Introduction
Malaria remains one of the most serious infectious
diseases of the 21st century. The development of an 80% efficacious
malaria vaccine by 2025, which could offer protection for ≥4 years,
is a worldwide goal. RTS,S/AS01 is raising the bar in the
development and approval of a first-generation malaria vaccine that
has >50% protective efficacy against severe disease and
mortality and lasts longer than one year. RTS,S/AS01 is based on
the major Plasmodium circumsporozoite surface protein and
has reached phase III clinical trials; however, it is known that
this vaccine does not confer complete sterile protection (1).
The parasite invasion process involves the duffy
binding-like protein family (2,3) and
Plasmodium falciparum reticulocyte-binding protein homologs
(PfRHs) (4). Among these proteins,
PfRH5 is vital for erythrocyte invasion (5) and has become a promising vaccine
target (6). The protein basigin
has been identified as the erythrocyte receptor of PfRH5 and is
essential for the invasion of multiple strains of the pathogen
(7,8). In a previous study, PfRH5 had an
effect on several strains of P. falciparum, yet an antibody
of a part of PfRH5 was considered not to inhibit the P.
falciparum invasion (6).
Despite this, a small fragment may be more suitable for vaccine
design. Due to the lack of a suitable animal model (9), the in vivo immune effects of
partial and full-length fragments of PfRH5 remain unknown. The
present study established a rodent model to evaluate the effects of
P. falciparum.
Materials and methods
Animals and parasites
The protocols in this study were approved by Xiking
Hospital Medical Ethics Committee of the Fourth Military Medical
University (Xi’an, China). The animals were well cared for and
immunized using narcotic drug treatment to alleviate the pain. The
female BALB/c mice were obtained from the Experimental Animal
Center of the Fourth Military Medical University. A total of 40
mice used in the experiment were sacrificed by cervical vertebra
dislocation. Cryopreserved P. berghei ANKA parasites
(maintained in our laboratory) were thawed and passaged once in
vivo prior to being used to infect the experimental animals.
The P. falciparum CY and 3D7 strains (maintained in our
laboratory) were cultured with human blood cells in RPMI-1640
medium containing 10% human sera as designed by Trager and Jensen
(9). Two rounds of sorbitol
treatment were used to synchronize the asexual stages as described
previously (10).
Prediction of B-cell epitopes of
PfRH5
Bioinformatics methods, including Jameson-Wolf
(11), Garnier-Robson (12), Chou-Fasman (13) and Karplus-Schulz (14), were used to predict the signal
peptides, transmembrane domains, hydrophobicity, secondary and
tertiary structures, potential B-cell epitopes and other properties
of the B-cell epitope of PfRH5. The ability to predict the B-cell
epitopes of PfRH5 provided a basis for the preparation of the PfRH5
vaccines.
Expression and purification of
recombinant (r)PfRH5 protein
Antigenic prediction showed that the highest score
of the linear epitope region of PfRH5 was concentrated at amino
acid positions 200–400, suggesting that these amino acids may play
an important role in the function of PfRH5; therefore, the
corresponding DNA sequences for amino acids 200–300 (PfRH5-23),
300–400 (PfRH5-34) and the full-length amino acid sequence
(PfRH5-FL) were cloned into the prokaryotic expression vector
pET32a(+) (Novagen, Darmstadt, Germany) with BamHI and
HindIII sites. The nucleic acid sequences of interest were
codon optimized to permit expression in Escherichia coli.
Recombinant proteins were induced in the E. coli BL21 strain
(maintained in our laboratory) by the addition of 1 mM isopropyl
β-D-1-thiogalactopyranoside (IPTG) for 4 h. The fusion protein was
isolated by nitrilotriacetic acid (NTA) affinity chromatography.
PfRH5-34 and PfRH5-FL were expressed in the inclusion body of the
insoluble protein. Protein isolated from the inclusion bodies was
refolded and purified on Ni2+-NTA agarose resin (Qiagen,
Valencia, CA, USA) under native conditions. The expression and
purification of recombinant proteins was confirmed by western
blotting with His tag antibody (mouse monoclonal; 1:1,000; Beyotime
Institute of Biotechnology, Shanghai, China).
Antibody generation
Immunizations were conducted in eight-week-old
BALB/c female mice. Mice were immunized three times on days 0, 14,
28 and 40 by intraperitoneal injection of 20 μg fusion
protein (PfRH5-23, PfRH5-34 or PfRH5-FL) and an equal volume of
Freund’s adju vant (F5881; Sigma-Aldrich, St. Louis, MO, USA). The
mice sera were collected on day 40 and used in all of the following
experiments.
ELISA and indirect immunofluorescent
assay (IFA)
Anti-PfRH5 specific antibody titers in the sera were
determined using a standard ELISA procedure using rPfRH5 (prepared
by our department) (15). ELISA
was performed three times on each serum sample in 96-well
polyvinylchloride microtiter plates. The plates were coated with
0.2 μg/well purified rPfRH5 and left overnight at 4°C.
