Introduction
Glutathione (GSH) is a tripeptide that functions as
an antioxidant. It is a coenzyme that is involved in the
detoxification of endogenous and exogenous compounds (1). GSH conjugation with oxidants is
catalyzed by the enzyme glutathione S-transferase.
GSH-dependent detoxifying reactions protect cells from oxidative
damage, but consequently reduce intracellular GSH levels. The
replenishment of GSH is achieved by recycling and biosynthesis
(2), which is regulated by
substrate availability and the synthesis rate. Intracellular
synthesis of GSH occurs by two consecutive adenosine triphosphate
(ATP)-dependent enzymatic reactions. In the first reaction,
glutamate is coupled with cysteine to form γ-glutamylcysteine
(γ-GC). This is catalyzed by glutamate cysteine ligase (GCL), which
is the rate-limiting enzyme of GSH biosynthesis. In the second
reaction, γ-GC is coupled with glycine to form GSH and this is
catalyzed by GSH synthetase (2).
Aging is a process of chronic oxidative stress and
has been shown to affect GSH levels in a tissue-specific manner
(3). In type I skeletal muscles,
GSH levels increase with aging (4,5). In
the soleus muscle under stress, aging impairs the expression of the
catalytic subunit of GCL (GCLC) (2). This decreased GCL activity is
associated with the reduction of GCLC protein expression (2)
A previous study observed that certain plant
extracts affect GCLC content (6).
Resveratrol (RSV; 3,5,4′-trihydroxystilbene) is a polyphenolic
bioactive substance with multiple functions that occurs naturally
in several plant species, including grapevines and berries
(7). Accumulating evidence has
indicated that RSV has anticarcinogenic, anti-inflammatory,
antimicrobial, antiviral and antioxidant properties that may be
relevant to chronic diseases and/or longevity in humans (8,9). In
addition, RSV has been hypothesized to possess anti-aging activity.
For example, RSV has been shown to prolong the lifespan and retard
the onset of age-associated markers in a short-lived fish, as well
as in invertebrate nematode worms and fruit flies (10–14).
Therefore, the aim of the current study was to evaluate the effect
that a dietary intake of RSV had on the expression of GCLC and the
modifier subunit of GCL (GCLM) in the kidneys of aged rats.
Materials and methods
Animals and treatments
All animal experiments were conducted according to
the methods of Yuan et al (7). The animal experimental procedures
were approved by the Ethics Committee of the Laboratory Animal
Administration of Shandong Province (permit number, SD2007695;
Jinan, China). Specific pathogen-free male Wistar rats (Rattus
norvegicus), aged 8 weeks (no. 20080002), were purchased from
Shandong Lukang Pharmaceutical Co., Ltd. (Jining, China) and housed
in an environmentally controlled atmosphere (temperature, 22°C;
relative humidity, 56%) with a 12-h light/dark cycle. The rats had
free access to water and their respective diets and were provided
with shredded wood floor bedding for social activity. Five rats
were housed in each cage until the age of 3 months and thereafter
two rats were housed per cage. Wistar male rats have a life span of
24–30 months; thus, 12- and 21-month-old rats were used as models
of middle-aged and aged rats, respectively.
Young (2 months old; n=20), middle-aged (12 months
old; n=20) and aged (21 months old; n=20) rats were each randomly
divided into two groups of 10 animals. Rats in the control groups
(n=10 per age group) were fed a controlled diet (complete
semisynthetic columniformed diet containing 18% crude proteins and
5% cellulose, following the recommendations of the Chinese
Association For Laboratory Animal Sciences), while rats in the
experimental groups (n=10 per age group) were fed the controlled
diet supplemented with RSV (Tianjin Jianfeng Natural Product
R&D Co., Ltd., Tianjin, China) at a dose of 22 mg/kg of body
weight continuously for 45 days. RSV was dissolved in distilled
water to reach a concentration of 10 mg/ml. A volume of RSV
solution that correlated with the body weight of each rat was
injected into a small piece of the controlled diet. The food
injected with RSV was then administered to the rats, ensuring that
the rats ate the food completely. Drinking bottles with fresh
mineral water were replaced daily. Throughout the 45-day study
period, rats were allowed free access to the selected diet and
drinking water. At the end of the study, rats were sacrificed under
anesthesia and the kidneys were removed under sterile
conditions.
GCL activity assay
GCL activity assays were performed according to the
method of Chen et al (2).
GCL activity levels were determined by a fluorescence assay. Frozen
rat kidneys were homogenized in 20 mM Tris, 1 mM EDTA, 250 mM
sucrose, 20 mM sodium borate and 2 mM serine (TES/SB buffer).
