Introduction
It is well known that genetic variation within the
cells initiating a tumor leads to neoplasia; however, the tumor
microenvironment theory suggests that host-derived components also
participate in malignant tumor progression (1). It has previously been demonstrated that
glioma stem progenitor cells have an important role in glioma
tissue remodeling, and that host-derived components, including an
altered vascular system, and glial cells that are transformed by
glioma stem progenitor cells, may contribute substantially to tumor
development (2,3). These results markedly improve the
understanding of tumor-host interactions.
The earliest theory linking host-derived components
with tumor progression was Paget's ‘seed and soil’ hypothesis
(4), which proposed that tumor
cells, or ‘seeds’, randomly disseminate within the blood flow,
although they can only survive in permissive organs (i.e., the
‘soil’).
Fidler (5–7) further developed the ‘seed and soil’
hypothesis, and claimed that the tumor microenvironment is able to
promote tumor growth. Furthermore, Fidler hypothesized that the
biological characteristics of the microenvironments of various
organs are distinct, and that the growth of metastatic cells
depends on their interactions with host cells; however, the
mechanisms underlying these interactions are not well understood,
due to the lack of an appropriate animal model.
Hoffman developed a novel BALB/c nude mouse model
that expressed green fluorescent protein (GFP), and subsequently
established a red fluorescent protein (RFP)/GFP human cancer
xenograft model by transplanting numerous solid tumor cells
expressing RFP into the mice. These xenograft models permitted
in vivo visualization and facilitated the study of
host-tumor interactions in histological sections (8–11). Our
previous study succeeded in cultivating two nude mouse strains,
NC-C57BL6J-EGFP and BALB/c-C57BL6J-EGFP (12), which were subsequently used to study
the efficacy of using dual-color fluorescence tracing in the
investigation of glioma.
The present study compared dual-color fluorescence
tracing with traditional methods in order to analyze tumor
characteristics, including localization, angiogenesis, cellular
fusion, and the tumor microenvironment. The dual-color fluorescence
tracing method is seldom used to study brain tumors.
Materials and methods
Materials
The SU3 human glioma cell line was established in
our laboratory. The C6 rat glioma cell line was obtained from the
Institute of Biochemistry and Cell Biology Shanghai Institutes for
Biological Sciences, Chinese Academy of Sciences (Shanghai, China).
The CM-Dil dye was purchased from Invitrogen Life Technologies
(Carlsbad, CA, USA) and the RFP lentiviral vector was purchased
from Yingweixing Biological Science Technologies Co. (Shanghai,
China). The nude NC-C57BL6J-EGFP and BALB/c-C57BL6J-EGFP mice (age,
6–8 weeks; weight, 22–24 g) were established and reared by our
group in independent ventilation cages. The in vivo FX Pro
Imaging system was purchased from Eastman Kodak (Rochester, NY,
USA), and a fluorescence torch (DFP-1) was purchased from Nightsea
(Lexington, MA, USA). The Eclipse TE2000U fluorescence inverted
microscope was obtained from Nikon Corporation (Tokyo, Japan), and
the BB16 UV CO2 cell incubator was produced by Heraeus
Holding GmbH (Hanau, Germany). The stereotactic apparatus was
obtained from Zhenhua Bioinstrumentation LTD (Huaibei, China), and
the SLY animal cerebral section mold was purchased from Suolinyuan
Technology Company (Beijing, China). A freezing microtome was
purchased from Leica Microsystems GmbH (Wetzlar, Germany).
Mouse-anti-CNP-2′,3′-cAMP-3′-phosphodiesterase (CNPase) monoclonal
antibody, rabbit-anti-Nestin polyclonal antibody, rat-anti-CD68
monoclonal antibody and rabbit-anti-Ki67 monoclonal antibody were
purchased from Abcam (Cambridge, UK). Mouse-anti-glial fibrillary
acidic protein (GFAP) was purchased from BD Biosciences (Oxford,
UK).
Labeling of C6 cells
C6 cells were cultivated in Dulbecco's modified
Eagle's medium (DMEM)/F12, supplemented with 10% fetal calf serum
(Gibco Life Technologies, Carlsbad, CA, USA) in 5%
CO2/95% air at 37°C. Once the cells reached the
logarithmic growth phase, the culture medium was removed, and the
cells were washed twice with phosphate-buffered saline (PBS). Prior
to labeling, CM-Dil was diluted in DMEM/F12 to a final
concentration of 2 µM, and the C6 cells were incubated in this
solution for 10 min at 37°C, and 15 min at 4°C. CM-Dil-stained
cells (C6-CM-Dil) were washed with PBS three times and suspended in
fresh culture medium. The SU3 human glioma cell line was
transfected with the RFP gene using a lentivirus-mediated gene
transfection kit (Genechem, Co., Ltd., Shanghai, China), according
to the manufacturer's instructions.
Establishment of the dual-color
orthotopic model of transplantable xenograft glioma
Transgenic female mice (BALB/c C57BL6J EGFP)
expressing enhanced GFP (eGFP) were established in our laboratory
(13), and cultivated in the
Experiment Animals Center, Soochow University (Suzhou, China). The
mice were used for research when they were aged 6–8 weeks and had a
body weight of ~25 g. All of the mice were bred and maintained in
the Specific Pathogen-Free Animal Care Facility. Following
successful general anesthesia of the mice via intraperitoneal
injection of 10% chloral hydrate (200 mg/kg), a small burr hole (2
mm in diameter) was made 2.5 mm right of the midline and 0.5 mm
anterior to the bregma using a microskull drill, as outlined in
Kaye et al (14). C6-CM-Dil
and SU3-RFP (1×105) cells were injected into the right
caudate nucleus at a depth of 3.5 mm, and a stereotactic apparatus
assisted this procedure. The skull hole was then sealed with bone
wax, and the scalps were sutured.
