Introduction
Early intervention in developmental seizure-induced
brain damage is a focus area in recent pediatric clinical and
animal studies. The autophagic/lysosomal pathway has been recently
considered as a promising therapy for preventing and treating
neurodegenerative diseases, in addition to seizure-induced brain
injuries (1–3). However, the underlying molecular
mechanism of its efficacy remain unclear.
Developmental seizure-induced long-term adverse
effects include neurodysfunction and pathological phenomena such as
regenerative aberrant mossy fiber sprouting in the hippocampus,
which is positively correlated with long-term neurofunctional
impairment (4–6). Hippocampal mossy fibers are the axons
of dentate gyrus granule cells. Overstimulation of mossy fibers in
epileptic activities leads to aberrant regenerative sprouting,
which is associated with aberrant zinc homeostasis (7,8). In a
previous study the present authors showed that recurrent neonatal
seizures could cause long-term cognitive dysfunction and disturbed
hippocampal expression of plasticity related gene 1 (PRG-1), which
could be reversed by pretreatment with the autophagy inhibitor
cathepsin B inhibitor (CBI) (9). The
results of another study showed that the autophagy inhibitor
3-methyladenine (3-MA) regulated the acute phase expression of
autophagy and zinc transporter-associated genes at 1.5, 3, 6 and 24
h after the last seizures (10).
These results showed linear correlations among
microtubule-associated protein light chain 3 (LC3), beclin-1, zinc
transporter 1 (ZnT-1) and ZnT-2 transcripts in the hippocampus and
cerebral cortex in the recurrent seizure group, which could be
modulated by 3-MA. However, there are no reports investigating the
long-term alterations in the mRNA expression levels of ZnT signals
in hippocampus pretreatment with autophagy inhibitor following
developmental seizures.
The aim of the present study was to characterize the
long-term expression profiles of ZnT-associated genes in the
hippocampus following developmental seizures. It was hypothesized
that the disturbed expression of ZnTs would correlate with
increases in hippocampal axon sprouting which could be inhibited by
pretreatment with E-64d, another autophagy inhibitor.
Materials and methods
Animals and grouping
A total of 130 Sprague-Dawley male rats (obtained
from the Experimental Animal Center of Zhejiang University,
Hangzhou, China) at postnatal day 21 (P21) were used in all
experiments. Animals were treated in accordance with the guidelines
set by the National Institutes of Health for the humane treatment
of animals and were approved by the animal care and use committee
of Soochow University. Attempts were made to minimize the number of
animals used. Rats were randomly assigned to three groups: Control
(CONT; n=30); penicillin-induced recurrent seizures group (RS;
n=50); and the E-64d-treated seizure group (E64D; n=50). Penicillin
(5.1 U/mg dissolved in saline; RS and E64D; North China
Pharmaceutical Co., Ltd., Shijiazhuang, China) or saline (CONT) was
administered intraperitoneally (i.p.) to induce seizure, as
previously reported (8). Penicillin
was injected every other day for 11 consecutive days from postnatal
day 21 (P21). E64D group rats were pretreated with E-64d
(Sigma-Aldrich, St. Louis, MO, USA) prior to seizure induction (4
µl, 1 mg/µl/day, i.p.). Thirty-four animals died due to serious
generalized seizures. The animals suffering generalized seizures
(stages 5 and 6, bilateral forelimb clonus with rearing and loss of
postural control) (8) were used for
subsequent experiments (n=30 per seizure group).
Western blot analysis
At 3, 6 and 24 h after the final seizure, rats (n=30
per group) were sacrificed by 4% chloral hydrate (1 ml/100 g i.p.;
Sinopharm Chemical Reagent Co., Ltd., Shanghai, China; cat. no.
30037517) and the hippocampus tissues in every time point from each
group (n=6) were rapidly removed and immediately frozen in dry ice
for western blot analysis, as described previously by Luo et
al (11). Briefly, samples were
homogenized in western blot analysis buffer containing 10 mM
Tris-HCl (pH 7.4), 150 mM NaCl, 1% v/v Triton X-100, 1% sodium
deoxycholate, 5 mM EDTA (all chemicals obtained from
Sigma-Aldrich). The homogenate was then centrifuged at 10,000 rpm
for 10 min at 4°C to obtain the supernatant, which was stored at
−70℃ until further use. Subsequently, 30 µg protein from each
sample was subjected to 10% SDS-PAGE. After blocking overnight at
4°C with skimmed milk (cat. no. 232100; BD Biosciences, Franklin
Lakes, NJ, USA), the blots were incubated with one of the following
antibodies: Goat polyclonal anti-beclin-1 (1:1,000; cat. no.
sc-11427; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) and
rabbit polyclonal anti-B-cell lymphoma 2 (Bcl-2; 1:100; cat. no.
sc-492; Santa Cruz Biotechnology, Inc.) in Tris-buffered saline
containing 0.2% Tween-20 (TBST; Sinopharm Chemical Reagent Co.,
Ltd.; cat. no. 30189328) and 5% nonfat dry milk overnight at 4°C.
Following overnight incubation, membranes were washed and incubated
with horseradish peroxidase-conjugated secondary antibody in TBST
for 2 h (goat anti-rabbit IgG and rabbit anti-goat IgG; cat. no.
GAR0072 and RAG0072, respectively; dilution, 1:3,000; MultiSciences
Biotech Co., Ltd., Hangzhou, China). Immunoreactivity was detected
using enhanced chemiluminescent autoradiography (Western BrightECL
kit; cat. no. k-12045-D50; Advansta Inc., Menlo Park, CA, USA). The
relative changes in the intensity of each immunoreactive band were
evaluated using SigmaScan Pro 5.0 (Systat Software, Inc., San Jose,
CA, UK) and were normalized against the loading control
β-actin.
