Introduction
Increasing numbers of immunocompromised patients has
resulted in an increase in the incidence of fungal infections
worldwide (1–3), of which the Candida species
(Candida spp.) cause numerous clinical diseases associated
with significant mortality and morbidity in healthcare settings
(3–5). Candida albicans is the
predominant pathogen of the Candida spp., which causes
mucosal and invasive infections in humans (3,6).
However, numerous studies have reported that a growing number of
infections can be attributed to non-albicans species
(4,5,7), many of
which exhibit intrinsic resistance or reduced susceptibility to
antifungal agents (2,3,8). Among
them, Candida krusei is an opportunistic fungal pathogen
that exhibits intrinsic resistance to fluconazole (3,9). Despite
the low prevalence of C. krusei infections (accounting for
2–5% of all yeast infections), the intrinsic resistance of the
pathogen to fluconazole means that they are associated with the
highest mortality rate (30–60%) of all yeast (2,3,10). Therefore, the discovery of antifungal
compounds with novel modes of action that are effective against
C. krusei infections, is critical.
Previous studies have indicated that antimicrobial
peptides (AMPs) should be considered leading compounds in the
development of novel antimicrobials (11–14).
AMPs are produced by numerous species, in which they are important
components of the innate immune system (15–17). The
most significant feature of AMPs is their broad-spectrum activity
against microbes, including those exhibiting multi-drug-resistance
(12,14,18). In
addition, unlike traditional antibiotics that inhibit specific
biosynthetic pathways, including cell wall or protein synthesis,
the majority of AMPs conduct their respective functions by
disrupting the membranes of microbes, resulting in the leakage of
cell contents and cell death (12–14).
Furthermore, the development of therapeutic AMPs may overcome the
limitations associated with the current azoles.
In our previous study (19), we identified a novel antifungal
peptide, chromogranin A (CGA)-N46, corresponding to the N-terminal
Pro31-Gln76 sequence of human CGA, which exhibited antifungal
activity against C. albicans. CGA is a soluble protein that
is present in the majority of endocrine cells and neurons (20–23). The
endogenous CGA-derived peptides, including vasostatin-I and
catestatin, have natural defensive roles in the human body
(24–26), and have been shown to possess
antifungal activity; for instance, vasostatin-I (CGA1-76), an
N-terminal fragment of CGA, was able to kill numerous fungal and
yeast cells in the micromolar concentration range (23). In addition, Lugardon et al
(27) synthesized numerous derived
CGA N-terminal fragments, and demonstrated potent antifungal
activity for the shortest peptide, which corresponded to the
sequence Arg47-Leu66 and was named chromofungin. Subsequently, it
was proposed that destabilization of the fungal cell wall and
plasma membrane, alongside intracellular inhibition of
calmodulin-dependent enzymes, may be the underlying mechanism by
which vasostatin-I and chromofungin inhibit fungal growth (27).
In a related study, vasostatin-I engaged in
electrostatic, as well as hydrophobic, interactions with the
membrane phospholipids of fungi under physiological conditions, and
was demonstrated to enhance the fluidity of saturated species of
phosphatidylserine (28), suggesting
numerous mechanisms by which vasostatin-I may inhibit fungal
growth. Concordantly, numerous AMPs have been shown to exert their
effects on various fungal targets (29,30).
The authors of the present study hypothesized that
the underlying antifungal mechanism of the CGA N-terminal fragment
may not be limited to mechanisms previously demonstrated for other
AMPs. In our previous study (31),
we expressed numerous recombinant derived peptides of the CGA
N-terminus in order to elucidate the underlying antifungal
mechanism of the CGA N-terminal fragment. The results suggested
that the short CGA-N46 peptide, corresponding to the sequence
Pro31-Gln76, possessed antifungal activity against numerous
Candida spp., of which C. krusei was the most
sensitive strain. The secondary structure of CGA-N46 was predicted
to be an α-helix by the bioinformatic software ScanProsite
(http://www.expasy.org/tools/scanprosite), and its
isoelectric point was 7.38. As a hydrophilic protein, it had no
transmembrane domain. In the present study, the mechanisms of
action of CGA-N46 AMP against C. krusei were studied.
Materials and methods
Reagents
Triton X-100, and the fluorescent stains propidium
iodide (PI), 2′,7′-dichlorofluorescin diacetate (DCFH-DA) and
Rhodamine-123 (Rh-123) were purchased from Sigma-Aldrich (St.
Louis, MO, USA). All of the other reagents were of the highest
purity available from commercial sources, and all of the solvents
were of high-performance liquid chromatography grade. The water
used for all experiments was supplied by a Milli-Q® Water
Purification system (Merck Millipore, Beijing, China). Candida
krusei ATCC 6258 was supplied by the Chinese Academy of Medical
Sciences and Peking Union Medical College (Beijing, China). The
yeast cells were grown overnight in liquid Sabouraud medium (SD;
Difco, BD Biosciences, San Jose, CA, USA), at 30°C with
agitation.
Antifungal assay
The minimum inhibitory concentrations (MICs) of
CGA-N46 against five different Candida strains (C.
glabrata, C. parapsilosis, C. krusei, C.
tropicalis, and C. albicans) were determined using the
Clinical and Laboratory Standards Institute M-27A3 methodology
(32,33). All strains were obtained from
American Type Culture Collection (Manassas, VA, USA). CGA-N46 was
prepared and stored in our laboratory, then serially diluted in 20
mM phosphate-buffered saline (PBS; pH 6.0) to a final concentration
range of 0.62 µM-3.2 mM. C. krusei cultures that had not
been treated with CGA-N46, or incubated in SD, were employed as
positive and negative controls, respectively. Each experiment was
repeated three times.
Transmission electron microscopy
(TEM)
The effects of CGA-N46 on C. krusei
morphology and internal organelles were investigated using TEM.
Briefly, mid-log phase C. krusei cell pellets were washed
with 20 mM PBS (pH 6.0), and resuspended to a concentration of
106 CFU/ml in SD. Subsequently, the cell suspension was
incubated with CGA-N46, at a concentration of 0.8 mM at 30°C for 3
h, followed by washing with PBS. Fungal pellets were fixed in 500
µl 2.5% glutaraldehyde in PBS at 4°C overnight, and then further
fixed with 1 ml 4% potassium permanganate for 30 min at room
temperature. After being washed and re-suspended in 1 ml saturated
uranyl acetate for 30 min at room temperature, the samples were
dehydrated in ascending concentrations of acetone 2 times per
concentration for 5 min each (30, 50, 70 and 95%), and further
dehydrated in absolute acetone 5 times for 15 min each.
Subsequently, the pellets were gently stirred in a 1:1 mixture of
absolute acetone and final Spurr resin (Polysciences, Inc.,
Warrington, PA, USA) for 1 h at room temperature. Then, the pellets
were transferred to a 1:3 mixture of absolute acetone and Spurr
resin for 3 h, and subsequently to Spurr resin alone overnight.
Specimens were subsequently embedded in gelatin capsules and heated
at 70°C for 9 h to polymerize. Ultra-thin sections, obtained using
an Ultracut E ultramicrotome (Reichert-Jung, Inc., Austria), were
stained with uranyl acetate, followed by staining with lead
citrate. The specimens were observed using a transmission electron
microscope (Hitachi H-7650; Hitachi, Ltd., Tokyo, Japan). C.
krusei cells that had not undergone CGA-N46 treatment were used
as a control.
Langmuir-Blodgett experiment
To determine the effects of CGA-N46 on the stability
of the phospholipid monolayer of fungal cells, the variability of
the surface pressure (π) versus the mean molecular area (A)
isotherms was investigated using a modified Langmuir Blodgett
procedure (34). Briefly, CGA-N46
was dissolved in double distilled H2O to 0, 0.2, 0.8 and
1.6 mM solutions, which were used as the sub-phase. Phospholipids
in ethyl ether (40 µl of 0.32 mg/ml) were spread on the sub-phase
surface with a Hamilton microliter syringe, and were left standing
for 15 min to allow complete evaporation of the solvent.