Homogenates were centrifuged at 10,000 × g at 4°C for 10 min. The
supernatants were collected and then centrifuged again at 15,000 ×
g at 4°C for 20 min. Protein concentrations in the supernatants
were determined using a bicinchoninic acid (BCA) protein assay kit
(Beyotime Institute of Biotechnology, Shanghai, China) with bovine
serum albumin as the standard.
In the GCL activity assay, 30 μl homogenate was
added to 30 μl GCL reaction cocktail (400 mM Tris, 40 mM ATP, 40 mM
L-glutamic acid, 2 mM EDTA, 20 mM sodium borate, 2 mM serine and 40
mM MgCl2) and incubated at 37°C for 5 min. After
incubation for 5 min, 30 μl cysteine (30 mM; dissolved in TES/SB
buffer) was added and the mixture was incubated for 13 min at 37°C.
The enzymatic reaction in the mixture was stopped by precipitating
the proteins with 200 mM 5-sulfosalicylic acid (SSA). After placing
on ice for 20 min, the mixture was centrifuged at 2,000 × g at 4°C
for 10 min. Next, 20 μl supernatant containing the γ-GC product was
added to a 96-well plate designed for fluorescence detection. For
each assay, 20 μl γ-GC standards containing 30 μl GCL reaction
cocktail, 30 μl SSA (200 mM), 30 μl H2O and 30 μl γ-GC
standard solution (0, 20, 40, 60, 80, 100, 120 and 140 μM γ-GC in
TES/SB buffer) were added to the same 96-well plate to generate a
standard curve. Next, 180 μl 2,3-naphthalenedicarboxyaldehyde (NDA)
was added to each well. The plate was incubated in the dark at room
temperature for 30 min. Following incubation, the formation of
NDA-γ-GC was measured (472 nm excitation/528 nm emission) using a
fluorescent plate reader (GENios Plus; Tecan Ltd., Männedorf,
Switzerland). The quantity of γ-GC in each sample was calculated
using the standard curve. Values were expressed in mM/min/mg of
protein.
Protein content of GCLC and GCLM
The protein levels of GCLC and GCLM in the kidneys
were determined by western blot analysis. Kidneys were homogenized
in DNase buffer containing 20 mM Tris (pH 6.8), 1 mM
CaCl2, 5 mM MgCl2 and 150 U/ml DNase I
(Takara Bio, Inc., Dalian, China). The homogenate was then placed
on ice for 40 min. Next, urea buffer containing 6 M urea, 2% sodium
dodecyl sulfate (SDS) and 20 mM Tris (pH 6.8) was added to the
homogenate, which was then homogenized again. The homogenate was
centrifuged at 600 × g at 4°C for 15 min and the supernatant was
collected. Protein concentrations in the supernatant were
determined using the BCA protein assay kit. Equal amounts (20 μg
for GCLC and 50 μg for GCLM, determined by linear responses of the
respective antibody) of protein were loaded onto 12%
SDS-polyacrylamide gels and separated by electrophoresis using
Mini-Vertical Gel Electrophoresis Units (Bio-Rad, Hercules, CA,
USA). Proteins resolved on the gels were transferred to
polyvinylidene difluoride (PVDF) membranes using Mini Trans-Blot
Electrophoretic Transfer Cells (Bio-Rad) at 110 V for 3 h. A kidney
sample was used as an internal control and loaded and transferred
on each blot. The band intensity of all the samples was normalized
against the intensity of the internal control, thus permitting the
comparison of samples across multiple blots.
Protein-bound PVDF membranes were incubated
overnight at 4°C with polyclonal GCLC antibodies (1:3,000; Abcam,
Cambridge, UK) or monoclonal GCLM antibodies (1:3,000; Abcam).
Blots probed with GCLC or GCLM antibodies were then incubated with
secondary antibodies (goat anti-rabbit IgG; 1:3,000; Beijing
Kangwei Technology Group Co., Ltd., Beijing, China) conjugated with
peroxidase for 1 h at room temperature. The substrate,
5-bromo-4-chloro-3-indolyl phosphate p-toluidine-nitroblue
tetrazolium chloride (Beijing Kangwei Technology Group Co., Ltd.,
Beijing, China), was used for colorimetric visualization of the
immunoreactions on the membranes. The immunoblots were imaged using
a JS-680D automatic gel imaging analyzer (Shanghai Peiqing Science
and Technology Co., Ltd., Shanghai, China). The intensity of the
immunoreactions on the blots was quantified using Quantity One
software (SensiAnsys; Shanghai Peiqing Science and Technology Co.,
Ltd.).
