Introduction
Depending on its acute or chronic use and the dose,
alcohol has toxic effects on the mother and fetus when consumed
during pregnancy (1). Its toxic
effects have been a topic of investigation for approximately five
decades (2,3). At present, fetal alcohol spectrum
disorders (FASD) have been shown to include alcohol-related birth
defects (ARBD), alcohol-related neurological disorders (ARND) and
fetal alcohol syndrome (FAS) (3–5). FAS
causes mental retardation and growth deficits together with facial
anomalies in newborns from chronic alcohol-consuming mothers
(3).
Children born with FAS have certain difficulties
including learning, memory, attention span, communication, hearing
and vision (6,7). Extreme attention, rehabilitation,
education, medical support and major economic resources are
required to integrate these children into society (8,9). In
order to overcome this social problem, new strategies are required
to prevent the teratogenic effects of alcohol on the fetus.
Oxidative stress has been shown to be associated
with numerous diseases, including cardiovascular disease, cancer
and neurological and endocrinological disorders, and also FAS
(10,11). Oxidative stress is the imbalance
between the production and breakdown of reactive oxygen species
(ROS) by endogenous antioxidants (12). Oxidative stress results in
increasing lipid peroxidation (13,14)
and also ROS production. As a result, membrane and enzyme function
may be damaged via overproduction of ROS (11). Antioxidants such as superoxide
dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPx)
detoxify ROS.
Boric acid (BA) is an essential trace element for
plants, humans and animals to support metabolic events. BA is
monobasic but is not a proton donor. Instead, it acts as a Lewis
acid, accepting a hydroxyl ion from water and releasing a proton.
It has high affinity for S-adenosyl methionine (SAM) and
oxidized nicotinamide adenine dinucleotide (NAD+). BA
forms complexes with hydroxyl group-containing glycolipids,
glycoproteins and phosphoinositides. By this, it affects membrane
integrity, calcium chelator function and redox metabolism. When
boron is taken orally, it enters the blood stream quickly and
thoroughly. Its urinary excretion rate is ~100% (15,16).
Boron can be used for neutron capture therapy in certain types of
cancer (17). It decreases the
severity and incidence of inflammatory diseases (18). BA affects calcium, magnesium,
potassium, vitamin D, aldehyde dehydrogenase, xanthine oxidase,
cytochrome b reductase, insulin, estrogen, testosterone, T3, T4,
triglycerides and glucose metabolism (19). Apart from its diverse functions,
previous studies have shown that BA also has antioxidant activity
(19–21).
In the current study, the possible protective effect
of BA supplementation was evaluated in prenatally alcohol-exposed
rats at the cerebral cortex.
Materials and methods
Experimental procedure
Ten adult male and 30 adult female Sprague-Dawley
rats weighing between 200 and 250 g were created for breeding in
the present study. For mating, a male rat was randomly picked up
and placed into a female’s cage. They were placed in a secluded,
temperature- and humidity-controlled room (22±3°C and 60±5%
respectively) in which a 12:12 h light-dark cycle was maintained.
The females were checked each morning for the presence of a vaginal
plug. The presence of a vaginal plug was considered as evidence of
fertilization, and the day the plug was observed was regarded as
embryonic day (E) 0, the first embryonic day. Only in 15 females
was a vaginal plug observed, and these were used to set up three
experimental groups.
Five pregnant rats were selected for each of the
control, alcohol and alcohol + boric acid groups. The animal model
of prenatal alcohol consumption was modified from that of Uzbay and
Kayaalp (22). The control group
was pair-fed with an isocaloric modified liquid diet containing
sucrose as a caloric substitute for alcohol (96 g sucrose and 75 ml
cows’ milk replaced 60.75 g or 75 ml ethanol). Modified liquid diet
(MLD) was given to the alcohol and alcohol + boric acid groups. The
MLD comprised 925 ml low fat cows’ milk (Sütaş, Istanbul, Turkey),
from 25 to 75 ml alcohol (96.5% v/v ethanol; Merck Millipore,
Darmstadt, Germany) and 17 g sucrose (Merck Millipore). The
calorific content of the diet was 1,000.7 kcal/l. The weight of the
rats was recorded every day, and the daily alcohol intake was also
measured. The MLD was prepared daily and given to the animals while
fresh. Extra chow or water were not available during the
experimental period.