Subsequently, the floating monolayer was compressed continuously
with a linear movement of two barriers at 2 mm/min. The π-A
isotherms were measured on a KSV Minitrough Langmuir-Blodgett
system (KSV Instruments, Helsinki, Finland), operated using a
Wilhelmy platinum plate with a dynamic surface pressure range of
0–40 mN/m, and a resolution of 4 µN/m. The sub-phase temperature
was maintained within 20.0 ± 0.5°C using a thermostatic bath. The
trough and barriers were thoroughly cleaned using 95% ethanol and
hot water between each isotherm, and every π-A isotherm was
repeated ≥3 times, in order to ensure reliable results. Double
distilled H2O was used as the control.
Permeability of the outer
membrane
Membrane permeability was determined by the uptake
of PI, a high affinity nuclear stain that penetrates compromised
cell membranes and fluoresces upon binding to nucleic acids, using
the method outlined in Lv et al (14), with minor alterations. Briefly, an
overnight culture of C. krusei in SD broth was washed once
and resuspended to 106 CFU/ml in SD broth. CGA-N46 (0,
0.4 or 0.8 mM) was added to the C. krusei cells, and
incubated at 28°C for 3 h. Subsequently, the cells were washed in
PBS and resuspended in SD, after which they were incubated with 50
µg/ml PI in the dark at 37°C for 10 min. The fluorescence images
were observed using a fluorescent microscope (Eclipse TS100; Nikon
Corporation, Tokyo, Japan), with an excitation wavelength of 488
nm. C. krusei cells that had not undergone treatment with
CGA-N46, or were treated with 0.3% Triton-X100, were used as
negative and positive controls, respectively.
Measurement of levels of reactive
oxygen species (ROS)
The levels of ROS were determined using the
fluorescent dye DCFH-DA as a reference (35). Intracellular
H2O2 or low molecular weight peroxides are
able to oxidize DCFH-DA to the highly fluorescent compound
dichlorofluorescein (DCF). Briefly, mid-log phase C. krusei
cells were prepared at a density of 107 CFU/ml and
treated with CGA-N46 (0, 0.2, 0.4 or 0.8 mM) for 3 h at 30°C.
Subsequently, cells were washed with PBS and resuspended in SD,
after which DCFH-DA (final concentration, 10 µM) was added to the
cell suspensions in the dark for 20 min at room temperature. DCF
fluorescence was measured using laser scanning confocal microscopy
(LSM 710; Zeiss, Oberkochen, Germany), at an excitation wavelength
of 488 nm and an emission wavelength of 535 nm. C. krusei
cells that had not undergone treatment with CGA-N46, were used as a
control.
Measurement of the mitochondrial
membrane potential
The mitochondrial membrane potential was assessed
using a fluorescent probe, Rh-123. Briefly, mid-log phase C.
krusei cells at a density of 107 CFU/ml in 20 mM PBS
(pH 6.0), were treated with CGA-N46 (0, 0.2, 0.4 or 0.8 mM) for 3 h
at 30°C. After cells were washed and resuspended in SD, Rh-123
(final concentration, 10 µg/ml) was added to the cell suspensions
in the dark for 30 min at room temperature. The fluorescence images
were observed using a fluorescent microscope (Eclipse TS100, Nikon
Corporation), at an excitation wavelength of 507 nm and an emission
wavelength of 529 nm.
Analysis of in vitro DNA
synthesis
Polymerase chain reaction (PCR) assays were used to
evaluate the effects of CGA-N46 on DNA synthesis in vitro.
PCR was conducted according to the method outlined in Sambrook
et al (36), using reagents
purchases from (Sigma-Aldrich). Briefly, 3 µl 10X PCR buffer was
mixed with 2.4 µl 25 mM MgCl2, 0.6 µl 10 mM
deoxyribonucleoside triphosphates (dNTPs) mixture, 1.5 µl 20 µM
upstream primer, 1.5 µl 20 µM downstream primer of the desired gene
cga-N46, and 0.3 µl PCR template (2 ng/µl plasmid pSVTQ, which bore
the cga-N46 gene). Primers were synthesized by Sangon Biotech Co.,
Ltd. (Shanghai, China), and their sequences were as follows:
Upstream, 5-AA CCC ATG CCT GTC AGC AAC-3 and downstream, 5- ATG TGC
CCT CTC-3. In order to test the effects of CGA-N46 on the
Taq DNA polymerase, 0.15 µl Taq DNA polymerase (5
U/µl) was treated with 21 µl 0.4 mM CGA-N46 in 20 mM PBS (pH 6.0)
for 30 min at 37°C, prior to addition into the PCR mixture. The
same treatment of the Taq DNA polymerase in the absence of
CGA-N46 was used as a positive control, and 21 µl 20 mM PBS (pH
6.0) without Taq DNA Polymerase was used as the negative
control. A final control was the same reactions without the
incubation at 37°C for 30 min. PCR was conducted in an S1000
thermal cycler (Bio-Rad Laboratories, Inc., Hercules, CA, USA)
using the following parameters: 95°C for 5 min, followed by 30
cycles of 95°C denaturation for 1 min, 58°C annealing for 30 sec,
72°C extension for 1 min, and a final extension at 72°C for 10 min.
PCR products (10 µl) were separated using 1% agarose gel
electrophoresis.
Flow cytometry
Mid-log phase C. krusei cells
(1×106 CFU/ml), incubated with 0.8 mM CGA-N46 in 20 mM
PBS (pH 6.0), were incubated for 12, 24 and 48 h at 30°C, with
agitation. Subsequently, the cells were collected, washed,
resuspended and fixed in 70% ice cold ethanol for 24 h. The fixed
cells were then incubated with the PI solution (50 µg/ml PI, 100
µg/ml RNase (Sangon Biotech Co., Ltd.), and 0.2% Triton X-100
supplemented with PBS) for 30 min in the dark. Cell cycle
distribution analysis was performed using the Cell Quest Flow
Cytometer (FACScalibur; BD Biosciences), and the results were
analyzed using the modFit LT™ software, version 3.1 (Verity
Software House, Topsham, ME, USA).
Statistical analysis
Experimental data were analyzed using the PASW
Statistics software, version 18 (SPSS, Inc., Chicago, IL, USA) to
perform one-way analysis of variance followed by Least Significant
Difference and Duncans tests. The results are reported as the mean
± standard error of the mean. Differences between the treatment and
control groups were considered to be statistically significant at
P<0.05, and highly significant at P<0.01.
Results
Anti-candidal activity
The MIC of CGA-N46 against numerous Candida spp.
(Table I) was used as an indicator
of the peptide's anti-candidal activity. CGA-N46 was active against
all of the yeasts tested (MICs, 0.1–0.8 mM), with C. krusei
exhibiting the greatest sensitivity (MIC, 0.1 mM).
 | Table I.MIC of CGA-N46 against Candida
species. |
Table I.