Following the coating, and between each incubation step, the plates
were washed three times with phosphate-buffered saline-Tween 20
(PBS-T). The plates were then blocked with 4% fat milk in PBS-T for
1 h at 37°C. Diluted serum samples were added to individual wells
and incubated for 1 h. Subsequent to washing, 100 μl
horseradish peroxidase-conjugated polyclonal sheep anti-mouse
immunoglobulin G (IgG) (1:5,000; Beyotime Institute of
Biotechnology) was added and the mixture was incubated for 1 h at
37°C. Color development was performed by the addition of 100
μl/well 3,3,5,5-tetramethylbenzidine (Sigma, St Louis, MO,
USA). Absorbance was read at 450 nm after 30 min using a plate
reader (Model-680; Bio-Rad, Hercules, CA, USA).
Giemsa stain
The thick blood film was left to dry in an incubator
at 37°C for 1 h and a fresh 4–5% Giemsa solution was prepared in
PBS (pH 7.1). The slides were placed on a staining rack and 1 ml
Giemsa solution (G4507; Sigma) was added onto the slides; the
slides were subsequently stained for 20 min. Following gentle
rinsing, the slides were left to dry in an upright position and
were observed under a microscope (magnification, ×100; Olympus
BX51; Olympus Corporation, Tokyo, Japan) using immersion oil
(BA-7003A; Baso Diagnostics Inc., Zhuhai, China).
Western blot analysis
A total of ~10 μg recombinant PfRH5-FL, PfRH5-23 and
PfRH5-34 were separated on a 15% SDS-PAGE gel. Following
electrophoresis, proteins were transferred to a nitrocellulose
membrane. After transfer, the membrane was blocked with 5% milk in
PBS-T for 1 h at room temperature and washed three times with
PBS-T. Membranes were incubated overnight at 4°C with a His-tagged
monoclonal mouse antibody (1:1,000 dilution in PBS-T; Beyotime
Institute of Biotechnology). Following washing three times with
PBS-T, the membranes were incubated with an HRP-conjugated
polyclonal goat anti-mouse antibody (1:5,000 dilution in PBS-T;
Beyotime Institute of Biotechnology). Detection was carried out
using an ECL kit (Millipore Corporation, Billerica, MA, USA).
In vitro assay of P. falciparum invasion
inhibition
The effect of mouse sera and rPfRH5 on the parasite
invasion of human red blood cells (RBCs) was assessed in three
experiments using an invasion inhibition assay. Synchronized
trophozoites of the P. falciparum CY and 3D7 strains were
purified by centrifugation at 1,500 × g and 4°C for 15 min on
Percoll gradients (16). To obtain
a final hematocrit of 2% and parasitemia of 0.2%,
~7.2×105 schizonts in 180 ml complete medium with 10%
human serum were mixed with 4×107 human RBCs. Each well
received 180 μl of the culture and 20 μl
heat-inactivated (56°C, 30 min) mouse anti-PfRH5 sera (or rPfRH5
protein). The medium was changed every 24 h. After incubation for
72 h, the number of parasites was determined; the parasites were
collected by centrifugation (3 min at 1,000 × g) and the
supernatant was removed by aspiration (17). Hoechst 33258 stain (no. 861405;
Sigma-Aldrich) in PBS (0.1 ml) was added to each well to cover the
complete surface of the well and left for 10 min at 37°C. The stain
was removed by centrifugation and the RBCs were washed twice with
PBS. Fluorescence intensity was read at 450 nm after 10 min using a
GENios Pro Reader (Tecan, Männedorf, Switzerland). The percent
inhibition was calculated using the fluorescence intensity data in
the pre-immunization serum controls as 100% invasion.
In vivo assay of P. berghei ANKA invasion
inhibition
Mice were immunized three times on day 40 by
intraperitoneal injection of 20 μg fusion protein (PfRH5-23,
PfRH5-34 or PfRH5-FL) and an equal volume of Freund’s adjuvant. The
mice were challenged by 1×105 P. berghei ANKA
RBCs with parasitemia of 5%. The survival time and parasitemia
levels of the mice were observed every day.
Results
B-cell epitope prediction, expression and
purification of rPfRH5
The sequence of PfRH5 from the P. falciparum
3D7 strain has been reported previously (GenBank accession no.
XM_001351508). Prediction of the B-cell epitope for the PfRH5
antigen is useful for the preparation of PfRH5 antibodies using a
synthetic peptide approach or for fusion with another protein.
Screening for the B-cell epitope of the PfRH5 antigen with
bioinformatics revealed that an abundance of the B-cell epitope was
located at amino acids 200–400. The DNA sequences for PfRH5-23,
PfRH5-34 and PfRH5-FL were optimized to permit expression in E.
coli. rPfRH5 expression and purification were observed
subsequent to IPTG induction (Fig.
