Introduction
Vasculogenic mimicry (VM) is a unique blood supply
pattern present in malignant tumours. VM refers to the remodelling
of tumour cells into a luminal-like structure for blood flow,
although no endothelial cells are present on the inner luminal
surface. The formation of VM is associated with tumour cell
plasticity and the varying tumour microenvironment (1). The mechanisms underlying its
formation remain unclear, but a variety of proteins and
microenvironmental factors are known to contribute to VM (2–4).
The changing tumour microenvironment has a certain
promoting effect on VM. Hypoxia and tumour extracellular matrix
remodelling are also actively involved in VM formation (5,6).
Thus, inducing hypoxic conditions can promote the biological
behaviour and VM of malignant glioma (SHG-44) cells.
Hypoxia-inducible factor-1α (HIF-1α)-vascular endothelial growth
factor (VEGF)-Ephrin type-A receptor 2 (EphA2)-matrix
metalloproteinase (MMP)-VM may be a key channel for VM formation
under hypoxic conditions. In addition to HIF-1α and VEGF, which
contribute to the mechanisms of VM formation, another known factor
that promotes the VM formation of highly malignant glioma cells is
the hypoxic condition (7,8). The
3-(5-hydroxymethyl-2-furyl)-1-benzylindazole (YC-1)-induced
inhibition of HIF-1α has been found to downregulate the VM
formation of glioma cells (9)
under hypoxic conditions; other pathways, aside from the HIF-1α
pathway, may also affect the VM formation of glioma cells (10–12).
The overexpression of the anti-apoptotic protein
B-cell lymphoma 2 (Bcl-2) is considered to be an important factor
in the generation, metastasis and angiogenesis of a variety of
malignant tumours, including malignant glioma. Bcl-2 reportedly
affects the VM formation of human melanoma cells by regulating
VE-cadherin under an induced hypoxic condition (13–15).
In the present study, chemical hypoxia was used to induce a hypoxic
environment for human malignant glioma U87 cells, and the effects
of the Bcl-2-specific inhibitor ABT-737 on the VM formation of the
U87 cells was subsequently observed. The aim of the study was to
explore the function and mechanism of Bcl-2 in VM formation under
hypoxic conditions.
Materials and methods
Cell culture and establishment of
model
Human malignant glioma U87 cells were obtained from
the Neurosurgical Institute of Zhujiang Hospital (Guangzhou,
China). Matrigel™ (Becton-Dickinson, San Jose, CA, USA) and foetal
bovine serum (FBS; Gibco-BRL, Grand Island, NY, USA) were mixed at
a ratio of 1:1 and were added to 24-well plates at 20 μl/well.
Subsequent to solidifying the glue for 20 min at room temperature,
1×104/well U87 cells were added to the plates and then
incubated at 37°C for adherence. When the adherent cells reached
80% confluence, the medium was replaced by a serum-free medium. The
cells were subsequently divided into four groups: Control (blank),
YC-1, ABT-737 and YC-1 + ABT-737. A total of 50 μmol/l YC-1 (Cayman
Chemical Co., Ann Arbor, MI, USA) and/or 50 μmol/l ABT-737
(Shanghai Han Hong Biotechnology Co., Ltd., Shanghai, China) was
added to the wells accordingly. After 5 min, 100 μmol/l cobalt
chloride (CoCl2; Sigma-Aldrich, St. Louis, MO, USA) was
added to each group, and the plates were incubated for 24 h.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The mRNA expression levels of HIF-1α, MMP-2, MMP-14
and Bcl-2 in the cells of each group were analysed using RT-qPCR.
Following incubation with CoCl2 for 24 h, the cells were
harvested and dissolved in TRIzol® (Invitrogen Life
Technologies, Carlsbad, CA, USA) for total RNA extraction. The
isolated RNA was applied as a template to an RT reaction using an
RT-PCR kit (Promega Corporation, Madison, WI, USA). The qPCR
reaction was performed using an Mx3000P qPCR system (Agilent
Technologies, Waldbronn, Germany) for 3 min at 95°C and for 39
cycles of 10 sec at 95°C, 10 sec at 55°C and 30 sec at 72°C.
