Introduction
Osteoarthritis (OA) is a degenerative joint disease
characterized by the loss of chondrocyte function and extracellular
matrix (ECM) destruction (1). The
stability of articular cartilage depends on the biosynthetic
activities of chondrocytes, which synthesize appropriate ECM
molecules to maintain cartilage homeostasis (2,3).
Chondrocyte apoptosis alters the cartilage matrix synthesis,
leading to matrix degeneration and destruction, and finally to OA
(4). Cartilage ECM molecules
including type II collagen and sulfated proteoglycan play a crucial
role in regulating chondrocyte functions by facilitating
cell-matrix interactions (5). The
development and progression processes of OA are now considered to
involve inflammation (6).
Interleukin (IL)-1β, tumor necrosis factor (TNF)-α and nitric oxide
(NO) are produced by activated synoviocytes, mononuclear cells or
chondrocytes, and have been reported to induce apoptosis in
chondrocytes and ECM degradation (7,8). In
response to IL-1β, chondrocytes secrete matrix metalloproteinases
(MMPs), which induce chondrocyte apoptosis and inhibit ECM
biosynthesis (9,10). Additionally, the activity of MMPs
is also regulated by the inhibition of endogenous tissue inhibitors
of metalloproteinase (TIMPs), and the imbalance in the ratio of
TIMPs and MMPs results in continued destruction in OA (11).
Sanmiao formula (SM) is a compound prescription that
comprises three ingredients: Chinese Phellodendri Cortex,
Atractylodis Rhizoma and Achyranthis Bidentatae Radix, in a
specified ratio (12). SM has been
used in traditional Chinese medicine since the Ming Dynasty (15th
century common era) due to its function of clearing heat and
removing dampness. Currently SM and modified SM are commonly used
clinically for the treatments of gouty and rheumatoid arthritis
(13,14). However, the effects of SM on OA
remain uncharacterized. In the present study, the effects of SM on
articular cartilage damage were investigated, with a focus on
chondrocyte apoptosis and cartilage matrix degradation, as well as
the inflammatory response, using a rat model of OA.
Materials and methods
Herbal preparation
SM was prepared as described in the Chinese
Pharmacopeia of 2010 (12).
Atractylodis rhizome (600 g, baked with salt), Phellodendri Chinese
Cortex (400 g) and Achyranthis bidentatae Radix (200 g) were
purchased from Beijing Medicinal Herbs Co., Ltd. (Beijing, China),
pulverized to a fine powder, suspended in distilled water to a
concentration of 0.25 g/ml, and mixed well prior to administration.
The representative chemical compositions of atractylenolide
(0.0085%), berberine (0.734%), and β-ecdysone (0.012%) in SM were
determined by high-performance liquid chromatography analysis.
OA animal model and treatment
Ten-week-old male Sprague-Dawley rats (Animal
Science Laboratory of Peking University Health Science Department,
Beijing, China) were anesthetized with halothane, and subsequent to
being shaved and disinfected, the right knee joint was exposed
through a medial para-patellar approach. The patella was dislocated
laterally and the knee placed in full flexion, followed by anterior
cruciate ligament transection and medial meniscus resection (ACLT
plus MMx) using micro-scissors. In the sham-operated negative
controls, the right knee joint was exposed and incisions were
closed following subluxation of the patella and washing of the
joint surface with saline. The rats were randomly assigned to five
groups: ACLT plus MMx without treatment (model, n=12),
sham-operated (sham, n=12), and ACLT plus MMx rats treated with SM
intragastrically at a daily dose of 0.63, 1.25 or 2.50 g/kg (SM,
n=12 per dose) for three or six weeks. Dose calculations followed
guidelines correlating dose equivalents between humans and
laboratory animals on the basis of ratios of body surface area
(15); 2.5 g/kg is the dose
equivalent to a recommended dose of 0.4 g/kg in humans. Rats in the
model and sham groups received distilled water only. All animals
were maintained on a 12 h light/dark cycle under constant
temperature (24±2°C) and humidity (55±5%), and were allowed free
access to food and water. All procedures for the consideration of
animal welfare were reviewed and approved by the Ethical Committee
of the China Academy of Chinese Medical Sciences (Beijing,
China).
Histological analysis
Animals were sacrificed after three or six weeks of
treatment. The tibia and femur were dissected and fixed in 4%
paraformaldehyde for 24 h, decalcified in 10% ethylenediamine
tetraacetic acid (EDTA) and embedded in paraffin. Tissue sections
(4 μm) were mounted on common slides for staining with hematoxylin
and eosin, toluidine blue and Masson’s trichrome as described
previously (16,17). Cartilage histopathological features
were analyzed using the scoring system modified by Mankin et
al (18) (score range 0–12,
from normal to complete disorganization and hypocellularity).
Synovium histopathology was evaluated according to Yoshimi’s
histological grading (score range 0–18, between normal and most
severe reaction) (19). The
Image-Pro Plus 6.0 System (IPP) image analysis system (Media
Cybernetics, Rockville, MD, USA) was used for quantitative
analysis. The positive index was calculated as the integral of the
optical density. All sections were randomized and evaluated by a
trained observer who was blinded to the treatment groups.
Terminal deoxynucleotidyl
transferase-mediated dUTP nick end labeling (TUNEL)
Apoptotic cells in specimens were recognized using
In Situ Cell Apoptosis Detection kits (Boster Biological
Technology, Ltd., Wuhan, China) according to the manufacturer’s
instructions (20). Briefly,
deparaffinized sections were permeabilized in 0.1% Triton X-100,
incubated in fluorescein-labeled dUTP and terminal deoxynucleotidyl
transferase (TdT) mixture, then probed using an antifluorescein
antibody conjugated with alkaline phosphatase. Sections were
developed using substrate solution containing fast red. Sections
without primary TdT were used as negative controls for the TUNEL
staining. TUNEL-positive cells (apoptosis index, AI) in three
different areas were counted under an Olympus OX31 microscope
(Olympus, Tokyo, Japan). The Image-Pro Plus 6.0 System (IPP) image
analysis system was used for quantitative analysis.
Immunoblotting analysis
Paraffin sections (4 μm) of joint tissue were
mounted on poly-L-lysine-coated slides. The paraffin sections were
dewaxed by a routine method and incubated for 10 min with 3%
H2O2. Each section was incubated with
blocking serum (Vectastain® ABC kit, Vector
Laboratories, Burlingame, CA, USA) at room temperature for 30 min
and then with primary rabbit monoclonal antibody against type II
collagen (dilution 1/30; Beijing Biosynthesis Biotechnology Co.,
Ltd., Beijing, China), rabbit polyclonal antibody against MMP-13
(dilution 1/50; Abcam, Cambridge, UK) and rabbit monoclonal
antibody against TIMP-1 (dilution 1/80; Abcam) overnight at 4°C.
Sections incubated in phosphate-buffered saline (PBS) without
antibody served as negative controls. Following incubation with
biotinylated secondary antibody (Zhonshan Golden Bridge
Biotechnology, Beijing, China) and avidin-biotin complex reagent
containing horseradish peroxidase (Vector Laboratories Ltd.) for 30
min, the sections were then stained with 3,3′-diaminobenzidine
(DAB; Sigma, St. Louis, MO, USA). The Image-Pro Plus 6.0 System
(IPP) image analysis system was used for quantitative analysis.
Serum radioimmunoassay (RIA) and
enzyme-linked immunosorbent assay (ELISA) analysis
Animal blood was collected from the abdominal aorta
and the serum was analyzed for IL-1β and TNF-α by RIA, and for bone
morphogenetic protein (BMP) and transforming growth factor-β
(TGF-β) by ELISA. NO and inducible NO synthase (iNOS) were detected
by assay kits (Beijing 4A Biotech Co., Ltd., Beijing, China).
Chondrocyte culture and sample
collection
Normal human chondrocytes were obtained commercially
(ScienCell Research Laboratories, Carlsbad, CA, USA). The
chondrocytes were maintained in a specific chondrocyte medium
(ScienCell Research Laboratories) with 5% heat-inactivated fetal
bovine serum at 37°C in a humidified atmosphere of 95% air and 5%
CO2. At the third passage, cells were seeded at a
density of 1×105 cells per well in a six-well plate and
cultured to ~80% confluence, then made quiescent in serum-free
medium for 24 h. Chondrocytes were treated with SM at 0.83, 2.08
and 5.21 μg/ml for 1 h, and then treated with IL-1β at 10 ng/ml for
another 24 h of incubation. The controls consisted of chondrocytes
incubated in the culture medium without IL-1β or SM. The
chondrocytes were collected for further study.