1A). The PfRH5-34 proteins detected with sodium dodecyl
sulfate-polyacrylamide gel electrophoresis were slightly larger
than the expected size. Their diffuse appearance may indicate
misfolding of the rPfRH5 in the E. coli cells. The
recombinant PfRH5 bands were also detected using anti-His tag
antibody (Fig. 1B).
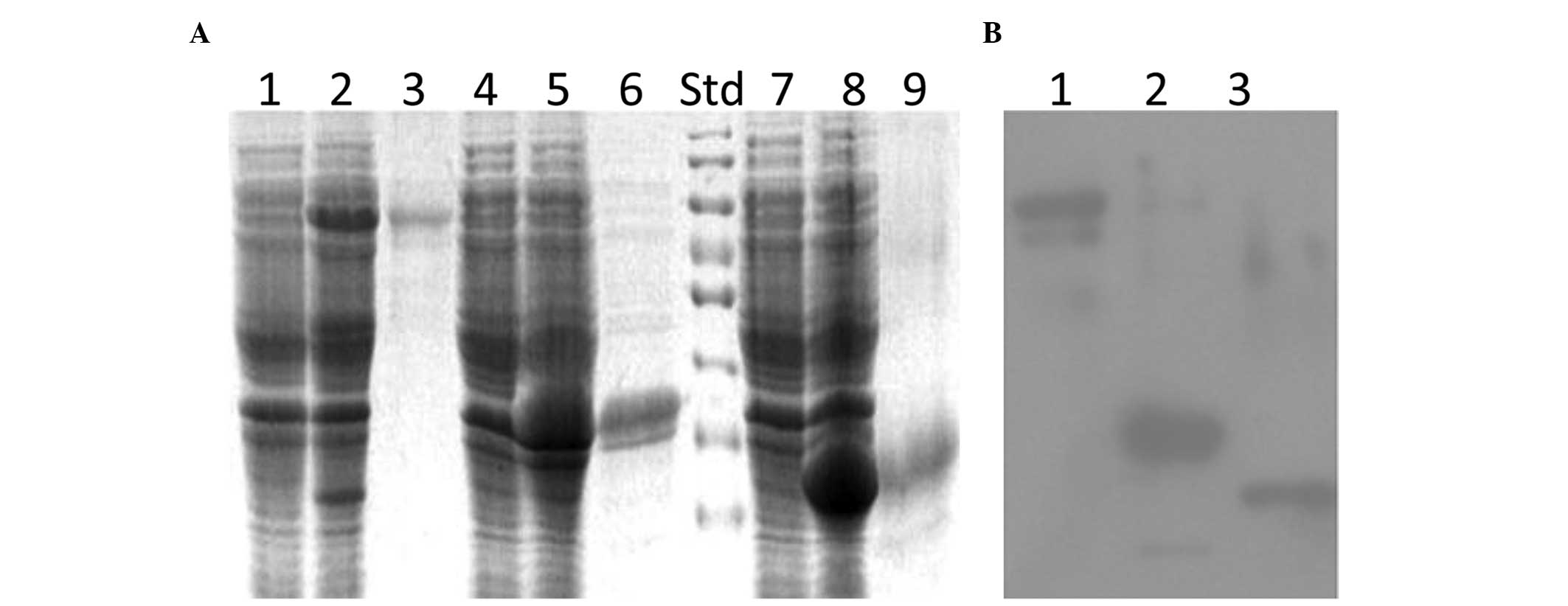 | Figure 1Production of recombinant PfRH5. (A)
Coomassie blue-stained sodium dodecyl sulfate-polyacrylamide gel
electrophoresis gel showing rPfRH5 fragment expression in
Escherichia coli and their purification by
Ni2+-NTA-agarose. Lanes 1, 4, and 7, uninduced; lanes 2,
5 and 8, rPfRH5-FL, rPfRH5-34 and rPfRH5-23 induced with 1 mM
isopropyl β-D-1-thiogalactopyranoside, respectively; lanes 3, 6 and
9, rPfRH5-FL, rPfRH5-34 and rPfRH5-23 protein purified under native
or denaturing conditions by Ni2+-NTA-agarose,
respectively. (B) Detection of rPfRH5 expression. Lanes 1
(rPfRH5-FL), 2 (rPfRH5-34) and 3 (rPfRH5-23), mouse anti-His
antibody. Std, protein standard; NTA, nitrilotriacetic acid;
