Introduction
Schwann cells are specialised glial cells and
myelin-forming cells in the peripheral nervous system, which serve
a critical role in nerve remyelination after injuries (1). Following peripheral nerve damage,
immature Schwann cells differentiate into a myelinating phenotype
secreting myelin proteins, including myelin-associated glycoprotein
(MAG) and myelin basic protein (MBP), and provide a
growth-permissive environment for nerve regeneration (2). It has been reported that the seeding of
Schwann cells significantly promotes nerve regeneration following
lesion development (3), while damage
to Schwann cells induces delay of nerve growth (4). Therefore, differentiated Schwann cells
with myelinating properties are required following nerve injury. In
addition, the use of bioactive substances, such as fingolimod and
methylcobalamin, that are capable of promoting Schwann cell
proliferation and improving their myelinating properties is an
important strategy for improving nerve regeneration (5,6).
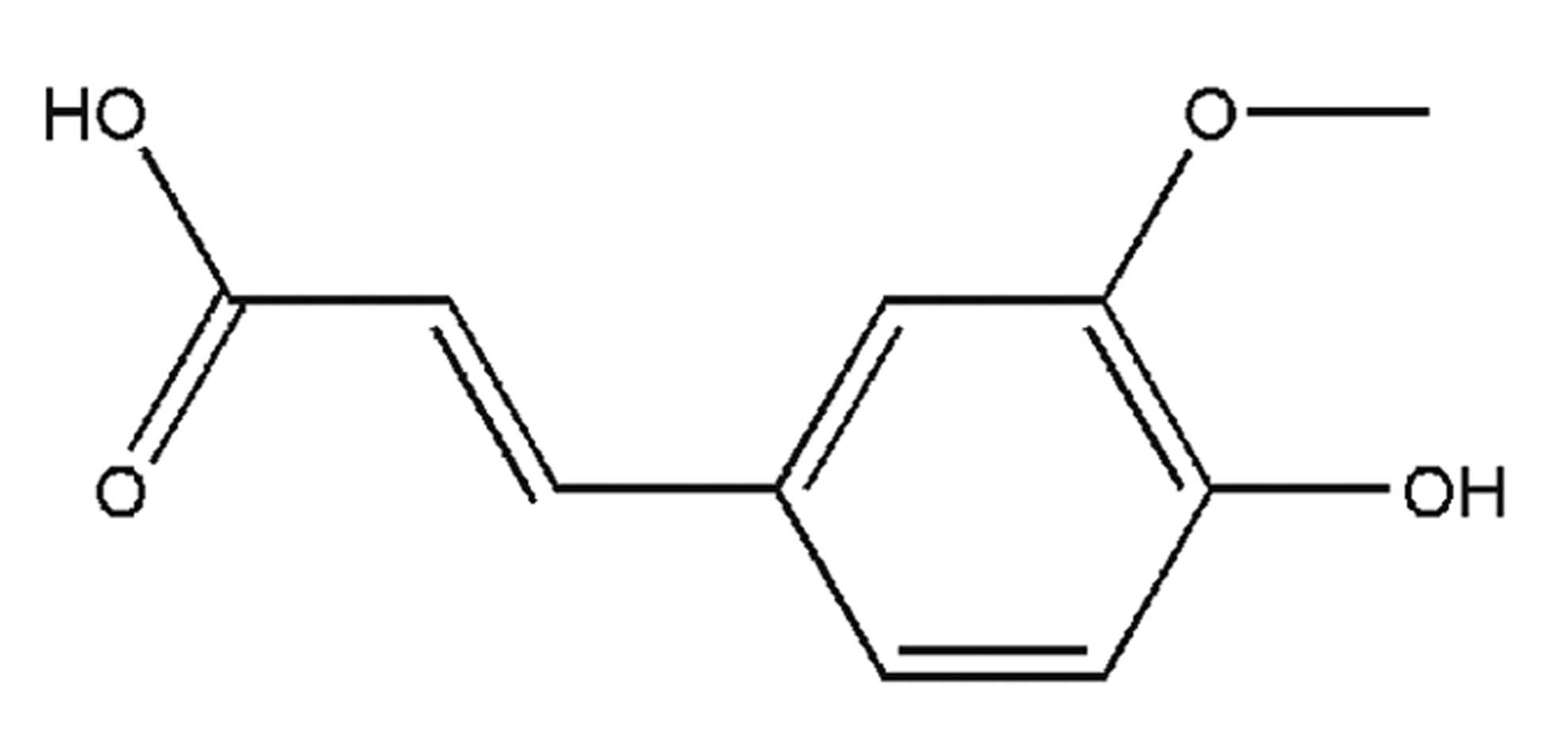
Ferulic acid (FA) is a phenolic compound isolated
from several plants, such as Ferula assa-foetida L. The
molecular structure of FA is shown in Fig. 1. FA has potent anti-oxidant and
anti-inflammatory properties, and exhibits a wide range of
therapeutic effects, including anticancer, antidiabetic and
cardioprotective activities (7).