Chondrocyte apoptosis by flow
cytometry
To quantify the percentage of cells undergoing
apoptosis, the Annexin V-FITC Apoptosis Detection kit (Nanjing
KeyGen Biotech. Co., Ltd., Nanjing, China) was used according to
the manufacturer’s instructions. Briefly, chondrocytes were
collected and the cells were washed twice with cold PBS. The cells
were then resuspended in binding buffer at a concentration of
5×106 cells/ml. Following incubation, 100 ml of the
solution was transferred to a 5 ml culture tube, and 5 μl Annexin
V-FITC and 10 μl propidium iodide (PI) were added. The tube was
gently vortexed and incubated for 15 min at room temperature in the
dark. At the end of the incubation, 400 ml binding buffer was
added, and the cells were analyzed immediately by flow cytometry. A
total of 1×106 chondrocytes were collected and the cells
were fixed using cold 75% ethanol at 4°C overnight, washed in PBS
and incubated with 50 μg/ml PI and 5 μg/ml RNase at room
temperature for 30 min. The cell cycle analysis was performed using
flow cytometry (provided by the Department of Cell Biology, School
of Medicine, Shanghai Jiao Tong University, Shanghai, China).
Western blot analysis
Following treatment, all chondrocytes were harvested
in cold PBS. The pellet was resuspended in lysis buffer (50 mm Tris
pH 8.0, 150 mm NaCl, 5 mm EDTA, 0.1% sodium dodecyl sulfate, 0.5%
nonyl phenoxypolyethoxylethanol-40) containing 10 mm
phenylmethylsulfonyl fluoride and 2 mg/ml aprotinin. Protein was
obtained to detect the levels of MMP-13 and TIMP-1 in the
chondrocytes by western blotting. The western blotting protocol and
semi-quantitative analysis were carried out as described previously
(21). The following antibodies
were used: Rabbit anti-MMP-13 monoclonal antibody (dilution 1/100,
Abcam), rabbit anti-TIMP-1 polyclonal antibody (dilution 1/50,
Boster Biological Technology, Ltd.) and GAPDH antibody (internal
control, rabbit polyclonal antibody, dilution 1/200, Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA). All experiments were
carried out in triplicate. The relative quantity of each antibody
was measured using AlphaEase FC (FluorChem FC2) software (Cell
Biosciences Inc., Santa Clara, CA, USA). The density ratio of
protein to GAPDH was calculated from the band density.
Quantitative polymerase chain reaction
(qPCR)
Following treatment, total RNA was extracted from
the chondrocytes with TRIzol® reagent (Invitrogen Life
Sciences, Carlsbad, CA, USA) according to the manufacturer’s
instructions. The total RNA (2 μg) was reverse transcribed to cDNA
using the High Capacity cDNA Reverse Transcription kit (Applied
Biosystems, Inc., Foster City, CA, USA) according to the
manufacturer’s instructions. The specific transcripts were
quantified by qPCR using a QuantiTect SYBR Green PCR kit (Qiagen,
Tokyo, Japan) and analyzed with an Applied Biosystems®
7500 Fast Real-Time PCR system (Applied Biosystems Inc.).
Gene-specific primers were used for MMP-13 (forward,
TGAGGATACAGGCAAGACTCT; reverse, CAATACGGT TACTCCAGATGC), TIMP-1
(forward, CTTCTGGCATCC TGTTGTTG; reverse, AGAAGGCCGTCTGTGGGT) and
GAPDH (forward, GAAGGTGAAGGTCGGAGTC; reverse,
GAAGATGGTGATGGGATTTC). The mRNA levels of MMP-13 and TIMP-1 were
normalized to the GAPDH mRNA level. PCR was performed at 95°C for
10 min, followed by 40 cycles of 95°C for 30 sec and 60°C for 1
min. The quantification data was analyzed with ABI
Prism® analysis software (Applied Biosystems, Inc.). The
relative mRNA expression was calculated with the comparative
threshold cycle method (22).
Statistical analysis
All data are expressed as the mean ± standard
deviation. The contents of collagen and proteoglycans were analyzed
with non-parametric statistics (Kruskal-Wallis one-way analysis of
variance). The other data were analyzed using a two-tailed
Student’s t-test, two-sample assuming unequal variance, within the
MS Excel 2007 software package (Microsoft Corp., Redmond, WA, USA).
Values were considered as significantly different if P<0.05.
Results
Effects of SM on the histopathology of
articular cartilage and synovium in the knee joints of ACLT plus
Mmx rats
To evaluate the anti-OA effect of SM, the isolated
knee joints from the five groups were analyzed microscopically. The
articular cartilage histopathological results are shown in Fig. 1. The sham group rats showed
smoothly surfaced articular cartilage and normal cellularity. By
contrast, the knee joints from the untreated rats with ACLT plus
MMx-induced OA revealed significant ongoing histopathological
changes, as indicated by surface irregularity, disorganization of
the articular cartilage with apparent cloning of chondrocytes in
the transitional and radial zones, and a disrupted tidemark. As
expected, the overall Mankin score was significantly increased by
ACLT plus MMx. These histomorphological changes in the cartilage
were reduced in the SM-treated OA animals, particularly with SM at
the dose of 2.5 g/kg, which significantly restored cartilage
morphology as indicated by normal cartilage surfaces with normal
cellularity in the transitional and radial zones, and a normal
tidemark. SM (2.5 g/kg) significantly decreased the overall
modified Mankin scores compared with those of the untreated OA
model rats at three and six weeks of treatment (both
P<0.001).
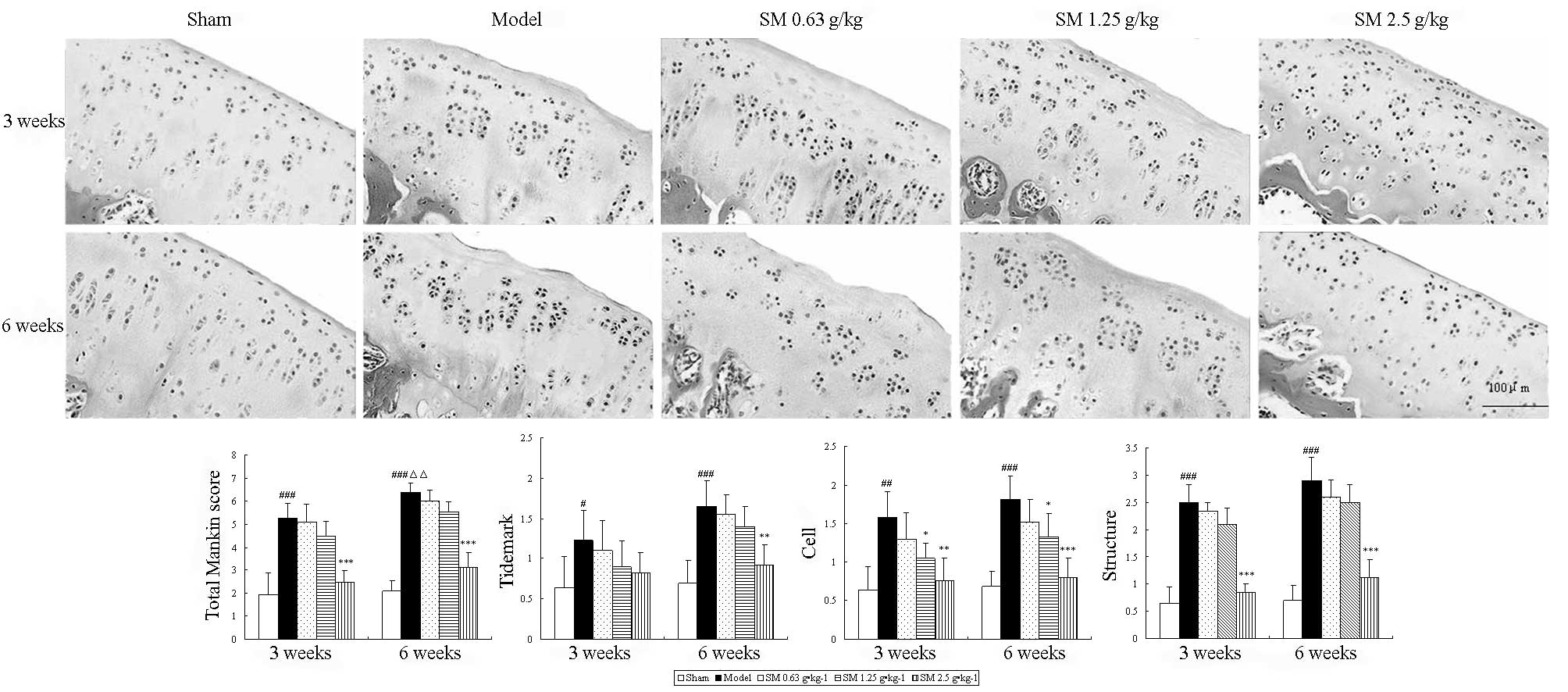 | Figure 1Effects of SM on cartilage histology
and Mankin score in the knee joints of rats following ACLT plus
Mmx. Cartilage histology was observed in ACLT plus MMx model and
sham-operated rats at the end of a three- or six-week treatment
period. Photomicrographs captured at ×200 magnification of
representative cartilage sections from one animal per treatment
group are shown: Sham, untreated ACLT plus MMx (model) and ACLT
plus MMx treated with SM at the doses of 0.63, 1.25 and 2.5 g/kg
(SM). At three or six weeks, significant histopathological changes
were evident in the model rats, as indicated by surface
irregularity, disorganization of articular cartilage with apparent
cloning of chondrocytes in the transitional and radial zones, and
an intact tidemark. Treatment with SM (2.5 g/kg) significantly
prevented damage to the cartilage structure, reduced cellular
abnormalities, and prevented change of the tidemark. The Mankin
scores of the SM (2.5 g/kg) group were significantly decreased
compared with those of the model group. Data are presented as the
mean ± standard deviation of samples from six rats per group.
