Introduction
Lentinus (L.) edodes (also know as shiitake)
is a popular mushroom known to possess beneficial bioactivities.
This mushroom is an important ingredient in Asian cuisine, due to
its desirable taste and odor. In addition, the fungus has high
levels of vitamins B and D (1).
One of the bioactive compounds in L. edodes is eritadenine,
which has been demonstrated to exert anti-hypercholemic effects in
previous studies (2,3). In addition, eritadenine may regulate
lipid metabolism by inhibiting S-adenosyl homocysteine hydrolase
activity (4–6).
The conversion of cholesterol into bile acids in the
liver is important for eliminating cholesterol from the body. Bile
acids are essential for the absorption and transport of
lipid-soluble vitamins, steroids and xenobiotics (7–9). The
main bile acid biosynthetic pathway is initiated by cholesterol
7-α-hydroxylase 1 (CYP7A1), a cytochrome P450 heme enzyme (10). The transcription of CYP7A1 is
inhibited by bile acid feedback and stimulated by dietary factors,
such as cholesterol (11,12). The expression of CYP7A1 is
downregulated by sterol regulatory element-binding proteins
(SREBPs) when plasma cholesterol levels are low and upregulated by
the nuclear receptor liver X receptor (LXR) when cholesterol levels
are high (13).
Dyslipidemia and hypercholesterolemia are among the
numerous risk factors associated with diabetes and metabolic
diseases, including fatty liver, obesity and atherosclerosis
(14–16). Eritadenine (an adenosine analog
alkaloid) and lentinacin (a purine alkaloid) have been shown to
reduce cholesterol levels in rats by 25% following 7 days of oral
administration at doses as low as 0.005% of the feed intake
(17). The hypocholesterolemic
action of eritadenine appears to be associated with the
modification of hepatic phospholipid (PL) metabolism, through the
induction of phosphatidylethanolamine N-methyltransferase
deficiency (4). Dietary
eritadenine also alters the fatty acid and molecular profiles of
the liver and plasma by suppressing the conversion of linoleic acid
into arachidonic acid and decreasing Δ6-desaturase activity
(5). In the present study, the
effects of L. edodes were investigated in a mouse model of
hypercholesterolemia induced by the consumption of a high-fat diet
(HFD) during the growth stage of the animals (between 4 and 6 weeks
of age). The serum levels of total cholesterol (T-CHO) and
triglyceride (TG) were measured. In addition, the expression of
CYP7A1 in the liver was evaluated in order to study potential
mechanisms underlying the hypocholesterolemic effects of L.
edodes.
Materials and methods
Experimental animals
Imprinting Control Region (ICR) mice (5-week-old)
were obtained from Koatech (Pyeongtaek, Republic of Korea). All
animals were housed in polycarbonate cages and acclimated in an
environmentally controlled room (23±2°C, 50±10% relative humidity,
frequent ventilation and a 12-h light/dark cycle) prior to use. The
mice (n=60) were divided into six groups (n=10 per group).
Hypercholesterolemia was induced in five of the groups by the
consumption of an HFD (D12336; Research Diets, Inc., New Brunswick,
NJ, USA) for 2 weeks (from 4 to 6 weeks of age).
To assess the ability of L. edodes to protect
against hypercholesterolemia, the mice were fed the HFD alone as a
negative control (NC), HFD with 10 mg/kg eritadenine (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) as a positive control
(PC) and HFD supplemented with 5% (LE5; w/w), 10% (LE10; w/w) or
20% (LE20; w/w) L. edodes for 4 weeks (from 5 to 9 weeks of
age). The remaining group (vehicle) was fed with D12337 pellets
(Research Diets, Inc.) as a vehicle control. The compositions of
the HFD (D12336) and control diet (D12337) are shown in Table I. Body weight (BW) was measured
prior to and following the experimental feeding period. All animal
experimental procedures were approved by the Ethics Committee of
Chungbuk National University (Cheongju, Republic of Korea).