Dual-color tracing with whole-body in
vivo fluorescence imaging of xenografts
The tumor-bearing mice were anesthetized by chloral
hydrate injection (200 mg/kg) at 3, 5, 7, 9, 11, 13 and 15 days
following tumor inoculation for in vivo fluorescent imaging
using a live imaging system, with excitation and emission
wavelengths of 470 and 535 nm for GFP, and 558 and 583 nm for
RFP.
Generation of serial frozen tumor
sections
Following completion of live imaging, the chests of
the mice were opened under anesthesia, and 1–2 ml of normal saline
was slowly injected into the left ventricle. A small incision was
made on the auricle of the right atrium to allow blood to flow out.
This step was immediately followed by the perfusion of 5–10 ml of
4% paraformaldehyde into the left ventricle. The whole brain was
obtained and fixed in 4% formaldehyde for 7–8 h, following which it
was dehydrated in a 20%, and then a 30%, sucrose solution. The
brain was ready for coronal slicing upon sinking to the bottom of
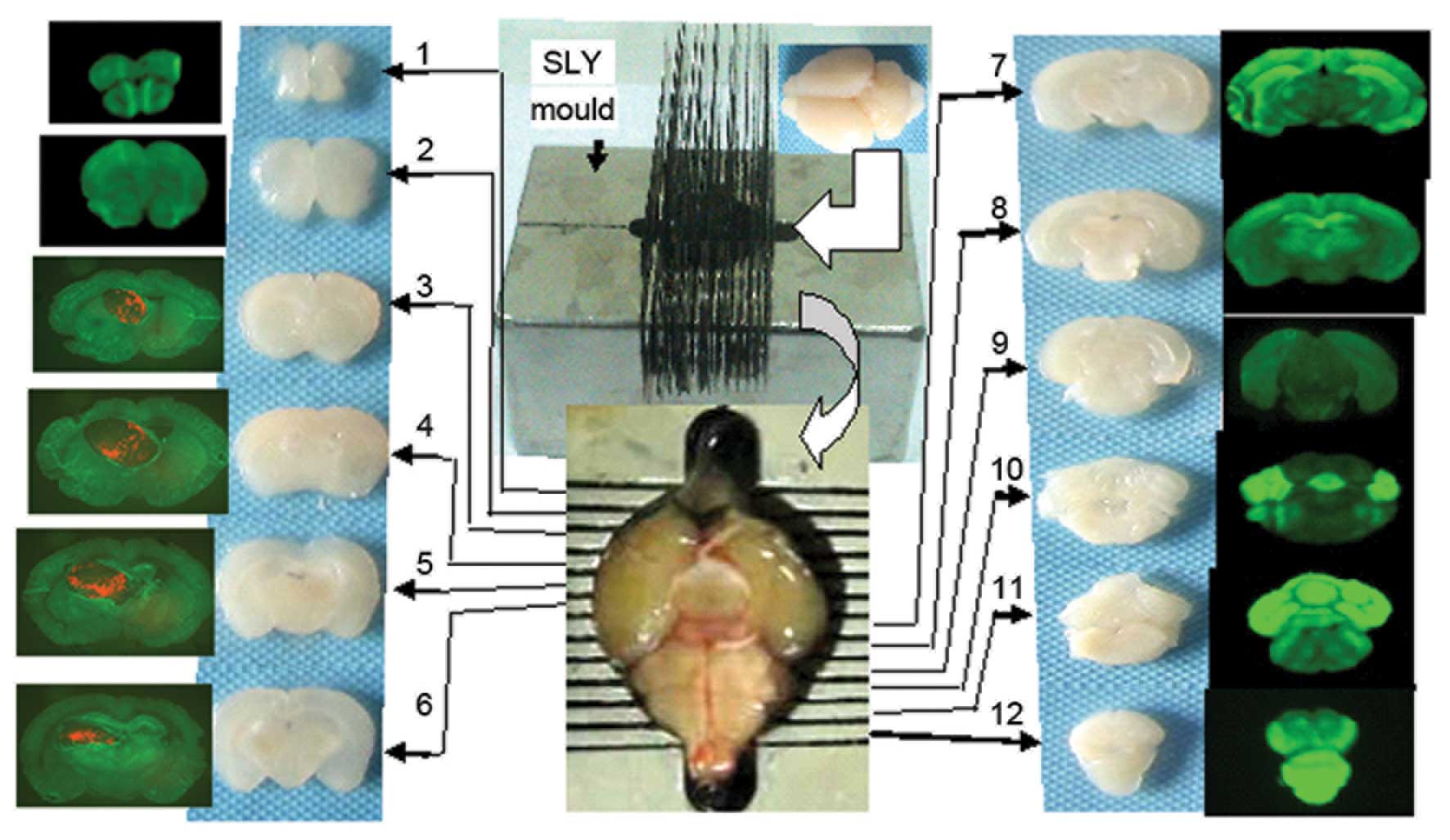
the sucrose solution, and this was performed using the SLY mold, by
which continual, coronal cerebral sections of 1 mm thickness could
be obtained (Fig. 1).