Timm staining
A subset of rats (n=6 each group) underwent Timm
staining on P51 according to the method previously described
(12). The contents of the Timm
staining solution were obtained from Sinopharm Chemical Reagent
Co., Ltd., and were as follows: Gum arabic powder (cat. no.
9000-01-5); hydroquinone (cat. no. 10011317); citric acid
monohydrate (cat. no. 10007118); trisodium citrate dihydrate (cat.
no. 10019418); and silver nitrate (cat. no. 10018461). Sprouting
was analyzed at a magnification of ×10 using an Olympus CX-40 light
microscope (Olympus Corp., Tokyo, Japan).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
A total of 6 rats from each group were sacrificed
using 4% chloral hydrate (1 ml/100 g i.p.) at P51. Hippocampal
samples from each group were subjected to RT-qPCR analysis, as
described previously (8). Briefly,
total RNA was extracted using TRIzol reagent (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) according to the
manufacturer's instructions. The concentration, purity and quantity
of the total isolated RNA was determined by measuring the optical
density at 260 and 280 nm by UV spectrophotometry using a
NanoPhotometer UV/Vis spectrophotometer (Implen, Munich, Germany).
Next, 2 µg total RNA was reverse transcribed into cDNA using 1 µg
random primers (Promega Corp., Madison, WI, USA), 1.5 µl M-MLV
Reverse Transcriptase and 0.6 µl RNase inhibitor (Thermo Fisher
Scientific, Inc.). For RT, the total reaction volume was 40 µl and
the sample was incubated at 37° for 60 min, followed by 95°C for 5
min. Subsequently, qPCR was performed with an Applied Biosystems
7500 Real-Time PCR system (Thermo Fisher Scientific, Inc.) using
TaqMan Gene Expression Master Mix (Thermo Fisher Scientific, Inc.).
The total volume of the PCR reaction mixture was 25 µl, including
2.5 µl 10X buffer; 1.5 µl Mg2+ (25 mM); 0.5 µl dNTP (10
mM); 0.25 µl hot start Taq DNA polymerase; 0.5 µl of each primer;
and 0.3 µl TaqMan probe. The primers and probes used in qPCR for
the zinc transporter genes (ZnT-2, ZnT-4, ZnT-5, ZnT-6 and ZnT-7),
divalent cation transporter 1 (DCT-1), Zrt-Irt-like protein-6
(ZIP-6), ZIP-7, cathepsin D and cathepsin L were designed against
the GenBank sequences using the Primer Express 2.0 software
(Applied Biosystems; Thermo Fisher Scientific, Inc.), and are shown
in Table I. The PCR cycling
conditions were 95°C for 10 min (initial denaturation), and 40
cycles of 95°C for 15 sec and 60°C for 60 sec. Quantitative
fluorescent data were collected and analyzed using SDS software,
version 1.3 (Thermo Fisher Scientific, Inc.). Threshold cycle data
were determined by setting the default threshold to 0.08; when the
fluorescence intensity reaches this threshold value, the PCR cycle
number was regarded to be the threshold cycle (Cq) value.
Initially, the RT-qPCR Cq value of the target mRNAs and the