 | Table IComposition of the purified high-fat
(D12337) and control (D12336) diets. |
Table I
Composition of the purified high-fat
(D12337) and control (D12336) diets.
| Diet |
|---|
|
|
|---|
| D12336 | D12337 |
|---|
|
|
|
|---|
| Ingredient | grams | kcal | grams | kcal |
|---|
| Casein, 30 Mesh | 75 | 300 | 75 | 300 |
| Soy protein | 130 | 520 | 130 | 520 |
| DL-Methionine | 2 | 8 | 2 | 8 |
| Corn starch | 275 | 1100 | 522.5 | 2090 |
| Maltodextrin | 150 | 600 | 150 | 600 |
| Sucrose | 30 | 120 | 30 | 120 |
| Cellulose, BW200 | 90 | 0 | 90 | 0 |
| Soybean oil | 50 | 450 | 50 | 450 |
| Cocoa butter | 75 | 675 | 0 | 0 |
| Coconut oil, 76 | 35 | 315 | 0 | 0 |
| Mineral mix
S10001 | 35 | 0 | 35 | 0 |
| Calcium
carbonate | 2.2 | 0 | 5.5 | 0 |
| Sodium chloride | 8 | 0 | 8 | 0 |
| Potassium
citrate | 10 | 0 | 10 | 0 |
| Vitamin mix
V10001 | 10 | 40 | 10 | 40 |
| Choline
bitartrate | 2 | 0 | 2 | 0 |
| Cholesterol,
USP | 12.5 | 0 | 0.3 | 0 |
| Sodium cholic
acid | 5 | 0 | 0 | 0 |
| FD&C Red Dye
#40 | 0.1 | 0 | 0 | 0 |
| FD&C Blue Dye
#1 | 0 | 0 | 0.1 | 0 |
| Total | 1000.1 | 4126 | 1120.4 | 4126 |
Serum and urine collection and
biochemical analysis
Blood was collected from the abdominal aorta in each
mouse, transferred to serum separator tubes (Microtainer tubes;
Becton-Dickinson Co., San Jose, CA, USA) and centrifuged at 400 × g
for 15 min. The serum was collected and stored in 200-ml aliquots.
T-CHO, high-density lipoprotein (HDL), low-density lipoprotein
(LDL) and TG levels in the serum were measured using a Hitachi 7080
auto-analyzer (Hitachi Science System Ltd., Ibaraki, Japan).
RNA extraction and quantitative
polymerase chain reaction (qPCR)
Total RNA was extracted from the mouse liver using
TRIzol® reagent (Invitrogen Life Technology, Carlsbad,
CA, USA), in accordance with the manufacturer’s instructions. RNA
concentrations were measured with a microplate spectrophotometer
(Epoch; BioTek Inc., Winooski, VT, USA) at 260 nm. RNA quality was
evaluated using electrophoresis in 1% agarose gels. Total RNA (1
mg) was reverse transcribed into first-strand complementary DNA
(cDNA) using Moloney murine leukemia virus reverse transcriptase
(Invitrogen Life Technology) and random primers (9-mer; Takara Bio,
Otsu, Japan). Each cDNA sample (1 ml) was amplified with 10 ml 2X
SYBR® Premix Ex Taq™ (Takara Bio) and 10 pmol of each
primer. Amplification was performed in a 7300 Real-Time PCR System
(Applied Biosystems, Foster City, CA, USA) with the following
parameters: denaturation at 95°C for 5 min followed by 40 cycles of
denaturation at 95°C for 30 sec, annealing at 60°C for 30 sec and
extension at 72°C for 45 sec. The sequences of the oligonucleotide
primers used for this study were 5′-TCC ACC TTT GAT GAC ATG GA-3′
(sense) and 5′-TTG GCC AGC ACT CTG TAA TG-3′ (antisense) for CYP7A1
(product size, 171 bp); and 5′-CCA GGG TTT GGA ATT ATT TC-3′
(sense) and 5′-GAA GAT AAA CCC TAA GGC TC-3′ (antisense) for 1A
(product size, 297 bp). The relative expression levels of CYP7A1 in
each sample (normalized to that of 1A) were determined using RQ
software (Applied Biosystems). All qPCR experiments were repeated
twice.
