Introduction
Cancer is a major human disease that causes a
considerable economic burden to health institutions worldwide.
Although significant progress has been made in cancer treatment,
the incidence and mortality rates for almost every type of cancer
remain high (1). Thus, further
research is required to develop safe and effective drugs for the
treatment of human cancers. Among the various types of cancer,
primary malignant osteosarcoma predominantly affects the growing
bones of adolescents and children, and is characterized by locally
aggressive growth and an early metastatic potential (2). The incidence rate of osteosarcoma among
younger individuals (<20 years-old) is 8.7 per million, and this
rate is higher in males compared with females (3). Respiratory failure due to lung
metastasis is the major cause of mortality in patients, despite
significant improvements in clinical treatments with combination
intensive chemotherapy and surgical resection (4,5).
However, patients frequently suffer from intolerable, acute and
long-term toxicities following the administration of approved
chemotherapeutic agents (6). Thus,
more effective and improved treatment strategies are required to
prevent the progression of osteosarcoma.
Withanolides (steroidal lactones) have been isolated
from various species of the Solanaceae family, including
Withania somnifera (W. somnifera), which is widely
distributed across the South Asian subcontinent (7). In the Ayurveda system of medicine,
Withania is noted for its aphrodisiac, sedative and
life-prolonging properties (8).
Extracts from different parts of W. somnifera have been
evaluated for various biological activities, such as cholinesterase
inhibition (9,10), anti-inflammatory properties via
cyclooxygenase-2 enzyme inhibition from leaf extracts (11), antibacterial properties (12) and sex hormone deficiency regulation
in rats with diabetes (13). The
isolation and identification of several withanolides with
anti-glycation (14) and antipyretic
activities (15) from
Withania has been reported. Among the reported withanolides,
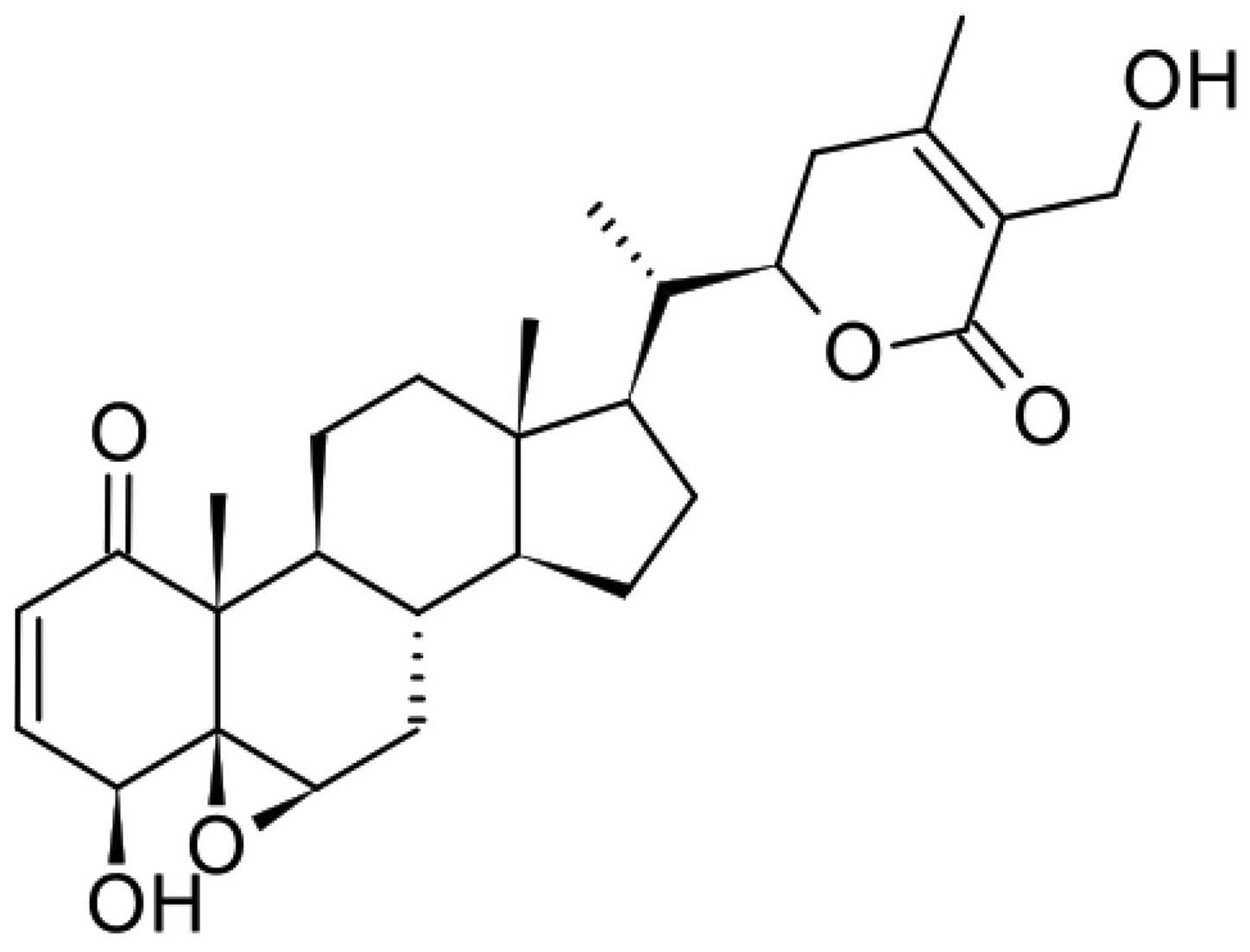
withaferin A (WA) (16) (as seen in
Fig. 1) and ashwagandhanolide
(17), obtained from the leaves and
roots, respectively, are well-known potent secondary metabolites of
W. somnifera and are reported to possess anticancer
properties. Antiproliferative, antimetastatic, antiangiogenic,
anti-invasive and proapoptotic activities of withanolides have been
reported, with the underlying mechanism found to be associated with
the suppression of nuclear factor (NF)-κB and NF-κB-regulated gene
products (18). Previous studies
have demonstrated the anticancer activity of WA in prostate
(19,20), breast (21), leukemia (22) and melanoma cancer cells (23). In addition, a previous study
demonstrated that WA induces apoptosis in prostate cancer cells
through Par-4 induction (19) and
inhibits IκB kinase activation via a thioalkylation-sensitive redox
mechanism (24); by covalently
modifying the cysteine residue, WA targets the intermediate
filament protein, vimentin (25).