Previous studies have indicated that FA also exerts potent
neuroprotective effects, such as protecting cultured neurons
against glutamate neurotoxicity (8),
preserving neuronal cells against cerebral ischaemia (9) and inducing proliferation of neuronal
progenitor cells (10). However, the
majority of current studies concerning FA have focused on
neuroprotection of neurons, while little is known about the effects
of FA on Schwann cells and the signalling pathway by which FA
brings about these effects.
Nerve crush injury is a well-established animal
model used to investigate the impact of pharmacological treatments
for nerve regeneration (11).
Different chemical drugs have been investigated to improve the
remyelination of the peripheral nerve in animal experiments, such
as clozapine, olanzapine and quetiapine (12). However, functional recovery
subsequent to peripheral injury is often poor, and severe
anatomical and functional disorders are observed. Currently,
naturally occurring components have gained emerging interest for
their potentially beneficial use in neurodegeneration, and thus it
is critical to develop pharmacological approaches that may assist
in the promotion of remyelination in the regrowth of nerves.
The aims of the present study were to evaluate the
impact of FA on the cell proliferation and differentiation of
Schwann cells in vitro and in vivo, and to determine
the effect of FA on nerve remyelination in a rat model of
peripheral nerve crush injury. The current study provided evidence
that FA significantly promoted the proliferation and expression of
MAG and MBP in cultured Schwann cells through MEK1/ERK1/2
signalling transduction. The study also identified that systematic
FA administration effectively promoted sciatic nerve remyelination
in the crushed nerves through increasing Schwann cell proliferation
and differentiation. Therefore, the present study revealed the
effects of FA on Schwann cell proliferation and differentiation,
and suggested a potential role for FA in the therapy of peripheral
nerve injuries.
Materials and methods
Animals
Adult Sprague Dawley (SD) rats weighing 250–275 g,
aged 6 months (n=5 per group), were provided by the Experimental
Animal Center at the Chengdu Military General Hospital (Chengdu,
China) and maintained with a 12 h light/dark cycle and ad
libitum access to food and water. All procedures were performed
in accordance with the Guidelines for the Care and Use of
Laboratory Animals (National Institutes of Health, Bethesda, MD,
USA) and were approved by the Administration Committee of
Experimental Animals of Sichuan Province, China. All efforts were
made to minimise the number of animals used and their suffering
during the experiments.
In vitro isolation and culture of
primary Schwann cells
Primary cultures of Schwann cells were obtained from
the sciatic nerves of 3-day-old SD rats using the method previously
described by Fei et al (13).
Schwann cells were then maintained in complete Dulbecco's modified
Eagle's medium (DMEM) at 37°C and 5% CO2. The purity and
viability of cultured primary rat Schwann cells were determined by
flow cytometry with FITC-anti-S100 antibody (1:1,000; ab76749;
Abcam, Cambridge, MA, USA) for Schwann cells and 7-aminoactinomycin
D (BioLegend, San Diego, CA, USA) for cell viability.
Schwann cell proliferation assay
In Schwann cells in vitro proliferation
assays, cells were cultured (1×106 cells/ml for 48 h) in
5% fetal bovine serum in triplicate in 96-well flat-bottom
microtiter plates and treated with FA (Sigma-Aldrich, St. Louis,
MO, USA). Cell proliferation was determined by cell counting kit-8
(CCK-8) assay (Dojindo Molecular Technologies, Inc., Kumamoto,
Japan), as previously described (14). In 3H-methyl-thymidine
proliferation experiments, cells were pulsed with 0.25 µCi
3H-methyl-thymidine (Shenggong Trade Co., Shanghai,
China) at the last 16 h of cell culture and cell proliferation was
expressed as the average counts per minute of
3H-thymidine uptake, as previously described (15).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA of Schwann cells and sciatic nerves was
isolated using TRIzol LS reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) and reverse transcribed into
cDNA using NCode™ miRNA First-Strand cDNA Synthesis kit
(Invitrogen; Thermo Fisher Scientific, Inc.). The resulting cDNA
was used to quantitatively measure the expression of genes using
Power SYBR® Green Master Mix (Invitrogen; Thermo Fisher
Scientific, Inc.). Primers (Wuhanboshide, China) used to measure
gene expression levels were as follows: GAPDH sense,
GACATGCCGCCTGGAGAAAC, and antisense, AGCCCAGGATGCCCTTTAGT; MAG
sense, ACAGCGTCCTGGACATCATCAACA, and antisense,
ATGCAGCTGACCTCTACTTCCGTT; MBP sense, TTGACTCCATCGGGCGCTTCTTTA, and
antisense, GCTGTGCCACATGTACAAGGACTCA.