Gene-specific primers for human HIF-1α, MMP-2, MMP-14 and Bcl-2
were designed by Sangon Biotech Co., Ltd. (Shanghai, China), as
shown in Table I.
 | Table IGene-specific primer pairs used in
this study. |
Table I
Gene-specific primer pairs used in
this study.
| Genes | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| MMP-2 |
CGCATCTGGGGCTTTAAACAT |
CCATTAGCGCCTCCATCGTA |
| MMP-14 |
CCGATGTGGTGTTCCAGACA |
TCGTATGTGGCATACTCGCC |
| HIF-1α |
GCGCGAACGACAAGAAAAAGA |
GTGGCAACTGATGAGCAAGC |
| Bcl-2 |
CTTTGAGTTCGGTGGGGTCA |
GGGCCGTACAGTTCCACAAA |
| GAPDH |
AAGGTGAAGGTCGGAGTCAA |
AATGAAGGGGTCATTGATGG |
Western blot analysis
The protein extraction reagent and the extraction
method used for the extraction of total protein from the cells of
each group were as described previously (2). The proteins were separated by 8%
sodium dodecyl sulphate-polyacrylamide gel electrophoresis and
transferred onto polyvinylidene difluoride membranes for immunoblot
analysis. Subsequent to blocking with 5% skimmed milk powder for 2
h at room temperature, the membranes were incubated with monoclonal
rabbit anti-human HIF-1α (ab51608; 1:500; Abcam, Cambridge, UK),
mouse monoclonal anti-human MMP-2 (sc-13594; 1:500 Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA), polyclonal rabbit
anti-human MMP-14 (ab109745, 1:500; Abcam), polyclonal rabbit
anti-human Bcl-2 (sc-492-G; 1:500; Santa Cruz Biotechnology, Inc.)
and GAPDH (ab14247; 1:3,000; Abcam) antibodies at 4°C overnight.
The membranes were then incubated with peroxidase-conjugated
secondary antibodies (EMD Millipore, Billerica, MA, USA) for 1 h at
room temperature. The specific protein bands on the membranes were
visualised by the enhanced chemiluminescence method and analysed