The third to sixth coronal sections (the location of
the transplantation tumor) were embedded in optimal cutting
temperature media, and continual tissue sections of 5 µm thickness
were prepared using a freezing microtome. Subsequently, the
sections were stained with DAPI (1 µg/ml), sealed with anti-fade
mounting medium and stored at −20°C. Routine hematoxylin and eosin
(HE) staining and immunohistochemistry staining with antibodies
against GFAP (1:400), 11-5bCNPase (1:600), CD68 (1:100), Nestin
(1:400) and Ki67 (1:500), were performed for all slices.
Results
Whole-body in vivo fluorescence
imaging of the xenografts
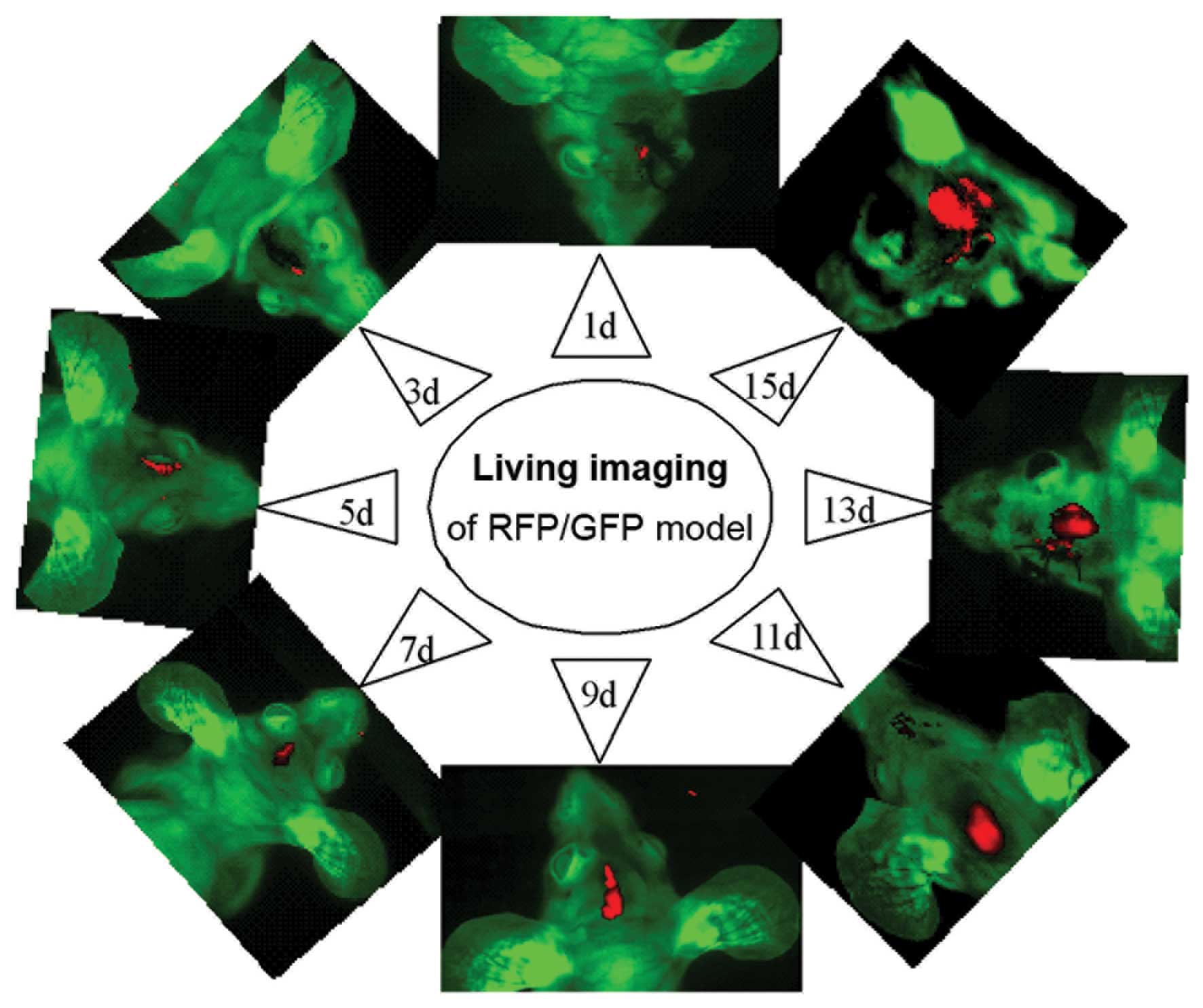
In vivo imaging of the xenografts was
performed prior to pathological study at days 1, 3, 5, 7, 9, 11, 13
and 15, following inoculation of SU3-RFP and C6-CM-Dil cells into
the right caudate nucleus. Dual-color images demonstrated that the
tumor masses emitted red fluorescence from the corresponding
inoculated site, and that the size of the tumors gradually
increased over time (Fig 2).
However, the tumors initiated by SU3-RFP cells grew at a slightly
slower rate, as compared with the tumors consisting of C6-CM-Dil
cells.
Anatomical location of xenografts with
red fluorescence
The section containing the largest tumor was
selected using the SLY coronal mold, and the relative positions of
the tumor and host tissues were detected using a fluorescence
microscope. The exact position and size of the tumor was determined
according to the Paxinos atlas (15). The stark contrast between the red
fluorescence emitted by the transplanted tumor cells and the green
fluorescence emitted by host tissues enabled the identification of
even small tumor masses.
Histological characteristics of the
transplanted tumor
C6-CM-Dil cells inoculated into the caudate nucleus
were able to grow and easily migrate to the cerebral parenchyma,
ventricles, choroid plexus, and subarachnoid cavity. The
histological characteristics of the transplanted tumor resembled
those of the host tissue where the tumor was located at that time,
particularly in the choroid plexus (Fig.