P-values are from the Student’s t-test comparing the SM group with
the model group. *P<0.05, **P<0.01 and
***P<0.001 compared with the model group;
#P<0.05, ##P<0.01 and
###P<0.001 compared with the sham group;
ΔΔP<0.01 compared with the model group at the three
weeks. SM, SianMiao formula; ACLT plus MMx, anterior cruciate
ligament transection and medial meniscus resection. |
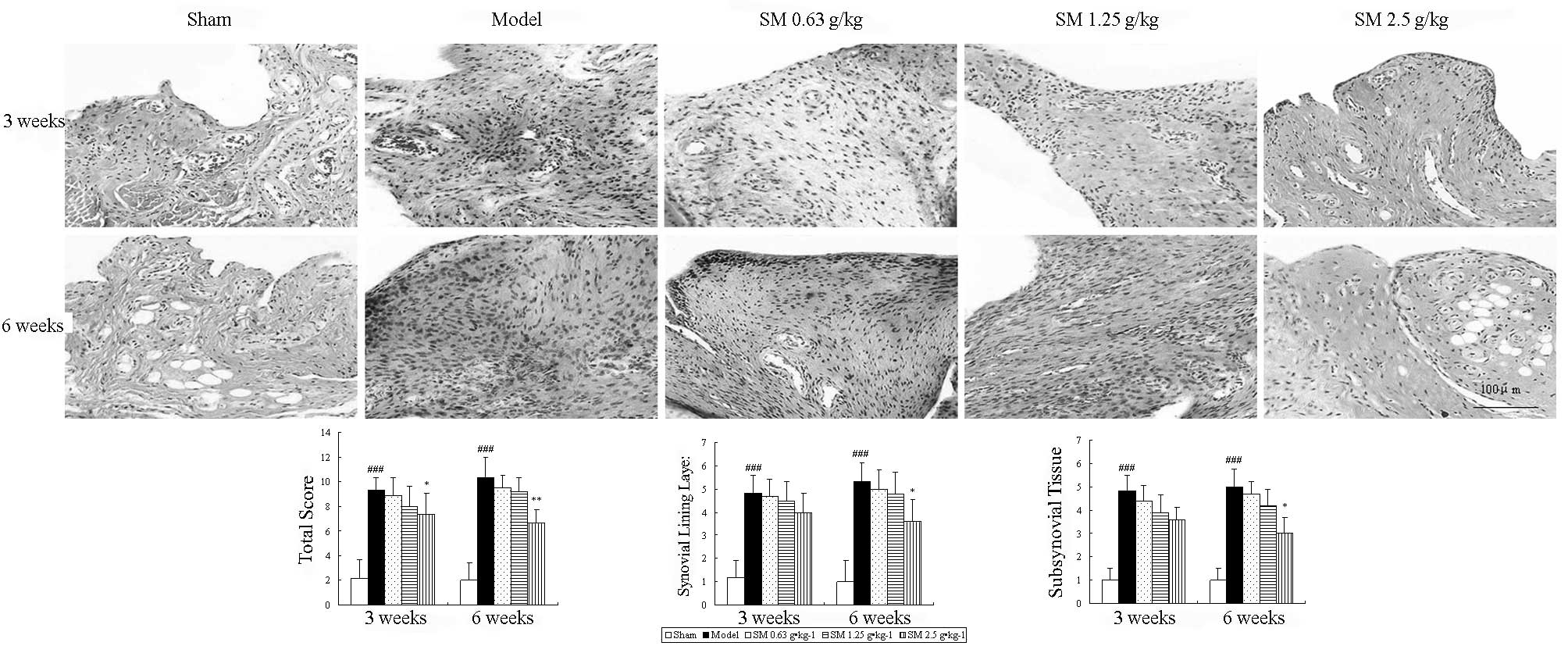
Histopathological features of the synovium are shown
in Fig. 2. The untreated ACLT plus
Mmx model rats exhibited increased hyperplasia of synovial lining
cells, hypertrophy of the synovial lining layer and an increase in
the infiltration of inflammatory cells in synovial tissue compared
with that observed in the sham rats. SM (2.5 g/kg) suppressed
hypertrophy of the synovial lining layer and inflammatory cellular
infiltration, and significantly reduced the histological severity
scores in rats following ACLT plus Mmx compared with those in the
untreated ACLT plus Mmx model rats at three and six weeks of
treatment (P<0.05 and P<0.01, respectively).
The above results provide evidence that SM exhibits
significant anti-OA activity. Notably, the largest changes in these
parameters were identified at the highest dose. These data prompted
further studies to elucidate the mechanism of SM (2.5 g/kg)
activity.