Furthermore, WA has been shown to induce actin microfilament
aggregation by targeting annexin II (26). The aim of the present study was to
investigate the antiproliferative effects of WA on two osteosarcoma
cell lines (U2OS and MG-63) and determine the mechanism underlying
the induction of cell cycle arrest.
Materials and methods
Reagents
WA, bovine serum albumin (BSA), ribonuclease A
(RNase A), propidium iodide (PI), sulforhodamine B (SRB) and a
rabbit polyclonal anti-β-actin antibody (1:1,000; cat. no. A2668)
were purchased from Sigma-Aldrich (St. Louis, MO, USA). Fetal
bovine serum (FBS), RPMI 1640 medium, trypsin-EDTA and
antibiotic-antimycotic were purchased from Gibco Life Technologies
(Grand Island, NY, USA). The following antibodies: Rabbit
polyclonal Cdk2 (1:1,000; cat. no. sc-163), rabbit polyclonal
cyclin A (1:1,000; sc-751), mouse monoclonal cyclin B1 (1:1,000;
cat. no. sc-245) and mouse monoclonal Cdc2 (1:1,000; cat. no.
sc-54) were purchased from Santa Cruz Biotechnology, Inc. (Santa
Cruz, CA, USA). The following antibodies: Rabbit polyclonal p-Cdc2
(Tyr15; 1:1,000; cat. no. P06493), rabbit polyclonal p-Chk1
(Ser345; 1:1,000; cat. no. 2341S), rabbit polyclonal p-Chk2 (Thr68;
1:1,000; cat. no. 2661), rabbit polyclonal Chk1 (1:1,000; cat. no.
2344S) and rabbit polyclonal Chk2 (1:1,000; cat. no. 2662) were
obtained from Cell Signaling Technology, Inc. (Danvers, MA,
USA).
Ethics and cell lines
U2OS and MG-63 human osteosarcoma cell lines were
obtained from the China Center for Type Culture Collection (Wuhan,
China), following standard ethical procedures (27). Following isolation, the osteosarcoma
cells were identified based on histological type and grade. The
cell lines were grown in high glucose Dulbecco's modified Eagle's
medium supplemented with 10% FBS, 100 U/ml penicillin G and 100
U/ml streptomycin (Sigma-Aldrich). The cells were subsequently
incubated in a humidified incubator at 37°C with 5% CO2.
All cells used in the experiments were in the logarithmic
phase.
Cell proliferation assay
The antiproliferation potential of WA in U2OS and
MG-63 cells was examined using an SRB assay (11). Cell lines were seeded at a density of
9×103 cells/well into 96-well plates and allowed to
adhere for 72 h. The cells were subsequently treated with WA at
various concentrations (0.1, 1 and 10 µM). Dimethyl sulfoxide
(DMSO) was used as the experimental control. The results were
presented as percentages relative to the solvent-treated control
incubations. Using nonlinear regression analysis, the
IC50 values were calculated.
Cell cycle analysis
U2OS and MG-63 cell lines were plated into 35-mm
culture dishes at a density of 1×106 cells/100 mm and
incubated for 24 h. The medium was replaced with fresh medium
containing WA (at concentrations of 0.1, 1 and 10 µM) and 0.1% DMSO
alone. Following incubation for 24 h at 37°C, the cells were
harvested, washed with phosphate-buffered saline and fixed with 70%
(v/v) ice-cold ethanol for 1 h at 4°C. Subsequently, 50 µg/ml RNase
A and 50 µg/ml PI were added to the fixed cells. In total, ~10,000
cells were analyzed by flow cytometry (BD Accuri™ C6; BD
Biosciences, Franklin Lakes, NJ, USA). All experiments were
repeated three times and data analysis was recorded using BD
CellQuest™ cell cycle analysis software (BD Biosciences).
Western blot analysis
U2OS and MG-63 cell lines were incybated with
varying concentrations of WA (0.1, 1 and 10 µM) for 24 h. The cell
samples were lysed in sample buffer [150 mM Tris (pH 6.8), 8 M
urea, 50 mM DTT, 2% sodium dodecyl sulfate, 15% sucrose, 2 mM EDTA,
0.01% bromophenol blue and 1% protease and phosphatase inhibitor
cocktails], sonicated (Ultrasonic Homogenizer 300MP; BioLogics,
Manassas, VA, USA) and the protein concentrations were recorded
using the Lowry protein assay method (28). Subsequently, samples of 40–45 µg were
refluxed, run through an 8–12% Bis/Tris gel (Invitrogen Life
Technologies, Carlsbad, CA, USA) using 5X
2-(N-morpholino)ethanesulfonic buffer (Sigma-Aldrich) and
transferred to an Immobilon-nitrocellulose membrane
(Sigma-Aldrich). The membrane was blocked with 5% skimmed milk or
3% BSA in Tris-buffered saline Tween-20 [TBST; 150 mM NaCl, 50 mM
Tris (pH 7.5) and 0.1% Tween-20] (blocking buffer). Next, the
membrane was probed with antibodies targeted against cyclin A,
cyclin B1, Cdk2, p-Cdc2 (Tyr15), Cdc2, p-Chk1 (Ser345), Chk1,
p-Chk2 (Thr68), Chk2 and β-actin (1:1,000) overnight at 4°C in
blocking buffer. Following washing with TBST, the membranes were
probed with anti-rabbit (cat. no. 7074; Cell Signalling Technology,
Inc.) and goat anti-mouse (cat. no. 11011MP; Life Technologies,
Grand Island, NY, USA) immunoglobulin G secondary antibodies
(1:5,000) in blocking buffer at room temperature for 90 min, and
after washing again, the fluorescence was subsequently detected
using a Bio-Rad imaging system (Bio-Rad Laboratories, Hercules CA,
USA).