Quantitative polymerase chain reaction
(qPCR)
Kidney samples were pulverized using liquid
nitrogen. Total RNA was extracted from the samples using TRIzol
reagent (Takara Bio, Inc.) and treated with RNase-free DNase
(Sangon Biotech, Co., Ltd., Shanghai, China) to remove genomic DNA
contamination. Next, 1 μg RNA was reverse transcribed to cDNA using
a reverse transcription system kit (Sangon Biotech Co., Ltd.). Gene
expression levels of GCLC and GCLM were evaluated by qPCR. Primers
for GCLC were as follows: Forward, 5′-CTGAGGCAAGATACCTTTATGACC-3′
and reverse, 5′-GTAGCTATCTATTGAGTCATACCGAGAC-3′. Primers for GCLM
were as follows: Forward, 5′-CTGTACCAGTGG GCACAGGTAA-3′ and
reverse, 5′-TTGGGTCATTGTGAG TCAGTAGC-3′. The mRNA expression level
of β-actin was also detected as an internal control for each
sample. For β-actin, the following primers were used: Forward,
5′-ACA TCCGTAAAGACCTCTATGCCAACA-3′ and reverse,
5′-GTGCTAGGAGCCAGGGCAGTAATCT-3′.
qPCR was performed using a SYBR Green I PCR kit
(Takara Bio, Inc.), according to the manufacturer’s instructions,
in an ABI PRISM 7500 sequence detection system (PerkinElmer,
Norwalk, CT, USA). Amplification conditions were as follows: 95°C
for 10 sec, followed by 40 cycles of 95°C for 5 sec and 60°C for 41
sec. Each experiment was performed in triplicate. The PCR products
were run in an agarose gel and were confined to a single band of
the expected size in all cases. Melting-curve analysis was also
performed to ensure the specificity of the products. Relative mRNA