Effects of SM on chondrocyte apoptosis in
the knee joints of ACLT plus Mmx rats
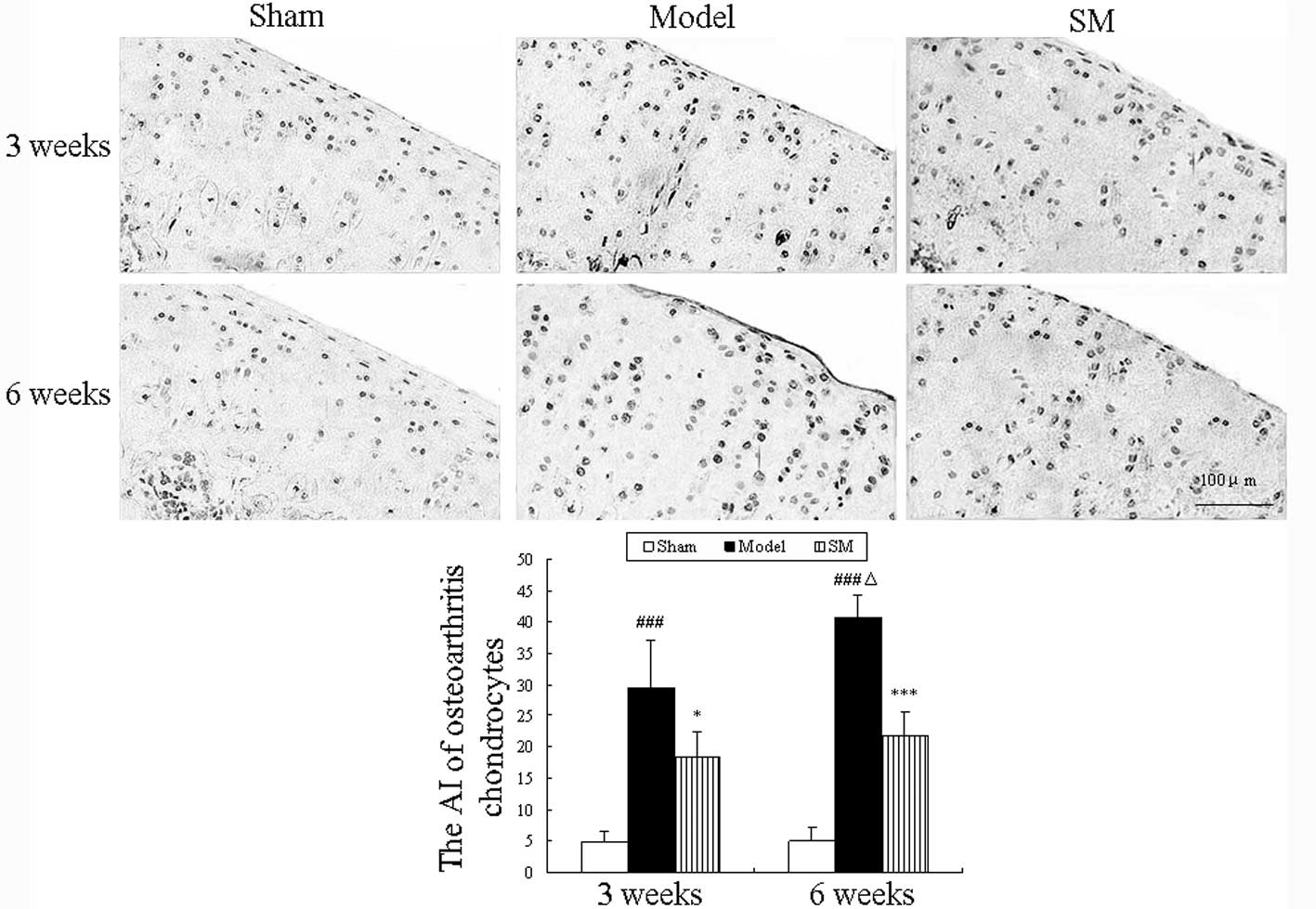
Apoptotic chondrocytes were observed using the TUNEL
assay, and the results are shown in Fig. 3. Increased numbers of apoptotic
chondrocytes were observed in the cartilage of untreated ACLT plus
Mmx model rats and the percentages of apoptotic chondrocytes were
29.5 and 40.75% at three and six weeks, respectively. Compared with
the untreated model group, SM (2.5 g/kg) treatment significantly
decreased the percentage of apoptotic chondrocytes at three and six
weeks (P<0.05 and P<0.01, respectively) and caused a 19%
reduction in the proportion of TUNEL-positive cells among the total
cells at six weeks compared with that in the untreated group.
Effects of SM on cartilage matrix in the
knee joints of ACLT plus Mmx rats
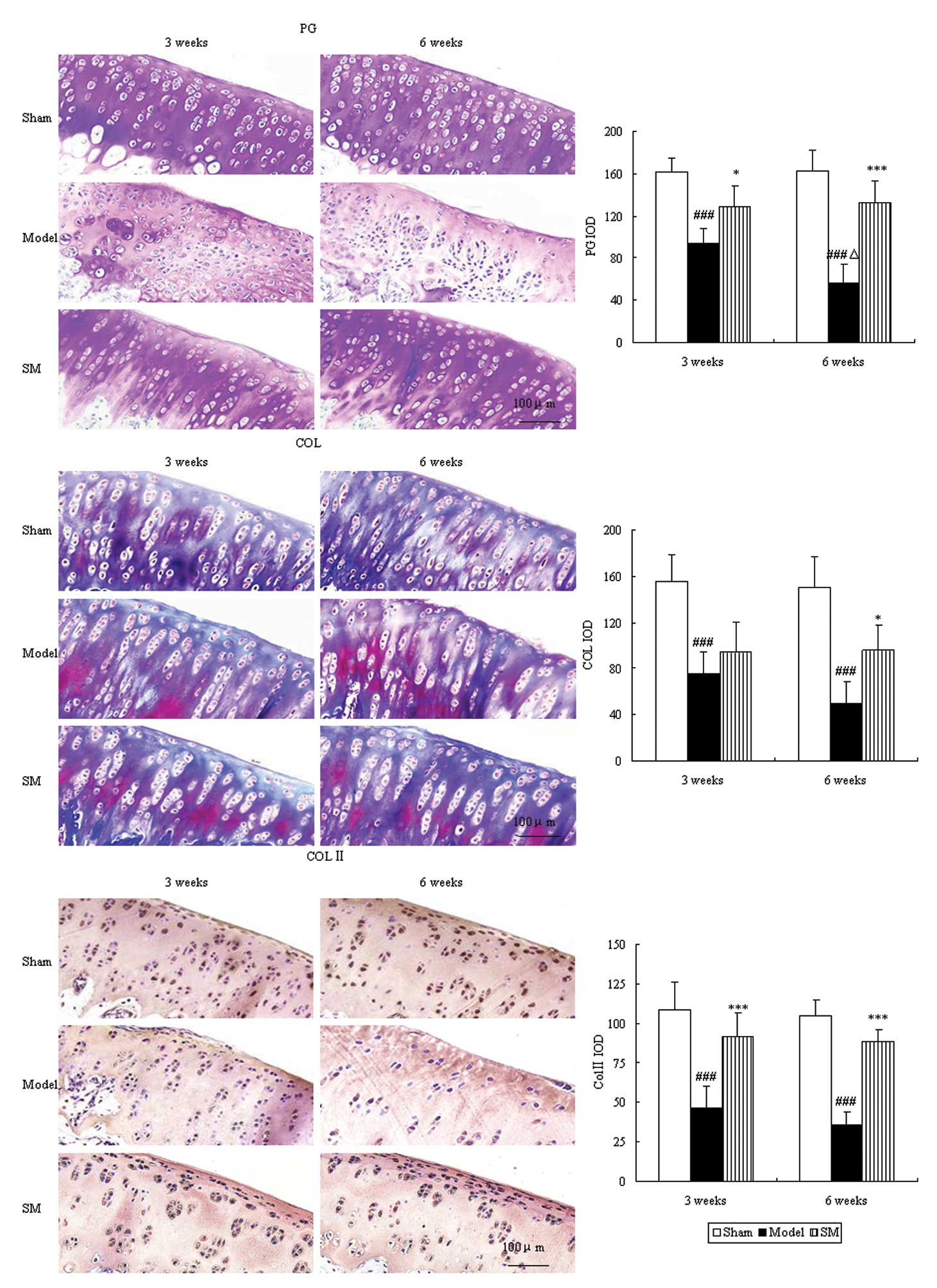
Proteoglycan and collagen are major constituents of
articular cartilage matrix, which were evaluated by toluidine blue
and Masson’s trichrome staining. The control group showed intensive
toluidine blue and Masson’s trichrome staining. In the untreated
ACLT plus Mmx model rats, toluidine blue and Masson’s trichrome
staining shown a significant ongoing reduction, which indicated
loss of proteoglycan and collagen. Furthermore,
immunohistochemistry revealed a significant reduction in type II
collagen levels. Treatment with SM (2.5 g/kg) significantly
inhibited cartilage matrix degradation, as indicated by attenuation
of the loss of proteoglycan and collagen and upregulation of the
expression of type II collagen at three and six weeks. SM (2.5
g/kg) induced ~2.3-, 1.9- and 2.5-fold increases in the expression
levels of proteoglycan (P<0.001), collagen (P<0.05) and type
II collagen (P<0.001), respectively, compared with those of
untreated ACLT plus Mmx model rats at six weeks (Fig. 4).