internal control (β-actin) were determined, and then the ΔCq method
of relative quantification was used to determine the fold changes
in expression. The fold change in expression was obtained using the
2−ΔΔCq method (13).
 | Table I.Oligonucleotide primers for reverse
transcription-quantitative polymerase chain reaction analysis. |
Table I.
Oligonucleotide primers for reverse
transcription-quantitative polymerase chain reaction analysis.
| Gene | Genbank accession
no. | Primer sequence |
|---|
| ZnT-2 | RNU50927 | F,
5′-GGCTGGATCCTGGACTAATGTT-3′ |
|
|
| R,
5′-ACACCCCAAAATCCCTTTCTG-3′ |
|
|
| Probe,
5′-FAM-CTCACACCACAGCTGGAGAGACACTGAGG-TAMRA-3′ |
| ZnT-4 | NM_172066.1 | F,
5′-GCTGAAGCAGAGGAAGGTGAA-3′ |
|
|
| R,
5′-TCTCCGATCATGAAAAGCAAGTAG-3′ |
|
|
| Probe,
5′-FAM-CAGGCTGACCATCGCTGCCGT-TAMRA-3′ |
| ZnT-5 | NM_001106404.1 | F,
5′-CCAGCACATGTCTGGCCTAA-3′ |
|
|
| R,
5′-TTTGCAGTACTTCATGGATTCCA-3′ |
|
|
|
Probe,5′-FAM-CACTGGCTTCCACGATGTCCTGGCTAT-TAMRA-3′ |
| ZnT-6 | NM_001106708.1 | F,
5′-CGGCATTATCCCAGGACTCA-3′ |
|
|
| R,
5′-CCAGCAAGATCGATCAGAACAA-3′ |
|
|
| Probe,
5′-FAM-TTCTTGCCCCGCATGAACCCG-TAMRA-3′ |
| ZnT-7 | XM_001073594.1 | F,
5′-TTGGGATCCGCGTCTGA-3′ |
|
|
| R,
5′-CCCTCTAGAAGTGACTCGGTATGG-3′ |
|
|
| Probe,
5′-FAM-TCGTCTCTGCTGTCACTGCCGCC-TAMRA-3′ |
| DCT-1 | NM_001108855.1 | F,
5′-TGGCTGCCTCAGCATCTG-3′ |
|
|
| R,
5′-CCCAGTTGTTGAGTGGTTTGG-3′ |
|
|
| Probe,
5′-FAM-AAAACCAGAGCCCCTTCCCTACGCA-TAMRA-3′ |
| ZIP-6 | NM_001024745.1 | F,
5′-CTCACTGCTGGCTTGTTCATG-3′ |
|
|
| R,
5′-CGTGATCACTGGCATCATTGT-3′ |
|
|
| Probe,
5′-FAM-TCGCTCTGGTCGACATGGTACCCG-TAMRA-3′ |
| ZIP-7 | NM_001008885.1 | F,
5′-GACTTGGCACACAACTTCACAGA-3′ |
|
|
| R,
5′-GGACTGTCATCGTGGTCAGGAT-3′ |
|
|
| Probe,
5′-FAM-CTGGCCATTGGTGCTTCCTTCCG-TAMRA-3′ |
| Cathepsin D | NM_134334 | F,
5′-CATCGCAGCCAAGTTTGATG-3′ |
|
|
| R,
5′-CCGGGAGCACATTGTTAACA-3′ |
|
|
| Probe,
5′-FAM-CATCTTGGGCATGGGCTACCCTTTTATC-TAMRA-3′ |
| Cathepsin L | NM_013156 | F,
5′-GGGTATGGATGGCTACATCAAAA-3′ |
|
|
| R,
5′-GGCTGCGGTGGCAAGTC-3′ |
|
|
| Probe,
5′-FAM-AGCCAAAGACCGGAACAACCACTGC-TAMRA-3′ |
Statistical analysis
The protein and mRNA expression levels
(2−ΔΔCq) were analyzed with post-hoc comparisons using
analysis of variance, followed by Bonferroni test. Timm staining
scores were compared using a non-parametric Kruskal-Wallis test.
SAS 8.0 statistical software (SAS Institute, Cary, NC, USA) was
used to perform statistical analysis. Data is presented as the mean
± standard deviation. P<0.05 was considered to indicate a
statistically significant difference.
Results
Western blot analysis
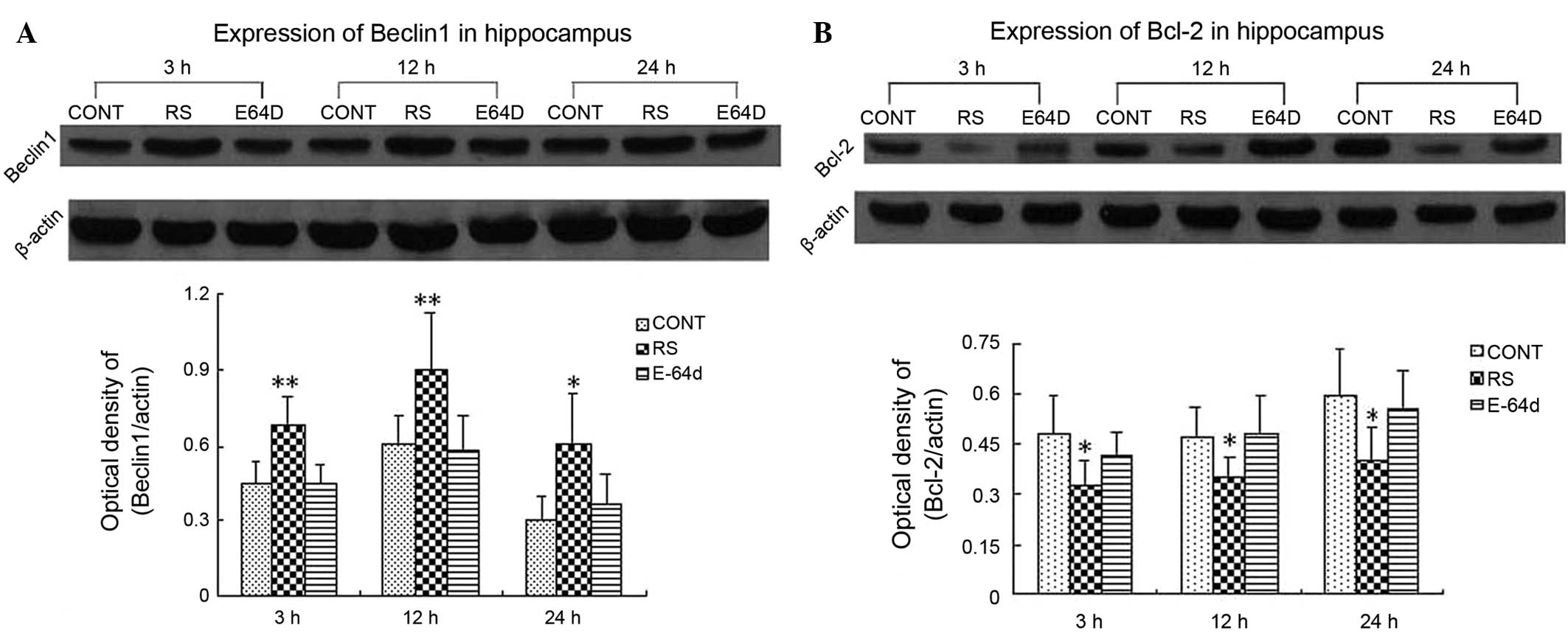
Significant increases in the hippocampal protein
expression levels of beclin-1 (Fig.
1A) and reductions in those of Bcl-2 (Fig. 1B) were observed at 3, 12 and 24 h
after the last seizures in the RS group. beclin-1 upregulation and
Bcl-2 downregulation were mitigated by pretreatment with E-64d.