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR) assay
In the RT-qPCR analysis, the two cell lines were
treated with 10 µM WA for 12 h. Extraction of the total cellular
RNA was performed using TRIzol® reagent (Invitrogen Life
Technologies), according to the manufacturer's instructions. For
reverse transcription of the RNA into cDNA, a SuperScript III
First-Strand Synthesis kit (Invitrogen Life Technologies) was used.
Subsequently, the qPCR was conducted using an ABI Prism 7900HT
Real-Time PCR system (PerkinElmer, Inc., Waltham, MA, USA) with
SYBR Green PCR Master Mix (Applied Biosystems Life Technologies,
Foster City, CA, USA). The primers used in the RT-qPCR analysis are
presented in Table I. The mRNA
expression levels were recorded as the fold change relative to the
control. Following completion of the RT-qPCR, cycle numbers (Ct
values) in which the signal intensity was equal to the threshold
value were obtained from the software (SDS Plate Utility v2.3;
Applied Biosystems Life Technologies). The relative expression
values were calculated using the 2−ΔΔCt method (29).
 | Table I.Primer sequences used for reverse
transcription quantitative polymerase chain reaction. |
Table I.
Primer sequences used for reverse
transcription quantitative polymerase chain reaction.
| Gene | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| Cdk2 |
GAAACTCTGAAGCCGACCAG |
GCCCTCTCAGTGTCCAGAAG |
| Cyclin B1 |
CGGGAAGTCACTGGAAACAT |
AAACATGGCAGTGACACCAA |
| Cyclin A1 |
GTCAGAGAGGGGATGGCAT |
CCAGTCCACCAGAATCGTG |
| Cdc2 |
GGTTCCTAGTACTGCAATTCG |
TTTGCCAGAAATTCGTTTGG |
| Chk1 |
GGTGCCTATGGAGAAGTTCAA |
TCTACGGCACGCTTCATATC |
| Chk2 |
CGGATGTTGAGGCTCACGA |
TATGCCCTGGGACTGTGAGG |