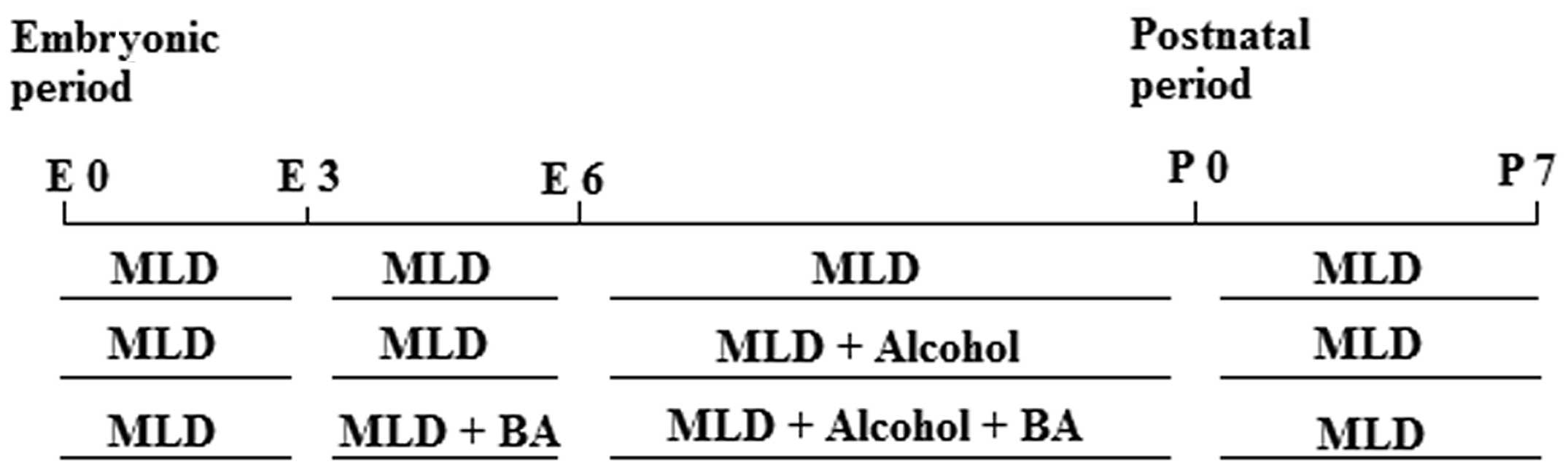
At the beginning of the study, the rats were given
MLD without alcohol for 6 days (E0–E5). BA was administered prior
to alcohol administration (E3–E5) in order to accustom the rats to
drinking the supplements. The dosage of BA (Merck Millipore) was
100 mg/kg (21). Then, a liquid
diet with 2.4% alcohol was administered for 3 days (E6–E8). The
alcohol concentration was increased to 4.8% during the following 3
days (E9–E11) and finally reached 7.2% over the next 11 days
(E11–E22; Fig. 1).
Following delivery, the rats were given MLD without
alcohol on postnatal days (P) 0–7. A pool was prepared by selecting
two pups from each group of five rats, frozen tissue from this
group of pups was used for biochemical analysis. A total of 30 pups
(n=10 per group) at P7 were used for the study. Pups were
sacrificed by decapitation on day P7. The cerebral cortex was
surgically removed. Tissues used for biochemical studies were
frozen in liquid nitrogen and kept at −80°C until they were tested.
All experiments were carried out in accordance with institutional
guidelines for animal welfare (Eskişehir Osmangazi University
Animal Care and Use Committee, Eskişehir, Turkey) and were approved
by the Ethics Committee of the Medical and Surgical Experimental
Research Center of Osmangazi University.
Blood alcohol concentration (BAC)
measurement
On days E15, E18 and E20, every morning 6 and 22 h
after the administration of fresh diets, a 20-μl blood sample was
taken from the tails of the rats. Plasma was immediately separated
following centrifugation and quantified at 340 nm to determine the
BAC (NAD-ADH Reagent Multiple Test Vial, Sigma-Aldrich, St. Louis,
MO, USA) using a Shimadzu UV-1201 spectrophotometer (Shimadzu
Corporation, Kyoto, Japan). For each group, on days E15, E18 and
E20, a total of six values were obtained as a result of two
applications per day (23). The
highest of these values was defined as the peak BAC for each
group.
Biochemical measurement
Lipid peroxidation was quantified at 532 nm by the
measurement of malondialdehyde (MDA) reacted with thiobarbituric
acid (TBA) according to the method of Ohkawa et al (24). The results were expressed in
nmol/mg protein. SOD activity was determined according to the
method of Winterbourn et al (25). One unit of SOD expressed in U/mg
protein was designated as the amount of enzyme that inhibits the
reduction of nitroblue tetrazolium reduction by 50%. CAT activities
were calculated using the method of Beutler (26). The reduction in optical density per
minute was determined and the enzyme activity was expressed in U/mg
protein. GPx activity in U/g protein was spectrophotometrically
determined at 340 nm using the methods of Paglia and Valentine
(27). The protein concentration
of homogenates gathered from brain tissues were determined using
the Bradford assay (28).
Statistical analysis
SPSS software, version 15.0 for Windows (SPSS, Inc.,
Chicago, IL, USA) was used for the statistical analysis of
biochemical data. In order to assess differences between groups,
one-way analysis of variance (ANOVA) and Tukey’s multiple
comparison test were used. Results are presented as mean ± standard
deviation and P<0.05 was considered to indicate a statistically
significant result.