expression levels of GCLC and GCLM were determined using the
comparative (2−ΔΔCt) method.
Statistical analysis
Statistical analysis was performed using the
Student’s t-test with SPSS software, version 13.0 (SPSS, Inc.,
Chicago, IL, USA). Analysis was conducted separately for the young,
middle-aged and aged rat studies since they were performed
independently. Data are expressed as the mean ± standard error.
P<0.05 was considered to indicate a statistically significant
difference. All figures were created with GraphPad Prism software,
version 5.0 (GraphPad Software, Inc., La Jolla, CA, USA).
Results
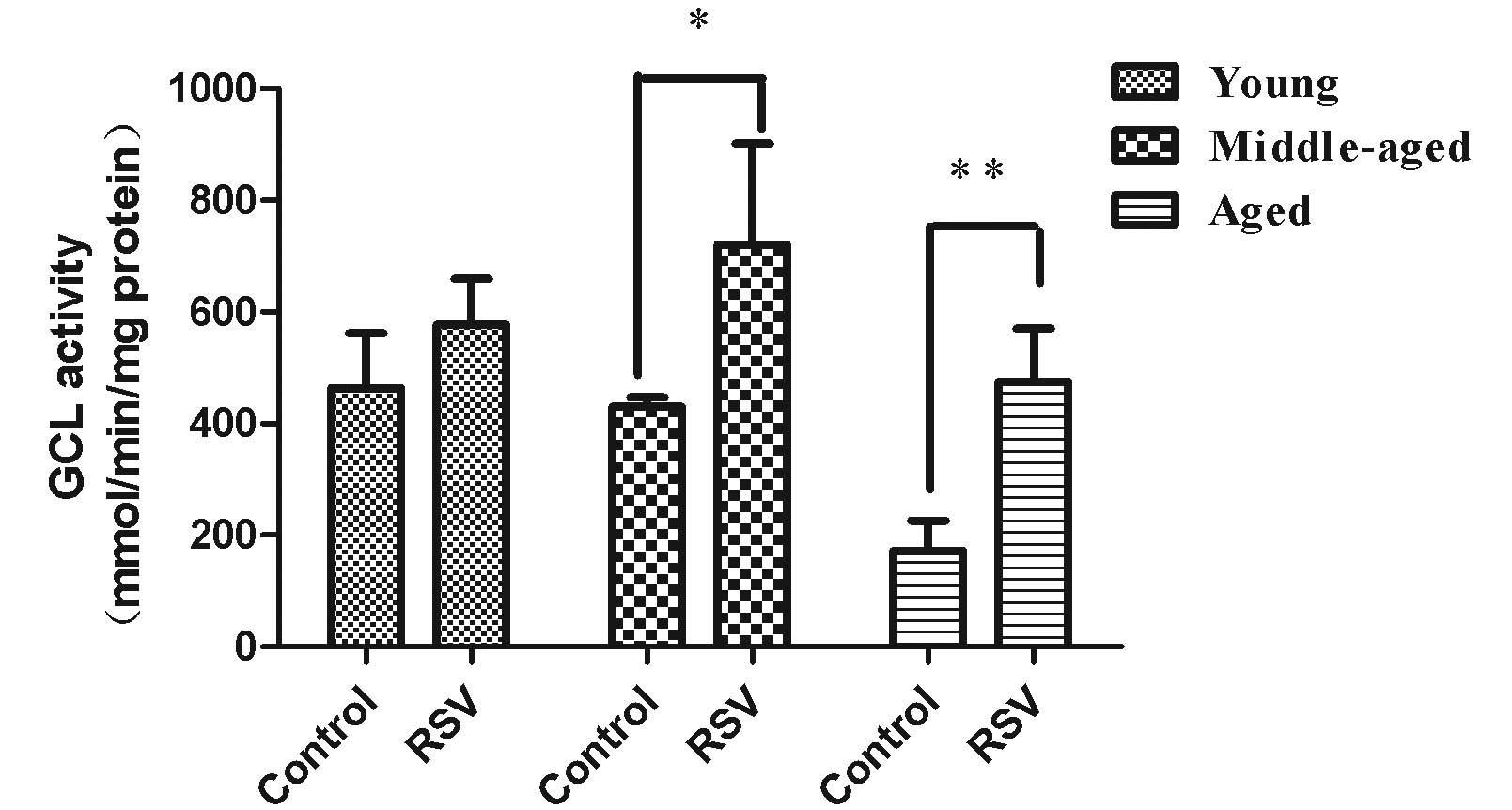
GCL activity
GCL is the rate-limiting enzyme of GSH synthesis
(15). RSV supplementation was not
observed to significantly affect the GCL activity levels (Fig. 1) in the kidneys of the young rats
(578 ± 83 mmol/min/mg protein) compared with those in the
respective control group (425 ± 92 mmol/min/mg protein). As shown
in Fig. 1, the GCL activity levels
were significantly increased in the kidneys of middle-aged (668 ±
110 mmol/min/mg protein, p=0.026) and aged rats (508 ± 94
mmol/min/mg protein, p=0.001) that were fed the RSV-supplemented
diet compared with those in the respective control groups (401 ± 62
mmol/min/mg protein for middle-aged rats and 189 ± 81 mmol/min/mg
protein for aged rats).
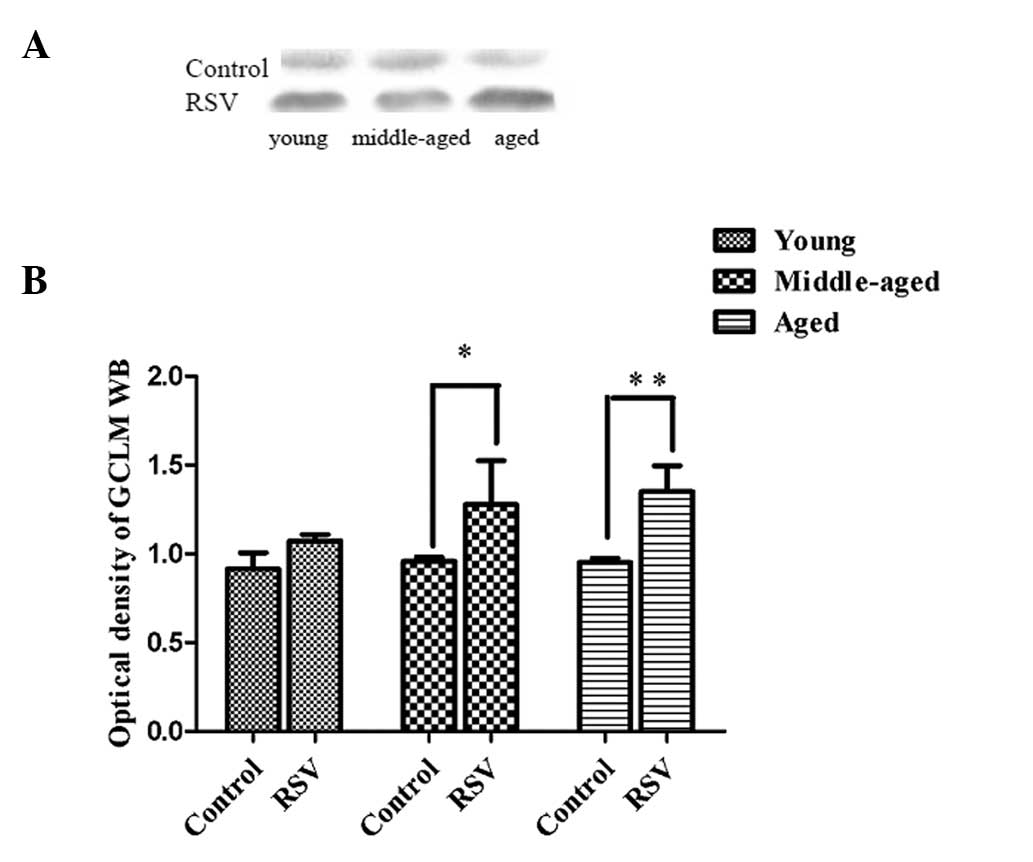
Protein content of the GCL subunits
In order to understand the increase of GCL activity
levels in the kidneys of aged rats, the protein content of the two
subunits of GCL was determined.