| β-actin |
GCTCGTCGTCGACAACGGCTC |
CAAACATGCTCTGGGTCATCTTCTC |
Statistical analysis
All experiments were repeated a minimum of three
times. Data are expressed as the mean ± standard deviation. The
treated groups were compared using one-way analysis of variance
with SPSS 12.0 software (SPSS, Inc., Chicago, IL, USA), where
P<0.05 was considered to indicate a statistically significant
difference. Relative percentages were calculated using GraphPad
Prism software, version 4.0 for Windows (GraphPad Software, Inc.,
San Diego, CA, USA).
Results
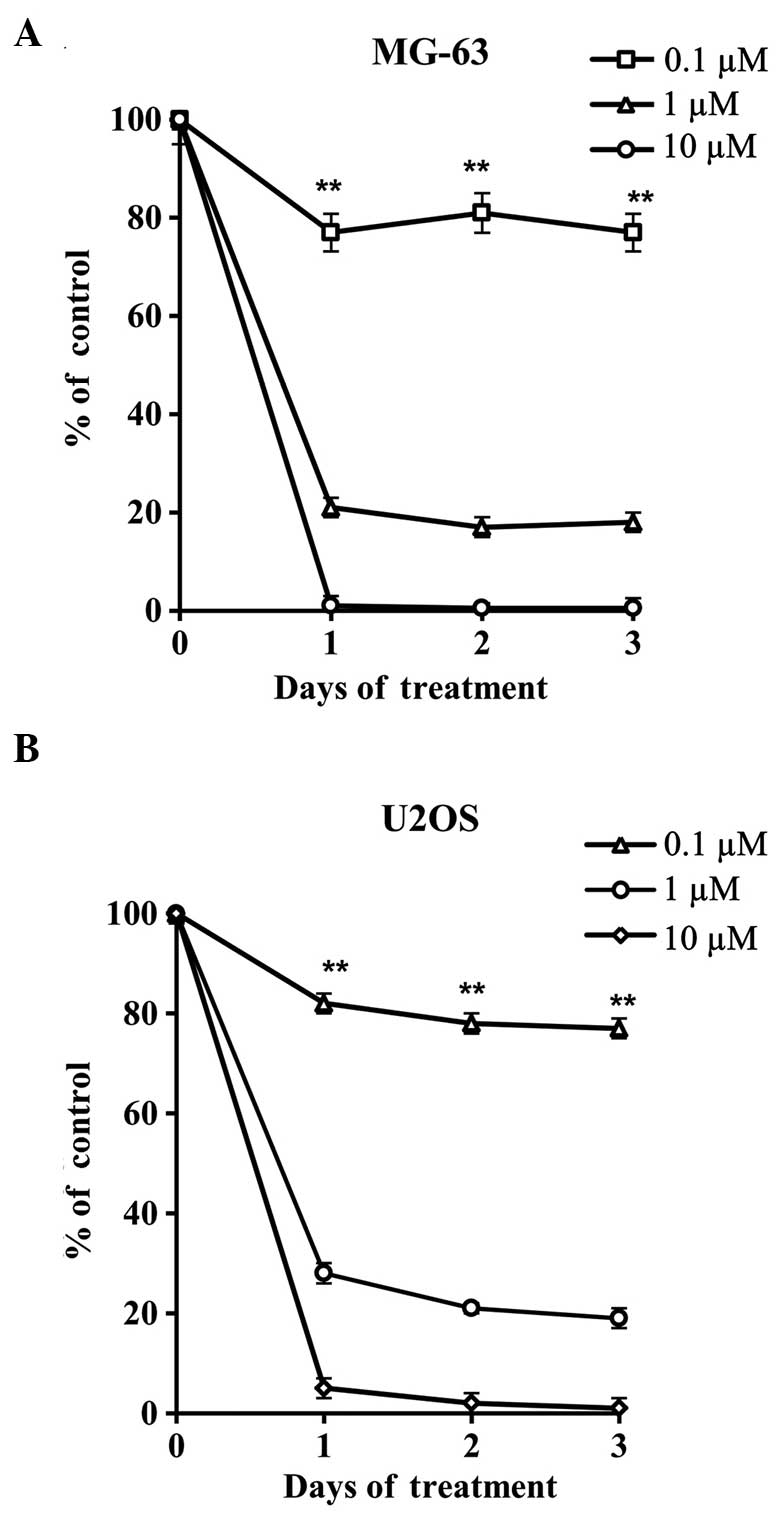
WA prevents the proliferation of MG-63
and U2OS osteosarcoma cells
Two human osteosarcoma cells lines, MG-63 (Fig. 2A) and U2OS (Fig. 2B), were treated with varying
concentrations of WA for 72 h to assess the antiproliferative
effect of WA. The cell proliferation of each of the tested samples
was recorded using an SRB assay. Higher concentrations of WA were
shown to exhibit potent antiproliferative effects. Thus, the
results revealed that WA produced a potent antiproliferative effect
by significantly inhibiting the growth of the tested cell
lines.
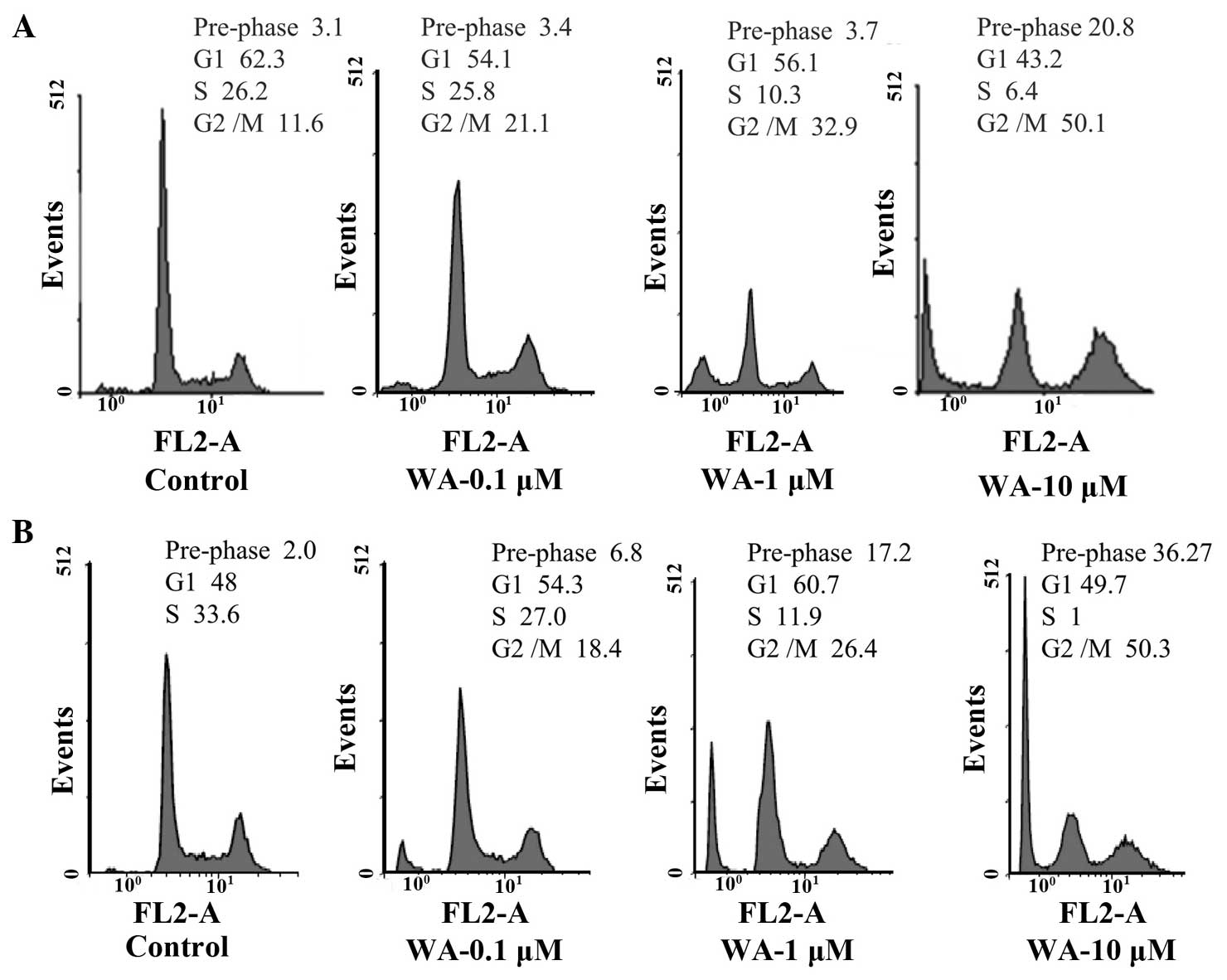
Effect of WA on cell cycle
distribution
MG-63 and U2OS cells were treated with varying
concentrations of WA for 24 h and the underlying antiproliferative
mechanisms were investigated via flow cytometric analysis with PI
staining. Cell cycle distribution analysis revealed that WA
administration resulted in the significant arrest of MG-63 and U2OS
human osteosarcoma cells at the G2/M phase in a dose-dependent
manner (Fig. 3). At a concentration
of 10 µM WA, more than half of the cells were arrested at the G2/M
phase.