Results
MDA levels
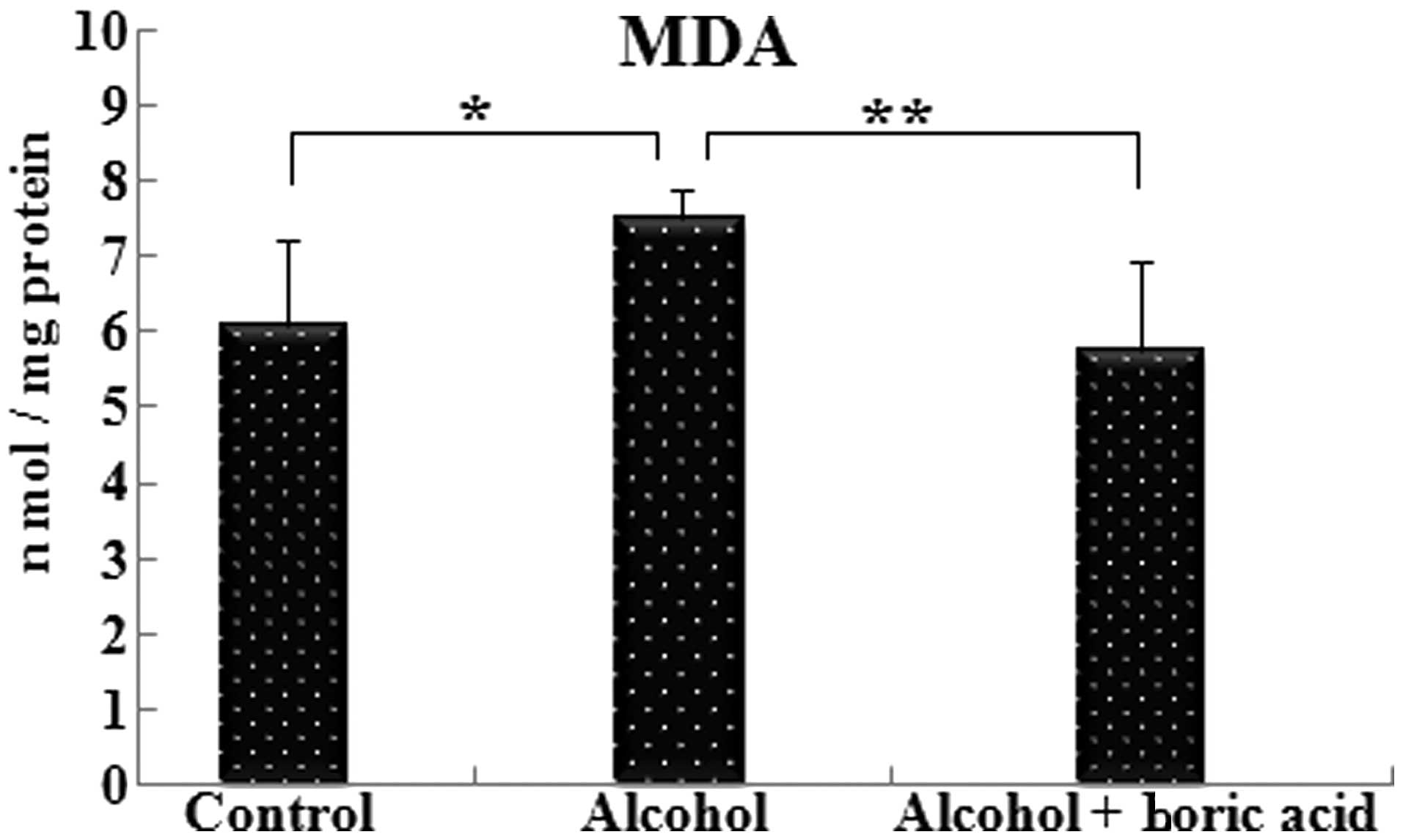
MDA levels in the alcohol group (7.53±1.41 nmol/mg
protein) were significantly increased (P<0.05) compared with the
control level (6.11±1.11 nmol/mg protein). The MDA level in the
alcohol + boric acid group (5.74±1.22) exhibited a significant
reduction (P<0.01) compared with that in the alcohol group
(Fig. 2).
Enzyme activities
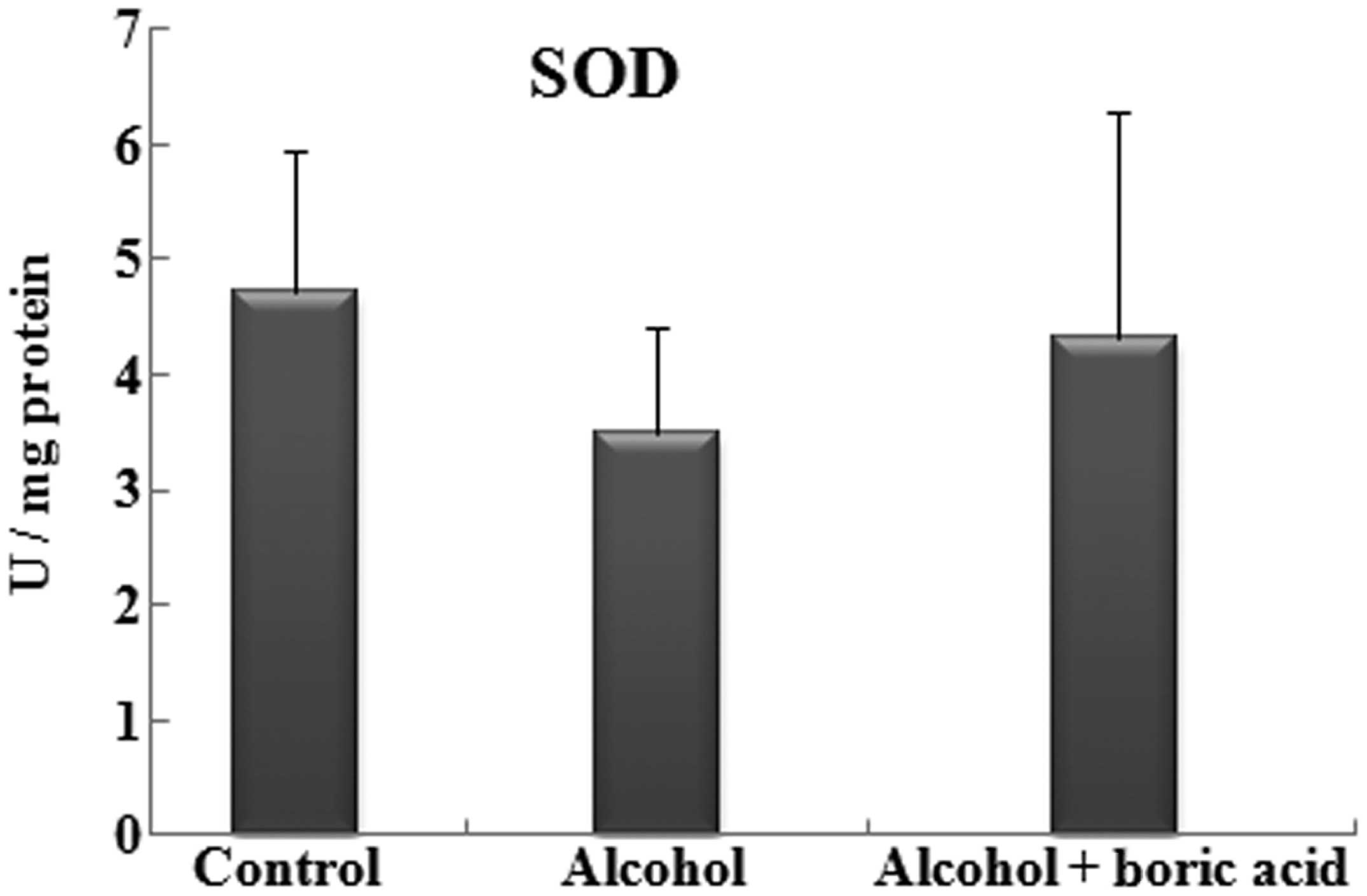
No statistically significant differences in SOD