using the Tanon 3500 (Tanon 3500R) Gel Imaging System (Jinglai
Laboratory Equipment Co., Ltd., Shanghai, China). The absorbance of
each band was measured, and the ratio of each target band to GAPDH
was used to show the expression of each target protein.
Statistical analysis
Data are shown as the mean ± standard deviation and
were analysed using SPSS version 11.5 software (SPSS, Inc.,
Chicago, IL, USA). Comparisons between two groups were performed
using the Student’s t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
mRNA expression of HIF-1α, MMP-2, MMP-14
and Bcl-2
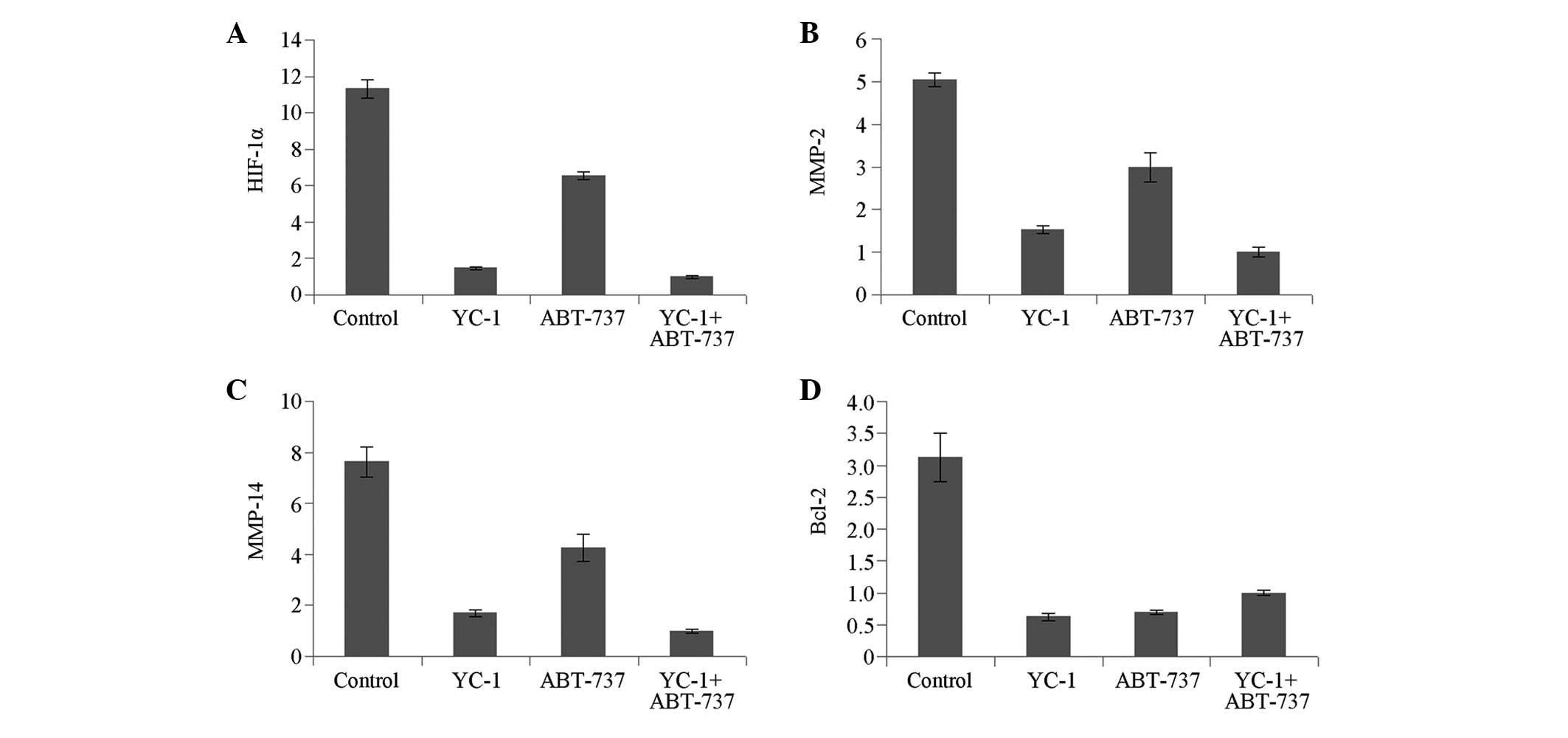
Compared with that in the control group, the mRNA
expression of HIF-1α was downregulated significantly in the YC-1
group (P<0.05); this downregulation was not significant in the
ABT-737 group. By contrast, the mRNA expression levels of MMP-2,
MMP-14 and Bcl-2 were all significantly reduced in the YC-1 and
ABT-737 groups (P<0.05) (Fig.
1).
Compared with that in the YC-1 + ABT-737 group, the
mRNA expression of HIF-1α was increased significantly in the
ABT-737 group (P<0.05) but increased slightly in the YC-1 group,
without significance (P>0.05). The mRNA expression of MMP-2 and
MMP-14 showed an increasing trend but that of Bcl-2 showed a
decreasing trend in the YC-1 and ABT-737 groups compared with that
in the YC-1 + ABT-737 group, although no significant differences
were observed (P>0.05) (Fig.
1). The mRNA expression levels of HIF-1α, MMP-2 and MMP-14 in
the YC-1 group were significantly different from those in the
ABT-737 group (P<0.01) (Fig.
1A–C); however, no statistically significant difference was
observed in the mRNA expression of Bcl-2 (Fig. 1D).