3).
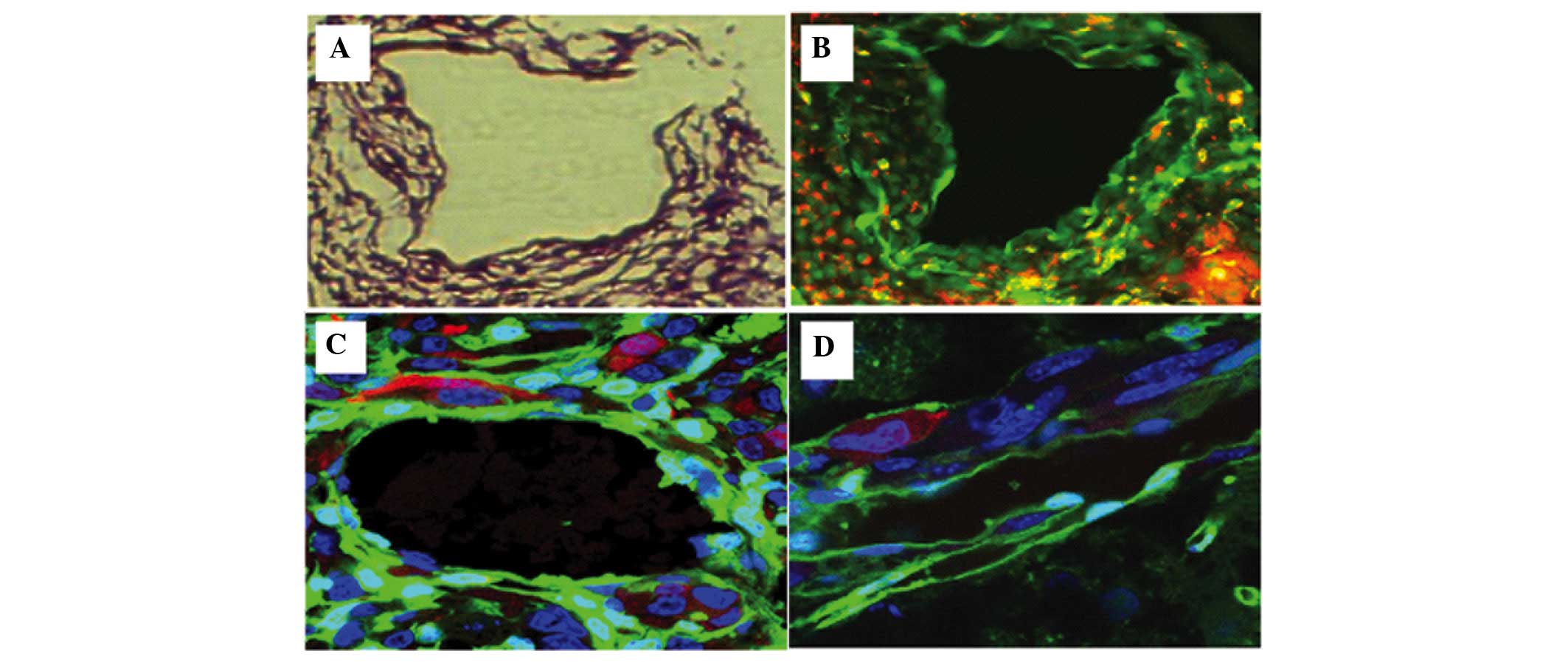
Invasion of tumor cells
SU3 cells expressing RFP were highly invasive, and
were demonstrated to have migrated out and around the tumor in
flocks, with some single cells being scattered away from the edge
of the tumor where the host tissue appeared normal. Dual-color
fluorescence permitted visualization of tumor cells invading the
smooth muscle septum of the host vessel wall, which could not
easily be identified in histological sections without fluorescence
tracing (Fig.4).
Blood vessels of xenografts
The transplanted tumor tissue was observed to
possess numerous blood vessels. The origin of the cells in the
tumor blood vessels could be distinguished by their fluorescence:
Green cells from the host and red cells from the tumor. Yellow
cells corresponded to fusions of tumor and host cells. Furthermore,
fluorescence imaging permitted identification of the origins of the
blood vessel endothelium, elastic fibers and perithelial cells of
the vessel wall, which in turn demonstrated the occurrence of
vascular mimicry.
Cell fusions between the transplanted
tumor and host cells
Wherever the tumors grew, they consisted of red,
green and yellow fluorescent cells. Although the number of yellow
cells was fewer, as compared with the red and green cells, they
could be found throughout the tumorigenesis and development
processes. Under a fluorescence microscope, the SU3-RFP and
C6-CM-Dil cells were red, whereas the host cells were green;
therefore, the yellow cells corresponded to cellular fusions. These
fused cells were detected wherever the tumor had migrated,
including the choroid plexus (Fig.