Histological staining and analysis of
liver tissues
Liver tissues were recovered, frozen in liquid
nitrogen and stored at −80°C until use. Frozen liver tissues were
cut with a cryomicrotome (CM3050S; Leica Biosystems, Nussloch,
Germany) into 5-mm thick sections, stained with Oil Red O
(Sigma-Aldrich, St. Louis, MO, USA) and counterstained with Harris
hematoxylin (Sigma-Aldrich). Aortic tissues were embedded in
paraffin prior to sectioning (5-μm thick). The sections were
deparaffinized in xylene, hydrated in descending grades of ethanol
and then stained with Harris hematoxylin and eosin (Sigma-Aldrich).
The stained sections were viewed and photographed with a microscope
(BX51; Olympus, Tokyo, Japan) equipped with a digital camera (DP71;
Olympus). All images were captured at ×200 magnification.
Data analysis
Data are presented as the mean ± standard error of
the mean (SEM) and were analyzed with a one-way analysis of
variance (ANOVA) followed by Tukey’s multiple comparison test.
Statistical analyses were performed using Prism Graph Pad (version
4.0; GraphPad Software Inc., San Diego, CA, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Effects of L. edodes on BW and daily
dietary intake
The differences in BW and daily food intake between
mice fed the control diet and those given an HFD for 4 weeks were
evaluated. The mice from all six groups had similar BW gains, as
shown in Table II. The BWs of the
mice in the vehicle, NC and PC groups were all the same at the end
of experiment. Groups fed with L. edodes (5, 10 and 20%)
also had similar BW gains compared with those of the other groups
at 9 weeks of age.
| Table IIBody weights and daily food
intake. |
Table II
Body weights and daily food
intake.
| Group | Baseline body
weight (g) | Body weight 4 weeks
later (g) | Daily food intake
(g) |
|---|
| Vehicle | 36.9±0.5 | 41.5±1.8 | 3.77±0.8 |
| NC | 36.0±1.3 | 42.3±0.4 | 3.24±0.8 |
| PC | 35.6±2.1 | 43.5±0.3 | 3.83±1.7 |
| LE5 | 35.8±2.8 | 42.3±0.3 | 4.16±1.1 |
| LE10 | 35.4±2.1 | 42.1±0.6 | 4.00±1.2 |
| LE20 | 35.3±1.5 | 41.7±0.4 | 4.12±1.1 |
Serum levels of T-CHO, HDL, LDL and TG in
the hypercholesterolemic mice
Dietary supplementation with L. edodes
reduced the serum levels of T-CHO, HDL, LDL and TG, as shown in
Table III. At 4 weeks, the mice
fed the HFD showed a marked increase in T-CHO levels (to 285.2±23.3
compared with 151.5±17.7 mg/dl in the vehicle group). Dietary
supplementation with eritadenine (10 mg/kg) or L. edodes (5,
10 and 20%) attenuated the rise in serum T-CHO, LDL and TG levels
caused by consumption of an HFD. In the groups fed with L.
edodes (5, 10 and 20%), the levels of T-CHO, LDL and TG were
reduced in a dose-dependent manner compared with those in the NC
group.
| Table IIIValues of serum chemistry. |
Table III
Values of serum chemistry.