rPfRH5, recombinant Plasmodium falciparum
reticulocyte-binding protein homolog 5. |
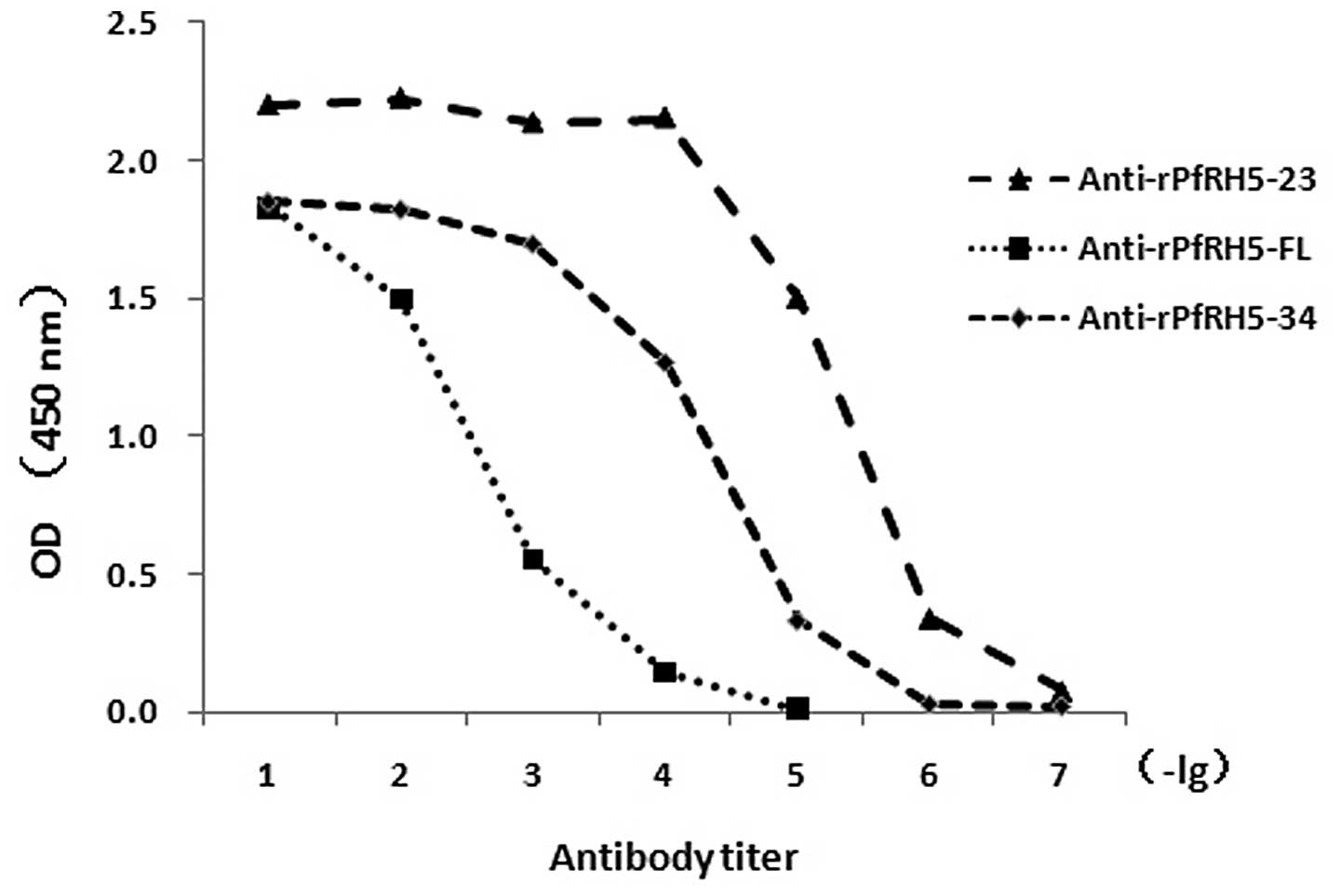
rPfRH5 vaccination
PfRH5-specific antibody titers increased over the
entire immunization course, and the final titers on day 40 reached
104–106 (Fig.
2). In addition, thin blood smears of P.
falciparum-infected RBCs were prepared for examination of the
antibody specificity against PfRH5 by IFA. Polyclonal anti-rPfRH5
antibodies (1:1,000) generated from mice were used as primary
antibodies. Following washing with PBS, the cells were incubated
with fluorescein isothiocyanate-conjugated anti-mouse IgG antibody
(Sigma). Immunofluorescence was visualized using fluorescence
microscopy (Olympus BX51; Olympus Corporation). Bright fluorescence
was observed when merozoites were released and contained in mature
schizonts using the rPfRH5 immune sera, whereas pre-immunization
sera showed only background fluorescence. These data suggested that
antibodies against rPfRH5 could detect native epitopes on PfRH5
(data not shown).