GCLM decreases the Km (increases the affinity) for
glutamate and ATP and increases the concentration of GSH required
for GCL inhibition (Ki) (16).
Fig. 2 shows the representative
immunoblots and summary of densitometric analysis of GCLM from the
kidneys of the rats. In young rats, the relative GCLM content of
the kidneys did not change significantly when the RSV-supplemented
diet was administered. In middle-aged (1.44 ± 0.83, P=0.016) and
aged rats (1.39 ± 0.56, P=0.001) fed the controlled diet
supplemented with RSV, the relative GCLM contents in kidneys were
greater than the values observed in the respective control
groups.
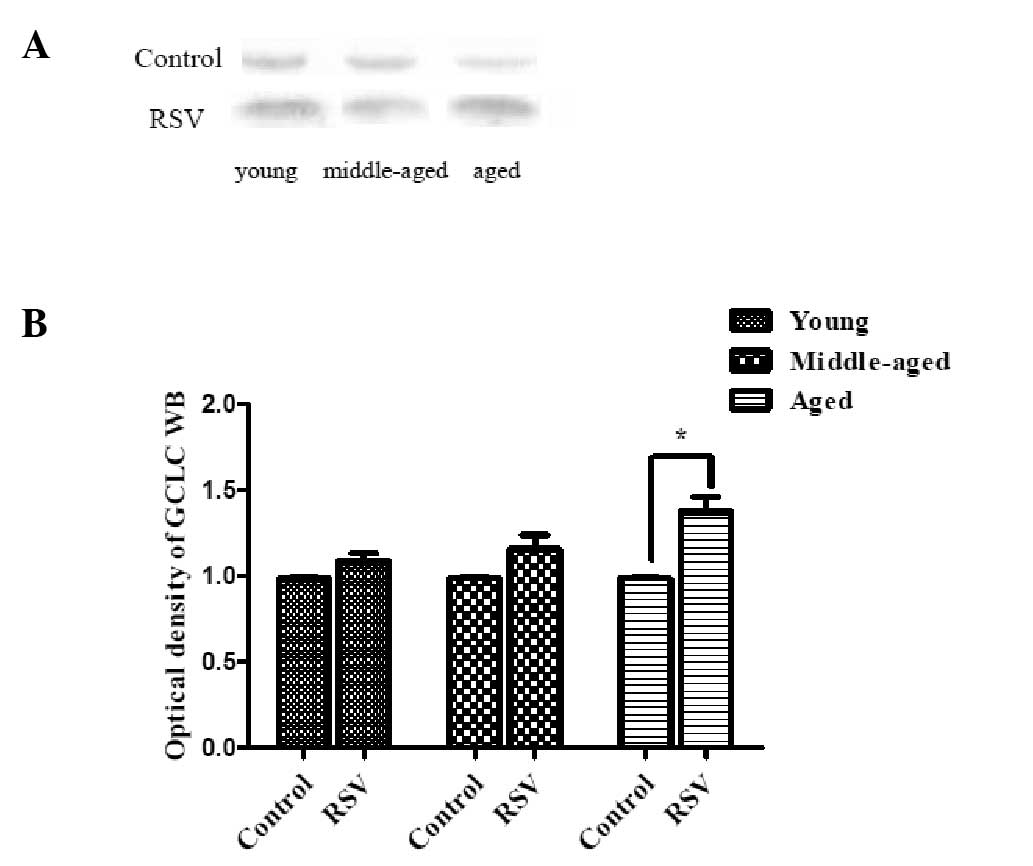
GCLC produces the catalytic function of GCL
(16). Fig. 3 shows the representative
immunoblots and densitometric analysis of GCLC from the kidneys of
the rats. As shown in Fig. 3, the
GCLC content of the kidneys was significantly increased in aged
rats (1.37 ± 0.38, P=0.012) fed RSV-supplemented diet compared with
those in the respective control group. However, no significant
change was observed in the young and middle-aged rats.
These results demonstrate the age-associated
differences in the protein levels of the GCL subunits in the
kidneys from aged rats fed an RSV-supplemented diet.