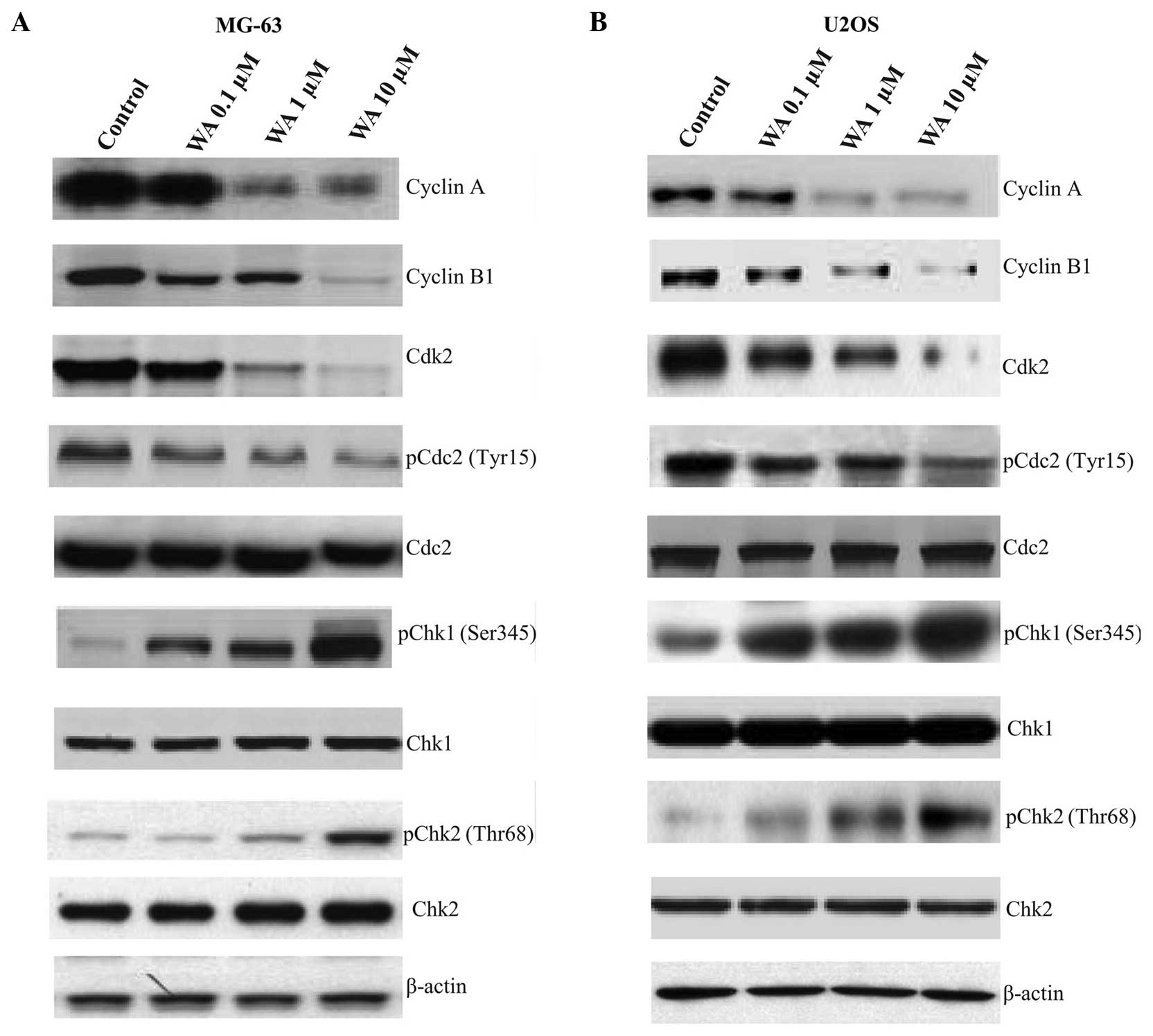
Western blot analysis on the
expression levels of G2/M phase cell cycle checkpoint proteins
Western blot analysis was used to examine the
expression levels of G2/M cell cycle regulatory proteins in MG-63
(Fig. 4A) and U2OS (Fig. 4B) human osteosarcoma cell lines
treated with WA, in order to determine whether the cell cycle
arrest was associated with the regulation of cell cycle checkpoint
proteins. There was a significant decrease in the levels of cyclin
A, cyclin B1, Cdk2 and p-Cdc2 (Tyr15) in the MG-63 and U2OS cells;
however, the protein expression levels of p-Chk1 (Ser345) and
p-Chk2 (Thr68) increased. No statistically significant changes were
observed in the levels of Cdc2 and Chk2 in the two cell lines
following WA treatment.