MIC of CGA-N46 against Candida
species.
| Strains | MICs (mM) |
|---|
| Candida
glabrata | 0.8 |
| Candida
parapsilosis | 0.8 |
| Candida
krusei | 0.1 |
| Candida
tropicalis | 0.2 |
| Candida
albicans | 0.2 |
Effects of CGA-N46 on cell
morphology
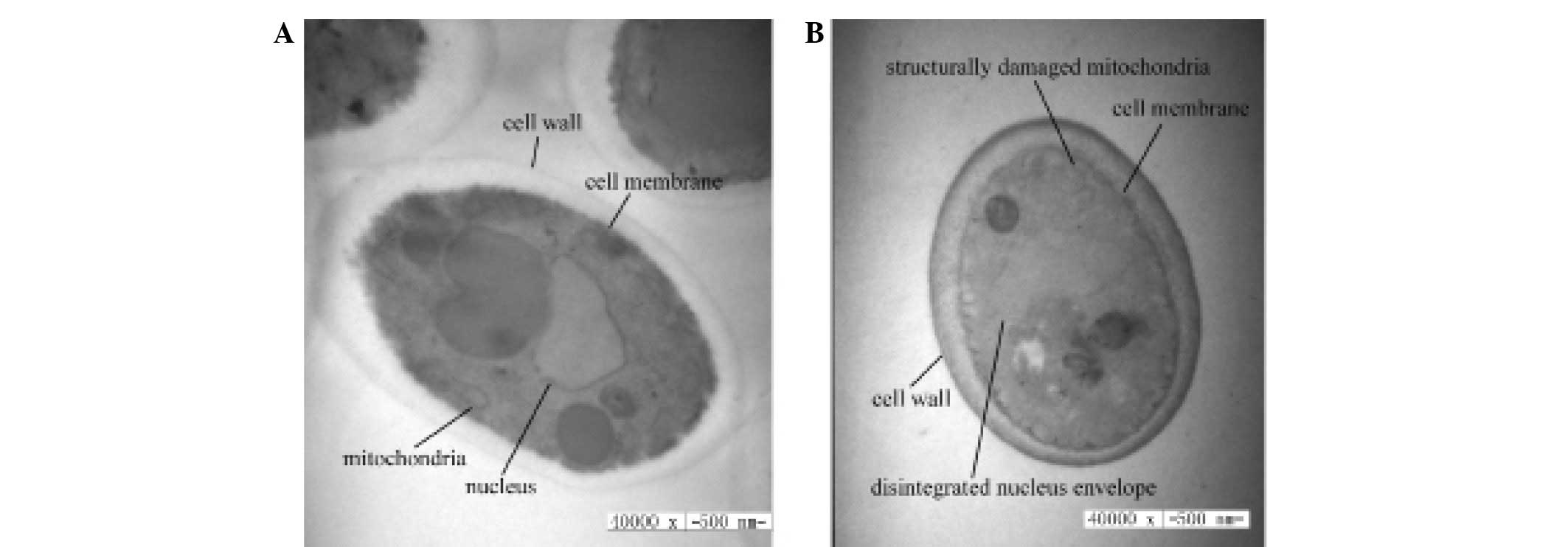
The effects of CGA-N46 on the cellular morphology
and organelles of C. krusei were investigated using TEM. As
compared with the control (Fig. 1A),
the cell wall and outer membrane were clear in CGA-N46-treated
C. krusei cells, but significant cytoplasmic vacuolization
was observed. Furthermore, mitochondrial structural damage was
observed, and the integrity of the nuclear envelope was disrupted,
as demonstrated by the visible pores (Fig. 1B). These results suggest that CGA-N46
may penetrate the cell membrane and subsequently interact with
cellular organelles.
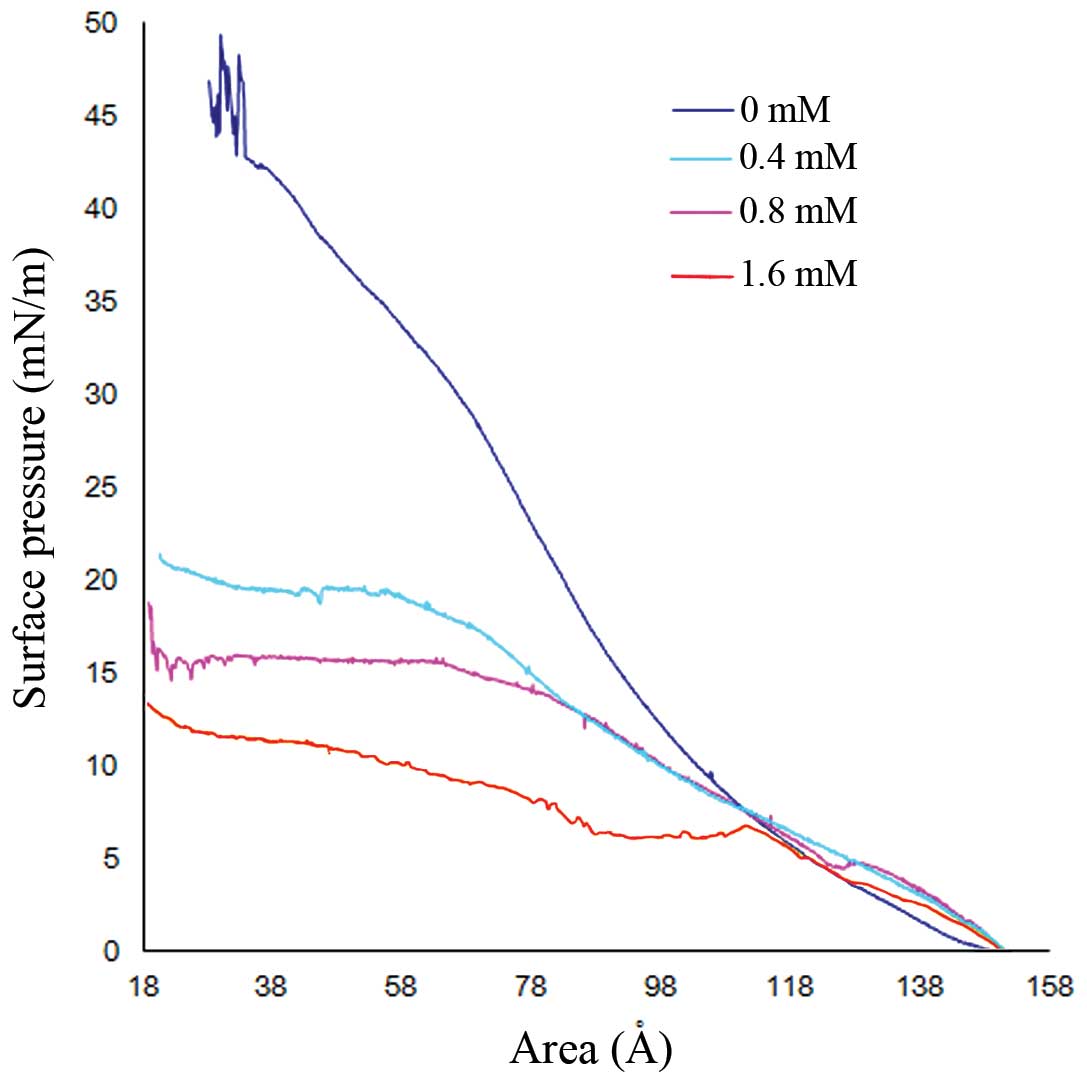
Effects of CGA-N46 on the stability of
the phospholipid monolayer
The effects of CGA-N46 on the stability of the cell
membrane were demonstrated by the variability of the π-A isotherms
of the phospholipid monolayer (Fig.
2). The limiting area of the phospholipid monolayer was
estimated by extrapolating the straight portion of the π-A isotherm
to zero surface pressure. As compared with the control,
CGA-N46-treatment (0, 0.4, 0.8 or 1.6 mM) was associated with
marked alterations to the shape of the π-A isotherms. As the
concentration of CGA-N46 in the sub-phase increased, the mean
molecular areas increased, which suggests that CGA-N46 was able to
lower the density of phospholipids in the lipid monolayer. This
indicates that CGA-N46 treatment may promote disorder of membrane
phospholipids, leading to a reduction in membrane stability.