Western blotting
Similarly to the cell culturing in the proliferation
assay, the Schwann cells were treated with FA in the absence or
presence of PD98059 (Sigma-Aldrich). Schwann cells and sciatic
nerves were lysed in ice-cold radioimmunoprecipitation assay buffer
(Shenggong Trade Co.). The lysed cells were centrifuged (12,000 ×
g, 25 min, 4°C) and the supernatants were collected. A standard
western blotting protocol was used, as previously described
(16). Next, the samples were
incubated with primary antibodies at 4°C overnight, including MAG
(1:500; sc-15324), MBP (1:500; sc-809), MEK1 (1:1,000; sc-219),
ERK1/2 (1:1,000; sc-292838), phosphorylated (p)-MEK1 (1:500;
sc-293106), p-ERK1/2 (1:500; sc-101760), which were purchased from
Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA), as well as
GAPDH (1:500; AB-P-R 001), from Goodhere Biotechnology Co., Ltd.
(Hangzhou, China). Subsequently, the samples were incubated with
the horseradish peroxidase-conjugated goat anti-rabbit IgG
secondary antibody (1:500; A0208), obtained from Beyotime Institute
of Biotechnology (Haimen, China). The blots were then detected by
chemiluminescence using enhanced chemiluminescence reagent (BeyoECL
Plus; Beyotime Institute of Biotechnology). Images of the blots
were captured by Carestream Image Station 4000MM (Carestream Health
Inc., Woodbridge, CT, USA), and the band density was detected by
Quantity One (version 4.6.2; Bio-Rad Laboratories, Inc., Hercules,
CA, USA).
Surgical procedure and treatment
Crush injury of rat sciatic nerves was performed
according to the method previously described by Varejão et
al (17), with minor
modifications. Briefly, rats were anaesthetised, and the right
sciatic nerve was carefully exposed through the gluteal-splitting
approach and then crushed with a haemostatic clamp at a
standardised force at moderate level for 30 sec. Animals in the
treatment group (n=5) received FA (50 mg/kg, intraperitoneal
injection) daily for 7 days after the injury, while the control
group rats (n=5) received the same amount of phosphate-buffered
saline (PBS) injection at the same time periods. At 2 or 4 weeks
after the beginning of the experiments, rats were euthanised and
the right sciatic nerve segment distal to the lesion was removed
and stored. Schwann cells in sciatic nerves were harvested using a
rat Schwann cell isolation kit from Miltenyi Biotec GmbH (Bergisch
Gladbach, Germany). The cell proliferation of isolated Schwann
cells was determined by flow cytometry with a proliferation marker
PE-conjugated anti-Ki-67 antibody from eBioscience Inc. (1:200;
cat. no. 12-5698; San Diego, CA, USA).
Evaluation and statistical
analysis
The data are expressed as the mean ± standard error
of the mean. The Mann-Whitney U test was performed to determine
statistically significant differences between FA-treated and
non-treated groups. GraphPad Prism version 5.0 software (GraphPad
Software, Inc., La Jolla, CA, USA) was used to perform statistical
analysis. Statistically significant differences were determined at
P<0.05.
Results
FA increases primary Schwann cell
proliferation in vitro
Primary rat Schwann cells were successfully isolated
and cultured, as demonstrated by flow cytometric analysis (Fig. 2A). In order to study the influence of
FA on Schwann cells proliferation in vitro, Schwann cells
were treated with various concentrations of FA (0, 25, 50 and 100
µM) and cultured in 5% serum DMEM for 0, 24, 48 or 72 h. As shown
in Fig. 2B, Schwann cell viability
was greatly increased following FA treatment compared with that in
the control cells, as demonstrated by CCK-8 assay (P<0.05).
Subsequently, the radioactive 3H-thymidine incorporation
assessment was applied in order to precisely determine the effects
of FA on Schwann cell proliferation. The results demonstrated that
FA significantly induced Schwann cell proliferation in a
dose-dependent manner (Fig. 2C).
These results confirmed that FA is involved in the induction of a
cell proliferative reaction of Schwann cells.