Protein expression of HIF-1α, MMP-2,
MMP-14 and Bcl-2
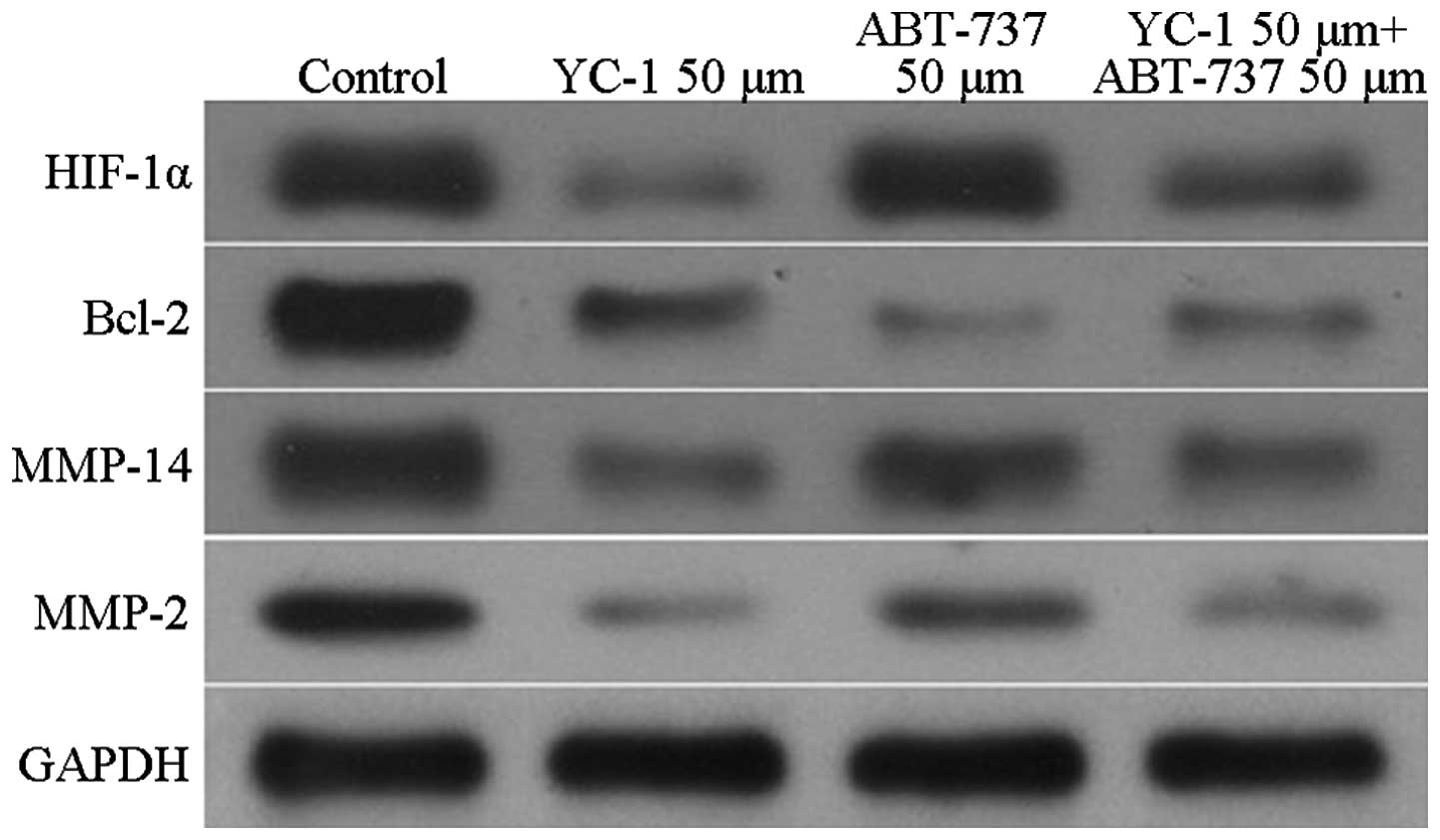
The expression levels of HIF-1α, MMP-2, MMP-14 and
Bcl-2 in the U87 cells underwent changes that were similar to those
observed for the mRNA expression. Compared with the HIF-1α protein
expression in the control group, the expression of the U87 cells in
the YC-1 group was significantly decreased (P<0.05). By
contrast, the expression of HIF-1α in the U87 cells in the ABT-737
group showed no significant reduction. In the YC-1 and ABT-737
groups, the expression levels of MMP-2, MMP-14 and Bcl-2 were all
significantly reduced compared with those in the control group
(P<0.05) (Fig. 2). Furthermore,
compared with HIF-1α protein expression in the YC-1 + ABT-737
group, the expression was increased significantly in the ABT-737
group (P<0.05) but not in the YC-1 group. The expression levels
of MMP-2, MMP-14 and Bcl-2 showed no significant differences
between the YC-1 + ABT-737 group and the YC-1 and ABT-737 groups
(P>0.05, Fig. 2).