3), tumor vessels (Fig. 4 and
5) and other cellular interstitialis
of the tumor. Furthermore, laser confocal microscopy demonstrated
that these fusions typically appeared within a portion of or within
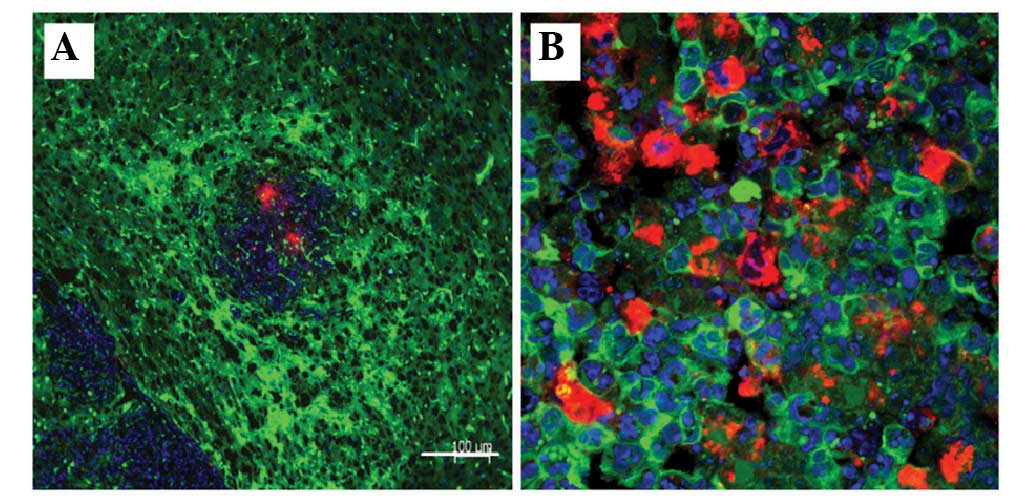
the entire cytoplasm and in the nucleolus (Fig. 6).
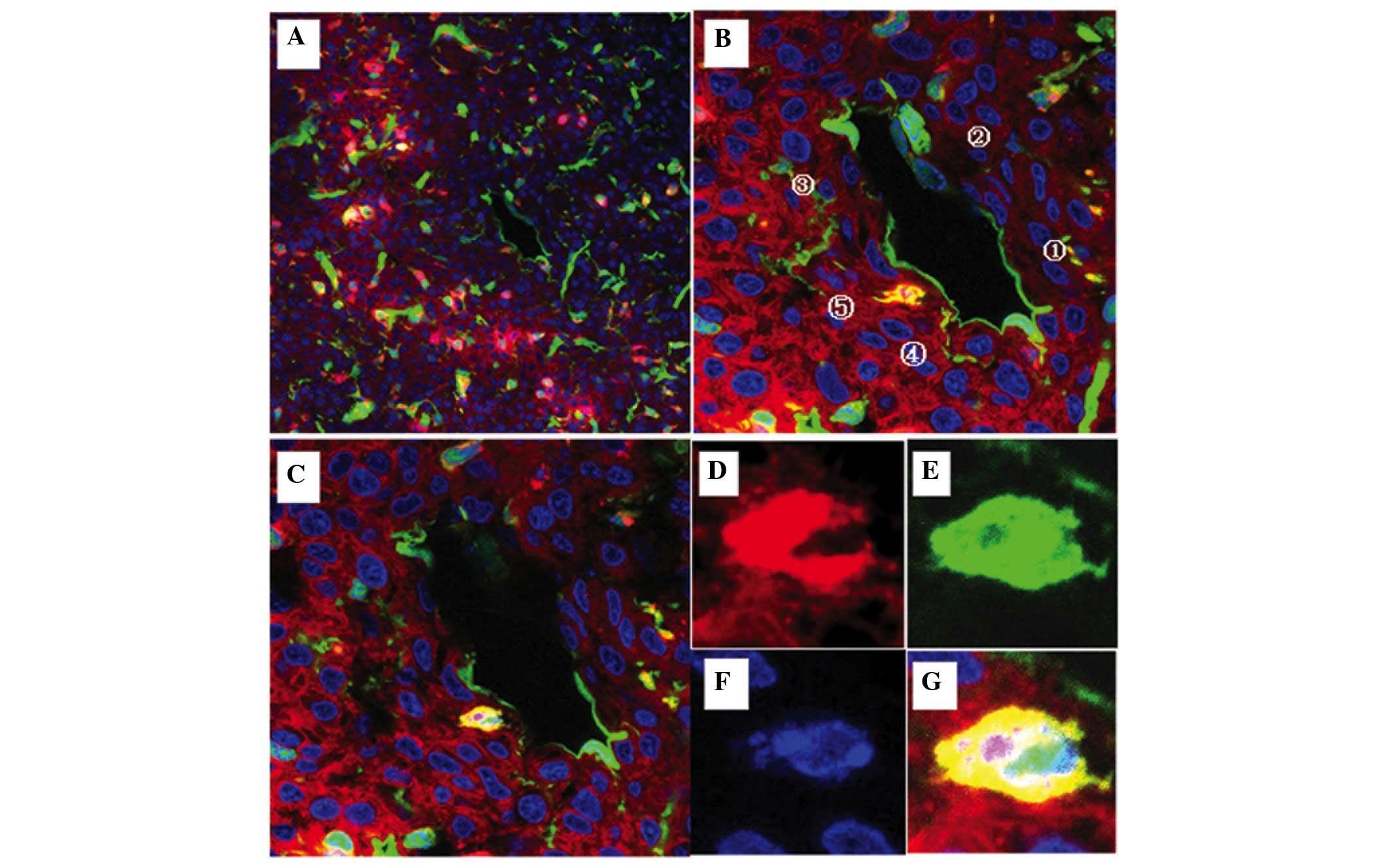
 | Figure 6.Visualization of cell fusion in an
SU3-RFP orthotopic transplantation tumor using fluorescence
tomographic scanning (section thickness, 20 µm) and laser confocal
microscopy. The tumor cells, host cells, karyon and fused cells are
red, green, blue and yellow, respectively. (A-I) Four fusion cells
with various morphologies were observed in 9 sections according to
DAPI staining. (J-M) correspond to amplified portions of (I): (J)
Cell 1, which was entirely fused with another cell, was round, with
a central nucleus and a predominantly yellow cytoplasm; (K) cell 2
was a green cell with a stretched out parapodium, as if attempting
to phagocytose the adjacent yellow cell; (L) cell 3 was a green
cell that was being put under pressure by an adjacent red cell,
with green-cell derived debris apparent (white arrow); and (M) cell
4 was a long fusiform with an eccentric nucleus, which contained
large red granules and small green/yellow granules in the
cytoplasm. |
Tumor microenvironment imaging
The tumor microenvironment contains stromal cells
and interstitial fluid, and mainly comprises the intratumoral and
peripheral microenvironments. In conventional tumor histological
sections, the tumor microenvironment cannot be easily studied due
to the lack of a proper tracing method. However, the method used in
the present study was able to distinguish the cellular origin of
every cell in the tumor mass, and this was particularly important
in areas containing few or no tumor cells. Furthermore as the tumor
color and luster changed, sites where tumor cells were remodeling
their microenvironment, which were called ‘emergency reaction
zones’ in the present study, could be detected (Fig. 7).