| Serum level
(mg/dl) |
|---|
|
|
|---|
| Variable | Vehicle | NC | PC | LE5 | LE10 | LE20 |
|---|
| Total
cholesterol | 151.5±17.7 | 285.2±23.3a | 239.2±15.8b | 231.5±23.4b | 182.7±18.0b,c | 147.5±20.2b,c |
|
HDL-cholesterol | 92.8±7.2 | 92.1±16.4 | 108.7±13.3 | 82.8±19.7 | 97.2±12.0 | 88.5±15.6 |
|
LDL-cholesterol | 7.1±3.0 | 53.8±1.5a | 45.6±2.0b | 42.6±7.4b | 37.8±3.6b,c | 24.5±3.7b,c |
| Triglyceride | 8.2±1.4 | 33.5±9.4a | 23.2±5.0b | 13±3.0b,c | 9.6±1.1b,c | 7.3±0.7b,c |
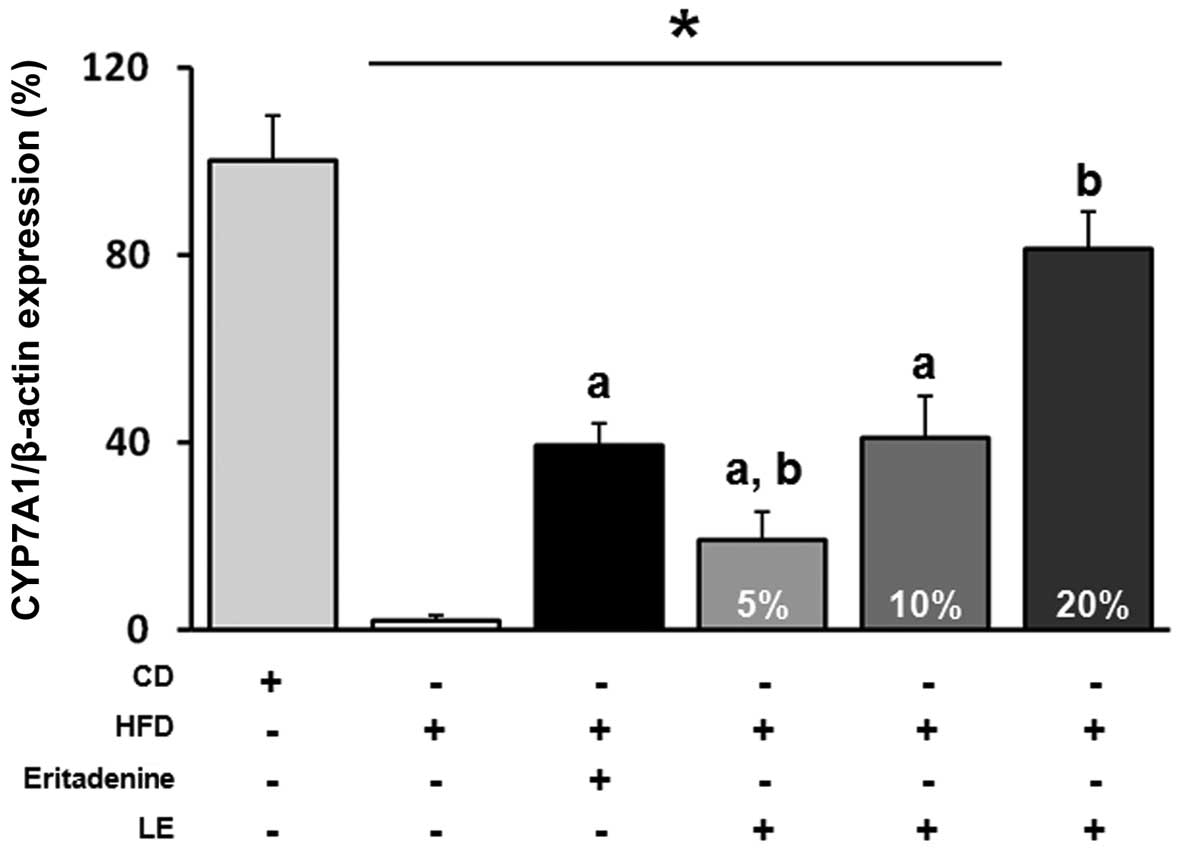
Expression of CYP7A1 mRNA in the liver of
hypercholesterolemic mice
The expression of hepatic CYP7A1 was measured in the
six groups of mice. As seen in Fig.