activities were identified among the groups. The SOD values in the
control, alcohol and alcohol + boric acid groups were 4.73±1.22,
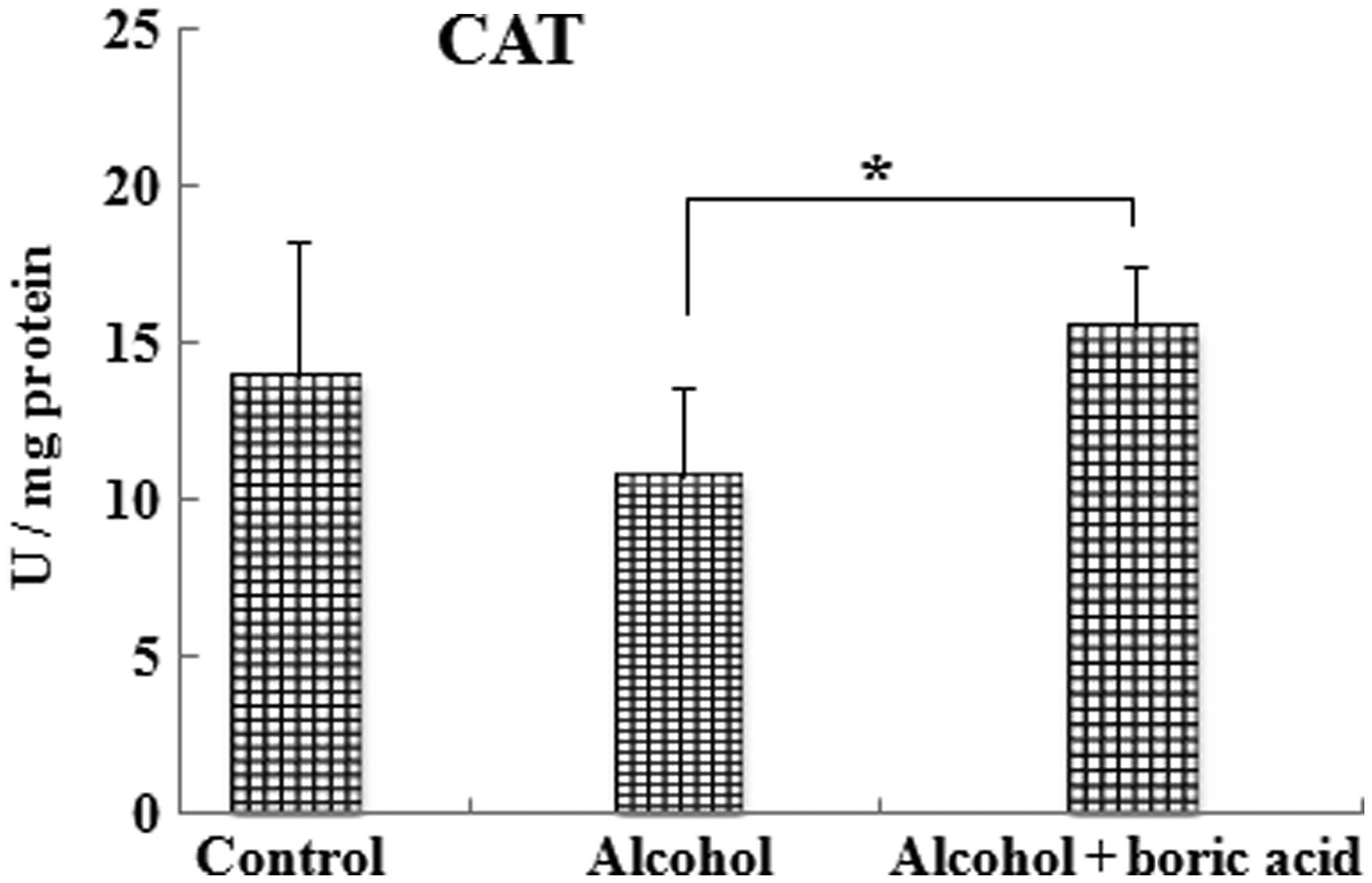
3.50±0.92 and 4.31±1.96 U/mg protein, respectively (Fig. 3). The CAT activity of the alcohol +
boric acid group (15.55±3.65 U/mg protein) was found to be higher
(P<0.05) compared with that of the alcohol group (10.80±2.79
U/mg protein). There was no statistically significant difference