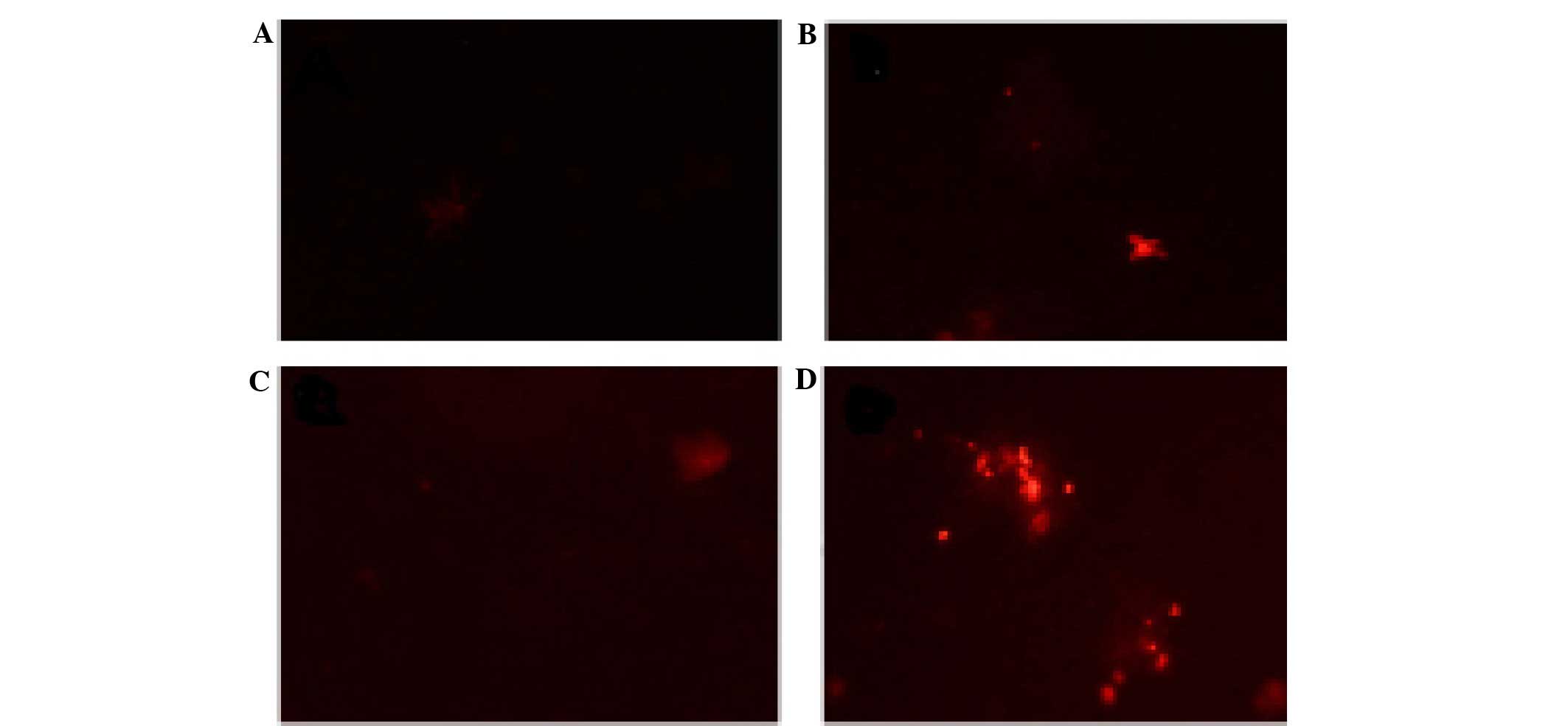
Effects of CGA-N46 on the outer
membrane permeability of C. krusei cells
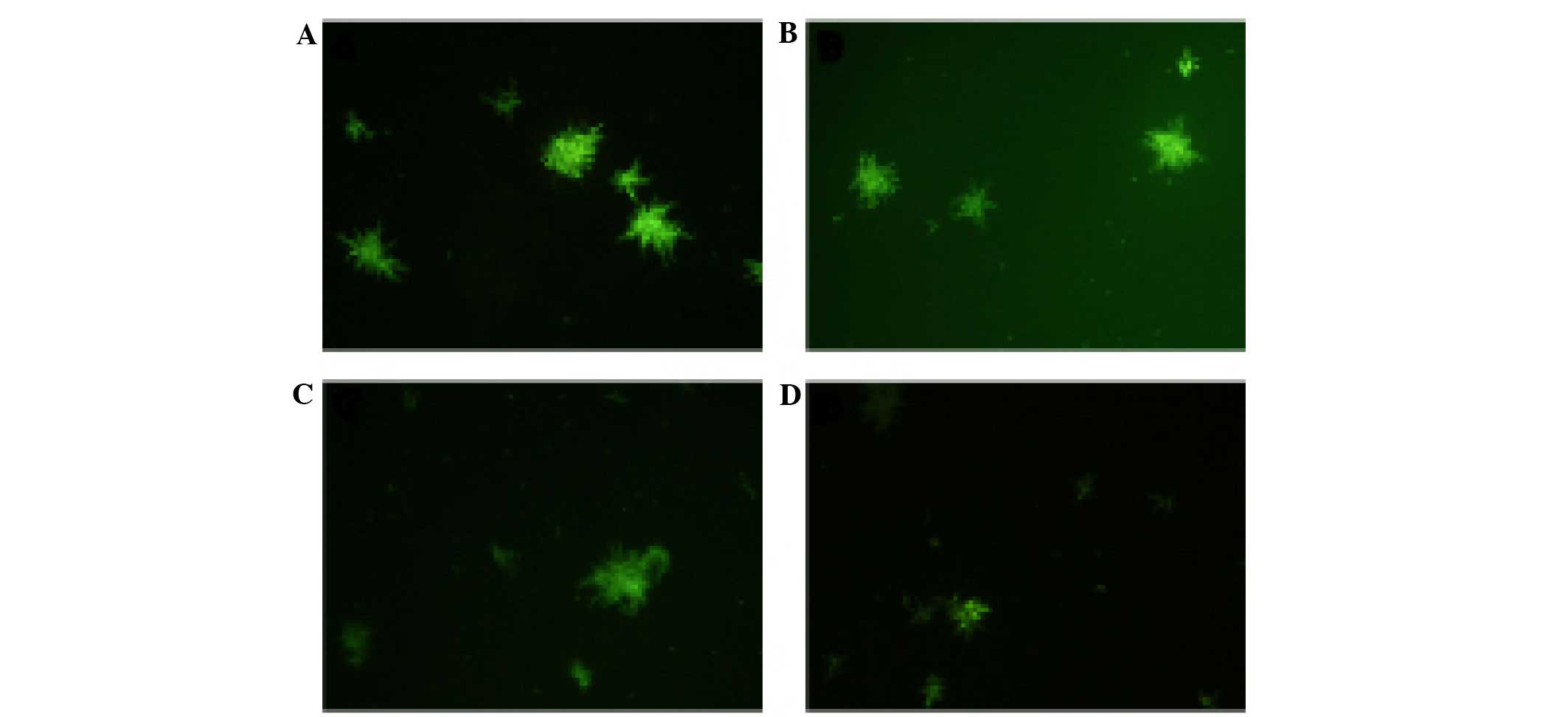
The PI uptake assay was used to detect the effects
of CGA-N46 on the permeability of the outer membrane of C.
krusei cells. PI is a small hydrophobic molecule that
fluoresces weakly in aqueous solution but strongly when it enters a
hydrophobic environment, including a biomembrane; therefore, PI is
widely used to detect the disruption of the outer membrane of
microbes. The PI fluorescent probe was unable to enter the outer
membrane of C. krusei cells following treatment with CGA-N46
(0.4 or 0.8 mM; Fig. 3). This result
suggests that CGA-N46 does not affect the permeability of the outer
membrane of C. krusei cells.