Timm staining
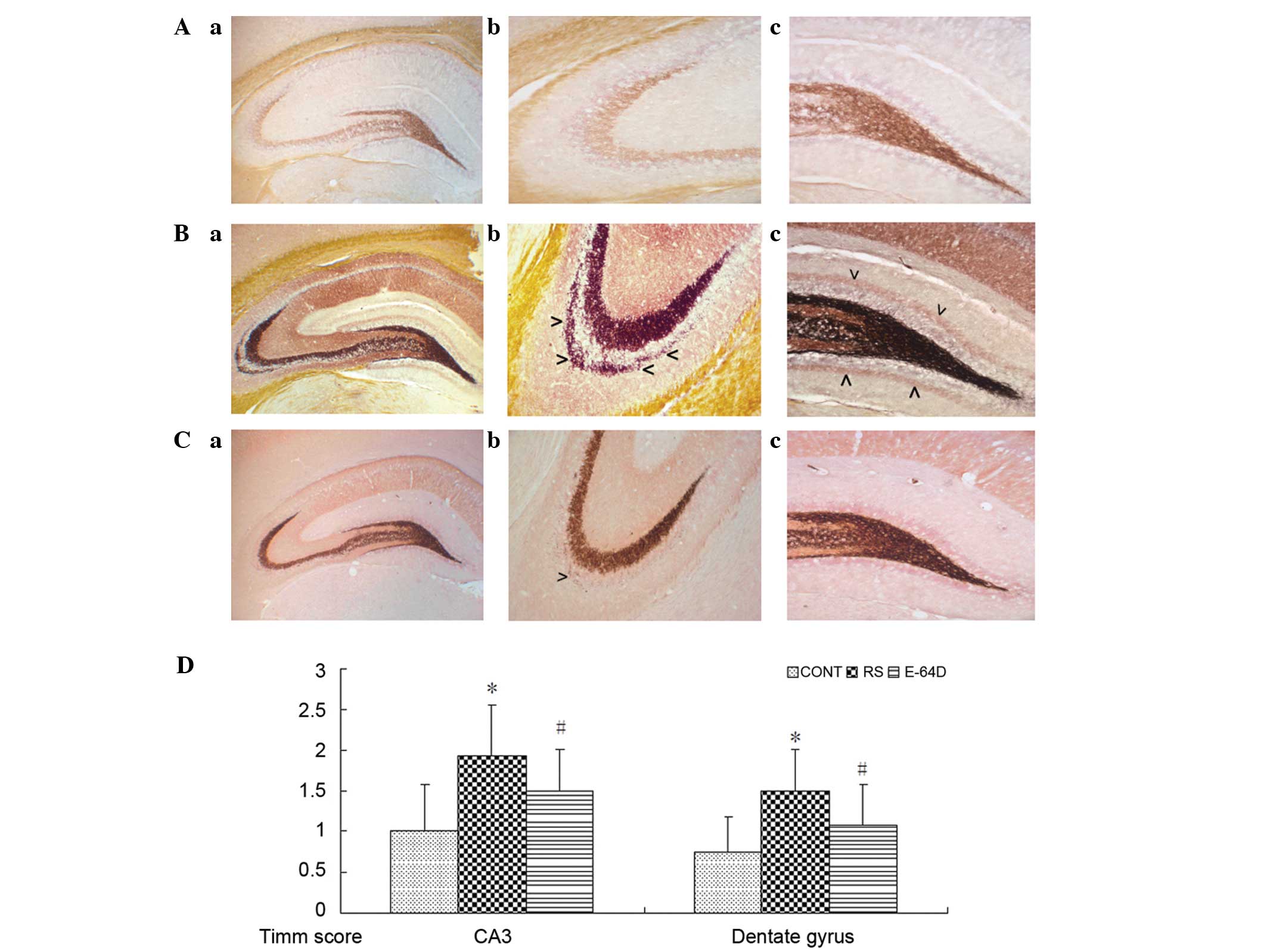
As shown in Fig. 2A,
there were no evident mossy fiber terminals in the control
hippocampus. There was prominent aggregation of mossy fiber
terminals in the stratum pyramidale of CA3 subfield (Fig. 2B-b) and the dentate gyrus (Fig. 2B-c) compared with the CONT group. In
the E64D group rats (Fig. 2C), the
aggregation of mossy fiber terminals was markedly decreased in the
supragranular region of dentate gyrus and CA3 subfield (Fig. 2C-b and 2C-c) compared with the RS
group.
RT-qPCR analysis
RT-qPCR was employed to evaluate the relative mRNA
expression levels of ZnT-2, ZnT-4, ZnT-5, ZnT-6, ZnT-7, DCT-1,
ZIP-6, ZIP-7, cathepsin D and cathepsin L in the hippocampus at
P51. As shown in Fig. 3A, RS group
rats showed a significant upregulation of the mRNA expression of
ZnT-4, ZnT-5, ZIP-7 and cathepsin D compared with the CONT group
rats (P<0.05). In addition, there was long-term downregulated
expression of ZnT-4, ZIP-6, ZIP-7 and cathepsin L in the E64D group
rats compared with the RS group (P<0.05).