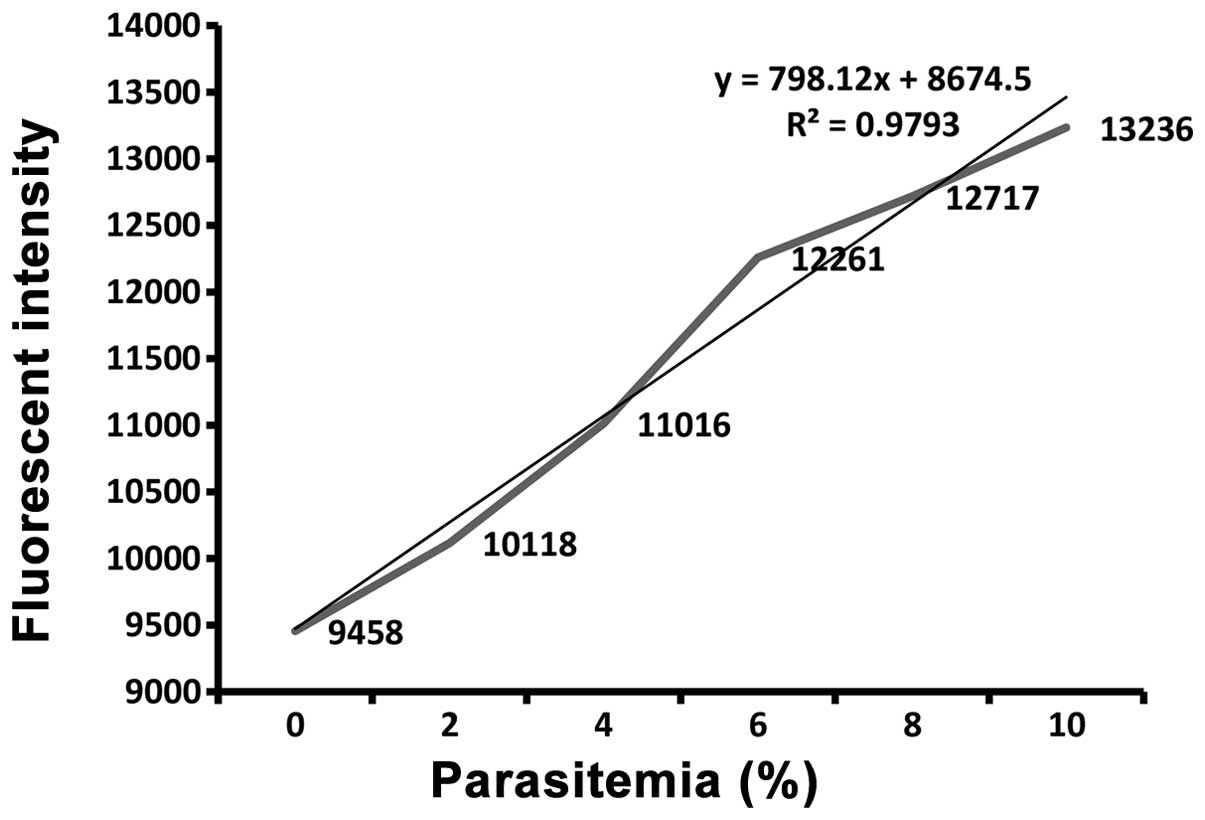
Easy detection of parasitemia
Fluorescence intensity-based evaluation of Hoechst
33258-stained parasites correlated well with the microscopic counts
of the Giemsa-stained smears (Fig.
3). The relevance of the Hoechst 33258 stain-based fluorescence
intensity method for detecting the parasite infection rates was
tested. The consistency of detection using fluorescence intensity
and counting under an oil immersion lens (18) was good.
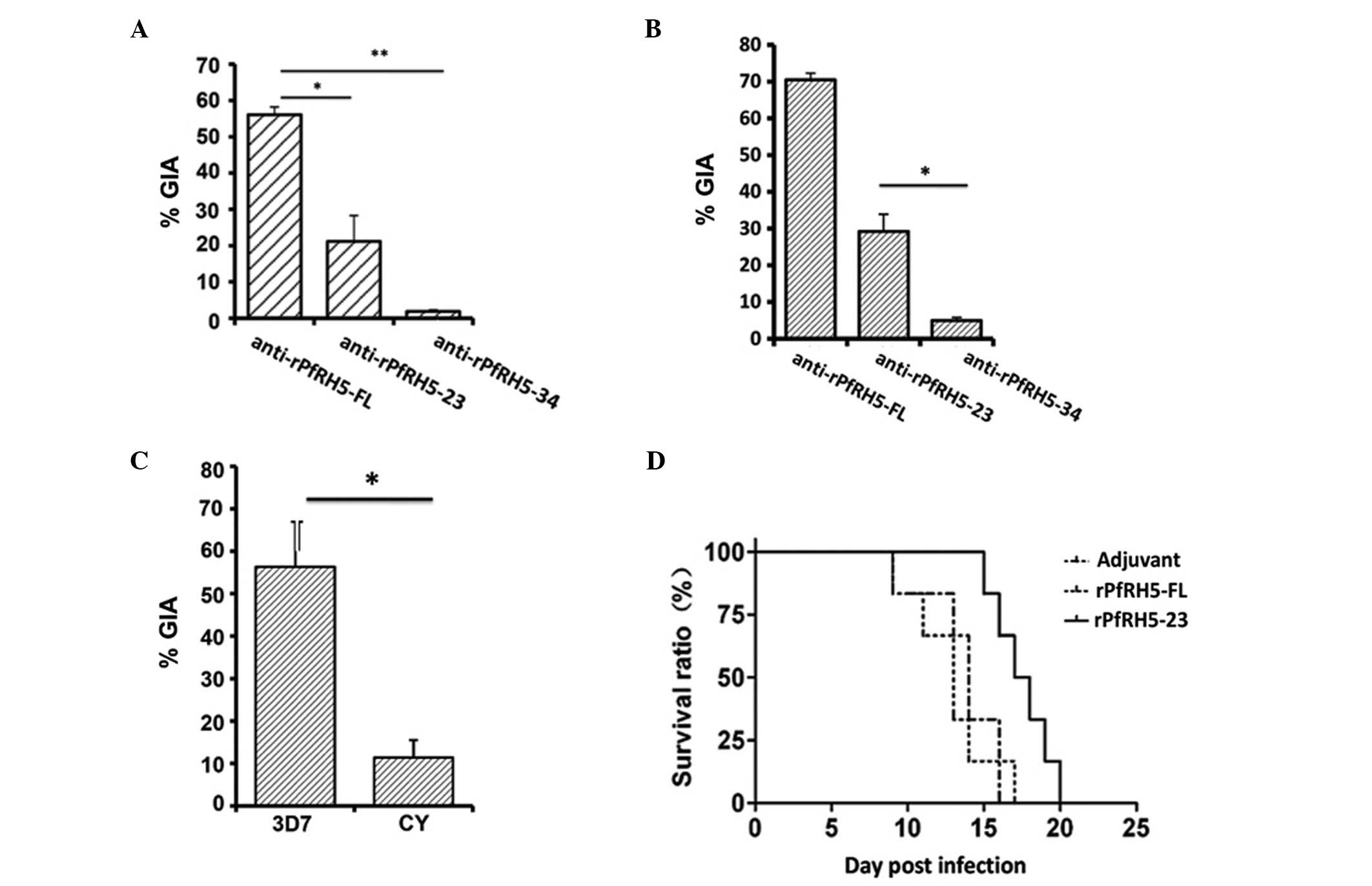
Inhibition of parasite invasion
In order to determine the effect of the immune sera
on merozoite invasion inhibition, schizonts from the P.
falciparum 3D7 strain were purified and used in an RBC invasion
assay. The results showed that antisera from the PfRH5-FL group
more effectively inhibited the merozoite invasion than antisera
from the PfRH5-23 and PfRH5-34 groups (Fig. 4A), suggesting that the in
vitro merozoite invasion inhibition effect is potentially
associated with the PfRH5-FL antigen. Of note, the PfRH5-23 was
more effective in merozoite invasion inhibition than the control
protein rPfRH-34 (Fig. 4B), and
antisera from the PfRH5-FL group were not found to be effective in
the RBC invasion assay for the P. falciparum CY strain
(Fig. 4C). The mice were
challenged by P. berghei ANKA. The parasitemia of the
PfRH5-34 and PfRH5-FL groups was higher than that of the control
(data not shown), while the parasitemia of the PfRH5-23 group was
lower than that of the control. Furthermore, the survival time of
the PfRH5-23 group was longer than that of any other group,
including the control, suggesting that any in vivo merozoite
invasion inhibition effect is likely to be associated with the