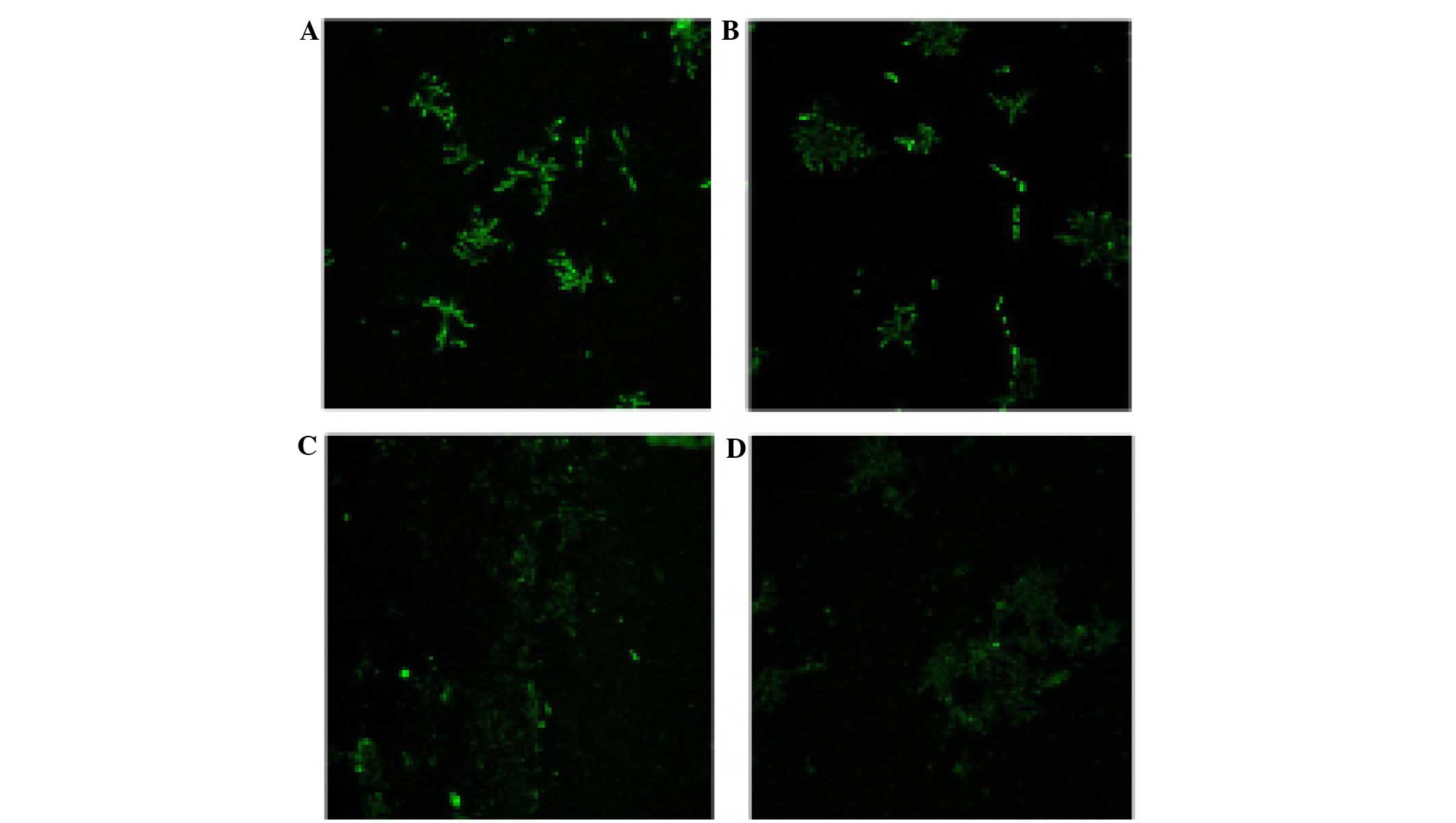
Effects of CGA-N46 on ROS levels
To investigate the effects of CGA-N46 on
intracellular ROS levels, the C. krusei cells were treated
with CGA-N46 (0, 0.2, 0.4 or 0.8 mM) for 3 h at 30°C. Intracellular
ROS oxidize DCFH-DA to the highly fluorescent DCF. CGA-N46
treatment of C. krusei cells decreased DCF fluorescence in a
concentration-dependent manner (Fig.
4). This result suggests that CGA-N46 is able to reduce the
levels of intracellular ROS.
Effects of CGA-N46 on mitochondrial
membrane potential
To investigate the effects of CGA-N46 on the
mitochondrial membrane potential of C. krusei cells, the
cells were treated with CGA-N46 (0, 0.2, 0.4 or 0.8 mM) for 3 h at
30°C. The mitochondrial membrane potential was analyzed using the
fluorescent probe Rh-123, and fluorescence microscopy. The Rh-123
fluorescence emitted by CGA-N46-treated C. krusei cells
decreased in a concentration-dependent manner (Fig. 5). This result suggests that CGA-N46
is able to reduce the mitochondrial membrane potential of C.
krusei cells.
Effects of CGA-N46 on DNA
synthesis
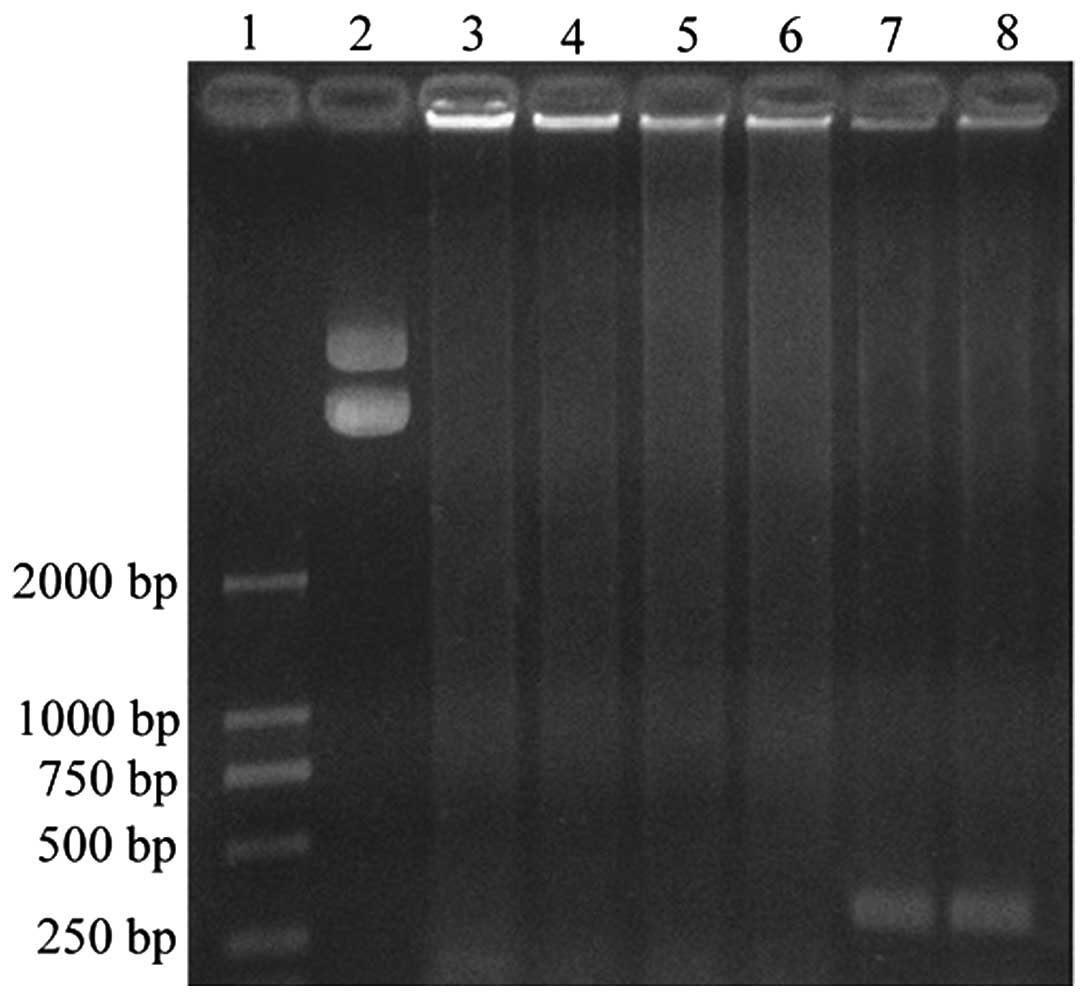
In order to determine whether CGA-N46 inhibits DNA
synthesis, the effects of CGA-N46 on the Taq polymerase in a
PCR assay were analyzed. CGA-N46 was incubated with Taq DNA
polymerase for 30 min at 37°C prior to PCR, and this was compared
to a reaction without prior incubation. The agarose gel
electrophoresis separation of the PCR products suggested that, as
expected, there was no PCR product when Taq DNA polymerase
was absent from the PCR system (Fig.