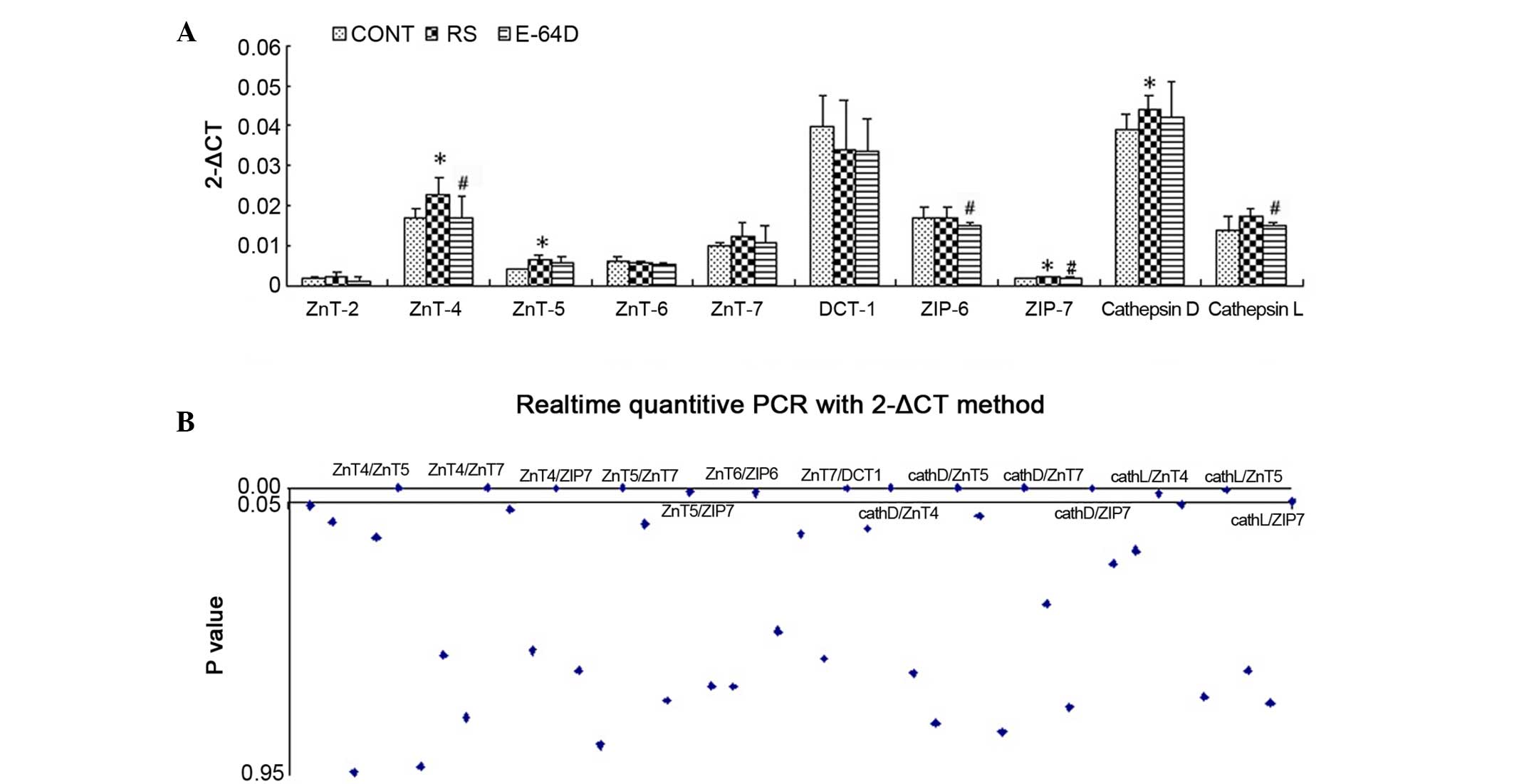 | Figure 3.mRNA expression levels as identified
using reverse transcription-quantitative polymerase chain reaction
analysis. (A) Expression of ZnT-2, ZnT-4, ZnT-5, ZnT-6, ZnT-7,
DCT-1, ZIP-6, ZIP-7, cathepsin D and cathepsin L in the rat
hippocampus after behavioral analysis. (B) Pearson's linear
correlation analyses of the ten genes. Data are expressed as the
mean ± standard deviation. Data was analyzed with post hoc
comparisons using a Bonferroni test after analysis of variance.
*P<0.05 vs. CONT group; #P<0.05 vs. RS group.
CONT, control group; RS, recurrent seizure group; E64D,
E-64d-treated group; Znt, zinc transporter; DCT-1, divalent cation
transporter 1; ZIP, Zrt-Irt-like protein. |
Linear correlation analysis showed a total of 45
Pearson correlation coefficients among the ten genes. As shown in
Fig. 3B and Table II, 14 of the Pearson correlation
coefficients showed significant inter-relationship (P<0.05);
primarily among ZIP-7, ZnT-4, ZnT-5, ZnT-7, cathepsin D and
cathepsin L. The results suggest that the ZnTs/ZIP7/cathepsin
signaling pathway may serve a key role in the neuroprotective
effects of E-64d.
| Table II.Linear correlation analysis among the
10 genes, showing the 14 Pearson correlation coefficients
(r) with statistically significant inter-associations. |
Table II.
Linear correlation analysis among the
10 genes, showing the 14 Pearson correlation coefficients
(r) with statistically significant inter-associations.