PfRH5-23 antigen (Fig. 4D).
Discussion
Vaccination against malaria is generally presumed to
be the most cost-effective way of protecting against the disease,
and it is hoped that it can potentially eradicate malaria from the
world. Studies (19–21) have shown different transcription
and expression patterns for RH genes in several P.
falciparum parasite strains and isolates. PfRH5 does not change
its expression levels in P. falciparum strains among
different invasion routes and it is recognized in sera from
patients suffering from natural malarial infection (15), which suggests that it is an
excellent candidate for inclusion as a component in a fully
effective anti-malarial vaccine. Reports published during the past
decade (15,22,23)
have shown that PfRH5 fragments bind to receptors on the RBC
membrane.
In the present study, antibodies against PfRH5 and
PfRH5 fragments were obtained in mice for the evaluation of P.
falciparum 3D7 and CY strain schizont inhibition. High titers
of PfRH5 and PfRH5 fragment antibodies that specifically recognized
PfRH5 in late-stage schizonts were revealed by IFAs similar to
those reported for other RH family members (data not shown)
(24–26). Studies have shown the crucial role
of PfRH5 in RBC invasion due to the association between PfRH5 and
basigin (7,8), suggesting that PfRH5 may be a
candidate for inclusion in a fully effective anti-malarial vaccine.
In this light, recent studies (27–30)
showing that vaccine-induced anti-PfRH5 antibodies can potently
inhibit different parasite strain invasions in vitro are
encouraging and suggest that future malaria vaccine research may
focus heavily on this basigin-PfRH5 interaction.