6, lanes 5 and 6). Furthermore, PCR product was observed when
Taq DNA polymerase was included in the reaction mixture
(Fig. 6, lanes 7 and 8); however,
there was no PCR product detected when Taq DNA polymerase
was treated with CGA-N46 (Fig. 6,
lanes 3 and 4). The results suggest that CGA-N46 inactivates the
Taq DNA polymerase, thereby inhibiting DNA synthesis.
Effects of CGA-N46 on cell cycle
distribution
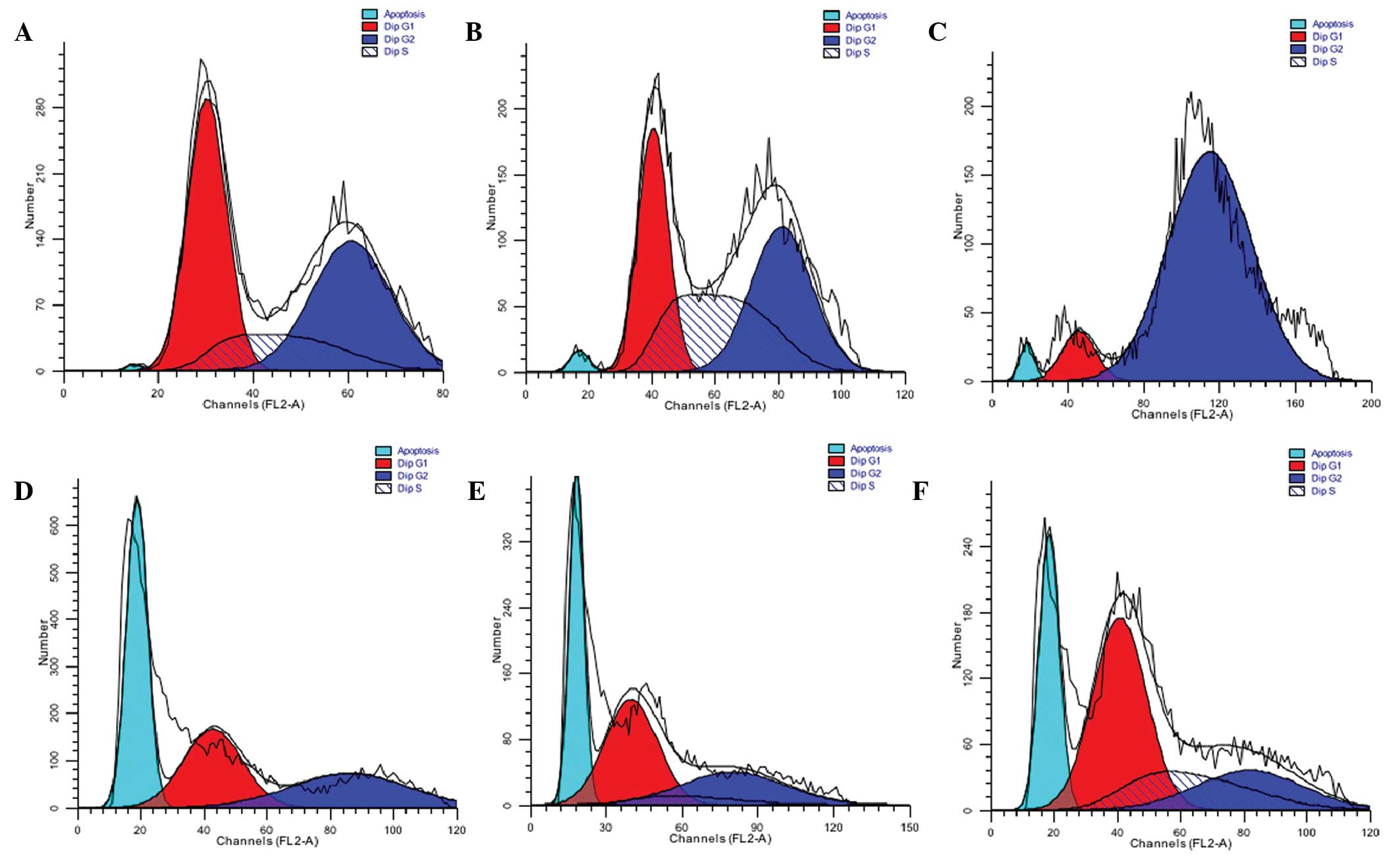
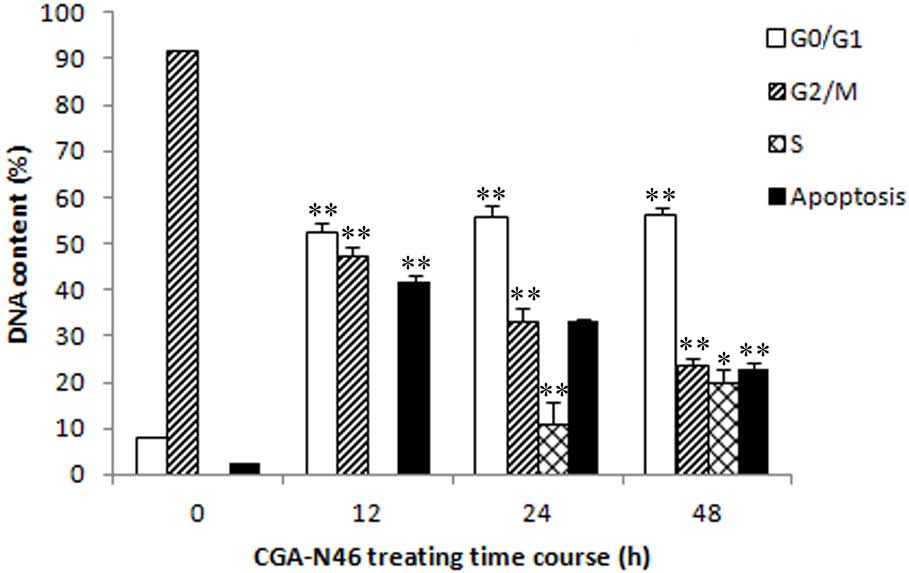
In order to investigate the effects of CGA-N46 on
DNA synthesis in vivo, cell cycle and DNA distribution
analyses were performed for C. krusei cells treated with 0.8
mM CGA-N46 for 12, 24 and 48 h. Following treatment with CGA-N46,
the proportion of C. krusei cells in the
G0/G1 phase remained constant (Fig. 7), which indicated that CGA-N46
arrested cell proliferation at the G0/G1
phase by blocking DNA synthesis. This result is consistent with the
in vitro PCR results. In addition, the DNA content in the
apoptotic phase significantly increased following treatment with
CGA-N46 (cells in the sub-G0/G1 phase are
considered apoptotic) (Fig. 8).