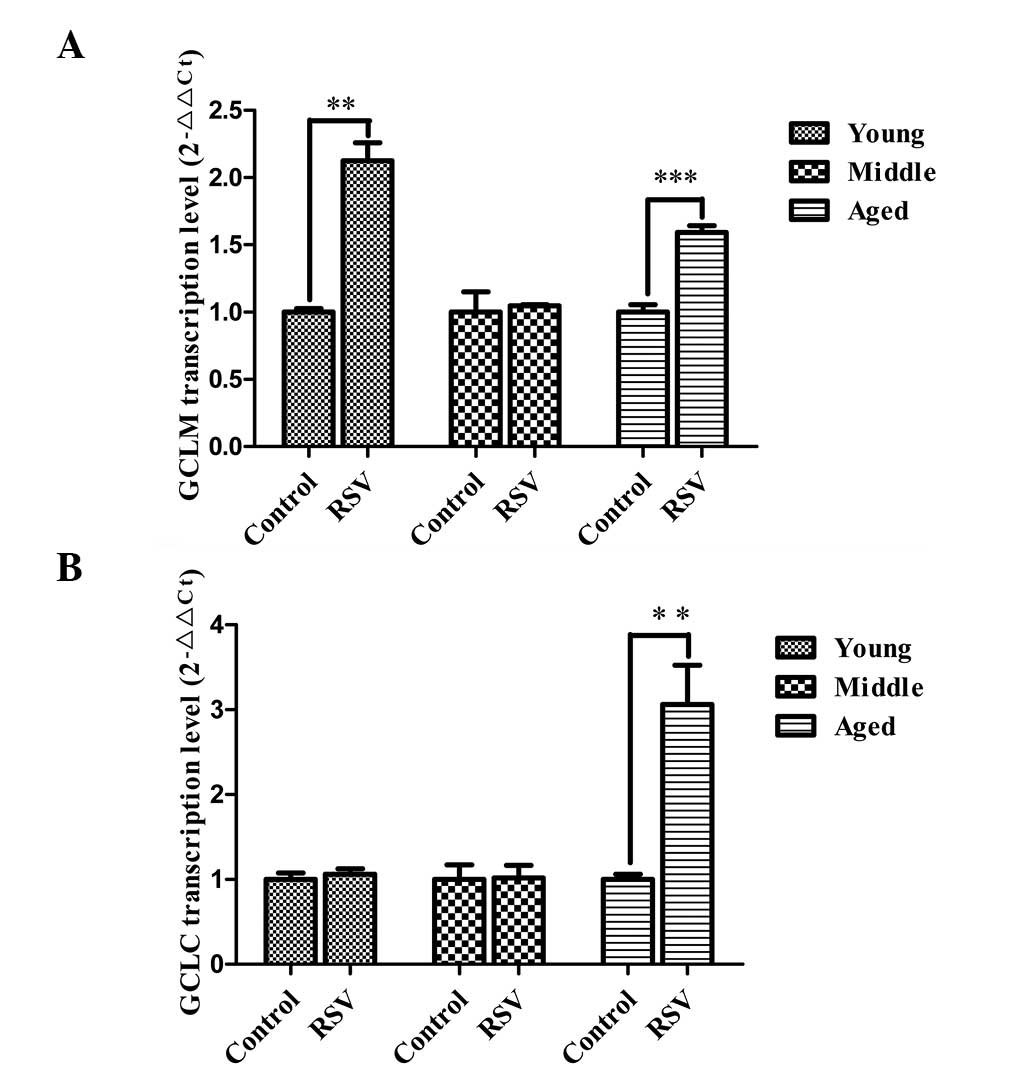
Quantification of the mRNA expression
levels of the GCL subunits
The mRNA expression levels of the GCL subunits were
examined in the kidneys of rats fed the control diet or a diet
supplemented RSV using qPCR. The results showed that the relative
expression levels of GCLM mRNA (Fig.
4A) in kidneys of aged rats (1.55 ± 0.15, P=0.001) fed a diet
supplemented with RSV were significantly higher than those in the
respective controls; they were also markedly increased in young
rats (2.13 ± 0.23, P=0.001) fed the RSV-supplemented diet. A
similar effect was not observed in the middle-age rats. As shown in
Fig. 4B, the levels of GCLC mRNA
expression were significantly increased in aged rats fed an
RSV-supplemented diet (2.86 ± 0.31, P=0.001) compared with those in
the control; however, a similar change was not observed in the
young and middle-aged rats. These results indicate that dietary RSV
supplementation is able to induce a significant increase in the
mRNA expression levels of GCLC and GCLM in aged rats.
Discussion
Alterations in GSH metabolism with aging (a
condition of chronic oxidative stress) are tissue specific
(4,17–23).
With aging, organisms exhibit a reduction in the ability to adapt
to stress (24–26). Identifying the age-associated
changes of GSH homeostasis is important since GSH regulates the
redox balance of the cells (2).
GCL is the rate-limiting enzyme of GSH synthesis. While the brain
and liver are the most studied tissues, few studies have
investigated the age-associated changes of GCL in the kidneys.
Accumulating evidence indicates that RSV has anticarcinogenic,
antiinflammatory, antimicrobial, antiviral and antioxidant
properties that may be relevant to chronic diseases and/or
longevity in humans (8,9). Thus, the aim of the present study was
to determine the mechanism behind the age-associated differences in
kidney GCL levels of rats fed a diet supplemented with RSV.