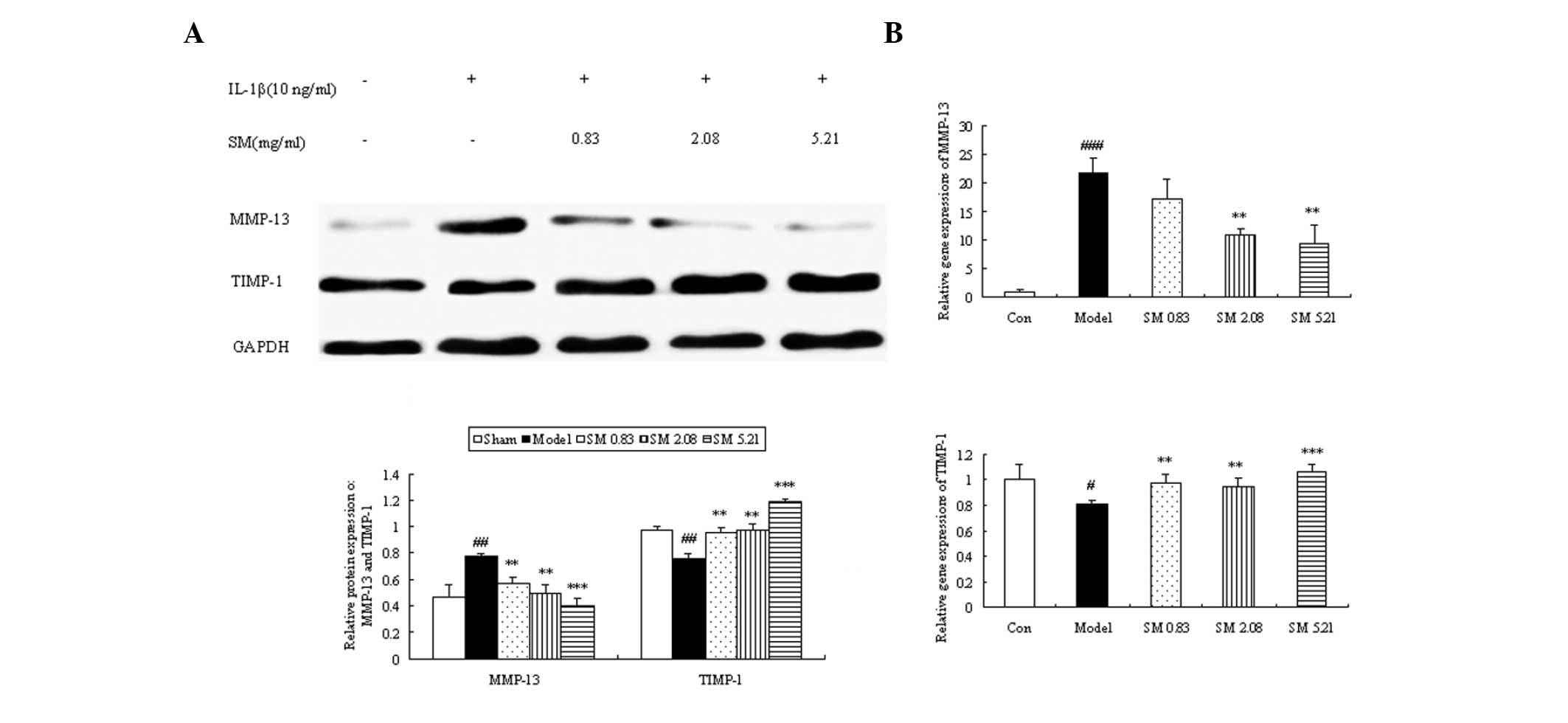
Effects of SM on MMP-13 and TIMP-1
expression levels in the knee joints of ACLT plus Mmx rats
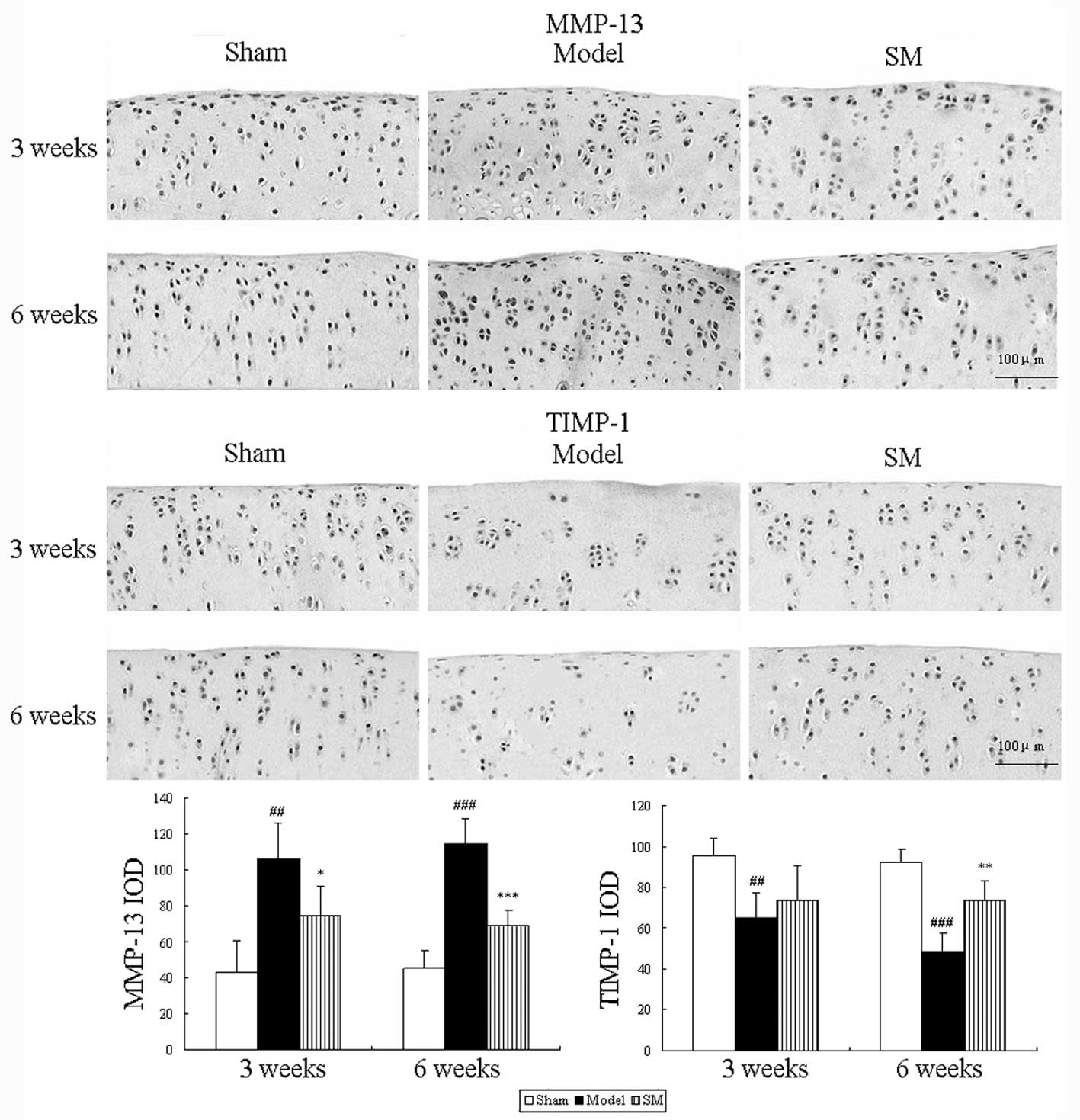
The effects of SM (2.5 g/kg) treatment on the
expression levels of MMP-13 and TIMP-1 in rats following ACLT plus
Mmx were assessed by immunohistochemistry. Representative joint
sections from all groups and the findings from the quantitative
analysis are shown in Fig. 5. ACLT
plus MMx induced increased the expression levels of MMP-13 and
decreased the expression levels of TIMP-1 in the joint. Treatment
with SM (2.5 g/kg) induced significant differences compared with
untreated ACLT plus Mmx model rats at three and six weeks. SM
decreased the expression of MMP-13 by 39% (P<0.001) and
increased that of TIMP-1 by 51% (P<0.01) compared with that in
the untreated ACLT plus Mmx model rats at six weeks. MMP-13 and
TIMP-1 were expressed in the chondrocytes of the knee joint in all
groups. The positive staining was predominantly cytoplasmic or
cytosolic in the chondrocytes. These results support the indication
that SM mediates anti-OA activity through the MMP/TIMP system.
Effects of SM on the levels of IL-1β,
TNF-a, NO, iNOS, BMP and TGF-β in the serum of ACLT plus Mmx
rats
To determine whether SM induced changes in the
levels of inflammation-related cytokines, chondrocytes and
cartilage matrix metabolism-related cytokines in serum, the levels
of IL-1β, TNF-α, NO, iNOS, BMP and TGF-β in serum were examined.