In the present study, however, contradictory results
were obtained: i) Inhibition of the parasite invasion was not
observed in the P. falciparum CY strain; ii) the PfRH5-23
protein could prolong survival time in vivo, but the
PfRH5-FL protein did not exhibit the same results, despite having
the ability to inhibit parasite invasion in vitro. Although
studies in mice do not meet the evaluation criteria for human P.
falciparum vaccines, the murine malaria model is useful.
Furthermore, in vitro studies of vaccine-induced mouse
antibodies are easier to conduct than in vivo studies.
Whereas known quantities of antibodies in studies in vitro
can be administered, the quantity and specificity of animal
antibodies produced against each vaccine cannot be controlled in
vivo and may vary substantially between individuals. In
conclusion, PfRH5-FL may not be as promising as was hoped. By
blocking parasite invasion through PfRH5, the vaccine appears to be
some distance away from being able to halt malaria. The
identification of basigin, a well-characterized membrane protein,
as a receptor that is essential for malaria infection, is likely to
contribute significantly to the prevention and treatment of
malaria.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (nos. 30600527 and 30901370).
Valuable comments from other members of the laboratories were
appreciated.
References
|
1
|
Agnandji ST, Lell B, Soulanoudjingar SS,
Fernandes JF, Abossolo BP, Conzelmann C, Methogo BG, Doucka Y,
Flamen A, Mordmüller B, et al; RTS,S Clinical Trials Partnership.
First results of phase 3 trial of RTS,S/AS01 malaria vaccine in
African children. N Engl J Med. 365:1863–1875. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tolia NH, Enemark EJ, Sim BK and
Joshua-Tor L: Structural basis for the EBA-175 erythrocyte invasion
pathway of the malaria parasite Plasmodium falciparum. Cell.
122:183–193. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jaśkiewicz E, Graczyk J and Rydzak J:
Proteins involved in invasion of human red blood cells by malaria
parasites. Postepy Hig Med Dosw (Online). 64:617–626. 2010.(In
Polish).
|
|
4
|
Lopaticki S, Maier AG, Thompson J, Wilson
DW, Tham WH, Triglia T, Gout A, Speed TP, Beeson JG, Healer J and
Cowman AF: Reticulocyte and erythrocyte binding-like proteins
function cooperatively in invasion of human erythrocytes by malaria
parasites. Infect Immun. 79:1107–1117. 2011. View Article : Google Scholar :
|
|
5
|
Nolan T: Identifying an essential
interaction between malaria parasites and erythrocytes unlocks the
door to promising vaccine targets. Pathog Glob Health. 106:642012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Douglas AD, Williams AR, Illingworth JJ,
Kamuyu G, Biswas S, Goodman AL, Wyllie DH, Crosnier C, Miura K,
Wright GJ, et al: The blood-stage malaria antigen PfRH5 is
susceptible to vaccine-inducible cross-strain neutralizing
antibody. Nat Commun. 2:6012011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Crosnier C, Bustamante LY, Bartholdson SJ,
Bei AK, Theron M, Uchikawa M, Mboup S, Ndir O, Kwiatkowski DP,
Duraisingh MT, Rayner JC and Wright GJ: Basigin is a receptor
essential for erythrocyte invasion by Plasmodium falciparum.
Nature. 480:534–537. 2011.PubMed/NCBI
|
|
8
|
Muramatsu T: Basigin: a multifunctional
membrane protein with an emerging role in infections by malaria
parasites. Expert Opin Ther Targets. 16:999–1011. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Trager W and Jensen JB: Human malaria
parasites in continuous culture. 1976. J Parasitol. 91:484–486.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cui L, Rzomp KA, Fan Q, Martin SK and
Williams J: Plasmodium falciparum: differential display analysis of
gene expression during gametocytogenesis. Exp Parasitol.
99:244–254. 2001. View Article : Google Scholar
|
|
11
|
Jameson BA and Wolf H: The antigenic
index: a novel algorithm for predicting antigenic determinants.
Comput Appl Biosci. 4:181–186. 1988.PubMed/NCBI
|
|
12
|
Garnier J, Osguthorpe DJ and Robson B:
Analysis of the accuracy and implications of simple methods for
predicting the secondary structure of globular proteins. J Mol
Biol. 120:97–120. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Creighton TE: Prediction of Protein
Structure and the Principles of Protein Conformation. Fasman Gerald
D: Plenum; New York: 1989, pp. xivpp. 798illus $95. Science. 247.
pp. 1351–1352. 1990
|
|
14
|
Karplus PA and Schulz GE: Prediction of
chain flexibility in proteins. Naturwissenschaften. 72:212–213.