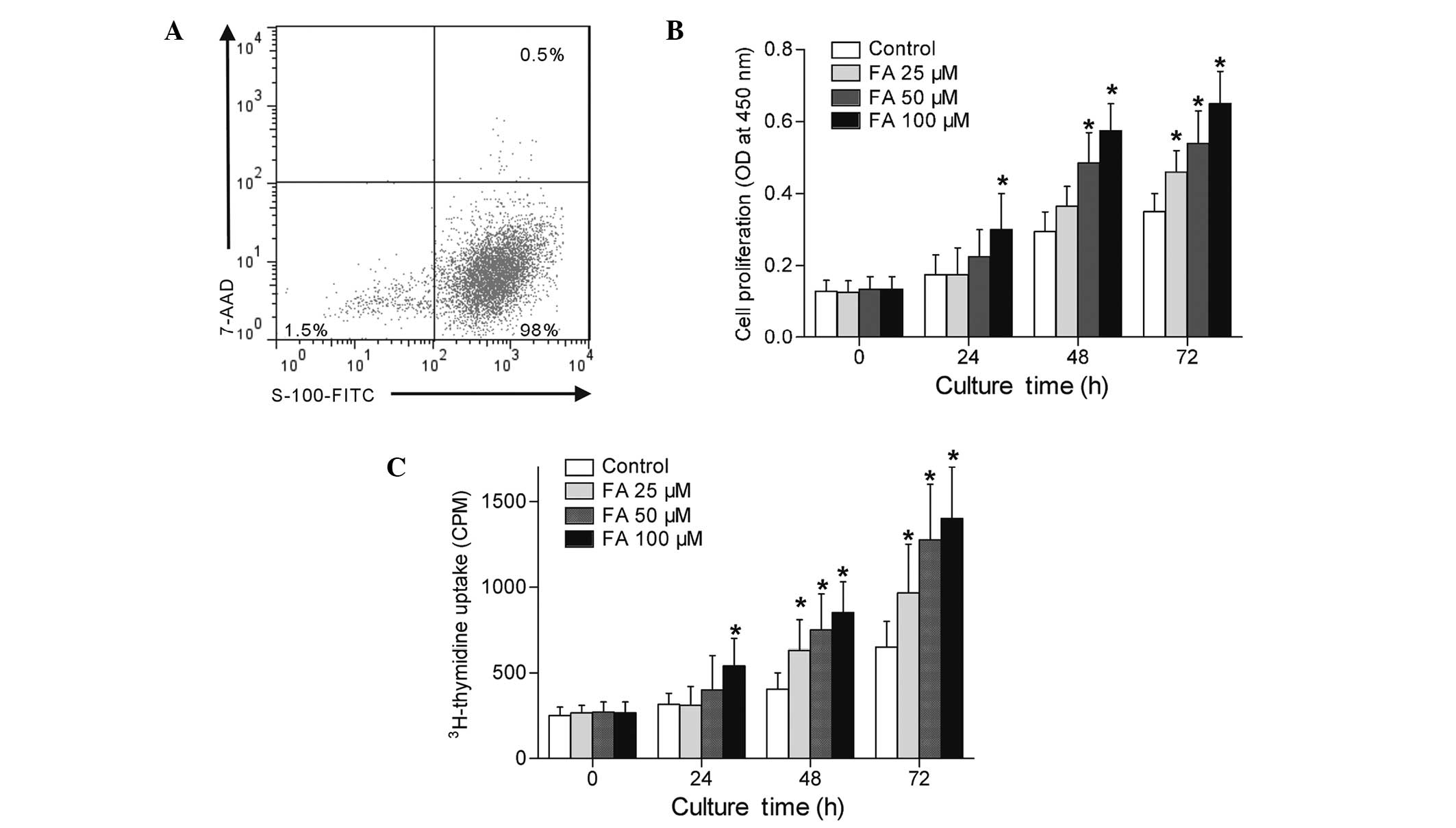 | Figure 2.FA increased cell proliferation of
primary Schwann cells in vitro. Schwann cells were treated
with FA (0, 25, 50 and 100 µM) and cultured for 0, 24, 48 or 72 h
in Dulbecco's modified Eagle's medium containing 5% fetal bovine
serum. (A) Flow cytometric identification of the purity and
viability of cultured primary Schwann cells (S-100 positive for
Schwann cells and 7-AAD negative for live cells). (B) Cell
proliferation determined by CCK-8 assay, with the results shown as
the OD at 450 nm. (C) Cell proliferation determined by
3H-thymidine uptake, with the results shown as CPM. Data
in the graphs are expressed as the mean ± standard error of three
separate experiments. *P<0.05, vs. control. FA, ferulic acid;
7-AAD, 7-aminoactinomycin D; CCK-8, cell counting kit-8; OD,
optical density; CPM, counts per minute. |
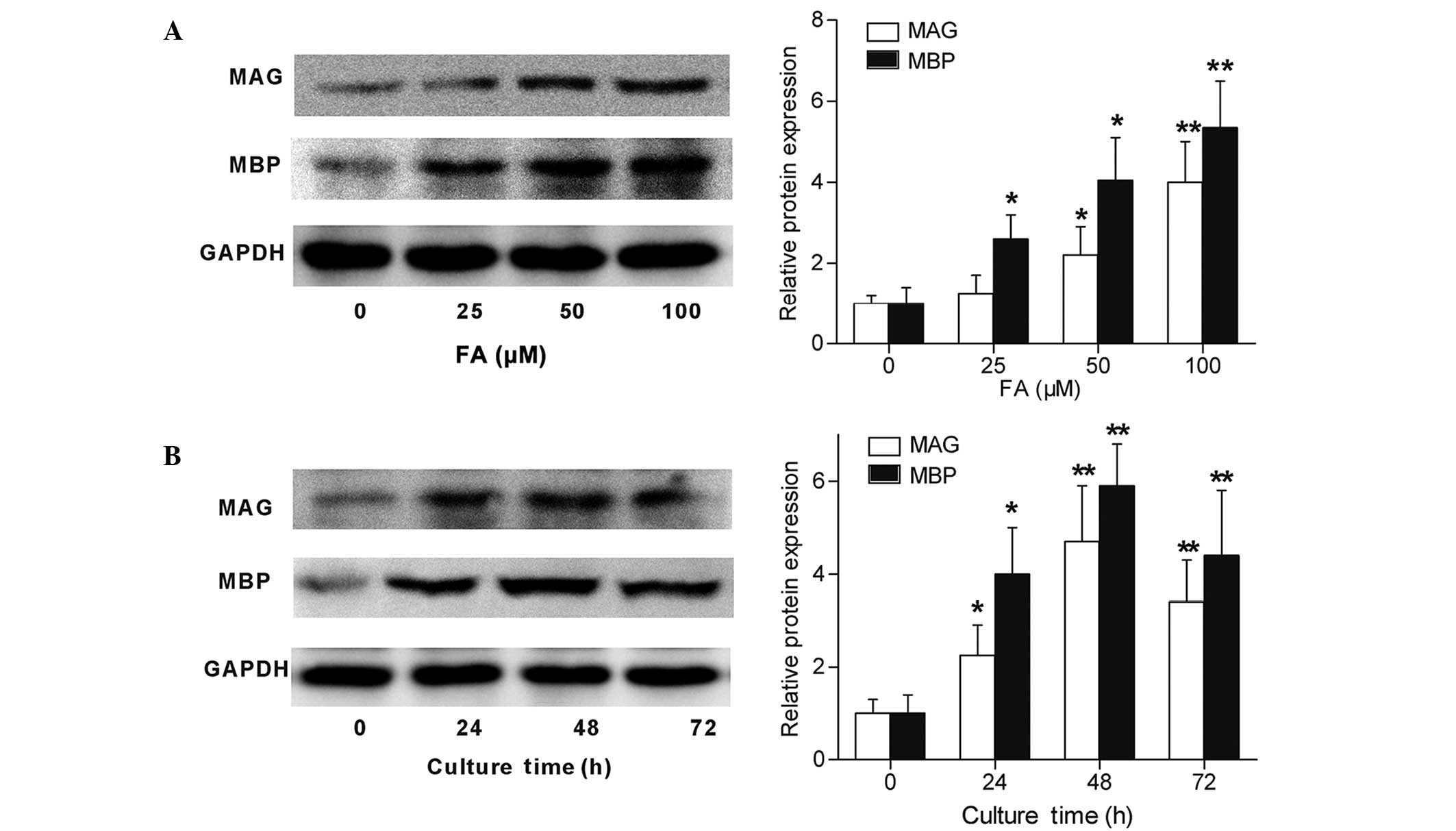
FA increases the mRNA and protein
levels of MAG and MBP expressed by Schwann cells
MAG and MBP are important myelin proteins in
peripheral nerves that are expressed by Schwann cells (18). The present study detected the MAG and
MBP expression levels of cultured Schwann cells following treatment
with FA by RT-qPCR and western blotting. As shown in Fig. 3, Schwann cells directly subjected to
FA treatment showed an increase in their MAG and MBP mRNA
expression levels (Fig. 3A and B).
Furthermore, Schwann cells were subjected to FA at a therapeutic
concentration (50 µΜ) for different time periods, and the results
demonstrated an upregulation of MAG and MBP mRNA expression levels
subsequent to FA exposure for 24, 48 and 72 h (Fig. 3C and D). In accordance with the
RT-qPCR results, the western blotting results also demonstrated
that the protein expression levels of MAG and MBP in Schwann cells
were directly promoted by FA in a concentration-dependent (Fig. 4A) and time-dependent (Fig. 4B) manner. Therefore, these data
revealed that FA, beside its ability to promote Schwann cell
proliferation, was also capable of inducing Schwann cell
differentiation in vitro.