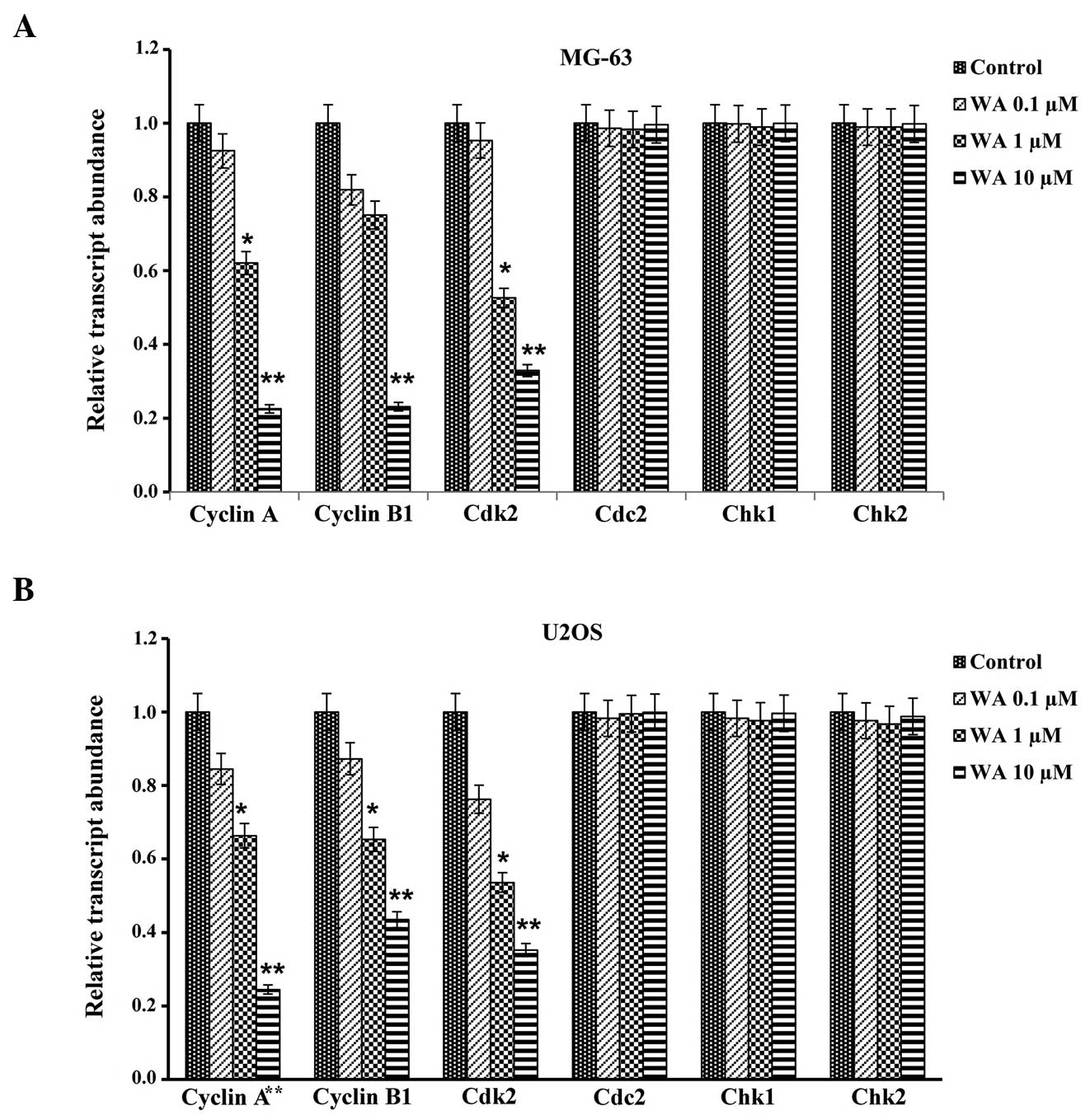
Effect of WA on the mRNA expression
levels of the cell cycle checkpoint proteins
In order to determine the effect of WA on the mRNA
expression levels of cell cycle checkpoint proteins, the expression
levels of G2/M cell cycle regulatory proteins, including cyclin A,
cyclin B1, Cdk2, Cdc2, Chk1 and Chk2, were elucidated by RT-qPCR
analysis for the MG-63 (Fig. 5A) and
U2OS (Fig. 5B) cell lines. There was
a significant decrease in the mRNA expression levels of cyclin A,
cyclin B1 and Cdk2, while no marked changes were observed in the
mRNA expression levels of Cdc2, Chk1 and Chk2 in the two cell
lines.
Discussion
WA is a well-known steroidal lactone from the
withanolides group. These are major constituents identified from
the important medicinal plant, Withania somnifera, and the
related Solanaceae family (30). A
number of biological properties of withanolides have been reported,
including anti-inflammatory, antitumor, antibacterial,
antidepressant, antioxidant, antiulcer, cytotoxic, quinone
reductase inducive, antileishmanial, antitrypanosomal,
immunosuppressive, cognition-enhancing and memory-improving
effects, as well as hypotensive, bradycardic and
respiratory-stimulating properties (31–33).