The serum levels of IL-1β, TNF-α, NO and iNOS exhibited a
significant ongoing increase. This was significantly suppressed by
SM with the following reductions at six weeks of treatment: IL-1β,
48%, P<0.05; TNF-α, 31%, P<0.05; NO, 18%, P<0.01 and iNOS,
23%, P<0.05. The levels of BMP and TGF-β were significantly
decreased in the model group compared with those in the sham group
during the treatment period. Treatment with SM (2.5 g/kg) caused
significant increases in the levels of BMP-2 (P<0.05) and
TGF-β1 (P<0.01), with 55 and 43% increases
respectively at six weeks of treatment (Fig. 6).
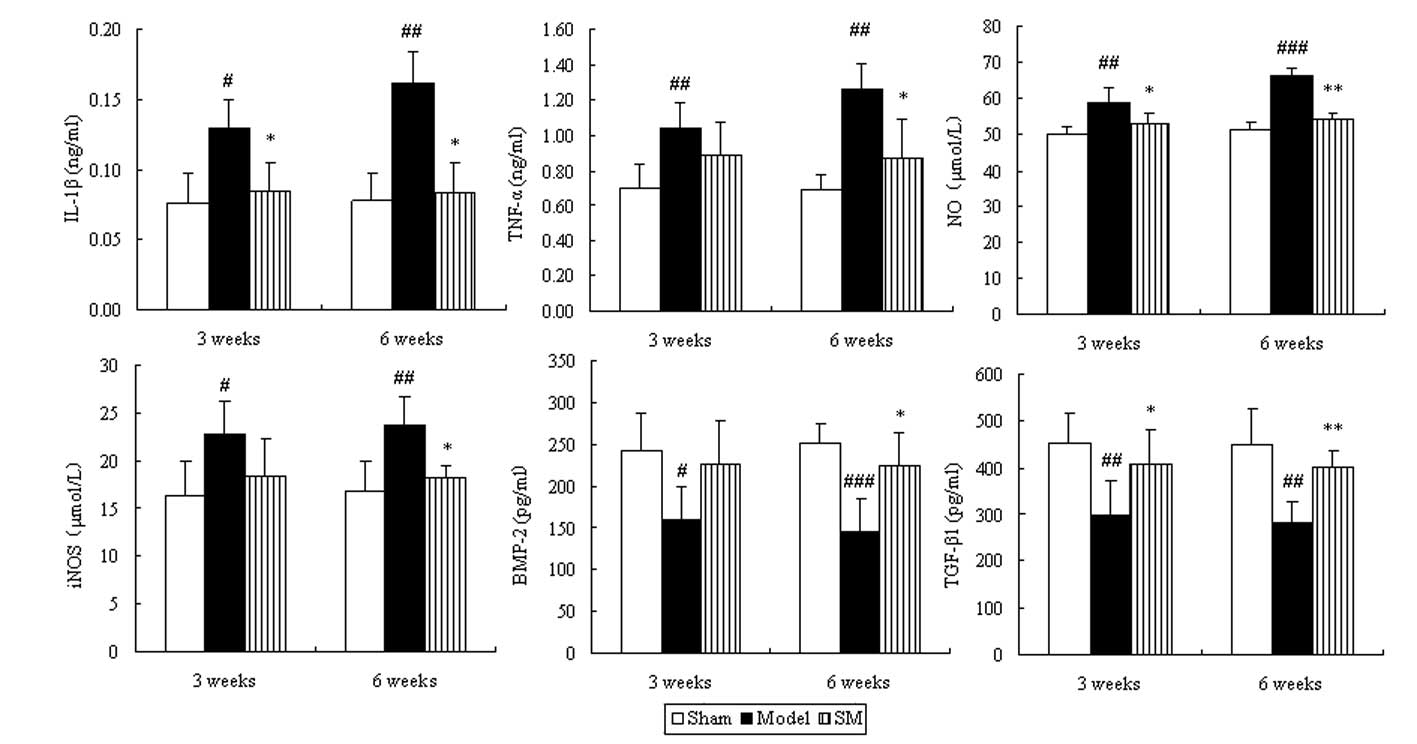 | Figure 6Effects of SM on IL-1β, TNF-α, NO,
iNOS, BMP-2 and TGF-β1 in the serum of rats with
osteoarthritis. Blood was obtained from the abdominal aorta at
three and six weeks of treatment following the anterior cruciate
ligament transection and medial meniscus resection and sham
surgeries. The serum was used for the assays of IL-1β, TNF-α, NO,
iNOS, BMP-2 and TGF-β1. Data are presented as the mean ±
standard deviation of samples from six rats in each group.
*P<0.05 and **P<0.01 compared with the
model group; #P<0.05, ##P<0.01 and
###P<0.001 compared with the sham group. SM, SanMiao
formula; IL-1β, interleukin-1β; TNF-α, tumor necrosis factor-α; NO,
nitric oxide; iNOS, inducible NO synthase; BMP-2, bone
morphogenetic protein-2; TGF-β1; transforming growth
factor-β1. |
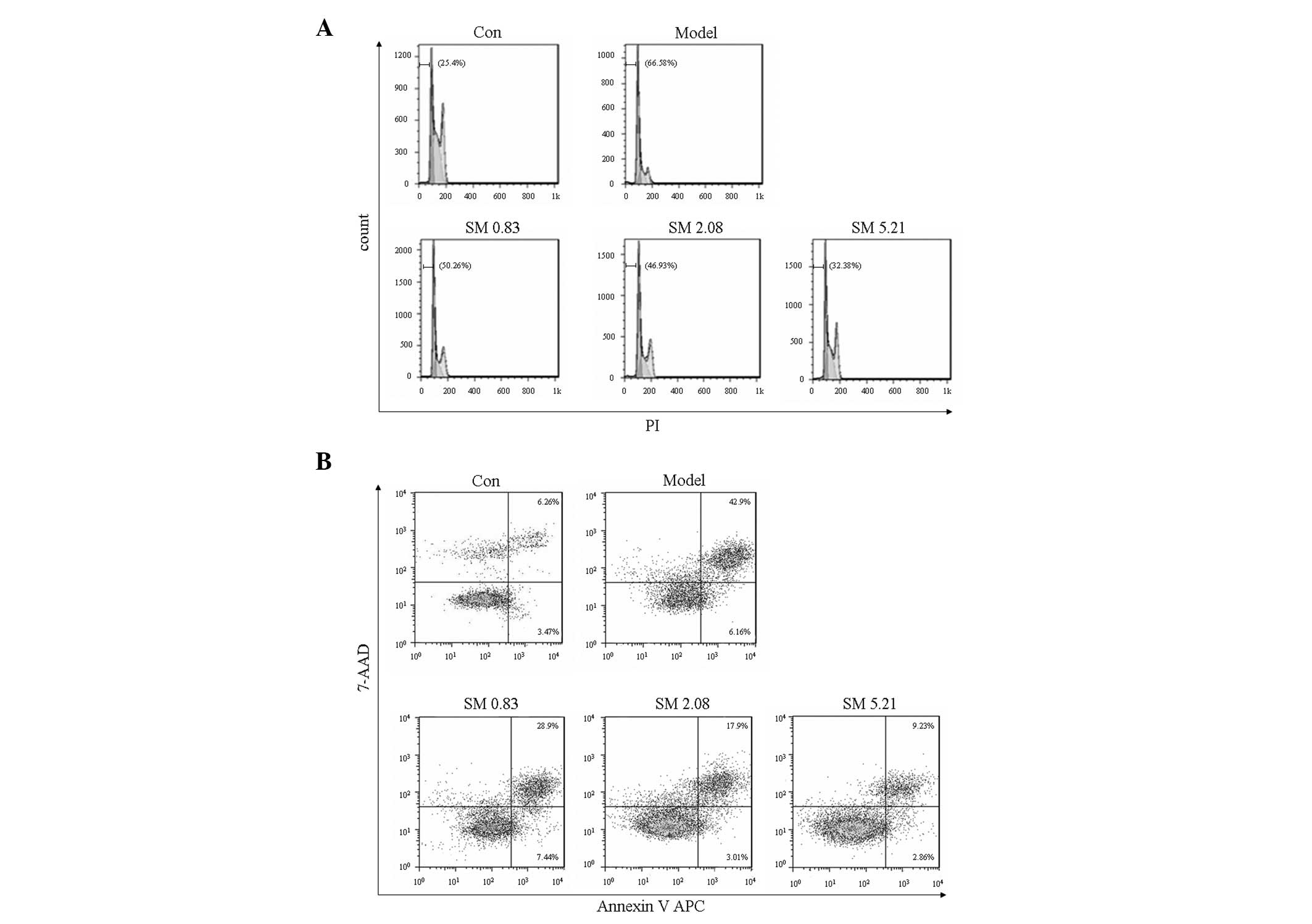
Effects of SM on chondrocyte apoptosis in
the flow cytometric analysis
IL-1β increased the proportion of apoptotic sub-G1
phase cells. SM significantly decreased the proportion of apoptotic
sub-G1 phase cells in a concentration-dependent manner as indicated
by PI staining using flow cytometry. SM at 5.21 μg/ml induced a
32.38% reduction in apoptotic sub-G1 phase cells (Fig. 7A). The inhibitory effect of SM on
apoptosis was confirmed using Annexin V staining to detect the
externalization of phosphatidylserine on the cell membrane. SM
markedly decreased early and late apoptotic cells, and SM at 5.21
μg/ml induced a 34% reduction in early and late apoptotic cells
(Fig. 7B).