Discussion
In 1999, Maniotis et al (1) found a special microcirculatory system
in the highly aggressive uveal melanoma, in which the channels were
covered not with endothelial cells but with a layer of tumour
cells. Blood flow was noted within the lumen. Occasionally, a layer
of periodic acid-Schiff-positive stromal membrane separating the
tumour cells from blood cells was observed; this mechanism was
eventually termed VM by Maniotis et al (1). VM was then gradually found in ovarian
(16), breast (17), liver (18) and stomach (19) cancer. Further experiments
demonstrated that VM only occurs in extremely malignant tumours
(20) and that hypoxia can promote
its formation in these tumours (15,20,21).
VM has also been found in malignant glioblastomas, and the induced
hypoxic condition has been found to affect the VM formation of
gliomas (22). Furthermore,
hypoxia can promote the VM generation of low-grade malignant glioma
cells. The HIF-1α and VEGF-EphA2-MMP-VM channels are reportedly key
channels for VM formation under hypoxic conditions. VM has also
been found among the three-dimensional culture of U-251 human glial
cells. In recent studies, hypoxia was confirmed to promote the VM
formation of glioma cells, and the induced HIF-1α and VEGF-A
expression was suggested a mechanism involved in this process
(10,23,24).
YC-1 has superior anti-angiogenic and tumour
therapeutic activities. Wohlfart et al (21) found that YC-1 could promote nitric
oxide (NO) synthesis in foetal bovine aortic endothelial cells in a
concentration-dependent manner; however, NO inhibits the stability
and transcriptional activity of HIF-1α under hypoxic conditions. NO
has also been shown to have inhibitory effects on VEGF and GPI
genes in human pancreatic cancer cells under hypoxic conditions
(8). Gariboldi et al
(9) reported that YC-1 exerts
inhibitory effects on the VM formation of U87 cells. Few reports,
however, have focused on the VM formation mechanisms of human
glioma U87 cells under the hypoxic condition (25,26).
The mechanisms underlying VM formation remain
unclear. Bcl-2 is currently considered to be an important factor
for the generation, metastasis and angiogenesis of malignant
tumours (5,9). In conditions of induced hypoxia,
Bcl-2 can affect the VM formation of human melanoma by regulating
the expression of VE-cadherin (27). An investigation into the possible
effect of Bcl-2 on VM formation inside hypoxic U87 cells is likely
to aid the understanding of VM formation mechanisms inside hypoxic
U87 cells.
In CoCl2-induced hypoxia, cobalt can
replace the chelated iron in haemoglobin and damage the oxygen
sensation of cells. CoCl2 has stimulated a hypoxic
environment in cultured cells that is strongly comparable to
hypoxia in vivo. Since glioblastoma cells are in an induced
hypoxic microenvironment, tumour cells fuse to form VM and then
provide nutrition for the tumour. Thus, during the inhibition of
tumour blood vessels, understanding the channels for VM and
applying comprehensive treatment programs can cut off the tumour
blood supply satisfactorily and consequently cure tumours (28–30).
In the present study, U87 cells were treated with
the specific inhibitors of Bcl-2 (ABT-737) and HIF-1α (YC-1). The
cells were then cultured under an induced hypoxic condition.