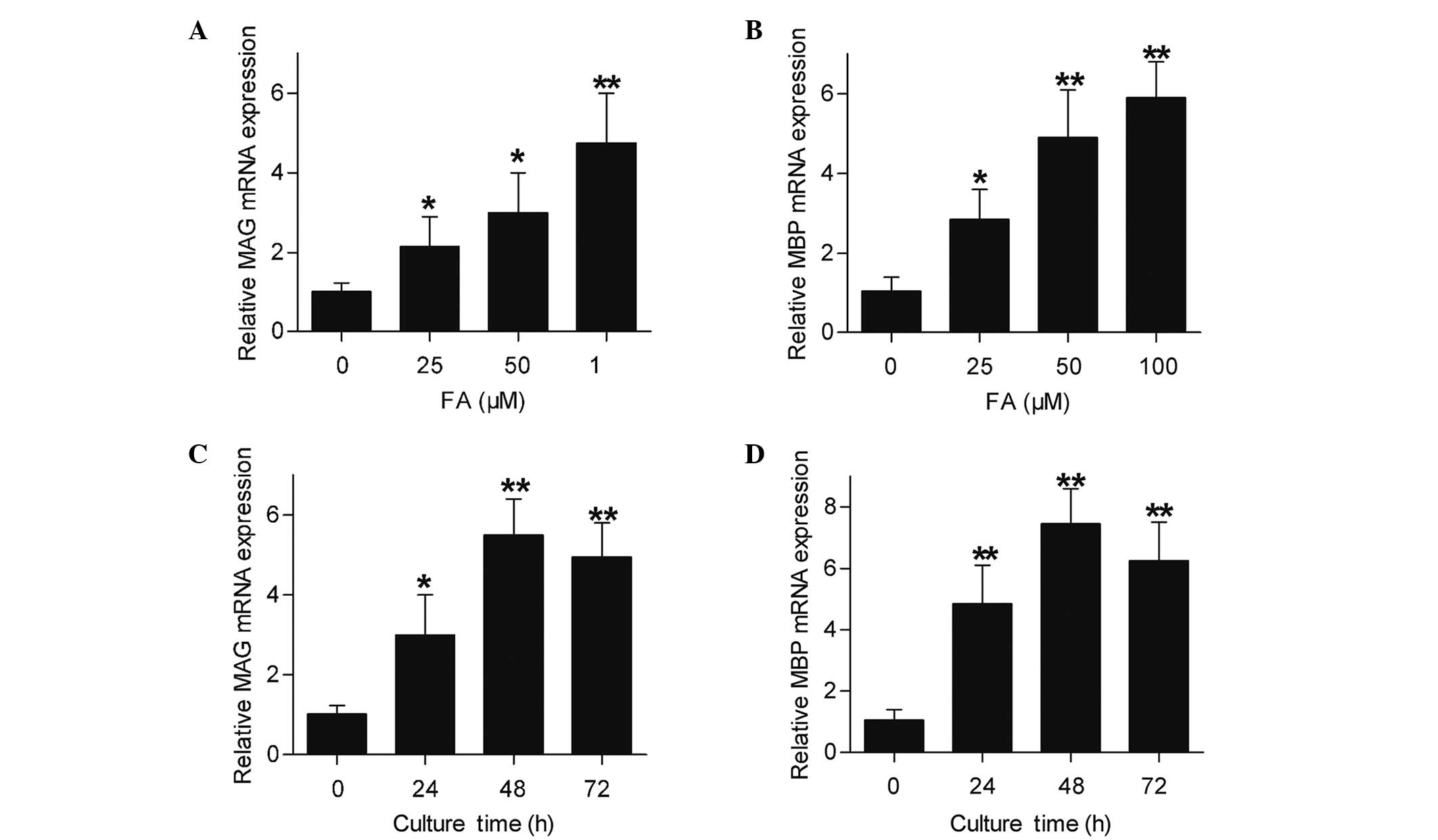 | Figure 3.mRNA levels of MAG and MBP expressed
by Schwann cells were upregulated by FA treatment, as determined
using reverse transcription-quantitative polymerase chain reaction.
(A) MAG and (B) MBP mRNA expression levels in Schwann cells treated
with 0, 25, 50 or 100 µΜ FA for 24 h. (C) MAG and (D) MBP mRNA
expression levels in Schwann cells treated with 50 µΜ FA for 0, 24,
48 or 72 h. β-actin was used as an internal control. Data are shown
as the mean ± standard error of three separate experiments,
*P<0.05 and **P<0.01, vs. 0 µΜ FA or 0 h incubation. MAG,
myelin-associated glycoprotein; MBP, myelin basic protein; FA,
ferulic acid. |
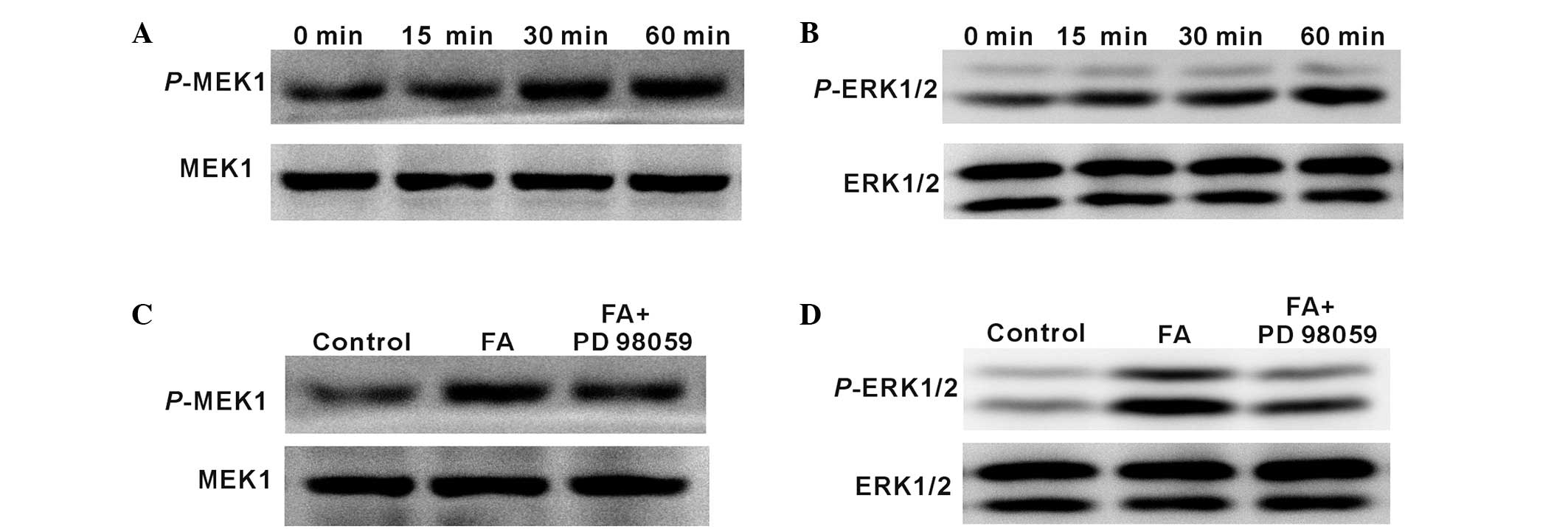
FA induces Schwann cell proliferation
and differentiation through MEK1/ERK1/2 signalling
The critical role of ERK signalling pathway in
Schwann cells has been previously demonstrated (19). Therefore, the present study attempted
to determine whether MEK1/ERK1/2 is also activated in Schwann cells
by FA. Schwann cells treated with 100 µM FA (as a result of