Effects of SM on the production and gene
expression of MMP-13 and TIMP-1 in human chondrocytes
The expression of MMP-13 was significantly
downregulated when the chondrocytes were incubated with SM at 0.83,
2.08 or 5.21 μg/ml. The downregulating effects were augmented as
the concentration of SM increased, and SM at 5.21 μg/ml induced a
49% reduction in the level of MMP-13 (P<0.001) compared with
that in untreated IL-1β-induced chondrocytes. The expression of
TIMP-1 was clearly upregulated following incubation with SM at all
three concentrations. The largest increases were found at 5.21
μg/ml SM (P<0.001) with a 1.6-fold upregulation (Fig. 8A). To further validate the above
mechanisms, the effects of SM on the mRNA expression of MMP-13 and
TIMP-1 in IL-1β-induced chondrocytes were also assessed by qPCR. SM
significantly inhibited the IL-1β-induced gene expression of MMP-13
in a concentration-dependent manner. SM at the highest
concentration induced a 57% reduction in MMP-13 mRNA levels
(P<0.01) compared with those in the model group. SM upregulated
the gene expression of TIMP-1, and SM at 5.21 μg/ml induced a
1.3-fold upregulation in TIMP-1 mRNA levels compared with those in
the model group (P<0.001; Fig.
8B).
Discussion
OA is a degenerative joint disease with multiple
underlying pathogenic mechanisms caused by various risk factors.
The loss of chondrocyte function, degradation of the ECM and
inflammation play crucial roles in OA progression, and thus
inhibiting the development of these factors is an important issue
for the treatment of OA. To the best of our knowledge, the effects
of SM on OA have not been characterized. In the present study, the
anti-OA effect of SM was evaluated using a classical ACLT plus MMx
rat model. The new findings demonstrate than SM significantly
inhibited articular cartilage damage and synovial inflammation in
rats following ACLT plus Mmx, by inhibiting chondrocyte apoptosis,
cartilage matrix degradation and the release of proinflammatory
cytokines and inflammatory mediators.
Chondrocyte apoptosis plays an key role in the
degeneration and degradation of articular cartilage in cases of OA.
Reduced cellularity is a characteristic feature of OA cartilage and
apoptosis has been proposed as an underlying cause of the
hypocellularity (23). In the
present study, it was shown that the cell scores in knee cartilage
following ACLT plus MMx were markedly increased compared with those
in nonarthritic control cartilage, and that cell scores correlated
positively with the percentage of apoptotic chondrocytes,
suggesting that the reduced cellularity in OA cartilage may be at
least partially attributable to cell death by apoptosis. The
results from the TUNEL analysis showed that SM suppressed
chondrocyte apoptosis in the superficial and middle zones of
cartilage of ACLT plus Mmx rats. These results are confirmed by the
in vitro experiments in the present study, which revealed
that SM inhibited the increase in the proportion of early and late
apoptotic cells and sub-G1 phase cells in IL-1β-induced
chondrocytes, suggesting that SM prevents the degeneration and
degradation of articular cartilage by inhibiting chondrocyte
apoptosis.
Cartilage matrix consists largely of proteoglycan
and collagen; the loss of proteoglycan occurs early in the process
of cartilage degeneration and is followed by the catabolism of
collagen fibrils (24). The
tensile strength of articular cartilage is provided by a network of
fibrils which exhibit a slow turnover that may occur over years,
rendering collagen damage a particular problem (25). Type II collagen is known to be the
most common type of collagen in hyaline articular cartilage
(26). MMP-13, a primary
collagenase in OA, degrades type II collagen and is inhibited by
TIMP-1 (27,28). Under normal conditions, TIMPs bind
to active MMPs in a 1:1 ratio to make an inactive complex. An
imbalance in the ratio of TIMPs to MMPs causes the continued matrix
destruction in OA (11). In the
present study, it was demonstrated that SM significantly interfered
with the OA-induced expression of MMP-13, while augmenting that of
TIMP-1 in the joints of OA model rats. These results are supported
by the in vitro experiments in the present study, which
revealed that the IL-1β-augmented expression of MMP-13 was
inhibited and the protein and gene expression levels of TIMP-1 were
upregulated by SM in the IL-1β-induced chondrocytes. A previous
study indicated that the important ingredient berberine in SM
decreased glycosaminoglycan release, inhibited the expression of
MMP-1, -3 and -13, and increased the level of TIMP-1 at the mRNA
level in IL-1β-induced rat articular chondrocytes (29). Therefore, regulating the balance of
MMPs/TIMPs is one of the mechanisms by which SM prevents cartilage
matrix degradation.
The progression of OA is currently considered to be
associated with inflammation in the early stages of the disease.
Among the proinflammatory cytokines involved in OA, IL-1β and TNF-α
are considered the major participants. In patients or animals with
OA, levels of IL-1β and TNF-α are elevated in the synovial fluid
and serum (30). NO, a gaseous
free radical, is an important inducer of apoptosis as well as the
upregulation of MMP-13 (31) and
is synthesized by iNOS. The results of the present study showed
that SM significantly decreased the secretion of IL-1β and TNF-α in
serum. Additionally, SM markedly inhibited the release of NO in the
circulation through suppression of iNOS production. Sun et
al (32) reported that SM
effectively inhibited arthrocele and synovial inflammation in rats
with adjuvant arthritis (AA). Thus, SM may be able to suppress the
inflammatory response of arthritis. However, the dose of SM used in
the rats with AA was higher than that used in the OA model rats in
the present study. All of the three ingredients of SM contain
anti-inflammatory substances, including berberine and β-eudesmol
(33,34). SM may inhibit the inflammatory
response in OA by interfering with the secretion of proinflammatory
cytokines and inflammatory mediators. TGF-β is one of the most
potent mediators of cartilage matrix synthesis (35). It upregulates the expression of
several types of collagens and proteoglycans. Other members of the
TGF-β superfamily, BMPs, are also known to stimulate cartilage
matrix synthesis. In particular, BMP-2 has been shown to stimulate
the anabolic activity of chondrocytes as well as being present in
articular cartilage (36). The
results of the present study revealed that SM may exert anti-OA
activity by upregulating the levels of TGF-β and BMP-2.
In conclusion, SM effectively exerted anti-OA
activity in ACLT plus Mmx model rats. The effect of prevention of
cartilage articular damage was mainly via the direct suppression of
chondrocyte apoptosis and cartilage matrix degradation, and
interference with the secretion of proinflammatory cytokines and
inflammatory mediators as well as chondrocytes and cartilage matrix
metabolism-related cytokines in the OA model rats. Furthermore, the
blocking of chondrocyte apoptosis was achieved by inhibiting the
proportion of early and late apoptotic and sub-G1 phase cells. The
effect of preventing cartilage matrix degradation was achieved by
the suppression of MMP-13 expression, simultaneous upregulation of
TIMP-1 production, and maintenance of the MMP to TIMP balance in
the knee joints and chondrocytes. This study, to the best of our
knowledge, provides the first evidence that SM can effectively
treat OA, as part of an ongoing effort to identify novel and potent
agents for the prevention and treatment of OA.
Acknowledgements
This study was supported by grants from the project
of the National Natural Science Foundation of China (no.