However, after 12 h, the DNA content of apoptotic cells decreased,
and this was accompanied by an increase in the DNA content of S
phase and G2/M phase cells. This result indicates that
the antagonistic effects of CGA-N46 may be lost by 24 h.
Discussion
AMPs are widely distributed in nature and have
important roles in the host defense against microbial pathogens.
Numerous organisms use these peptides as an innate defense
mechanism that protects against invading microorganisms (17,37–39); the
high cationic charge and strong amphipathic nature of AMPs enables
them to bind to the anionic microbial cell membrane and disrupt the
membrane bilayer via the formation of pores or channels (39–41),
which in turn leads to intracellular potassium ion leakage and cell
death (41). As compared with the
classical pore-forming peptides, CGA-N46 is a weak alkaline
α-helical peptide and does not kill Candida spp. via a
pore-forming mechanism. Conversely, in the present study, CGA-N46
was transported into the cytoplasm of Candida spp. via a
mechanism that disrupted the arrangement of phospholipids in the
cell membrane without disturbing its integrity.
Lugardon et al (28) proposed that N-terminal derived
fragments of CGA may promote destabilization of the fungal plasma
membrane. In the present study, CGA-N46 was able to promote the
destabilization of a phospholipid monolayer. Numerous antibacterial
peptides have been demonstrated to pass through the cell membrane
of microbial pathogens, without damaging the membrane integrity;
instead they interact with intracellular organelles and
macromolecules, and influence intracellular nucleic acid synthesis
and repair, protein synthesis, cell wall and membrane synthesis and
numerous physiological activities (42,43). In
a previous study (40), the human
salivary antimicrobial peptide Muc7 damaged the fungal
mitochondrial membrane, leading to vacuolation of the mitochondria
and deformation or degradation of the ridge, thereby resulting in
the cell death. These effects were similarly observed in the C.
krusei cells treated with CGA-N46 in the present study.
Unlike classical AMPs, CGA-N46 destroyed
mitochondrial structure and nuclear envelope integrity, and induced
a reduction in the levels of intracellular ROS. An abnormal
cellular ROS balance has previously been associated with
mitochondrial structural injury (35), and the results of the present study
corroborated this. In addition, treatment with CGA-N46 was
associated with a decreased mitochondrial membrane potential of
C. krusei cells. The present study hypothesized that CGA-N46
may disrupt the mitochondrial electron transfer chain by reducing
the mitochondrial membrane potential, following the reduction of
O2− and H2O2
generation. The results of the present study align with those
reported in Bensassi et al (44), and were also demonstrated in the
inhibitory effect of terbinafine, which reduced the ability of
C. albicans to generate ROS (45).
Numerous antibacterial peptides are able to inhibit
replication and transcription by binding to DNA, and this is one
example of a mechanism by which AMPs exert their antibacterial
activity (46–48). In the present study, a gel
retardation assay was performed to determine whether CGA-N46
associated with C. krusei DNA. The agarose gel
electrophoresis results demonstrated that the migration of DNA was
not retarded following CGA-N46 treatment, even at a weight ratio of
1:80 of CGA-N46 to C. krusei chromosomal DNA (data not
shown). This suggested that CGA-N46 did not associate with C.
krusei DNA, which may have been due to the near-neutral charge
of CGA-N46. The results of the PCR and cell cycle distribution
analyses suggested that CGA-N46 was able to inhibit the synthesis
of DNA in vitro and in vivo, and the results of the
in vitro PCR indicated that CGA-N46 may kill Candida
species by inactivating DNA polymerase.
In conclusion, AMPs have been demonstrated to kill
microbes via numerous mechanisms that exert effects on various
targets. Concordantly, the CGA-N46 peptide analyzed in the present
study exerted its anti-candidal effects on numerous intracellular
targets of C. krusei cells. Therefore, CGA-N46 may be
considered a promising candidate for the treatment of patients with
candidiasis.
Acknowledgements
The authors of the present study would like to thank
Mr. Liang Wang, College of Biological Engineering, Henan University
of Technology for the flow cytometry analyses. This study was
supported by grants from the National Natural Science Foundation of
China (grant no. 31071922) and the Henan University of Technology
(grant no. 11JCYJ10).
References
|
1
|
Pfaller MA and Diekema DJ: Epidemiology of
invasive mycoses in North America. Crit Rev Microbiol. 36:1–53.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bassetti M, Taramasso L, Nicco E, Molinari
MP, Mussap M and Viscoli C: Epidemiology, species distribution,
antifungal susceptibility and outcome of nosocomial candidemia in a
tertiary care hospital in Italy. PLoS One. 6:e241982011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Scorzoni L, de Lucas MP, Mesa-Arango AC,
Fusco-Almeida AM, Lozano E, Cuenca-Estrella M, Mendes-Giannini MJ
and Zaragoza O: Antifungal efficacy during Candida krusei
infection in non-conventional models correlates with the yeast in
vitro susceptibility profile. PLoS One. 8:e600472013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shorr AF, Gupta V, Sun X, Johannes RS,
Spalding J and Tabak YP: Burden of early-onset candidemia: Analysis
of culture-positive bloodstream infections from a large U.S.
database. Crit Care Med. 37:2519–2526. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Colombo AL, Tobón A, Restrepo A,
Queiroz-Telles F and Nucci M: Epidemiology of endemic systemic
fungal infections in Latin America. Med Mycol. 49:785–798.
2011.PubMed/NCBI
|
|
6
|
Pushpanathan M, Rajendhran J, Jayashree S,
Sundarakrishnan B, Jayachandran S and Gunasekaran P: Direct cell
penetration of the antifungal peptide, MMGP1, in Candida
albicans. J Pept Sci. 18:657–660. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Arendrup MC: Epidemiology of invasive
candidiasis. Curr Opin Crit Care. 16:445–452. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Leroy O, Gangneux JP, Montravers P, Mira
JP, Gouin F, Sollet JP, Carlet J, Reynes J, Rosenheim M, Regnier B
and Lortholary O: AmarCand Study Group: Epidemiology, management,
and risk factors for death of invasive Candida infections in
critical care: A multicenter, prospective, observational study in
France (2005–2006). Crit Care Med. 37:1612–1618. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Muñoz P, Sánchez-Somolinos M, Alcalá L,
Rodríguez-Créixems M, Peláez T and Bouza E: Candida krusei
fungaemia: Antifungal susceptibility and clinical presentation of
an uncommon entity during 15 years in a single general hospital. J
Antimicrob Chemother. 55:188–193. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Abbas J, Bodey GP, Hanna HA, Mardani M,
Girgawy E, Abi-Said D, Whimbey E, Hachem R and Raad I: Candida
krusei fungemia. An escalating serious infection in
immunocompromised patients. Arch Intern Med. 160:2659–2664. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bowdish DM, Davidson DJ and Hancock RE: A
re-evaluation of the role of host defence peptides in mammalian
immunity. Curr Protein Pept Sci. 6:35–51. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang J, Hao D and Chen Y, Xu Y, Tan J,
Huang Y, Li F and Chen Y: Inhibitory effects and mechanisms of
physiological conditions on the activity of enantiomeric forms of
an α-helical antibacterial peptide against bacteria. Peptides.
32:1488–1495. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Teixeira V, Feio MJ and Bastos M: Role of
lipids in the interaction of antimicrobial peptides with membranes.
Prog Lipid Res. 51:149–177. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lv Y, Wang J, Gao H, Wang Z, Dong N, Ma Q
and Shan A: Antimicrobial properties and membrane-active mechanism
of a potential α-helical antimicrobial derived from cathelicidin
PMAP-36. PLoS One. 9:e863642014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Auvynet C and Rosenstein Y:
Multifunctional host defense peptides: Antimicrobial peptides, the
small yet big players in innate and adaptive immunity. FEBS J.
