanGK BhatCurrent
concepts in apoptosis: the physiological suicide program
revisitedCell Mol Biol
Lett11506525200610.2478/s11658-006-0041-316977376
|
|
10.
|
F DunnAJ AverbuchWD O’Brien JrA primary
method for the determination of ultrasonic intensity with the
elastic sphere radiometerAcoustica3858611977
|
|
11.
|
NN OrieW ZidekM TepelChemoattractant- and
mitogen-induced generation of reactive oxygen species in human
lymphocytes: the role of calciumExp
Physiol84515520199910.1111/j.1469-445X.1999.01861.x10362849
|
|
12.
|
M KurokiK HachimineH AbeSonodynamic
therapy of cancer using novel sonosensitizersAnticancer
Res2736733677200717970027
|
|
13.
|
JH LiDY SongYG XuZ HuangW YueIn vitro
study of haematoporphyrin monomethyl ether-mediated sonodynamic
effects on C6 glioma cellsNeurol
Sci29229235200810.1007/s10072-008-0972-818810596
|
|
14.
|
AH BaratiM Mokhtari-DizajiUltrasound dose
fractionation in sonodynamic therapyUltrasound Med
Biol36880887201010.1016/j.ultrasmedbio.2010.03.01020510185
|
|
15.
|
XB WangQH LiuP WangK ZhangW TangBL
WangEnhancement of apoptosis by sonodynamic therapy with
protoporphyrin IX in isolate sarcoma 180 cellsCancer Biother
Radiopharm23238246200810.1089/cbr.2007.043618454693
|
|
16.
|
BC BarnhartEC AlappatME PeterThe CD95 type
I/type II modelSemin
Immunol15185193200310.1016/S1044-5323(03)00031-914563117
|
|
17.
|
MH AzizKE SundlingNE DreckschmidtAK
VermaProtein kinase C epsilon inhibits UVR-induced expression of
FADD, an adaptor protein, linked to both Fas- and TNFR1-mediated
apoptosisJ Invest
Dermatol12920112021200910.1038/jid.2008.45819194472
|
|
18.
|
SG KimHS JongTY KimTransforming growth
factor-beta 1 induces apoptosis through Fas ligand-independent
activation of the Fas death pathway in human gastric SNU-620
carcinoma cellsMol Biol
Cell15420434200410.1091/mbc.E03-04-020114595120
|
|
19.
|
SA ChoiSJ KimKC
ChungHuntingtin-interacting protein 1-mediated neuronal cell death
occurs through intrinsic apoptotic pathways and mitochondrial
alterationsFEBS
Lett58052755282200610.1016/j.febslet.2006.08.07616979168
|
|
20.
|
U Felderhoff-MueserM SifringerS
PesditschekPathways leading to apoptotic neurodegeneration
following trauma to the developing rat brainNeurobiol
Dis11231245200210.1006/nbdi.2002.052112505417
|
|
21.
|
AE MilnerDH PalmerEA HodgkinInduction of
apoptosis by chemotherapeutic drugs: the role of FADD in activation
of caspase-8 and synergy with death receptor ligands in ovarian
carcinoma cellsCell Death
Differ9287300200210.1038/sj.cdd.440094511859411
|
|
22.
|
Q LiuX WangP WangH QiK ZhangL
XiaoSonodynamic effects of protoporphyrin IX disodium salt on
isolated sarcoma 180
cellsUltrasonics455660200610.1016/j.ultras.2006.06.06316890264
|
|
23.
|
L Quan-hongS Shi-huiX Ya-pingSynergistic
anti-tumor effect of ultrasound and hematoporphyrin on sarcoma 180
cells with special reference to the changes of morphology and
cytochrome oxidase activity of tumor cellsJ Exp Clin Cancer
Res23333341200415354420
|
|
24.
|
S DaiS HuC WuApoptotic effect of
sonodynamic therapy mediated by hematoporphyrin monomethyl ether on
C6 glioma cells in vitroActa
Neurochir15116551661200910.1007/s00701-009-0456-519597759
|
|
25.
|
F ReineckeO LevanetsY
OlivierMetallothionein isoform 2A expression is inducible and
protects against ROS-mediated cell death in rotenone-treated HeLa
cellsBiochemistry395405415200610.1042/BJ2005125316402917
|
|
26.
|
CC SuJG LinTM LiCurcumin-induced apoptosis
of human colon cancer colo 205 cells through the production of ROS,
Ca2+ and the activation of caspase-3Anticancer
Res2643794389200617201158
|