81072900).
References
|
1
|
Loeser RF: Aging and osteoarthritis: the
role of chondrocyte senescence and aging changes in the cartilage
matrix. Osteoarthritis Cartilage. 17:971–979. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Goldring MB: The role of the chondrocyte
in osteoarthritis. Arthritis Rheum. 43:1916–1926. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Roughley PJ: Articular cartilage and
changes in arthritis: noncollagenous proteins and proteoglycans in
the extracellular matrix of cartilage. Arthritis Res. 3:342–347.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Aigner T and Kim HA: Apoptosis and
cellular vitality: issues in osteoarthritic cartilage degeneration.
Arthritis Rheum. 46:1986–1996. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Eyre D: Collagen of articular cartilage.
Arthritis Res. 4:30–35. 2002. View
Article : Google Scholar
|
|
6
|
Kapoor M, Martel-Pelletier J, Lajeunesse
D, Pelletier JP and Fahmi H: Role of proinflammatory cytokines in
the pathophysiology of osteoarthritis. Nat Rev Rheumatol. 7:33–42.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Aizawa T, Kon T, Einhorn TA and
Gerstenfeld LC: Induction of apoptosis in chondrocytes by tumor
necrosis factor-alpha. J Orthop Res. 19:785–796. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fernandes JC, Martel-Pelletier J and
Pelletier JP: The role of cytokines in osteoarthritis
pathophysiology. Biorheology. 39:237–246. 2002.PubMed/NCBI
|
|
9
|
Attur M, Al-Mussawir HE, Patel J, et al:
Prostaglandin E2 exerts catabolic effects in osteoarthritis
cartilage: evidence for signaling via the EP4 receptor. J Immunol.
181:5082–5088. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Abramson SB: Osteoarthritis and nitric
oxide. Osteoarthritis Cartilage. 16(Suppl 2): S15–S20. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Burger D, Rezzonico R, Li JM, et al:
Imbalance between interstitial collagenase and tissue inhibitor of
metalloproteinases 1 in synoviocytes and fibroblasts upon direct
contact with stimulated T lymphocytes: involvement of
membrane-associated cytokines. Arthritis Rheum. 41:1748–1759. 1998.
View Article : Google Scholar
|
|
12
|
China Pharmacopoeia Committee. Chinese
Pharmacopoeia. Chemical Industry Press; Beijing, China: 2010, (In
Chinese).
|
|
13
|
Zhang YH, Song YJ and Jia Y: Observation
of curative effect on the treatment of acute gouty arthritis with
SM in 45 cases. Yunnan Zhong Yi Zhong Yao Za Zhi. 24(4): 5–6.
2003.(In Chinese).
|
|
14
|
Liang GX and Duan JM: The curative effect
observation on 68 cases of rheumatoid arthritis treated with Si
long San Miao Formula. Guangming Journal of Chinese Medicine.
22:86–87. 2007.(In Chinese).
|
|
15
|
Guidance for Industry: Estimating the
maximum safe starting dose in initial clinical trials for
therapeutics in adult healthy volunteers. US Department of Health
and Human Services, Food and Drug Administration, Center for Drug
Evaluation and Research (CDER); Rockville, MD: 2005
|
|
16
|
Hayami T, Pickarski M, Wesolowski GA, et
al: The role of subchondral bone remodeling in osteoarthritis:
reduction of cartilage degeneration and prevention of osteophyte
formation by alendronate in the rat anterior cruciate ligament
transection model. Arthritis Rheum. 50:1193–1206. 2004. View Article : Google Scholar
|
|
17
|
Gruber HE, Marshall GJ, Nolasco LM,
Kirchen ME and Rimoin DL: Alkaline and acid phosphatase
demonstration in human bone and cartilage: effects of fixation
interval and methacrylate embedments. Stain Technol. 63:299–306.
1988.PubMed/NCBI
|
|
18
|
Mankin HJ, Dorfman H, Lippiello L and
Zarins A: Biochemical and metabolic abnormalities in articular
cartilage from osteo-arthritic human hips. II Correlation of
morphology with biochemical and metabolic data. J Bone Joint Surg
Am. 53:523–537. 1971.PubMed/NCBI
|
|
19
|
Yoshimi T, Kikuchi T, Obara T, et al:
Effects of high-molecular-weight sodium hyaluronate on experimental
osteoarthrosis induced by the resection of rabbit anterior cruciate
ligament. Clin Orthop Relat Res. 296–304. 1994.PubMed/NCBI
|
|
20
|
Kim KM, Kim JM, Yoo YH, Kim JI and Park
YC: Cilostazol induces cellular senescence and confers resistance
to etoposide-induced apoptosis in articular chondrocytes. Int J Mol
Med. 29:619–624. 2012.PubMed/NCBI
|
|
21
|
Xu Y, Zhang ZJ, Geng F, et al: Treatment
with Qing’E, a kidney-invigorating Chinese herbal formula,
antagonizes the estrogen decline in ovariectomized mice.
Rejuvenation Res. 13:479–488. 2010.
|
|
22
|
Pfaffl MW: A new mathematical model for
relative quantification in real-time RT-PCR. Nucleic Acids Res.
29:e452001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kühn K, D’Lima DD, Hashimoto S and Lotz M:
Cell death in cartilage. Osteoarthritis Cartilage. 12:1–16.
2004.
|
|
24
|
Jubb RW and Fell HB: The breakdown of
collagen by chondrocytes. J Pathol. 130:159–167. 1980. View Article : Google Scholar
|
|
25
|
Rosenberg AE: Bones, joints, and soft
tissue tumors. Robbins Pathologic Basis of Disease. Cotran RS,
Kumar C and Collins T: 6th edition. Philadelphia, PA: WB Saunders;
pp. 12531999
|
|
26
|
Naito K, Watari T, Muta T, et al:
Low-intensity pulsed ultrasound (LIPUS) increases the articular
cartilage type II collagen in a rat osteoarthritis model. J Orthop
Res. 28:361–369. 2010.PubMed/NCBI
|
|
27
|
Goldring MB, Otero M, Plumb DA, et al:
Roles of inflammatory and anabolic cytokines in cartilage
metabolism: signals and multiple effectors converge upon MMP-13
regulation in osteoarthritis. Eur Cell Mater. 21:202–220.
2011.PubMed/NCBI
|
|
28
|
Wetzel M, Li L, Harms KM, et al: Tissue
inhibitor of metalloproteinases-3 facilitates Fas-mediated neuronal
cell death following mild ischemia. Cell Death Differ. 15:143–151.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hu PF, Chen WP, Tang JL, Bao JP and Wu LD:
Protective effects of berberine in an experimental rat
osteoarthritis model. Phytother Res. 25:878–885. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hashimoto S, Nishiyama T, Hayashi S, et
al: Role of p53 in human chondrocyte apoptosis in response to shear
strain. Arthritis Rheum. 60:2340–2349. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Del Carlo M Jr and Loeser RF: Nitric
oxide-mediated chondrocyte cell death requires the generation of
additional reactive oxygen species. Arthritis Rheum. 46:394–403.
2002.PubMed/NCBI
|
|
32
|
Sun B, Lv L, Lu ZX and Yang SY: Study on
drug-guide effect of Achyranthes bidentata in Sanmiao pill in
arthritic rats. Zhongguo Zhong Yao Za Zhi. 33:2946–2949. 2008.(In
Chinese).
|
|
33
|
Xiao HB, Sun ZL, Zhang HB and Zhang DS:
Berberine inhibits dyslipidemia in C57BL/6 mice with
lipopolysaccharide induced inflammation. Pharmacol Rep. 64:889–895.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Seo MJ, Kim SJ, Kang TH, et al: The
regulatory mechanism of β-eudesmol is through the suppression of
caspase-1 activation in mast cell-mediated inflammatory response.
Immunopharmacol Immunotoxicol. 33:178–185. 2011.
|
|
35
|
Shuler FD, Georgescu HI, Niyibizi C, et
al: Increased matrix synthesis following adenoviral transfer of a
transforming growth factor beta1 gene into articular chondrocytes.
J Orthop Res. 18:585–592. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Aigner T, Soeder S and Haag J: IL-1beta
and BMPs - interactive players of cartilage matrix degradation and
regeneration. Eur Cell Mater. 12:49–56. 2006.PubMed/NCBI
|