previous results shown in Fig. 2B)
were harvested and analysed by western blotting. As shown in
Fig. 5A and B, p-MEK1 and p-ERK1/2
expression was induced following incubation with FA for 15, 30 and
60 min. Furthermore, when co-cultured with 20 µM PD98059, a
chemical inhibitor of MEK1, p-MEK1 and p-ERK1/2 expression levels
in Schwann cells induced by FA were significantly inhibited
(Fig. 5C and D). These results
indicated that MEK1/ERK1/2 signalling in Schwann cells can be
activated by FA treatment.
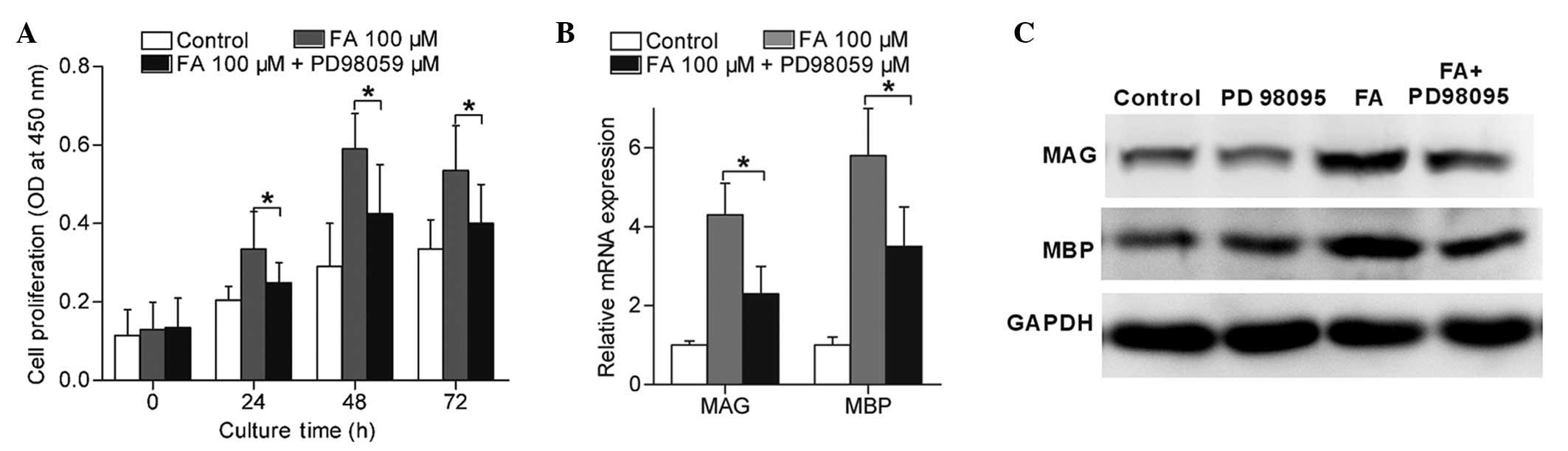
Furthermore, in order to determine the role of
MEK1/ERK1/2 activation in FA-induced Schwann cell differentiation
and proliferation, cells were incubated with 100 µM FA in the
presence of 20 µM PD98059. The results indicated that the
FA-induced Schwann cell proliferation (Fig. 6A) and expression of MAG and MBP
(Fig. 6B and C) were significantly
reduced in the presence of PD98059 (P<0.05). These findings
suggested that FA induced Schwann cell proliferation and
differentiation mainly through the MEK1/ERK1/2 signalling
pathway.