1, the level of CYP7A1 mRNA was reduced by HFD administration
in the NC group compared with the level in the vehicle group and
was significantly increased by eritadenine supplementation in the
PC group. Similar to the PC group, hepatic CYP7A1 mRNA expression
was increased by L. edodes supplementation in a
dose-dependent manner.
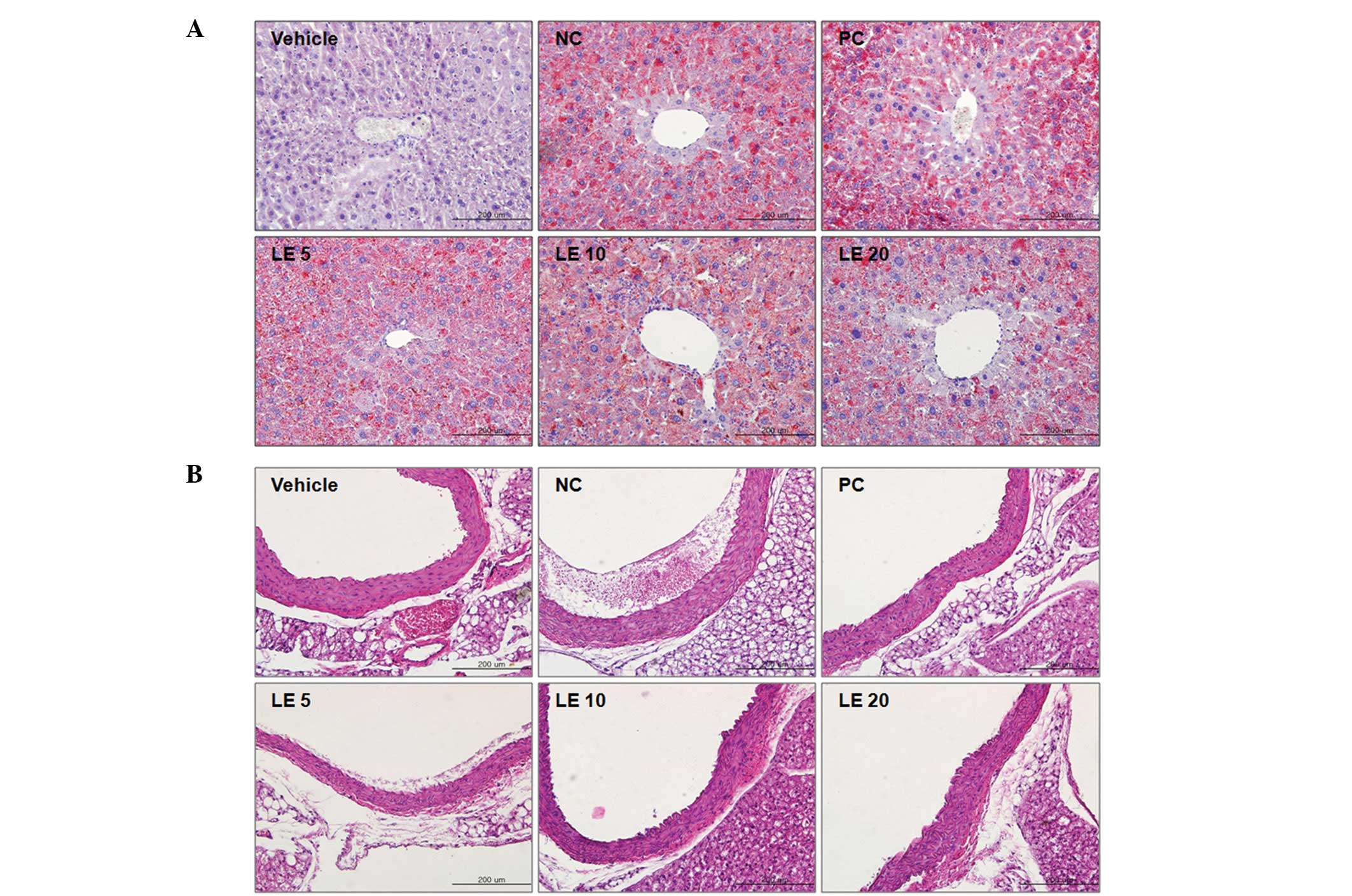
Effects of L. edodes on the histology of
hypercholesterolemic mouse liver
The effect of L. edodes on liver tissue
morphology in hypercholesterolemic mice is presented in Fig. 2A. Liver tissues were stained with
Oil Red O and hematoxylin without eosin. The hepatic cords were
arranged in a typical manner and located near the central vein in
all groups of mice. Lipid droplets, evidence of fat accumulation in
the cytoplasm of hepatocytes, were observed in the livers of
hypercholesterolemic mice fed the HFD. The lipid accumulation was
reduced by eritadenine or L. edodes supplementation.