Subsequent to the cells being cultured with CoCl2 for 24
h, the expression of HIF-1α, MMP-2, MMP-14 and Bcl-2 in the U87
cells was determined. In the current study, it was observed that
the expression of HIF-1α in the U87 cells in the YC-1 group was
significantly reduced (P<0.05) but the ABT-737 group exhibited
no significant reduction. In the YC-1 + ABT-737 group, the
expression levels of MMP-2, MMP-14 and Bcl-2 were all significantly
reduced compared with those in the control group (P<0.05). No
significant differences were observed in the expression of Bcl-2
among the YC-1 + ABT-737, ABT-737 and YC-1 groups. This finding
indicates that Bcl-2 does not play a role in VM via the HIF-1α
pathway. HIF-1α-MMP-2-MMP-14-VM has previously been confirmed as a
traditional pathway (2,5). Through a different pathway, the Bcl-2
inhibitor influenced the MMP-2 and MMP-14 expression levels, but
not HIF-1α expression levels, in hypoxic U87 cells significantly.
This finding indicates that Bcl-2 inhibitors were not involved in
VM formation via the traditional pathway. In conclusion, Bcl-2 may
be an important factor in the VM formation of human malignant
glioma U87 cells under hypoxic conditions. Bcl-2 may exert its
effects both through the HIF-1α-MMP-2-MMP-14-VM channel and through
other means that are independent of this channel. This finding
indicates that hypoxia affects VM formation not simply by
influencing the expression of HIF-1α. Further studies on the
mechanisms underlying the VM formation of glioma cells under
hypoxic conditions should therefore be carried out.
References
|
1
|
Maniotis AJ, Folberg R, Hess A, et al:
Vascular channel formation by human melanoma cells in vivo and in
vitro: vasculogenic mimicy. Am J Pathol. 155:739–752. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Paulis YW, Soetekouw PM, Verheul HM,
Tjan-Heijnen VC and Griffioen AW: Signalling pathways in
vasculogenic mimicry. Biochim Biophys Acta. 1806:18–28.
2010.PubMed/NCBI
|
|
3
|
Kirschmann DA, Seftor EA, et al: Molecular
pathways: vasculogenic mimicry in tumor cells: diagnostic and
therapeutic implications. Clin Cancer Res. 18:2726–2732. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mohla S: Tumor microenvironment. J Cell
Biochem. 101:801–804. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Florczyk SJ, Wang K, Jana S, et al: Porous
chitosan-hyaluronic acid scaffolds as a mimic of glioblastoma
microenvironment ECM. Biomaterials. 34:10143–10150. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tafani M, Di Vito M, et al:
Pro-inflammatory gene expression in solid glioblastoma
microenvironment and in hypoxic stem cells from human glioblastoma.
J Neuroinflammation. 8:32–40. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Heddleston JM, Wu Q, Rivera M, et al:
Hypoxia-induced mixed-lineage leukemia 1 regulates glioma stem cell
tumorigenic potential. Cell Death Differ. 19:428–439. 2012.
View Article : Google Scholar
|
|
8
|
Scully S, Francescone R, Faibish M, et al:
Transdifferentiation of glioblastoma stem-like cells into mural
cells drives vasculogenic mimicry in glioblastomas. J Neurosci.
32:12950–12960. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gariboldi MB, Ravizza R and Monti E: The
IGFR1 inhibitor NVP-AEW541 disrupts a pro-survival and
pro-angiogenic IGF-STAT3-HIF1 pathway in human glioblastoma cells.
Biochem Pharmacol. 80:455–462. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Neelam S, Brooks MM and Cammarata PR:
Lenticular cytoprotection. Part 1: the role of hypoxia inducible
factors-1α and -2α and vascular endothelial growth factor in lens
epithelial cell survival in hypoxia. Mol Vis. 19:1–15. 2013.
|
|
11
|
Kucharzewska P, Christianson HC, Welch JE,
et al: Exosomes reflect the hypoxic status of glioma cells and
mediate hypoxia-dependent activation of vascular cells during tumor
development. Proc Natl Acad Sci USA. 110:7312–7317. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nissou MF, El Atifi M, Guttin A, et al:
Hypoxia-induced expression of VE-cadherin and filamin B in glioma
cell cultures and pseudopalisade structures. J Neurooncol.