| Associated
genes | Coefficient
value | P-value |
|---|
| ZnT-4/ZnT-5 | 0.90 | <0.0001 |
| ZnT-4/ZnT-7 | 0.89 | <0.0001 |
| ZnT-4/ZIP-7 | 0.64 | <0.01 |
| ZnT-5/ZnT-7 | 0.90 | <0.0001 |
| ZnT-5/ZIP-7 | 0.57 | <0.05 |
| ZnT-6/ZIP-6 | 0.56 | <0.05 |
| ZnT-7/DCT-1 | 0.64 | <0.01 |
| Cathepsin
D/ZnT-4 | 0.72 | <0.001 |
| Cathepsin
D/ZnT-5 | 0.74 | <0.001 |
| Cathepsin
D/ZnT-7 | 0.70 | <0.01 |
| Cathepsin
D/ZIP-7 | 0.62 | <0.01 |
| Cathepsin
L/ZnT-4 | 0.53 | <0.05 |
| Cathepsin
L/ZnT-5 | 0.60 | <0.01 |
| Cathepsin
L/ZIP-7 | 0.47 | <0.05 |
Discussion
E-64d is an autophagy inhibitor that has been shown
to perform a neuroprotective function in spinal cord injury,
hypoxic-ischemic brain injury, acute optic neuritis (14–17), in
addition to kainic acid-induced excitotoxicity of primary striatal
neurons (18). However, the
molecular mechanisms underlying the efficacy of E-64d remain
unclear. The aim of the present study was to elucidate the
neuroprotective effects of autophagy inhibitors on developmental
seizure-induced brain damage by neuropathological and molecular
biology analyses, focusing on ZnT-associated gene expression in the
hippocampus.
In accordance with previous results (19), pretreatment with E-64d prior to the
induction of seizures markedly attenuated the long-term hippocampal
mossy fiber sprouting. Furthermore, E-64d treatment resulted in the
downregulated mRNA expression of ZnT-4, ZIP-6, ZIP-7 and cathepsin
L at the long-term time point compared with the RS group. To the
best of our knowledge, the present study is the first to
investigate the long-term effects of an autophagy inhibitor on the
expression profiles of ZnTs in hippocampus following developmental
seizures.
Previous results showed that CBI, an autophagy
inhibitor, could improve functional deficits and inhibit
hippocampal sprouting by modulating long-term PRG-1 expression in
the hippocampus and cerebral cortex (9). In another study (10), we reported for the first time that
pretreatment with 3-MA, another autophagy inhibitor, markedly
attenuated neonatal seizure-induced ZnT-1, ZnT-2, LC3 and beclin-1
elevations in the hippocampus at the acute phase time point
following seizures. However, the present study primarily examined
the ZnT-associated mRNA expression at a long-term time point
following seizures during adolescence (P51), a time point which is
in parallel with the genesis of mossy fiber sprouting in
hippocampus. The results suggest that the regulators for zinc
homeostasis may be involved in E-64d-mediated neuroprotection
following developmental seizure-induced excitotoxicity.
The divalent cation Zn2+, an essential
trace element that performs structural and cofactor functions, is
concentrated in synaptic vesicles, particularly in the hippocampal
zinc-rich mossy fiber pathway, which has the highest quantity of
zinc in brain (20). The presence of
this type of vesicular zinc in the mossy fiber system may be
detected using Timm staining, where its excessive synaptic movement
from pre- to post-synaptic neurons via ZnTs contributes to
regenerative sprouting following epilepsy (21). Among the ten ZnTs that have been
identified to date, ZnT-2, ZnT-4, ZnT-5, ZnT-6 and ZnT-7 are of
particular interest due to their function in intracellular
organelles. ZnT-2 and ZnT-4 transport zinc into endosomes and
lysosomes, while ZnT-5, ZnT-6 and ZnT-7 appear to be localized on
the Golgi apparatus (22). A study
by McCormick and Kelleher indicated that ZnT-4 provides zinc to
zinc-dependent proteins in the trans-Golgi network, which directly
contributed to labile zinc accumulation as ZnT-4 overexpression
increased FluZin3 (zinc probe used to monitor zinc fluxes during
fluorescent imaging) fluorescence (23). As FluZin3 is regarded as a marker for
detecting sprouting mossy fibers in the temporal lobe epileptic
hippocampus (24), it is possible
that ZnT-4 overexpression in the brain may be a molecular marker
for hippocampal mossy fiber sprouting. The present results showed
the upregulation of the transcription of ZnT-4 in the RS group
compared with the CONT group. Notably, this elevated mRNA
expression of ZnT-4 was significantly downregulated by pretreatment
with E-64d. As mentioned above, ZnT-4 transports zinc into
endosomes and lysosomes, which are also the target organelles for
autophagy inhibitors. The present results indicate that ZnT-4 may
contribute to E-64d-mediated improvement of morphological and
cognitive functions, which merits further investigation. In
addition, this study identified significantly increased mRNA
expression of ZnT-5 and a increasing trend of ZnT-7 in the RS group
compared with the CONT group. Furthermore, linear correlation
analysis showed significant inter-relationships among ZnT-4, ZnT-5
and ZnT-7. Combined with previous results, which suggest that ZnT-5
and ZnT-7 contribute synergic to the activation of phosphatases in
cytoplasmic membrane (25), it is
reasonable to speculate that ZnT-4, ZnT-5 and ZnT-7 may contribute
synergically to the neuroprotective effects of E-64d in an animal
model of penicillin-induced developmental seizures.
Apart from ZnTs, the regulation of cellular zinc
homeostasis is controlled closely by a number of different
mechanisms. Hence, we further investigated the expression of ZIP-6,
ZIP-7 and DCT-1 in hippocampus. ZIP is responsible for the movement
of zinc into a cell from the extracellular space, and transports
zinc into the cytoplasm from organelles that contain zinc (22). Among the 14 mammalian members of the
ZIP family, the ZIP transporters ZIP-6 and ZIP-7 have been detected
in the brain (26). DCT-1 (also
known as divalent metal transporter 1) is present in the
hippocampal pyramidal and granule cells, cerebellar granule cells
and pyramidal cells of the piriform cortex in high densities
(27). In the present study, ZIP-7
showed significantly upregulated mRNA expression in the RS group
compared with CONT group rats, while no significant differences
were observed in the mRNA levels of ZIP-6 and DCT-1 between the RS
and CONT groups. In addition, the upregulated expression of ZIP-7
was significantly inhibited by pretreatment with E-64d.