Effects of L. edodes on the aortae of
hypercholesterolemic mice
The effect of L. edodes on the descending
aortic tissues of hypercholesterolemic mice is shown in Fig. 2B. Descending aortic tissues were
stained with hematoxylin and eosin. Atherosclerotic plaques were
observed in mice fed the HFD and the atherosclerotic plaques
observed in the PC mice were smaller than those in the mice fed the
HFD alone. The atherosclerotic plaques observed in the mice fed
with the HFD and L. edodes supplementation were smaller
and/or less numerous compared with the plaques observed in the NC
(and/or PC) mice.
Discussion
L. edodes contains relatively high
concentrations of vitamins D, B6, B9 and B12, as well as other
beneficial compounds, such as eritadenine and dietary fiber
(1). This mushroom has been
proposed to be useful for treating hypercholesterolemia,
inflammation, hypertension and osteoporosis (18–20).
In a previous study, we examined the anti-osteoporotic effects of
L. edodes and the ability of this fungus to induce the
expression of duodenal and renal calcium transport channels in mice
with osteoporosis-like symptoms (20). In the present study, we
investigated the hypolipidemic effects of L. edodes and
determined whether this fungus was able to prevent increases in
T-CHO, LDL and TG levels in mice fed an HFD. Having fed the mice
the experimental diets for 4 weeks, it was observed that dietary
L. edodes supplementation reduced serum lipid levels, as
well as fat accumulation, in the liver and descending aorta of the
hypercholesterolemic mice. Eritadenine was demonstrated to have a
similar effect. Previously, eritadenine was shown to lower
phosphatidylcholine levels in rats (3,21).
An analysis of the eritadenine levels in L. edodes showed
that the concentration of this compound ranged between 3.2 and 6.3
mg/g of dried mushrooms (22).
The stimulation of murine CYP7A1 gene expression by
dietary cholesterol has been well studied (23–25).
Oxysterols regulate the activity of responsive promoters in this
gene by activating nuclear receptors known as LXRs (25). In addition, inactivation of the
murine gene encoding the LXRα isoform has been shown to result in
resistance to the stimulation of CYP7A1 gene expression by dietary
cholesterol (23). CYP7A1 is the
key regulatory enzyme of bile acid synthesis, plays a crucial role
in cholesterol metabolism and has been implicated in genetic
susceptibility to atherosclerosis (26). A CYP7A1 polymorphism has also been
demonstrated to increase the progression of atherosclerosis in male
patients (27). Furthermore, the
development of fatty liver and hypercholesterolemia has been
prevented by regulating the expression of CYP7A1 (28).
In the present study, atherosclerotic plaques were
observed in the descending aortae of hypercholesterolemic mice.
Plaque formation induced by the consumption of an HFD was
effectively reduced by L. edodes supplementation (Fig. 2B). Hepatic fat accumulation was
also observed in the livers of hypercholesterolemic mice. This was
inhibited by eritadenine and L. edodes supplementation.
Furthermore, CYP7A1 mRNA expression was decreased in the
hypercholesterolemic mice and upregulated by eritadenine, as well
as by L. edodes (5, 10 and 20%), in a dose-dependent manner
compared with the levels in the NC group.