1985. View Article : Google Scholar
|
|
15
|
Baum J, Chen L, Healer J, Lopaticki S,
Boyle M, Triglia T, Ehlgen F, Ralph SA, Beeson JG and Cowman AF:
Reticulocyte-binding protein homologue 5 - an essential adhesin
involved in invasion of human erythrocytes by Plasmodium
falciparum. Int J Parasitol. 39:371–380. 2009. View Article : Google Scholar
|
|
16
|
Wahlgren M, Berzins K, Perlmann P and
Björkman A: Characterization of the humoral immune response in
Plasmodium falciparum malaria. I Estimation of antibodies to P
falciparum or human erythrocytes by means of microELISA. Clin Exp
Immunol. 54:127–134. 1983.PubMed/NCBI
|
|
17
|
Miao J, Li X, Liu Z, Xue C, Bujard H and
Cui L: Immune responses in mice induced by prime-boost schemes of
the Plasmodium falciparum apical membrane antigen 1 (PfAMA1)-based
DNA, protein and recombinant modified vaccinia Ankara vaccines.
Vaccine. 24:6187–6198. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jogdand PS, Singh SK, Christiansen M,
Dziegiel MH, Singh S and Theisen M: Flow cytometric readout based
on Mitotracker Red CMXRos staining of live asexual blood stage
malarial parasites reliably assesses antibody dependent cellular
inhibition. Malar J. 11:2352012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Taylor HM, Grainger M and Holder AA:
Variation in the expression of a Plasmodium falciparum protein
family implicated in erythrocyte invasion. Infect Immun.
70:5779–5789. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Duraisingh MT, Triglia T, Ralph SA, Rayner
JC, Barnwell JW, McFadden GI and Cowman AF: Phenotypic variation of
Plasmodium falciparum merozoite proteins directs receptor targeting
for invasion of human erythrocytes. EMBO J. 22:1047–1057. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nery S, Deans AM, Mosobo M, Marsh K, Rowe
JA and Conway DJ: Expression of Plasmodium falciparum genes
involved in erythrocyte invasion varies among isolates cultured
directly from patients. Mol Biochem Parasitol. 149:208–215. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hayton K, Gaur D, Liu A, Takahashi J,
Henschen B, Singh S, Lambert L, Furuya T, Bouttenot R, Doll M, et
al: Erythrocyte binding protein PfRH5 polymorphisms determine
species-specific pathways of Plasmodium falciparum invasion. Cell
Host Microbe. 4:40–51. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rodriguez M, Lustigman S, Montero E, Oksov
Y and Lobo CA: PfRH5: a novel reticulocyte-binding family homolog
of Plasmodium falciparum that binds to the erythrocyte, and an
investigation of its receptor. PloS One. 3:e33002008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rayner JC, Galinski MR, Ingravallo P and
Barnwell JW: Two Plasmodium falciparum genes express merozoite
proteins that are related to Plasmodium vivax and Plasmodium yoelii
adhesive proteins involved in host cell selection and invasion.
Proc Natl Acad Sci USA. 97:9648–9653. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Triglia T, Duraisingh MT, Good RT and
Cowman AF: Reticulocyte-binding protein homologue 1 is required for
sialic acid-dependent invasion into human erythrocytes by
Plasmodium falciparum. Mol Microbiol. 55:162–174. 2005. View Article : Google Scholar
|
|
26
|
Stubbs J, Simpson KM, Triglia T, Plouffe
D, Tonkin CJ, Duraisingh MT, Maier AG, Winzeler EA and Cowman AF:
Molecular mechanism for switching of P. falciparum invasion
pathways into human erythrocytes. Science. 309:1384–1387. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bustamante LY, Bartholdson SJ, Crosnier C,
Campos MG, Wanaguru M, Nguon C, Kwiatkowski DP, Wright GJ and
Rayner JC: A full-length recombinant Plasmodium falciparum PfRH5
protein induces inhibitory antibodies that are effective across
common PfRH5 genetic variants. Vaccine. 31:373–379. 2013.
View Article : Google Scholar :
|
|
28
|
Williams AR, Douglas AD, Miura K,
Illingworth JJ, Choudhary P, Murungi LM, Furze JM, Diouf A, Miotto
O, Crosnier C, et al: Enhancing blockade of Plasmodium falciparum
erythrocyte invasion: assessing combinations of antibodies against
PfRH5 and other merozoite antigens. PLoS Pathog. 8:e10029912012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lopez-Perez M, Villasis E, Machado RL,
Póvoa MM, Vinetz JM, Blair S, Gamboa D and Lustigman S: Plasmodium
falciparum field isolates from South America use an atypical red
blood cell invasion pathway associated with invasion ligand
polymorphisms. PLoS One. 7:e479132012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Villasis E, Lopez-Perez M, Torres K,
Gamboa D, Neyra V, Bendezu J, Tricoche N, Lobo C, Vinetz JM and
Lustigman S: Anti-Plasmodium falciparum invasion ligand antibodies
in a low malaria transmission region, Loreto, Peru. Malar J.
11:3612012. View Article : Google Scholar : PubMed/NCBI
|