Furthermore, linear correlation analysis demonstrated significant
inter-relationships between ZIP-7 and ZnT-4 or ZnT-5. This result
suggests that ZIP-7 is correlated with the long-term hippocampal
mossy fiber sprouting and cognitive processing following
developmental seizures. Previous studies have reported that ZIP-7
plays a critical role in zinc homeostasis (28), and is implicated in aberrant cellular
biochemical signaling such as tyrosine kinase activation and growth
factor signaling (29,30). Collectively, these results suggest
that ZIP-7 functions as a critical node in zinc-mediated aberrant
mossy fiber sprouting in hippocampus and may be a novel target for
the intervention effect of E-64d following developmental
seizure-induced brain damage.
A previous study by Luo et al (31) showed that the inhibition of autophagy
by 3-MA and bafliomycin A1 could reduce traumatic brain injury
(TBI)-induced cell damage and attenuate behavioral outcomes via the
inhibition of the TBI-induced upregulation of LC3, beclin-1 and the
beclin-1/Bcl-2 ratio in the hippocampus. The present results also
observed the modulatory effects of E-64d on beclin-1 and Bcl-2,
which were observed at 3 and 12 h after the last seizures. Notably,
the present RT-qPCR results showed upregulated cathepsin D
expression in the RS group compared with the CONT group.
Furthermore, pretreatment with E-64d significantly inhibited the
elevated mRNA expression of cathepsin L in comparison with the RS
group rats. Cathepsin D and cathepsin L are acid lysosomal
hydrolases. It has been reported that the majority of
zinc-containing vacuoles of cultured retinal cells are lysosomes,
and that the accumulation of zinc in lysosomes induced the release
of cathepsin D into the cytosol, which may be a key mechanism of
ethambutol-induced retinal cell death (32). These results are in accordance withe
present study, in which the expression of ZnT-4, ZnT-5, ZIP-7 and
cathepsin D was upregulated in RS group rats compared with CONT
group rats. In addition, long-term downregulated expression of
ZnT-4, ZIP-6, ZIP-7 and cathepsin L was detected in E64D group rats
compared with the RS group. Linear correlation analysis further
indicated significant inter-relationships among ZIP-7, ZnT-4,
ZnT-5, ZnT-7, cathepsin D and cathepsin L. Based on these findings,
it is postulated that perturbations of intralysosomal zinc and
cathepsin metabolism, and the subsequent results in lysosomal
dysfunction, may underlie the neuronal dysfunction characteristic
of developmental seizure-induced brain damage. Thus, the lysosomal
modulation by E-64d represents a novel approach for treating brain
damage induced by developmental seizures.
In conclusion, the present study provides insights
into the abnormalities in signal transduction of zinc, reflected by
ZnTs and ZIP. These results suggest that the E-64d attenuates
hippocampal mossy fiber sprouting, at least in part, via the
modulation of the ZnT-4/ZIP-7/cathepsin signaling pathway. These
results may offer a novel strategy for the development of
therapeutic interventions for treatment of developmental
seizure-induced brain damage.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant nos. 81271458 and 81471337) and
the Jiangsu Province's Key Provincial Talents Program (grant no.
RC2011113), a project funded by the Priority Academic Program
Development of Jiangsu Higher Education Institutions.
References
|
1
|
Au AK, Bayir H, Kochanek PM and Clark RS:
Evaluation of autophagy using mouse models of brain injury. Biochim
Biophys Acta. 1802:918–923. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bae N, Ahn T, Chung S, Oh MS, Ko H, Oh H,
Park G and Yang HO: The neuroprotective effect of modified
Yeoldahanso-tang via autophagy enhancement in models of Parkinson's
disease. J Ethnopharmacol. 134:313–322. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jing CH, Wang L, Liu PP, Wu C, Ruan D and
Chen G: Autophagy activation is associated with neuroprotection
against apoptosis via a mitochondrial pathway in a rat model of
subarachnoid hemorrhage. Neuroscience. 213:144–153. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Holopainen IE: Seizures in the developing
brain: Cellular and molecular mechanisms of neuronal damage,
neurogenesis and cellular reorganization. Neurochem Int.
52:935–947. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dulac O, Nabbout R, Plouin P, Chiron C and
Scheffer IE: Early seizures: Causal events or predisposition to
adult epilepsy? Lancet Neurol. 6:643–651. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Holmes GL: Effects of seizures on brain
development: Lessons from the laboratory. Pediatr Neurol. 33:1–11.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ni H, Jiang YW, Bo T, Wang JM and Wu XR:
C-Fos, N-methyl-D-aspartate receptor 2C, GABA-A-alpha 1
immonoreactivity, seizure latency and neuronal injury following
single or recurrent neonatal seizures in hippocampus of Wistar rat.
Neurosci Lett. 380:149–154. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ni H, Jiang YW, Tao LY, Cen JN and Wu XR:
Effects of penicillin-induced developmental epilepticus on
hippocampal regenerative sprouting, related gene expression and
cognitive deficits in rats. Toxicol Lett. 188:161–166. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ni H, Yan JZ, Zhang LL, Feng X and Wu XR:
Long-term effects of recurrent neonatal seizures on neurobehavioral
function and related gene expression and its intervention by
inhibitor of cathepsin B. Neurochem Res. 37:31–39. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ni H, Feng X, Gong Y, Tao LY and Wu XR:
Acute phase expression pattern of ZnTs, LC3 and beclin-1 in rat
Hippocampus and its regulation by 3-methyladenine following
recurrent neonatal seizures. Biol Trace Elem Res. 143:320–331.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Luo CL, Chen XP, Yang R, Sun YX, Li QQ,
Bao HJ, Cao QQ, Ni H, Qin ZH and Tao LY: Cathepsin B contributes to
traumatic brain injury-induced cell death through a
mitochondria-mediated apoptotic pathway. J Neurosci Res.