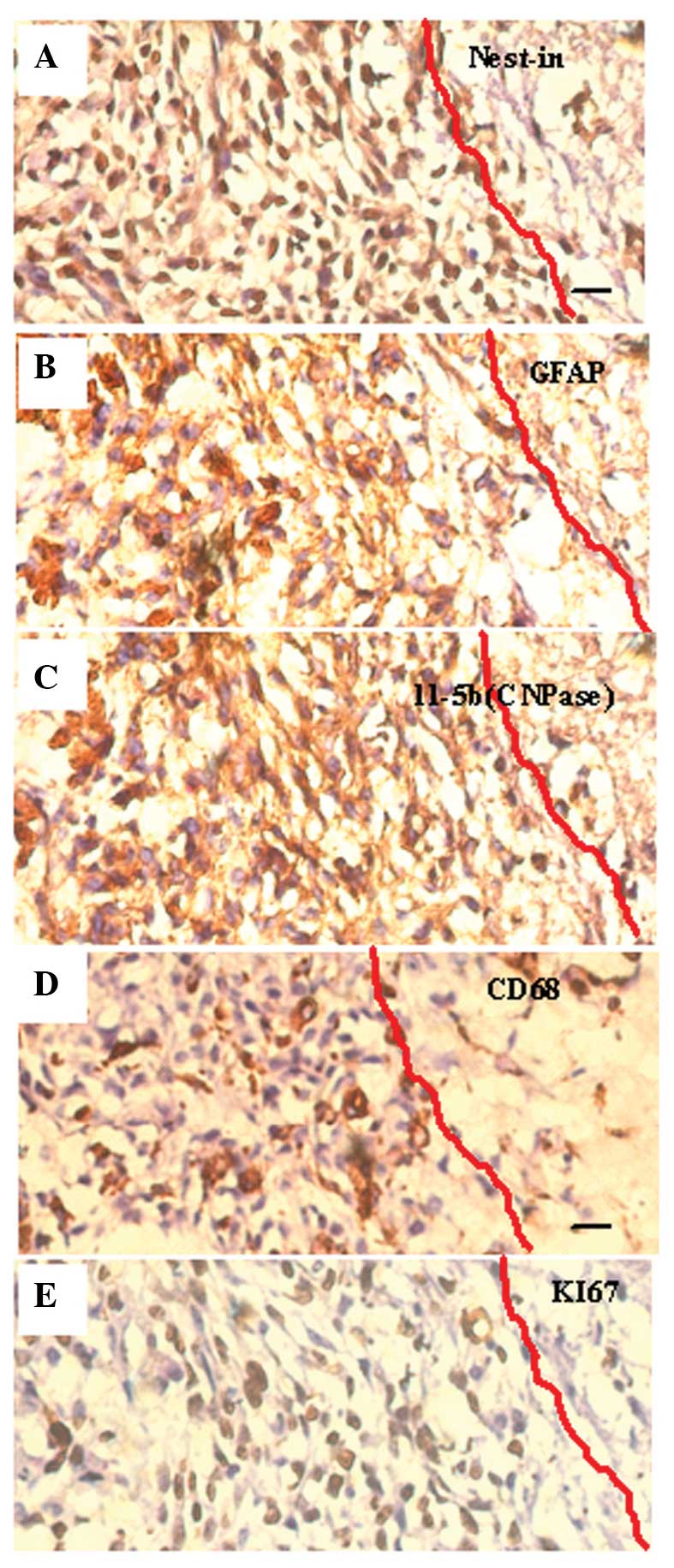
Immunohistochemistry demonstrated the existence of
cells strongly expressing the astrocyte marker protein GFAP,
oligodendroglia marker protein CNPase, and Nestin (the marker of
progenitors of astrocytes and oligodendroglia) in the emergency
reaction zone tissue sections. This phenomenon suggested that
neuralgia cells were forced to dedifferentiate into immature cells
with upregulated expression of Ki-67 under physiopathological
stress. Furthermore, the distribution of CD68+ (marker
protein of macrophage) cells suggests that immune-inflammatory
cells reaching and surrounding the tumor reaction zone may
participate in the physiopathological process (Fig. 8).