(P>0.05) in CAT activity between the control group (13.90±4.34
U/mg protein) and the alcohol group (10.80±2.79 U/mg protein;
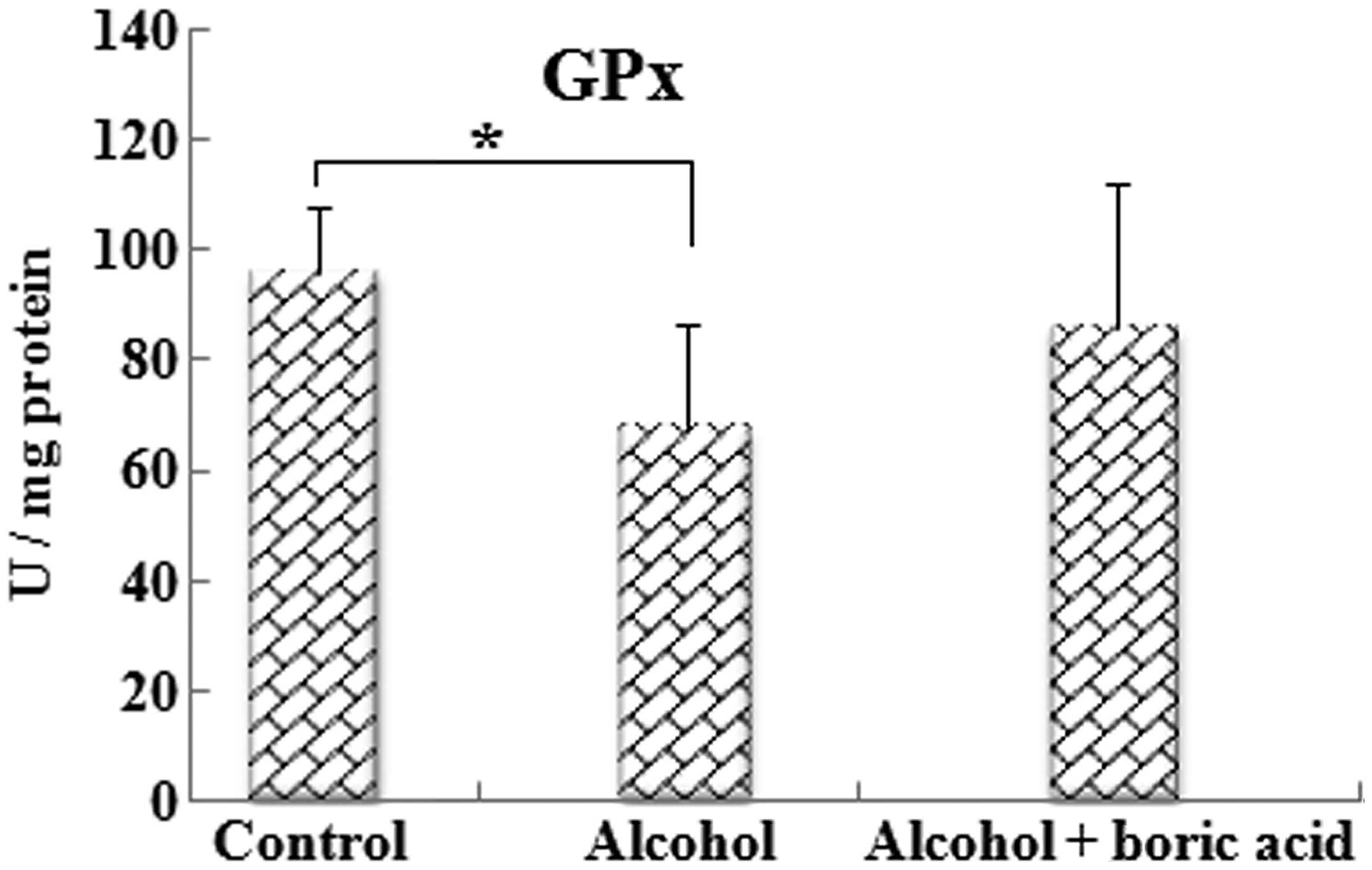
Fig. 4). GPx activity in the
alcohol group (67.49±18.88 U/mg protein) was decreased (P<0.05)
as compared with that in the control group (95.70±25.85 U/mg
protein). The GPx activity of the alcohol + boric acid group
(85.67±29.71 U/mg protein) was not significantly different
(P>0.05) from that on the other two groups (Fig. 5).