113:239–249. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Biroccio A, Candiloro A, Mottolese M, et
al: Bcl-2 overpression and hypoxia synergistically act to modulate
vascular endothelial growth factor expression and in vivo
angiogenesis in a breast carcinoma line. FASEB J. 14:652–660.
2000.PubMed/NCBI
|
|
14
|
Reed JC: Bcl-2-family proteins and
hematologic malignancies: history and future prospects. Blood.
111:3322–3330. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yip KW and Reed JC: Bcl-2 family proteins
and cancer. Oncogene. 27:6398–6406. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sood AK, Fletcher MS, Zahn CM, et al: The
clinical significance of tumor cell-lined vasculature in ovarian
carcinoma-: iImplications for anti-vasculogenic therapy. Cancer
Biol Ther. 1:661–664. 2002. View
Article : Google Scholar
|
|
17
|
Shirakawa K, Kobayashj H, et al:
Inflammatory breast cancer: vasculogenic mimicry and its
hemodynamics of an inflammatory breast cancer xenograft model.
Breast Cancer Res. 5:136–139. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guzman G, Cotler SJ, Lin AY, et al: A
pilot study of vasculogenic mimicry immunohistochemical expression
in hepatocellular carcinoma. Arch Pathol Lab Med. 131:1776–1781.
2007.PubMed/NCBI
|
|
19
|
Fujimoto A, Onodera H, Mori A, et al:
Tumor plasticity and extravascular circulation in EVC304 human
bladder carcinoma cells. Anticancer Res. 26:59–69. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Guzman G, Cotler SJ, et al: A pilot study
of vasculogenic mimicry immunohistochemical expression in
hepatocellular carcinoma. Arch Pathol Lab Med. 131:1776–1781.
2007.PubMed/NCBI
|
|
21
|
Wohlfart P, Malinski T, Ruetten H, et al:
Release of nitric oxide from endothelial cells stimulated by YC-1,
an activator of soluble guanylyl cyclase. Br J Pharmacol.
128:1316–1322. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
El Hallani S, Boisselier B, Peglion F, et
al: A new alternative mechanism in glioblastoma vascularization:
tubular vasculogenic mimicry. Brain. 133:973–982. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vaupel P and Mayer A: Hypoxia in cancer:
significance and impact on clinical outcome. Cancer Metastasis Rev.
26:225–239. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ahluwalia A and Tarnawski AS: Critical
role of hypoxia sensor - HIF-1α in VEGF gene activation.
Implications for angiogenesis and tissue injury healing. Curr Med
Chem. 19:90–97. 2012. View Article : Google Scholar
|
|
25
|
EI Hallani S, Boisselier B, Peglion F, et
al: A new alternative mechanism in glioblastoma vascularization:
tubular vasculogenic mimicry. Brain. 133:973–982. 2010. View Article : Google Scholar
|
|
26
|
Youle RJ and Strasser A: The BCL-2 protein
family: opposing activities that mediate cell death. Nat Rev Mol
Cell Biol. 9:47–59. 2008. View
Article : Google Scholar
|
|
27
|
Zhao N, Sun BC, Sun T, et al:
Hypoxia-induced vasculogenic mimicry formation via VE-cadherin
regulation by Bcl-2. Med Oncol. 29:3599–3607. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jain RK, di Tomaso E, Duda DG, et al:
Angiogenesis in brain tumours. Nat Rev Neurosci. 8:610–622. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sullivan R, Paré GC, Frederiksen LJ,
Semenza GL and Graham CH: Hypoxia-induced resistance to anticancer
drug is associated with decreased senescence and requires
hypoxia-inducible factor-1 activity. Mol Cancer Ther. 7:1961–1973.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mak P, Leav I, Pursell B, et al: ERbeta
impedes prostate cancer EMT by destabilizing HIF-1alpha and
inhibiting VEGF mediated snail nuclear localization: implications
for Gleason grading. Cancer Cell. 17:319–332. 2010. View Article : Google Scholar : PubMed/NCBI
|