Discussion
Prior to the 1960s, non-immunodeficient mice or rats
were widely used as brain tumor models, and were transformed using
carcinogenic agents, including chemical materials and a virus. Due
to the appearance of immunodeficient nude mice and severe combined
immunodeficiency mice in the 1960s, various types of human tumor
xenograft models were successfully established, which greatly
promoted the in vivo study of oncogenicity and the
experimental effectiveness of human cancer (16–18). At
that time, researchers typically used changes in the size of a
tumor as an indicator of effectiveness; however, describing the
precise anatomical location of tumors was challenging. The method
used in the present study permitted easy identification of the
anatomical location of a tumor and allowed the study of the tumor
microenvironment. Charles et al (1) reported that a number of cells in the
tumor microenvironment were recruited from the host bone marrow or
circulation, and had little association with the anatomic site of
the tumor. However, the use of traditional models may result in
some errors, including when determining cell origin using HE
staining, immunohistochemistry and even immunofluorescence
tracing.
The long, carbon-chain carbocyanine dye CM-Dil that
was used in the present study is lipid-soluble, and therefore
becomes incorporated into the plasma membrane. CM-Dil does not
affect the survival, development or basic physiological properties
of the labeled cells, and does not detectably spread from labeled
to unlabeled cells (19). As
compared with RFP, CM-Dil is not integrated into the cell
chromosome and may be lost during cell passaging. However, in a
previous study, cells could be traced in vitro and in
vivo for up to six weeks following CM-Dil labeling, and the
process of CM-Dil labeling was convenient and fast (20).
It is important to investigate every component of
the tumor microenvironment. All interstitial cells, with the
exception of transplanted tumor cells and their daughter cells, are
members of the tumor microenvironment, including host cells
surrounding tumor transplantation sites and host cells that migrate
to the site from the vicinity or from distant sites. When using
histological sections of traditional animal models it is often
difficult to distinguish between tumor and host cells. However, the
dual-color fluorescence-tracing glioma model used in the present
study allowed easy observation and discrimination of these cells,
based on the color of their fluorescence: Red for transplanted
tumor-derived cells and green for host-derived cells.
The dual-color fluorescence-tracing glioma model
facilitated the study of the tumor microenvironment by enabling
easy discrimination between tumor- and host-derived cells, and
permitting visualization of invading tumor cells, including their
remodeling of the host environment. Therefore, the present study
was able to identify the part of the host that was under pressure
from tumor remodeling, based on the precise location of the tumor
cell in SLY brain sections of this model.
In dual-color fluorescence tracing sections, yellow
fluorescent cells, which were expressing both RFP and GFP, and
which can not be easily identified using traditional tumor models,
corresponded to fusion cells of the RFP-expressing tumor cells and
GFP-expressing host cells. It has been suggested that
multinucleated giant cells are fused cells, although fused cells
are not necessarily multinucleated giant cells. Cell fusion has a
crucial function in the genesis and development of a tumor
(21–28), as the daughter cells of the fusion
cells may become immature cells by retrodifferentiation or
transdifferentiation into other blastodermic cells. These
observations have aided progress in the study of tumor
angiogenesis, tumor-associated cell carcinogenesis, tumor cell
heterogeneity and tumor cell remodeling of the microenvironment,
which is induced by tumor stem/progenitor cells. The results of the
present study demonstrated that tumor cells were able to fuse with
various types of cells within their microenvironment, including
gitter, vascular endothelial, vascular smooth muscle and choroid
plexus papillary cells.
Controversy remains with regards to the
transdifferentiation of glioma stem/progenitor cells into vascular
endothelial cells. Cheng et al (29) suggested that tumor cells may
transdifferentiate into pericytes but not into endothelial cells.
However, the results of the present study suggested that
tumor-derived RFP-expressing cells were able to transform into both
endothelial and perithelial cells.
In conclusion, the present study identified the
presence of red, green and yellow fluorescent cells in the vessel
walls of the neoformative vascular region, although their
proportion dynamically changed over time, particularly the number
of yellow, fusion cells. Future studies should endeavor to culture
yellow cells from the transplantation tumor and assess their
differentiation direction towards endothelial and/or perithelial
cells by analyzing their marker proteins.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (grant nos. 81071766, 81172400,
81272799, 81272793, and 81101909) and the Jiangsu Provincial
Special Program of Clinical Medical Science (no. BL2014040), Suzhou
Science and Technology Development Program (nos. SYS201269,
SYSD2013087 and SZS201509).
References
|
1
|
Charles NA, Holland EC, Gilbertson R,
Glass R and Kettenmann H: The brain tumor microenvironment. Glia.
60:502–514. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dong J, Zhang Q, Huang Q, Chen H, Shen Y,
Fei X, Zhang T, Diao Y, Wu Z, Qin Z, et al: Glioma stem cells
involved in tumor tissue remodeling in a xenograft model. J
Neurosurg. 113:249–260. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dong J, Zhao Y, Huang Q, Fei X, Diao Y,
Shen Y, Xiao H, Zhang T, Lan Q and Gu X: Glioma stem/progenitor
cells contribute to neovascularization via transdifferentiation.
Stem Cell Rev. 7:141–152. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Paget S: The distribution of secondary
growths in cancer of the breast. 1889. Cancer Metastasis Rev.
8:98–101. 1989.PubMed/NCBI
|
|
5
|
Fidler IJ: Seed and soil revisited:
Contribution of the organ microenvironment to cancer metastasis.