Blood alcohol levels
The highest blood alcohol levels recorded in the two
alcohol-administered groups were between 175 and 182 mg/dl.
Discussion
To the best of our knowledge, this is the first
study concerning the effect of BA administration on rats with FAS
and the possible antioxidant mechanisms. Alcohol may have direct
and indirect effects on the brain. The direct effect of alcohol
results in diffusion from the lipid barrier of the cell membrane
and increased flowability in the membrane. Oxidative and
non-oxidative metabolisms are associated with the indirect effects
of alcohol (29). Acetaldehyde is
generated, and as a result of forming protein adducts, acetaldehyde
disrupts mitochondria and restorative enzymes, leading to the
formation of reactive oxygen species. Furthermore, acetaldehyde
causes a reduction in the number of glutathione groups and
hydrophobic regions at the cell membrane and mitochondrial
membranes (30). Fatty acid ethyl
esters are produced as a result of non-oxidative metabolism,
preventing the esterification of cholesterol on neuronal membranes
and also disrupting mitochondrial and myelin metabolism (31). It is well-documented that oxidative
stress-induced by alcohol-associated mechanisms gives rise to cell
damage.
The constitutive formation of oxidants can be
balanced by the production of antioxidants at a similar rate. The
imbalance between oxidants and antioxidant species causes oxidative
stress, resulting from the peroxidation of lipids. MDA is one of
the end products in the lipid peroxidation process (32,33).
In the present study, prenatal alcohol intake increased the level
of lipid peroxidation. These findings are in line with previous
fetotoxicity studies (34,35). Enzymatic (SOD, CAT and GPx) and
non-enzymatic (thiol and GSH) antioxidants have important roles in
preventing the damage resulting from alcohol-induced ROS (11). When female rats were exposed to
alcohol during gestation, GPx activity decreased while SOD and CAT
activities were not significantly affected (36). In a study concerning free radical
formation in rat brains exposed to acute doses of ethanol, it was
demonstrated that in the early postnatal period while ROS levels
increased, GPx activity decreased (37). Accordingly, chronic alcohol
exposure has been shown to decrease cytosolic and mitochondrial GPx
activity by 40 and 30%, respectively, in rat liver (38). The steady activity levels of
enzymatic antioxidants, namely SOD and CAT, may be due to levels of
those enzymes being higher in an adult rat than in a fetus.
Furthermore, the activities of these enzymes have been shown to be
lower in the brain than other tissues, for example, in the liver
and kidney (11).
At present, the complete antioxidant mechanism of BA
is not fully understood (39);
however, BA is a well-known component of cell membrane functions
and enzymatic reactions (40,41).
In a previous study, boron supplementation (100 mg/kg/day) was
shown to decrease lipid peroxidation by enhancing antioxidant
activity (21). In another study,
a 100-mg/kg/day dose of BA was also used and shown not to be toxic
for rodents when administered for a short period (≤4 weeks)
(42). According to the proposed
hypothesis, a possible mechanism for the decline of alcohol-induced
lipid peroxidation might be the increasing levels of SAM with BA
administration. Being a Lewis acid, BA may form complexes with many
biological compounds through its hydroxyl groups. It has high
affinity for SAM, which is known as a methyl donor in biochemical
processes, a metabolite in the transsulfuration pathway and a
precursor in the synthesis of polyamines (43). In a previous study, chronic alcohol
consumption decreased SAM levels while increasing
S-adenosylhomocysteine (SAH) levels and decreasing the
methylation capacity indicator ratio, SAM/SAH (44). In another study on boron
deprivation in the liver, the level of SAM was reported to decrease
while the homocysteine level increased (41). As Nielsen (40) indicated, boron balances
transmembrane signaling and ion movements with its structural
and/or functional properties on the cell membrane. According to
this information, in the current study, it is hypothesized that a
decline in lipid peroxidation was associated with an increase in
the SAM/SAH ratio, resulting in balancing of the cell membrane
functions.