Withanolides have been studied extensively, and >130
withanolides are known and >40 withanolides have been isolated
(34). Certain newly-isolated
withanolides, including withangulatins B, C, G, H and I (33), have been shown to exert cytotoxic
activity. Withanolide A possesses a reactive enone moiety in the A
ring, an epoxy alcohol in the B ring, as well as a tetrasubstituted
unsaturated lactone side chain. The main synthetic challenges arise
from the stereoselective construction of the side chain, the
oxidation pattern of the A and B ring, and the diastereoselective
instalment of a tertiary alcohol at C20 (35). Furthermore, structure-activity
relationship studies have confirmed that the unsaturated ring A and
epoxide play an important role in the cytotoxic activity of
withanolides (32). Withangulatin A
has been shown to inhibit topoisomerase II and induce a heat shock
response (36,37). Zaarur et al (38) suggested that withangulatin A may
induce the heat shock response by inhibiting Hsp90 activity.
The present study investigated the anticancer
potential of WA in MG-63 and U2OS human osteosarcoma cancer cell
lines. The results revealed that WA exhibited potent
antiproliferative effects. Data from the cell cycle distribution
analysis showed that WA treatment at a concentration of 10 µM
caused a significant accumulation of treated cells that were
arrested in the G2/M phase; this effect was shown to be
dose-dependent. Furthermore, western blot analysis revealed that WA
significantly regulated the protein expression levels of G2/M cell
cycle regulatory proteins in the two cell lines.
The key component involved in G2 to M phase
transition is cyclin B1 (39). In
addition, cyclin A/Cdk2 complexes are known to participate in the
initiation of mitosis in human cancer cells (40). The DNA damage sensors, Chk1 and Chk2,
participate in G2/M checkpoint control through the
ataxia-telangiectasia mutated (ATM)/ATM-RAD3-related (ATR) pathway.
In the current study, a significant decrease in the expression
levels of cyclin A, cyclin B1, Cdk2 and p-Cdc2 (Tyr15) were
observed, whereas the expression levels of p-Chk1 (Ser345) and
p-Chk2 (Thr68) increased significantly in the two treated cell
lines. At a concentration of 10 µM, WA was found to exhibit the
maximal effect on protein expression and cell cycle arrest. At a
transcription level, decreases in the mRNA expression levels of
cyclin A, cyclin B1 and Cdk2 were observed, while no change was
observed in the mRNA expression levels of Cdc2, Chk1 and Chk2 in
the two treated cell lines.
In conclusion, the results of the present study
demonstrated that the antiproliferative effects of WA on the human
osteosarcoma cell lines, MG-63 and U2OS, at the G2/M phase were
primarily associated with cell cycle arrest. These observations
indicate that WA may be used in human osteosarcoma therapy
following further clinical investigation. The main limitation of
the present study was the recruitment of all participants from a
single centre. This study may serve as a basis for future
prospective studies. Future clinical investigation should be
conducted prospectively, and include a large number of patients
from various centres, so that results can be generalized.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, et
al: Cancer incidence and mortality worldwide: Sources, methods and
major patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Arndt CA and Crist WM: Common
musculoskeletal tumors of childhood and adolescence. N Engl J Med.
341:342–352. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rodriguez-Galindo C, Baez F and Alcasabas
AP: Bone TumorsPediatric Hematology-Oncology in Countries with
Limited Resources. Stefan DC and Rodriguez-Galindo C: Springer; New
York: pp. 323–335. 2014
|
|
4
|
Petrilli AS, de Camargo B, Filho VO, et
al: Brazilian Osteosarcoma Treatment Group Studies III and IV:
Results of the Brazilian Osteosarcoma Treatment Group Studies III
and IV: prognostic factors and impact on survival. J Clin Oncol.
24:1161–1168. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bielack SS, Kempf-Bielack B, Delling G, et
al: Prognostic factors in high-grade osteosarcoma of the
extremities or trunk: an analysis of 1,702 patients treated on
neoadjuvant cooperative osteosarcoma study group protocols. J Clin
Oncol. 20:776–790. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen Y, Han XZ, Wang W, Zhao RT and Li X:
Withaferin A inhibits osteosarcoma cells through inactivation of
Notch-1 signalling. Bangladesh J Pharmacol. 9:364–370. 2014.
View Article : Google Scholar
|
|
7
|
Said HM: Hamdard Pharmacopoeia of Eastern
Medicine. 1st. Hamdard Academy; Karachi, Pakistan: 1970
|
|
8
|
Williamson EM: Major herbs of Ayurveda.
Churchill Livingstone; Philadelphia, PA, USA: 2002
|
|
9
|
Choudhary MI, Yousuf S, Nawaz SA, Ahmed S
and Atta-ur-Rahman: Cholinesterase inhibiting withanolides from
Withania somnifera. Chem Pharm Bull (Tokyo). 52:1358–1361. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Choudhary MI, Nawaz SA, ul-Haq Z, Lodhi
MA, et al: Withanolides, a new class of natural cholinesterase
inhibitors with calcium antagonistic properties. Biochem Biophys
Res Commun. 334:276–287. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jayaprakasam B and Nair MG:
Cyclooxygenase-2 enzyme inhibitory withanolides from Withania
somnifera leaves. Tetrahedron. 59:841–849. 2003. View Article : Google Scholar
|
|
12
|
Owais M, Sharad KS, Shehbaz A and
Saleemuddin M: Antibacterial efficacy of Withania somnifera
(ashwagandha) an indigenous medicinal plant against experimental
murine salmonellosis. Phytomedicine. 12:229–235. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kiasalari Z, Khalili M and Aghaei M:
Effect of Withania somnifera on levels of sex hormones in the
diabetic male rats. Iran J Reprod Med. 7:163–168. 2009.
|
|
14
|
Maurya R, Akanksha, Jayendra, Singh AB and
Srivastava AK: Coagulanolide, a withanolide from Withania coagulans
fruits and antihyperglycemic activity. Bioorg Med Chem Lett.