Surg Oncol Clin N Am. 10:257–269, vii–viiii. 2001.PubMed/NCBI
|
|
6
|
Fidler IJ: The organ microenvironment and
cancer metastasis. Differentiation. 70:498–505. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fidler IJ: The pathogenesis of cancer
metastasis: The ‘seed and soil’ hypothesis revisited. Nat Rev
Cancer. 3:453–458. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hayashi K, Kimura H, Yamauchi K, Yamamoto
N, Tsuchiya H, Tomita K, Kishimoto H, Hasegawa A, Bouvet M and
Hoffman RM: Comparison of cancer-cell seeding, viability and
deformation in the lung, muscle and liver, visualized by
subcellular real-time imaging in the live mouse. Anticancer Res.
31:3665–3672. 2011.PubMed/NCBI
|
|
9
|
Suetsugu A, Katz M, Fleming J, Truty M,
Thomas R, Saji S, Moriwaki H, Bouvet M and Hoffman RM: Non-invasive
fluorescent-protein imaging of orthotopic pancreatic-cancer-patient
tumorgraft progression in nude mice. Anticancer Res. 32:3063–3067.
2012.PubMed/NCBI
|
|
10
|
Bouvet M and Hoffman RM: In vivo imaging
of pancreatic cancer with fluorescent proteins in mouse models.
Methods Mol Biol. 872:51–67. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Menen RS, Hassanein MK, Momiyama M,
Suetsugu A, Moossa AR, Hoffman RM and Bouvet M: Tumor-educated
macrophages promote tumor growth and peritoneal metastasis in an
orthotopic nude mouse model of human pancreatic cancer. Vivo.
26:565–569. 2012.
|
|
12
|
Dong J, Dai XL, Lu ZH, Fei XF, Chen H,
Zhang QB, Zhao YD, Wang ZM, Wang AD, Lan Q and Huang Q: Incubation
and application of transgenic green fluorescent nude mice in
visualization studies on glioma tissue remodeling. Chin Med J
(Engl). 125:4349–4354. 2012.PubMed/NCBI
|
|
13
|
Wu ZC, Huang Q, Shao YX, Xue ZM, Dong J,
Diao Y, Wang AD and Lan Q: Transplantation of human glioma stem
cells in nude mice with green fluorescent protein expression.
Zhonghua Yi Xue Za Zhi. 88:2317–2320. 2008.(In Chinese). PubMed/NCBI
|
|
14
|
Kaye AH, Morstyn G, Gardner I and Pyke K:
Development of a xenograft glioma model in mouse brain. Cancer Res.
46:1367–1373. 1986.PubMed/NCBI
|
|
15
|
Paxinos G and Franklin KBJ: The Mouse
Brain in Stereotaxic Coordinates (2nd). San Diego: Academic Press.
2001.
|
|
16
|
Rana MW, Pinkerton H, Thornton H and Nagy
D: Heterotransplantation of human glioblastoma multiforme and
meningioma to nude mice. Proc Soc Exp Biol Med. 155:85–88. 1977.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fei XF, Zhang QB, Dong J, Diao Y, Wang ZM,
Li RJ, Wu ZC, Wang AD, Lan Q, Zhang SM and Huang Q: Development of
clinically relevant orthotopic xenograft mouse model of metastatic
lung cancer and glioblastoma through surgical tumor tissues
injection with trocar. J Exp Clin Cancer Res. 29:842010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen H, Dong J and Huang Q: Xenograft
model of human brain tumor. Brain Tumors: Current and Emerging
Therapeutic Strategies. Abujamra AL: (Shanghai, China). InTech.
3–20. 2011.
|
|
19
|
Honig MG and Hume RI: Fluorescent
carbocyanine dyes allow living neurons of identified origin to be
studied in long-term cultures. J Cell Biol. 103:171–187. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Weir C, Morel-Kopp MC, Gill A, Tinworth K,
Ladd L, Hunyor SN and Ward C: Mesenchymal stem cells: Isolation,
characterisation and in vivo fluorescent dye tracking. Heart Lung
Circ. 17:395–403. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
He X, Tsang TC, Pipes BL, Ablin RJ and
Harris DT: A stem cell fusion model of carcinogenesis. J Exp Ther
Oncol. 5:101–109. 2005.PubMed/NCBI
|
|
22
|
Shinn-Thomas JH, Scranton VL and Mohler
WA: Quantitative assays for cell fusion. Methods Mol Biol.
475:347–361. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dittmar T, Nagler C, Schwitalla S, Reith
G, Niggemann B and Zänker KS: Recurrence cancer stem cells - made
by cell fusion? Med Hypotheses. 73:542–547. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lu X and Kang Y: Cell fusion hypothesis of
the cancer stem cell. Adv Exp Med Biol. 714:129–140. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nagler C, Zänker KS and Dittmar T: Cell
fusion, drug resistance and recurrence CSCs. Adv Exp Med Biol.
714:173–182. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Goldenberg DM: Horizontal transmission of
malignancy by cell-cell fusion. Expert Opin Biol Ther. 12(Suppl 1):
S133–S139. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Parris GE: Historical perspective of
cell-cell fusion in cancer initiation and progression. Crit Rev
Oncog. 18:1–18. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Harkness T, Weaver BA, Alexander CM and
Ogle BM: Cell fusion in tumor development: Accelerated genetic
evolution. Crit Rev Oncog. 18:19–42. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cheng L, Huang Z, Zhou W, Wu Q, Donnola S,
Liu JK, Fang X, Sloan AE, Mao Y, Lathia JD, et al: Glioblastoma
stem cells generate vascular pericytes to support vessel function
and tumor growth. Cell. 153:139–152. 2013. View Article : Google Scholar : PubMed/NCBI
|