In the present study, the blood alcohol level
reached 175–182 mg/dl in the alcohol group. This level might lead
to impairment in prenatal development since it has been reported
that a BAC >100 mg/dl in pregnant rats can impair embryo-fetal
development (45). The alcohol +
boric acid group had no significant change in SOD and GPx
activities compared with the alcohol group. SOD and GPx inactivate
superoxide and hydrogen peroxide, respectively. In an earlier study
of BA supplementation in rats, SOD activity was not affected in
erythrocytes or in heart and liver tissues (21). Paralleling the results of the
present study, SOD activity was found to be unchanged in another
study conducted to assess the effect of BA on the redox status of
the rat liver (46). However, in
those studies, BA increased GPx activity. Turkez et al
demonstrated that when boron compounds were administered at low
doses (15 mg/l), the CAT activity of erythrocytes increased,
whereas high doses (500 mg/l) of boron led to decreased CAT
activities (20). In a study of
carbon tetrachloride-induced toxicity in rats, it was shown that BA
administration at various doses (50, 100 and 200 mg/kg) increased
the CAT enzyme activity in the liver (19). Placental transfer from the mother
to fetus and the presence of CAT in the fetus are responsible for
the presence of aldehydes in fetal rat brains (47). In the present study, the increasing
CAT activity following BA administration may stem from the ability
of BA (a Lewis acid) to accept a hydroxyl ion. BA potentially has
two crucial functions in this mechanism. Firstly, it detoxifies
hydrogen peroxide derived from alcohol by converting it into
peroxyboric acid. Secondly, BA converts the CAT enzyme to its
native state and maintains the enzyme.
In summary, alcohol-related oxidative stress of cell
membranes can be decreased by BA supplementation. According to the
results of this study, BA has no impact on blood alcohol levels,
but may be a great candidate for use in the protection and
regulation of brain membranes. However, future studies are required
to evaluate this recovery effect of BA on FAS.
References
|
1
|
Abel EL and Hannigan JH: Maternal risk
factors in fetal alcohol syndrome: provocative and permissive
influences. Neurotoxicol Teratol. 17:445–462. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lemoine P, Harousseau H, Borteyru JP and
Menuet JC: Children of alcoholic parents - observed anomalies:
discussion of 127 cases. Ther Drug Monit. 25:132–136. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jones KL and Smith DW: Recognition of the
fetal alcohol syndrome in early infancy. Lancet. 302:999–1001.
1973. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
May PA, Gossage JP, Kalberg WO, et al:
Prevalence and epidemiologic characteristics of FASD from various
research methods with an emphasis on recent in-school studies. Dev
Disabil Res Rev. 15:176–192. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Burd L and Martsolf JT: Fetal alcohol
syndrome: diagnosis and syndromal variability. Physiol Behav.
46:39–43. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ahveninen J, Escera C, Polo MD, Grau C and
Jääskeläinen IP: Acute and chronic effects of alcohol on
preattentive auditory processing as reflected by mismatch
negativity. Audiol Neurootol. 5:303–311. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sampson PD, Streissguth AP, Bookstein FL
and Barr HM: On categorizations in analyses of alcohol
teratogenesis. Environ Health Perspect. 108(Suppl 3): 421–428.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Abel EL and Sokol RJ: A revised
conservative estimate of the incidence of FAS and its economic
impact. Alcohol Clin Exp Res. 15:514–524. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fast DK, Conry J and Loock CA: Identifying
fetal alcohol syndrome among youth in the criminal justice system.
J Dev Behav Pediatr. 20:370–372. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cornelli U: Antioxidant use in
nutraceuticals. Clin Dermatol. 27:175–194. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brocardo PS, Gil-Mohapel J and Christie
BR: The role of oxidative stress in fetal alcohol spectrum
disorders. Brain Res Rev. 67:209–225. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sies H: Oxidative stress: from basic
research to clinical application. Am J Med. 91(Suppl 3): 31S–38S.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cohen-Kerem R and Koren G: Antioxidants
and fetal protection against ethanol teratogenicity. I Review of
the experimental data and implications to humans. Neurotoxicol
Teratol. 25:1–9. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bandyopadhyay U, Das D and Banerjee RK:
Reactive oxygen species: Oxidative damage and pathogenesis. Current
Sci. 77:658–666. 1999.
|
|
15
|
Nielsen FH: Is boron nutritionally
relevant? Nutr Rev. 66:183–191. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hunt CD: Boron. Encyclopedia of Dietary
Supplements. 1st edition. Coates PM, Blackman MR, Cragg GM, Levine
M, Moss J and White JD: Marcel Dekker; New York, NY: pp. 55–65.
2005
|
|
17
|
Altieri S, Bortolussi S, Bruschi P, et al:
Neutron autoradiography imaging of selective boron uptake in human
metastatic tumours. Appl Radiat Isot. 66:1850–1855. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hunt CD and Idso JP: Dietary boron as a
physiological regulator of the normal inflammatory response: A
review and current research progress. J Trace Elem Exp Med.
12:221–233. 1999. View Article : Google Scholar
|
|
19
|
Ince S, Keles H, Erdogan M, Hazman O and
Kucukkurt I: Protective effect of boric acid against carbon
tetrachloride-induced hepatotoxicity in mice. Drug Chem Toxicol.