In conclusion, the present study demonstrated that
supplementation with eritadenine and L. edodes significantly
inhibited the development of hypercholesterolemia induced by an
HFD. The reductions in serum lipid levels, hepatic fat accumulation
and aortic atherosclerotic plaque formation in mice treated with
L. edodes illustrated the beneficial effects of this fungus.
In addition, dietary supplementation with eritadenine or L.
edodes increased the level of hepatic CYP7A1 expression in the
hypercholesterolemic mice up to that observed in the vehicle
control. Based on these results, we propose that the beneficial
compounds present in L. edodes may be used in the treatment
of hypercholesterolemia.
Acknowledgements
This study was supported by the National Research
Foundation of Korea (NRF; grant no. 2010-0011433), funded by the
Republic of Korea government (MEST).
References
|
1
|
Wasser SP: Shiitake (Lentinus
edodes). Encyclopedia of Dietary Supplements. View Article : Google Scholar : Marcel Dekker; New
York: pp. 653–664. 2005
|
|
2
|
Ngai PH and Ng TB: Lentin, a novel and
potent antifungal protein from shitake mushroom with inhibitory
effects on activity of human immunodeficiency virus-1 reverse
transcriptase and proliferation of leukemia cells. Life Sci.
73:3363–3374. 2003. View Article : Google Scholar
|
|
3
|
Shimada Y, Morita T and Sugiyama K:
Eritadenine-induced alterations of plasma lipoprotein lipid
concentrations and phosphatidylcholine molecular species profile in
rats fed cholesterol-free and cholesterol-enriched diets. Biosci
Biotechnol Biochem. 67:996–1006. 2003. View Article : Google Scholar
|
|
4
|
Sugiyama K, Akachi T and Yamakawa A:
Hypocholesterolemic action of eritadenine is mediated by a
modification of hepatic phospholipid metabolism in rats. J Nutr.
125:2134–2144. 1995.PubMed/NCBI
|
|
5
|
Shimada Y, Morita T and Sugiyama K:
Dietary eritadenine and ethanolamine depress fatty acid desaturase
activities by increasing liver microsomal phosphatidylethanolamine
in rats. J Nutr. 133:758–765. 2003.PubMed/NCBI
|
|
6
|
Fukada S, Setoue M, Morita T and Sugiyama
K: Dietary eritadenine suppresses guanidinoacetic acid-induced
hyperhomocysteinemia in rats. J Nutr. 136:2797–2802.
2006.PubMed/NCBI
|
|
7
|
Adachi R, Honma Y, Masuno H, et al:
Selective activation of vitamin D receptor by lithocholic acid
acetate, a bile acid derivative. J Lipid Res. 46:46–57. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Teo NH, Scott JM, Reed B, Neale G and Weir
DG: Bile acid inhibition of vitamin B12 binding by intrinsic factor
in vitro. Gut. 22:270–276. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Seo YK, Chung YT, Kim S, Echchgadda I,
Song CS and Chatterjee B: Xenobiotic- and vitamin D-responsive
induction of the steroid/bile acid-sulfotransferase Sult2A1 in
young and old mice: the role of a gene enhancer in the liver
chromatin. Gene. 386:218–223. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Russell DW and Setchell KD: Bile acid
biosynthesis. Biochemistry. 31:4737–4749. 1992. View Article : Google Scholar
|
|
11
|
Hubacek JA and Bobkova D: Role of
cholesterol 7alpha-hydroxylase (CYP7A1) in nutrigenetics and
pharmacogenetics of cholesterol lowering. Mol Diagn Ther.
10:93–100. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Moon MS, Lee MS, Kim CT and Kim Y: Dietary
chitosan enhances hepatic CYP7A1 activity and reduces plasma and
liver cholesterol concentrations in diet-induced
hypercholesterolemia in rats. Nutr Res Pract. 1:175–179. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chawla A, Saez E and Evans RM: Don’t know
much bile-ology. Cell. 103:1–4. 2000.
|
|
14
|
Liu Y, Wu H, Wu W, Li R and Huang Y:
Intervention of Huanglian Jiedu decoction on haemorheology and
cholesterol-supplemented diet. Zhongguo Zhong Yao Za Zhi.