Mechanisms underlying the age-associated
upregulation of GSH and GSH-dependent detoxifying enzymes are
unknown. However, upregulation is likely to be a compensatory
adaptation responding to chronic oxidative stress developed during
the aging process (24,27). In the control group in the current
study, the GCL activity in the kidneys of the aged rats was
observed to be lower compared with the values in the middle-aged
and young rats. Although the antioxidant system shows a
compensatory adaptation with aging, the ability of aged organisms
to positively respond to an additional stress appears to be
compromised. The results of the present study indicate that the
reduction in GCL activity contributes to a reduction in GSH levels.
GCL activity in the kidneys of aged rats fed the RSV diet
significantly increased compared with that in the aged rats fed the
control diet, and this increase was closely associated with
augmentation of the mRNA expression and protein levels of GCLC and
GCLM. GCL activity is regulated by the two subunits, GCLC and GCLM.
GCLC (73 kDa) contains the active site for the ATP-dependent bond
formation between glutamate and cysteine and possesses the
catalytic activity of GCL (28).
Studies have shown that GCLC alone is necessary and sufficient for
γ-GC formation (29,30). GCLM (31 kDa), although having no
catalytic activity, enhances enzyme activity by increasing the
affinity of GCLC to glutamate and ATP and increasing the
concentration of GSH required to inhibit GCL activity. Yang et
al (30) demonstrated that
GCLM homozygous knockout mice had lower GSH levels, increased Km
values of GCLC to glutamate and were more sensitive to oxidative
stress than their wild-type littermates were. These results
demonstrate the important contribution of GCLM in the regulation of
GSH levels (2).
In order to understand the increase in GCL activity
levels in the kidneys of aged rats fed an RSV diet, the protein
levels of the two subunits of GCL were investigated. In the aged
rats, it was observed that the GCLC and GCLM protein levels changed
significantly. Studies where the GCLC content has been maintained
have shown that the addition of GCLM protein increases GCL
activity, and conversely, a reduction in GCLM content decreases GCL
activity (23,30,31).
Previous studies have also indicated that the effect of GCLM on the
changes to GCL activity with stress is likely to be dependent on
the changes of GCLC content (2).
Increased GCL activity in the kidneys of aged rats
fed a diet supplemented with RSV is associated with an increase in
GCLC protein and mRNA expression. These results indicate that
dietary RSV supplementation is able to modulate GCL activity.
Therefore, the results of the present study
highlight the antioxidant properties of RSV mediated via the
modulation of GCL activity in the kidneys of aged rats. RSV is a
good candidate for further study of antioxidative activity. These
observations have important implications in the development of
therapeutic agents for the kidneys that may enable the elderly
population to combat oxidant stress.
References
|
1
|
Brown-Borg HM and Rakoczy SG: Glutathione
metabolism in long-living Ames dwarf mice. Exp Gerontol.
40:115–120. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen CN, Brown-Borg HM, Rakoczy SG,
Ferrington DA and Thompson LV: Aging impairs the expression of the
catalytic subunit of glutamate cysteine ligase in soleus muscle
under stress. J Gerontol A Biol Sci Med Sci. 65:129–137. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Maher P: The effects of stress and aging
on glutathione metabolism. Ageing Res Rev. 4:288–314. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Leeuwenburgh C, Fiebig R, Chandwaney R and
Ji LL: Aging and exercise training in skeletal muscle: responses of
glutathione and antioxidant enzyme systems. Am J Physiol.
267:R439–R445. 1994.PubMed/NCBI
|
|
5
|
Chen CN, Brown-Borg HM, Rakoczy SG and
Thompson LV: Muscle disuse: adaptation of antioxidant systems is
age dependent. J Gerontol A Biol Sci Med Sci. 63:461–466. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu XP, Goldring CE, Wang HY, et al:
Extract of Ginkgo biloba induces glutamate cysteine ligase
catalytic subunit (GCLC). Phytother Res. 22:367–371. 2008.
|
|
7
|
Yuan J, Lu L, Zhang Z and Zhang S: Dietary
intake of resveratrol enhances the adaptive immunity of aged rats.
Rejuvenation Res. 15:507–515. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pervaiz S and Holme AL: Resveratrol: its
biologic targets and functional activity. Antioxid Redox Signal.
11:2851–2897. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vang O, Ahmad N, Baile CA, et al: What is
new for an old molecule? Systematic review and recommendations on
the use of resveratrol. PLoS One. 6:e198812011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Valenzano DR, Terzibasi E, Genade T,
Cattaneo A, Domenici L and Cellerino A: Resveratrol prolongs
lifespan and retards the onset of age-related markers in a
short-lived vertebrate. Curr Biol. 16:296–300. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Evason K, Huang C, Yamben I, Covey DF and
Kornfeld K: Anticonvulsant medications extend worm life-span.