276:6497–6508. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shang D, Sun Y, Wang C, Wei S, Ma L and
Sun L: Membrane interaction and antibacterial properties of
chensinin-1, an antimicrobial peptide with atypical structural
features from the skin of Rana chensinensis. Appl Microbiol
Biotechnol. 96:1551–1560. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Vila-Farres X, de la Maria C Garcia,
López-Rojas R, Pachón J, Giralt E and Vila J: In vitro activity of
several antimicrobial peptides against colistin-susceptible and
colistin-resistant Acinetobacter baumannii. Clin Microbiol
Infect. 18:383–387. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gopal R, Seo CH, Song PI and Park Y:
Effect of repetitive lysine-tryptophan motifs on the bactericidal
activity of antimicrobial peptides. Amino Acids. 44:645–660. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li R, Zhang T, Luo J, Wang F, Gu Q, Gan J
and Xiao F: Antifungal activity fragments of N domain of
chromogranin A. Zhong Shan Da Xue Xue. 45:64–67. 2006.(In
Chinese).
|
|
20
|
Eiden LE: Is chromogranin a prohormone?
Nature. 325:3011987. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Simon JP and Aunis D: Biochemistry of the
chromogranin A protein family. Biochem J. 262:1–13. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Helle KB: Chromogranins: Universal
proteins in secretory organelles from paramecium to man. Neurochem
Int. 17:165–175. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lugardon K, Raffner R, Goumon Y, Corti A,
Delmas A, Bulet P, Aunis D and Metz-Boutigue MH: Antibacterial and
antifungal activities of vasostatin-1, the N-terminal fragment of
chromogranin A. J Biol Chem. 275:10745–10753. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Radek KA, Lopez-Garcia B, Hupe M, Niesman
IR, Elias PM, Taupenot L, Mahata SK, O'Connor DT and Gallo RL: The
neuroendocrine peptide catestatin is a cutaneous antimicrobial and
induced in the skin after injury. J Invest Dermatol. 128:1525–1534.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Akaddar A, Doderer-Lang C, Marzahn MR,
Delalande F, Mousli M, Helle K, Van Dorsselaer A, Aunis D, Dunn BM,
Metz-Boutigue MH and Candolfi E: Catestatin, an endogenous
chromogranin A-derived peptide, inhibits in vitro growth of
Plasmodium falciparum. Cell Mol Life Sci. 67:1005–1015.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mahata SK, Mahata M, Fung MM and O'Connor
DT: Catestatin: A multifunctional peptide from chromogranin A.
Regul Pept. 162:33–43. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lugardon K, Chasserot-Golaz S, Kieffer
A-E, Maget-Dana R, Nullans G, Kieffer B, Aunis D and Metz-Boutigue
MH: Structural and biological characterization of chromofungin, the
antifungal chromogranin A-(47–66)-derived peptide. J Biol Chem.
276:35875–35882. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Blois A, Holmsen H, Martino G, Corti A,
Metz-Boutigue MH and Helle KB: Interactions of chromogranin
A-derived vasostatins and monolayers of phosphatidylserine,
phosphatidylcholine and phosphatidylethanolamine. Regul Pept.
134:30–37. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zasloff M: Antimicrobial peptides of
multicellular organisms. Nature. 415:389–395. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li Y, Xiang Q, Zhang Q, Huang Y and Su Z:
Overview on the recent study of antimicrobial peptides: Origins,
functions, relative mechanisms and application. Peptides.
37:207–215. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li R, Lu Y, Chen S, Zhang H, Wang B and
Xiong Q: Bioinformatics analysis of animal endogenous peptides
CGA-N46. Dong Wu Yi Xue Jin Zhan. 34:43–46. 2013.(In Chinese).
|
|
32
|
Clinical and Laboratory Standards
Institute: Reference method for broth dilution antifungal
susceptibility testing of yeasts: Approved standard3rd. CLSI
document M27-A3. Clinical and Laboratory Standards Institute.
Wayne, PA, USA: 2008
|
|
33
|
Jurevic RJ, Traboulsi RS, Mukherjee PK,
Salata RA and Ghannoum MA: Oral HIV/AIDS Research Alliance Mycology
Focus group: Identification of gentian violet concentration that
does not stain oral mucosa, possesses anti-candidal activity and is
well tolerated. Eur J Clin Microbiol Infect Dis. 30:629–633. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen Y, Sun R and Wang B: Monolayer
behavior of binary systems of betulinic acid and cardiolipin:
Thermodynamic analyses of Langmuir monolayers and AFM study of
Langmuir-Blodgett monolayers. J Colloid Interface Sci. 353:294–300.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li D, Ma H, Ye Y, Ji C, Tang X, Ouyang D,
Chen J, Li Y and Ma Y: Deoxynivalenol induces apoptosis in mouse
thymic epithelial cells through mitochondria-mediated pathway.
Environ Toxicol Pharmacol. 38:163–171. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sambrook J and Rusell DW: Molecular
Cloning: A Laboratory Mannual. 3rd. Cold Spring Harbor Laboratory
press; New York: 2001
|
|
37
|
van Kan EJ, Demel RA, van der Bent A and
de Kruijff B: The role of the abundant phenylalanines in the mode
of action of the antimicrobial peptide clavanin. Biochim Biophys
Acta. 1615:84–92. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hancock RE and Scott MG: The role of
antimicrobial peptides in animal defenses. Proc Natl Acad Sci USA.
97:8856–8861. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mochon AB and Liu H: The antimicrobial
peptide histatin-5 causes a spatially restricted disruption on the
Candida albicans surface, allowing rapid entry of the
peptide into the cytoplasm. PLoS Pathog. 4:e10001902008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bobek LA and Situ H: MUC7 20-Mer:
Investigation of antimicrobial activity, secondary structure and
possible mechanism of antifungal action. Antimicrob Agents
Chemother. 47:643–652. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bolintineanu D, Hazrati E, Davis HT,
Lehrer RI and Kaznessis YN: Antimicrobial mechanism of pore-forming
protegrin peptides: 100 pores to kill E. coli. Peptides.
31:1–8. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Park CB, Kim HS and Kim SC: Mechanism of
action of the antimicrobial peptide buforin II: Buforin II kills
microorganisms by penetrating the cell membrane and inhibiting
cellular functions. Biochem Biophys Res Commun. 244:253–257. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kochervinskii VV, Yudin SG, Zanaveskina
IS, Arkharova HA, Klechkovskaya VV and Lokshin BV: Structure and
appearance of residual polarization in thin films of polyvinylidene
fluoride prepared via the Langmuir-Blodgett method. Polym Sci Ser
A. 52:40–48. 2010. View Article : Google Scholar
|
|
44
|
Bensassi F, Gallerne C, el Dein OS,
Hajlaoui MR, Bacha H and Lemaire C: Mechanism of Alternariol
monomethyl ether-induced mitochondrial apoptosis in human colon
carcinoma cells. Toxicology. 290:230–240. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sander CS, Hipler UC, Wollina U and Elsner
P: Inhibitory effect of terbinafine on reactive oxygen species
(ROS) generation by Candida albicans. Mycoses. 45:152–155.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Koshlukova SE, Lloyd TL, Araujo MW and
Edgerton M: Salivary histatin 5 induces non-lytic release of ATP
from Candida albicans leading to cell death. J Biol Chem.
274:18872–18879. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Moreno AB, Del Pozo A Martínez and Segundo
B San: Biotechnologically relevant enzymes and proteins. Antifungal
mechanism of the Aspergillus giganteus AFP against the rice
blast fungus Magnaporthe grisea. Appl Microbiol Biotechnol.
72:883–895. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang J, Wu X and Zhang SQ: Antifungal
mechanism of antibacterial peptide ABP-CM4, from Bombyx mori
against Aspergillus niger. Biotechnol Lett. 30:2157–2163.
2008. View Article : Google Scholar : PubMed/NCBI
|