18:6534–6537. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ali M, Shuaib M and Ansari SH:
Withanolides from the stem bark of Withania somnifera.
Phytochemistry. 44:1163–1168. 1997. View Article : Google Scholar
|
|
16
|
Jayaprakasam B, Zhang Y, Seeram NP and
Nair MG: Growth inhibition of human tumor cell lines by
withanolides from Withania somnifera leaves. Life Sci. 74:125–132.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Subbaraju GV, Vanisree M, Rao CV, et al:
Ashwagandhanolide, a bioactive dimeric thiowithanolide isolated
from the roots of Withania somnifera. J Nat Prod. 69:1790–1792.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ichikawa H, Takada Y, Shishodia S,
Jayaprakasam B, Nair MG and Aggarwal BB: Withanolides potentiate
apoptosis, inhibit invasion and abolish osteoclastogenesis through
suppression of nuclear factor-kappaB (NF-kappaB) activation and
NF-kappaB-regulated gene expression. Mol Cancer Ther. 5:1434–1445.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Srinivasan S, Ranga RS, Burikhanov R, Han
S and Chendil D: Par-4-dependent apoptosis by the dietary compound
withaferin A in prostate cancer cells. Cancer Res. 67:246–253.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang H, Shi G and Dou QP: The tumor
proteasome is a primary target for the natural anticancer compound
Withaferin A isolated from ʻIndian winter cherryʼ. Mol Pharmacol.
71:426–437. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Stan SD, Hahm E-R, Warin R and Singh SV:
Withaferin A causes FOXO3a- and Bim-dependent apoptosis and
inhibits growth of human breast cancer cells in vivo. Cancer Res.
68:7661–7669. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Malik F, Kumar A, Bhushan S, et al:
Reactive oxygen species generation and mitochondrial dysfunction in
the apoptotic cell death of human myeloid leukemia HL-60 cells by a
dietary compound withaferin A with concomitant protection by
N-acetyl cysteine. Apoptosis. 12:2115–2133. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Devi PU, Kamath R and Rao BS:
Radiosensitization of a mouse melanoma by withaferin A: in vivo
studies. Indian J Exp Biol. 38:432–437. 2000.PubMed/NCBI
|
|
24
|
Kaileh M, Vandin Berghe W, Heyerick A, et
al: Withaferin A strongly elicits IkappaB kinase β
hyperphosphorylation concomitant with potent inhibition of its
kinase activity. J Biol Chem. 282:4253–4264. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bargagna-Mohan P, Hamza A, Kim YE, et al:
The tumor inhibitor and antiangiogenic agent withaferin A targets
the intermediate filament protein vimentin. Chem Biol. 14:623–634.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Falsey RR, Marron MT, Gunaherath GM, et
al: Actin microfilament aggregation induced by withaferin A is
mediated by annexin II. Nat Chem Biol. 2:33–38. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Emanuel EJ, Wendler D, Killen J and Grady
C: What makes clinical research in developing countries ethical?
The benchmarks of ethical research. J Infect Dis. 189:930–937.
2004. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lowry OH, Rosebrough NJ, Farr AL and
Randall RJ: Protein measurement with the Folin phenol reagent. J
Biol Chem. 193:265–275. 1951.PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kuroyanagi M, Shibata K and Umehara K:
Cell differentiation inducing steroids from Withania somnifera L.
(Dun). Chem Pharm Bull (Tokyo). 47:1646–1649. 1999. View Article : Google Scholar
|
|
31
|
Mirjalili MH, Moyano E, Bonfill M, Cusido
RM and Palazón J: Steroidal lactones from Withania somnifera, an
ancient plant for novel medicine. Molecules. 14:2373–2393. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Damu AG, Kuo PC, Su CR, et al: Isolation,
structures and structure - cytotoxic activity relationships of
withanolides and physalins from Physalis angulata. J Nat Prod.
70:1146–1152. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee SW, Pan MH, Chen CM and Chen ZT:
Withangulatin I, a new cytotoxic withanolide from Physalis
angulata. Chem Pharm Bull (Tokyo). 56:234–236. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tursunova R, Maslennikova V and Abubakirov
N: Withanolides in the vegetable kingdom. Chem Nat Compd.
13:131–138. 1977. View Article : Google Scholar
|
|
35
|
Liffert R, Hoecker J, Jana CK, et al:
Withanolide A: Synthesis and structural requirements for neurite
outgrowth. Chem Sci. 4:2851–2857. 2013. View Article : Google Scholar
|
|
36
|
Juang J-K, Huang HW, Chen CM and Liu HJ: A
new compound, withangulatin A, promotes type II DNA
topoisomerase-mediated DNA damage. Biochem Biophys Res Commun.
159:1128–1134. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lee WC, Lin KY, Chen CM, Chen ZT, Liu HJ
and Lai YK: Induction of heat-shock response and alterations of
protein phosphorylation by a novel topoisomerase II inhibitor,
withangulatin A, in 9 L rat brain tumor cells. J Cell Physiol.
149:66–76. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zaarur N, Gabai VL, Porco JA Jr,
Calderwood S and Sherman MY: Targeting heat shock response to
sensitize cancer cells to proteasome and Hsp90 inhibitors. Cancer
Res. 66:1783–1791. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jackman M, Lindon C, Nigg EA and Pines J:
Active cyclin B1-Cdk1 first appears on centrosomes in prophase. Nat
Cell Biol. 5:143–148. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mitra J and Enders GH: Cyclin A/Cdk2
complexes regulate activation of Cdk1 and Cdc25 phosphatases in
human cells. Oncogene. 23:3361–3367. 2004. View Article : Google Scholar : PubMed/NCBI
|