Science. 307:258–262. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Howitz KT, Bitterman KJ, Cohen HY, et al:
Small molecule activators of sirtuins extend Saccharomyces
cerevisiae lifespan. Nature. 425:191–196. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kang HL, Benzer S and Min KT: Life
extension in Drosophila by feeding a drug. Proc Natl Acad
Sci USA. 99:838–843. 2002.
|
|
14
|
Wood JG, Rogina B, Lavu S, Howitz K,
Helfand SL, Tatar M and Sinclair D: Sirtuin activators mimic
caloric restriction and delay ageing in metazoans. Nature.
430:686–689. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lu SC: Regulation of glutathione
synthesis. Mol Aspects Med. 30:42–59. 2009. View Article : Google Scholar
|
|
16
|
Franklin CC, Backos DS, Mohar I, White CC,
Forman HJ and Kavanagh TJ: Structure, function, and
post-translational regulation of the catalytic and modifier
subunits of glutamate cysteine ligase. Mol Aspects Med. 30:86–98.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hollander J, Bejma J, Ookawara T, Ohno H
and Ji LL: Superoxide dismutase gene expression in skeletal muscle:
fiber-specific effect of age. Mech Ageing Dev. 116:33–45. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ji LL, Dillon D and Wu E: Alteration of
antioxidant enzymes with aging in rat skeletal muscle and liver. Am
J Physiol. 258:R918–R923. 1990.PubMed/NCBI
|
|
19
|
Kim HG, Hong SM, Kim SJ, et al:
Age-related changes in the activity of antioxidant and redox
enzymes in rats. Mol Cells. 16:278–284. 2003.PubMed/NCBI
|
|
20
|
Liu R and Choi J: Age-associated decline
in gamma-glutamylcysteine synthetase gene expression in rats. Free
Radic Biol Med. 28:566–574. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mosoni L, Breuillé D, Buffière C, Obled C
and Mirand PP: Age-related changes in glutathione availability and
skeletal muscle carbonyl content in healthy rats. Exp Gerontol.
39:203–210. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ohrloff C, Hockwin O, Olson R and Dickman
S: Glutathione peroxidase, glutathione reductase and superoxide
dismutase in the aging lens. Curr Eye Res. 3:109–115. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhu Y, Carvey PM and Ling Z: Age-related
changes in glutathione and glutathione-related enzymes in rat
brain. Brain Res. 1090:35–44. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lambertucci RH, Levada-Pires AC, Rossoni
LV, Curi R and Pithon-Curi TC: Effects of aerobic exercise training
on antioxidant enzyme activities and mRNA levels in soleus muscle
from young and aged rats. Mech Ageing Dev. 128:267–275. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Siu PM, Pistilli EE, Murlasits Z and Alway
SE: Hindlimb unloading increases muscle content of cytosolic but
not nuclear Id2 and p53 proteins in young adult and aged rats. J
Appl Physiol (1985). 100:907–916. 2006.PubMed/NCBI
|
|
26
|
Kayani AC, Morton JP and McArdle A: The
exercise-induced stress response in skeletal muscle: failure during
aging. Appl Physiol Nutr Metab. 33:1033–1041. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Franco AA, Odom RS and Rando TA:
Regulation of antioxidant enzyme gene expression in response to
oxidative stress and during differentiation of mouse skeletal
muscle. Free Radic Biol Med. 27:1122–1132. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Botta D, White CC, Vliet-Gregg P, et al:
Modulating GSH synthesis using glutamate cysteine ligase transgenic
and gene-targeted mice. Drug Metab Rev. 40:465–477. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dalton TP, Dieter MZ, Yang Y, Shertzer HG
and Nebert DW: Knockout of the mouse glutamate cysteine ligase
catalytic subunit (Gclc) gene: embryonic lethal when homozygous,
and proposed model for moderate glutathione deficiency when
heterozygous. Biochem Biophys Res Commun. 279:324–329. 2000.
View Article : Google Scholar
|
|
30
|
Yang Y, Dieter MZ, Chen Y, Shertzer HG,
Nebert DW and Dalton TP: Initial characterization of the
glutamate-cysteine ligase modifier subunit Gclm(−/−) knockout
mouse. Novel model system for a severely compromised oxidative
stress response. J Biol Chem. 277:49446–49452. 2002.PubMed/NCBI
|
|
31
|
Lee JI, Kang J and Stipanuk MH:
Differential regulation of glutamate-cysteine ligase subunit
expression and increased holoenzyme formation in response to
cysteine deprivation. Biochem J. 393:181–190. 2006. View Article : Google Scholar
|