88:2847–2858. 2010.PubMed/NCBI
|
|
12
|
Ni H, Jiang YW, Tao LY, Jin MF and Wu XR:
ZnT-1, ZnT-3, CaMKII, PRG-1 expressions in hippocampus following
neonatal seizure-induced cognitive deficit in rats. Toxicol Lett.
184:145–150. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Johnson MR, Wang K, Smith JB, Heslin MJ
and Diasio RB: Quantitation of dihydropyrimidine dehydrogenase
expression by real-time reverse transcription polymerase chain
reaction. Anal Biochem. 278:175–184. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ray SK, Matzelle DC, Wilford GG, Hogan EL
and Banik NL: E-64-d prevents both calpain upregulation and
apoptosis in the lesion and penumbra following spinal cord injury
in rats. Brain Res. 867:80–89. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ray SK, Matzelle DD, Wilford GG, Hogan EL
and Banik NL: Cell death in spinal cord injury (SCI) requires de
novo protein synthesis. Calpain inhibitor E-64-d provides
neuroprotection in SCI lesion and penumbra. Ann NY Acad Sci.
939:436–449. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tsubokawa T, Solaroglu I, Yatsushige H,
Cahill J, Yata K and Zhang JH: Cathepsin and calpain inhibitor E64d
attenuates matrix metalloproteinase-9 activity after focal cerebral
ischemia in rats. Stroke. 37:1888–1894. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tsubokawa T, Yamaguchi-Okada M, Calvert
JW, Solaroglu I, Shimamura N, Yata K and Zhang JH: Neurovascular
and neuronal protection by E64d after focal cerebral ischemia in
rats. J Neurosci Res. 84:832–840. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dong XX, Wang YR, Qin S, Liang ZQ, Liu BH,
Qin ZH and Wang Y: P53 mediates autophagy activation and
mitochondria dysfunction in kainic acid-induced excitotoxicity in
primary striatal neurons. Neuroscience. 207:52–64. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ni H, Ren SY, Zhang LL, Sun Q, Tian T and
Feng X: Expression profiles of hippocampal regenerative
sprouting-related genes and their regulation by E-64d in a
developmental rat model of penicillin-induced recurrent
epilepticus. Toxicol Lett. 217:162–169. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Takeda A: Zinc signaling in the
hippocampus and its relation to pathogenesis of depression. Mol
Neurobiol. 44:166–174. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mitsuya K, Nitta N and Suzuki F:
Persistent zinc depletion in the mossy fiber terminals in the
intrahippocampal kainate mouse model of mesial temporal lobe
epilepsy. Epilepsia. 50:1979–1990. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nakashima AS and Dyck RH: Zinc and
cortical plasticity. Brain Res Rev. 59:347–373. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
McCormick NH and Kelleher SL: ZnT4
provides zinc to zinc-dependent proteins in the trans-Golgi network
critical for cell function and Zn export in mammary epithelial
cells. Am J Physiol Cell Physiol. 303:C291–C297. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sutula TP and Dudek FE: Unmasking
recurrent excitation generated by mossy fiber sprouting in the
epileptic dentate gyrus: An emergent property of a complex system.
Prog Brain Res. 163:541–563. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Suzuki T, Ishihara K, Migaki H, Matsuura
W, Kohda A, Okumura K, Nagao M, Yamaguchi-Iwai Y and Kambe T: Zinc
transporters, ZnT5 and ZnT7, are required for the activation of
alkaline phosphatases, zinc-requiring enzymes that are
glycosylphosphatidylinositol-anchored to the cytoplasmic membrane.
J Biol Chem. 280:637–643. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lichten LA and Cousins RJ: Mammalian zinc
transporters: Nutritional and physiologic regulation. Annu Rev
Nutr. 29:153–176. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mims MP and Prchal JT: Divalent metal
transporter 1. Hematology. 10:339–345. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yan G, Zhang Y, Yu J, Yu Y, Zhang F, Zhang
Z, Wu A, Yan X, Zhou Y and Wang F: Slc39a7/zip7 plays a critical
role in development and zinc homeostasis in zebrafish. PLoS One.
7:e429392012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Taylor KM, Vichova P, Jordan N, Hiscox S,
Hendley R and Nicholson RI: ZIP7-mediated intracellular zinc
transport contributes to aberrant growth factor signaling in
antihormone-resistant breast cancer Cells. Endocrinology.
149:4912–4920. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hogstrand C, Kille P, Nicholson RI and
Taylor KM: Zinc transporters and cancer: A potential role for ZIP7
as a hub for tyrosine kinase activation. Trends Mol Med.
15:101–111. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Luo CL, Li BX, Li QQ, Chen XP, Sun YX, Bao
HJ, Dai DK, Shen YW, Xu HF, Ni H, et al: Autophagy is involved in
traumatic brain injury-induced cell death and contributes to
functional outcome deficits in mice. Neuroscience. 184:54–63. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chung H, Yoon YH, Hwang JJ, Cho KS, Koh JY
and Kim JG: Ethambutol-induced toxicity is mediated by zinc and
lysosomal membrane permeabilization in cultured retinal cells.
Toxicol Appl Pharmacol. 235:163–170. 2009. View Article : Google Scholar : PubMed/NCBI
|