34:600–604. 2009.(In Chinese).
|
|
15
|
Tuncer MA, Yaymaci B, Sati L, et al:
Influence of Tribulus terrestris extract on lipid profile
and endothelial structure in developing atherosclerotic lesions in
the aorta of rabbits on a high-cholesterol diet. Acta Histochem.
111:488–500. 2009.
|
|
16
|
von Holt K, Lebrun S, Stinn W, Conroy L,
Wallerath T and Schleef R: Progression of atherosclerosis in the
Apo E−/− model: 12-month exposure to cigarette mainstream smoke
combined with high-cholesterol/fat diet. Atherosclerosis.
205:135–143. 2009.
|
|
17
|
Chibata I, Okumura K, Takeyama S and
Kotera K: Lentinacin: a new hypocholesterolemic substance in
Lentinus edodes. Experientia. 25:1237–1238. 1969. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kabir Y, Yamaguchi M and Kimura S: Effect
of shiitake (Lentinus edodes) and maitake (Grifola
frondosa) mushrooms on blood pressure and plasma lipids of
spontaneously hypertensive rats. J Nutr Sci Vitaminol (Tokyo).
33:341–346. 1987.
|
|
19
|
Carbonero ER, Gracher AHP, Komura DL, et
al: Lentinus edodes heterogalactan: Antinociceptive and
anti-inflammatory effects. Food Chem. 111:531–537. 2008. View Article : Google Scholar
|
|
20
|
Lee GS, Byun HS, Yoon KH, Lee JS, Choi KC
and Jeung EB: Dietary calcium and vitamin D2 supplementation with
enhanced Lentinula edodes improves osteoporosis-like
symptoms and induces duodenal and renal active calcium transport
gene expression in mice. Eur J Nutr. 48:75–83. 2009.PubMed/NCBI
|
|
21
|
Takashima K, Sato C, Sasaki Y, Morita T
and Takeyama S: Effect of eritadenine on cholesterol metabolism in
the rat. Biochem Pharmacol. 23:433–438. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Enman J, Rova U and Berglund KA:
Quantification of the bioactive compound eritadenine in selected
strains of shiitake mushroom (Lentinus edodes). J Agric Food
Chem. 55:1177–1180. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Peet DJ, Turley SD, Ma W, et al:
Cholesterol and bile acid metabolism are impaired in mice lacking
the nuclear oxysterol receptor LXR alpha. Cell. 93:693–704. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Torchia EC, Cheema SK and Agellon LB:
Coordinate regulation of bile acid biosynthetic and recovery
pathways. Biochem Biophys Res Commun. 225:128–133. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Agellon LB, Drover VA, Cheema SK, Gbaguidi
GF and Walsh A: Dietary cholesterol fails to stimulate the human
cholesterol 7alpha-hydroxylase gene (CYP7A1) in transgenic mice. J
Biol Chem. 277:20131–20134. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lambrinoudaki I, Kaparos G, Papadimitriou
D, et al: Methylenetetrahydrofolate reductase C677T polymorphism is
associated with central adiposity and increased androgenicity in
healthy postmenopausal women. Eur J Endocrinol. 159:233–241. 2008.
View Article : Google Scholar
|
|
27
|
Hofman MK, Princen HM, Zwinderman AH and
Jukema JW: Genetic variation in the rate-limiting enzyme in
cholesterol catabolism (cholesterol 7alpha-hydroxylase) influences
the progression of atherosclerosis and risk of new clinical events.
Clin Sci (Lond). 108:539–545. 2005. View Article : Google Scholar
|
|
28
|
Yiu WF, Kwan PL, Wong CY, et al:
Attenuation of fatty liver and prevention of hypercholesterolemia
by extract of Curcuma longa through regulating the expression of
CYP7A1, LDL-receptor, HO-1, and HMG-CoA reductase. J Food Sci.
76:H80–H89. 2011. View Article : Google Scholar : PubMed/NCBI
|