35:285–292. 2012. View Article : Google Scholar
|
|
20
|
Turkez H, Geyikoğlu F, Tatar A, Keleş S
and Ozkan A: Effects of some boron compounds on peripheral human
blood. Z Naturforsch C. 62:889–896. 2007. View Article : Google Scholar
|
|
21
|
Ince S, Kucukkurt I, Cigerci IH, Fatih
Fidan A and Eryavuz A: The effects of dietary boric acid and borax
supplementation on lipid peroxidation, antioxidant activity, and
DNA damage in rats. J Trace Elem Med Biol. 24:161–164. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Uzbay IT and Kayaalp SO: A modified liquid
diet of chronic ethanol administration: validation by ethanol
withdrawal syndrome in rats. Pharmacol Res. 31:37–42. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiang Q, Hu Y, Wu P, et al: Prenatal
alcohol exposure and the neuroapoptosis with long-term effect in
visual cortex of mice. Alcohol Alcohol. 42:285–290. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ohkawa H, Ohishi N and Yagi K: Assay for
lipid peroxides in animal tissues by thiobarbituric acid reaction.
Anal Biochem. 95:351–358. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Winterbourn CC, Hawkins RE, Brian M and
Carrell RW: The estimation of red cell superoxide dismutase
activity. J Lab Clin Med. 85:337–341. 1975.PubMed/NCBI
|
|
26
|
Beutler E: Catalase. Red Cell Metabolism A
Manual of Biochemical Methods. 3rd edition. Grune and Stratton; New
York, NY: pp. 105–106. 1982
|
|
27
|
Paglia DE and Valentine WN: Studies on the
quantitative and qualitative characterization of erythrocyte
glutathione peroxidase. J Lab Clin Med. 70:158–169. 1967.PubMed/NCBI
|
|
28
|
Bradford MM: A rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal Biochem. 72:248–254.
1976. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sogut I and Kanbak G: In vitro effects of
ethanol with aspirin on rat brain synaptosomes: the potential
protective role of betaine. Int J Neurosci. 120:774–783. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zimatkin SM and Deitrich RA: Ethanol
metabolism in the brain. Addict Biol. 2:387–400. 1997. View Article : Google Scholar
|
|
31
|
Bora PS and Lange LG: Molecular mechanism
of ethanol metabolism by human brain to fatty acid ethyl esthers.
Alcohol Clin Exp Res. 17:28–30. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu D and Cederbaum AI: Alcohol, oxidative
stress, and free radical damage. Alcohol Res Health. 27:277–284.
2003.
|
|
33
|
Hagihara M, Nishigaki I, Maseki M and Yagi
K: Age-dependent changes in lipid peroxide levels in the
lipoprotein fractions of human serum. J Gerontol. 39:269–272. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Amini SA, Dunstan RH, Dunkley PR and
Murdoch RN: Oxidative stress and the fetotoxicity of alcohol
consumption during pregnancy. Free Radic Biol Med. 21:357–365.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Smith AM, Zeve DR, Grisel JJ and Chen WJ:
Neonatal alcohol exposure increases malondialdehyde (MDA) and
glutathione (GSH) levels in the developing cerebellum. Brain Res
Dev Brain Res. 160:231–238. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Brocardo PS, Boehme F, Patten A, Cox A,
Gil-Mohapel J and Christie BR: Anxiety- and depression-like
behaviors are accompanied by an increase in oxidative stress in a
rat model of fetal alcohol spectrum disorders: Protective effects
of voluntary physical exercise. Neuropharmacology. 62:1607–1618.
2012. View Article : Google Scholar
|
|
37
|
Shivakumar BR, Anandatheerthavarada HK and
Ravindranath V: Free radical scavenging systems in developing rat
brain. Int J Dev Neurosci. 9:181–185. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bailey SM, Patel VB, Young TA, Asayama K
and Cunningham CC: Chronic ethanol consumption alters the
glutathione/glutathione peroxidase-1 system and protein oxidation
status in rat liver. Alcohol Clin Exp Res. 25:726–733. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pawa S and Ali S: Boron ameliorates
fulminant hepatic failure by counteracting the changes associated
with the oxidative stress. Chem Biol Interact. 160:89–98. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nielsen FH: Nutritional requirements for
boron, silicon, vanadium, nickel, and arsenic: current knowledge
and speculation. FASEB J. 5:2661–2667. 1991.PubMed/NCBI
|
|
41
|
Hunt CD: The biochemical effects of
physiologic amounts of dietary boron in animal nutrition models.
Environ Health Perspect. 102(Suppl 7): 35–43. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Weir RJ Jr and Fisher RS: Toxicologic
studies on borax and boric acid. Toxicol Appl Pharmacol.
23:351–364. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Nielsen FH: Boron deprivation decreases
liver S-adenosylmethionine and spermidine and increases plasma
homocysteine and cysteine in rats. J Trace Elem Med Biol.
23:204–213. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Stickel F, Choi SW, Kim YI, et al: Effect
of chronic alcohol consumption on total plasma homocysteine level
in rats. Alcohol Clin Exp Res. 24:259–264. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Driscoll CD, Streissguth AP and Riley EP:
Prenatal alcohol exposure: comparability of effects in humans and
animal models. Neurotoxicol Teratol. 12:231–237. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Mohora MBL, Boghianu L, Muscurel C, et al:
Effects of boric acid on redox status in the rat liver. Romanian J
Biophys. 12:77–82. 2002.
|
|
47
|
Hamby-Mason R, Chen JJ, Schenker S, Perez
A and Henderson GI: Catalase mediates acetaldehyde formation from
ethanol in fetal and neonatal rat brain. Alcohol Clin Exp Res.
21:1063–1072. 1997. View Article : Google Scholar : PubMed/NCBI
|