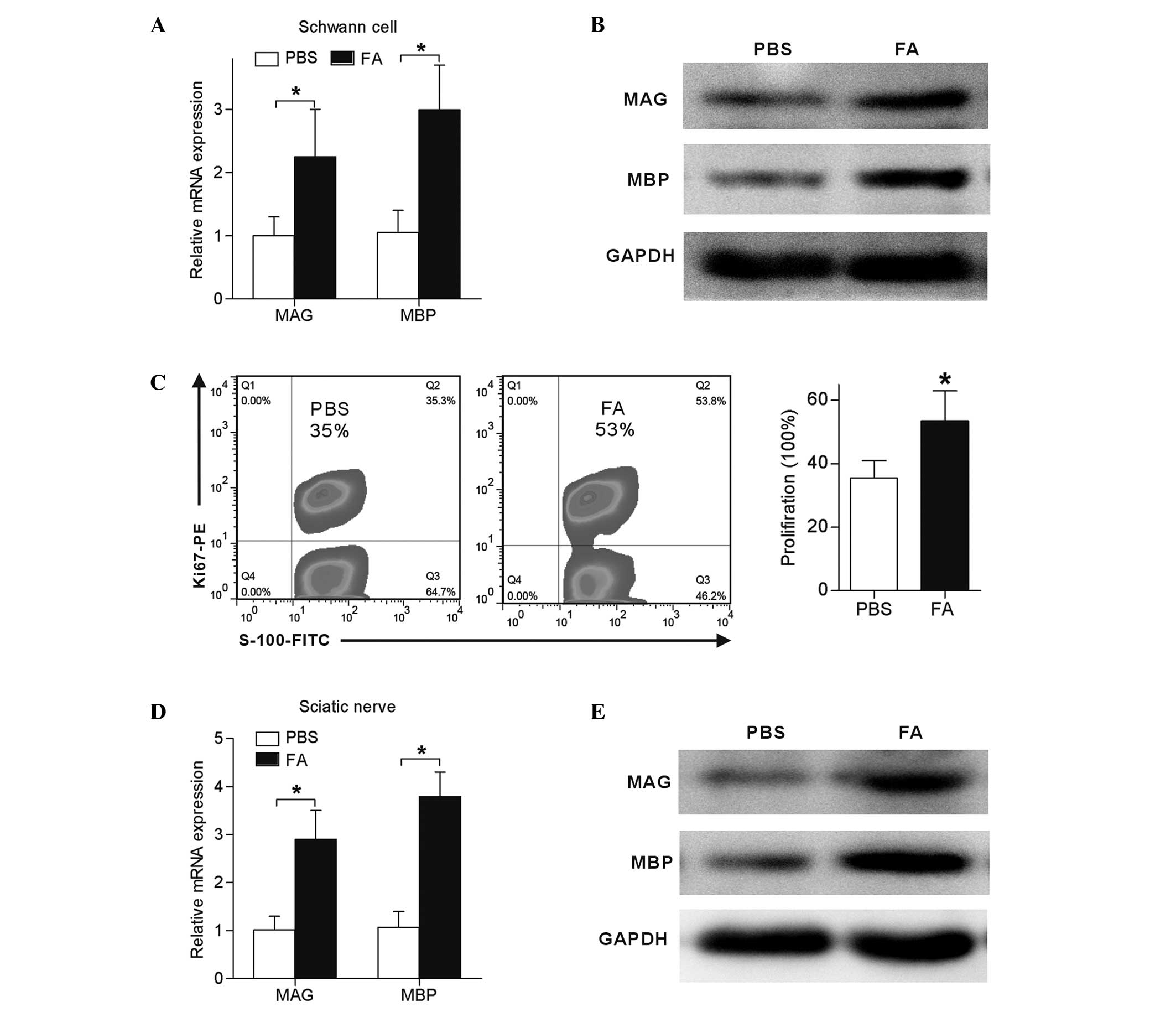
FA treatment promoted rat sciatic
nerve myelination following crush injury
Since proliferation and differentiation of Schwann
cells was induced by FA in vitro, it was suggested that FA
may exert similar nerve remyelination effects in vivo. Rats
with sciatic nerve crush were intraperitoneally treated with FA or
PBS daily for a total of 7 days. The results demonstrates that, at
4 weeks post-injury, Schwann cells isolated from FA-treated nerves
showed 1-fold increase in MAG expression and 2-fold increase in MBP
expression when compared with the expression levels in the PBS
group (P<0.05; Fig. 7A and B).
Furthermore, Ki-67, a cell proliferation marker was used to
determine the proliferation of sciatic nerve-derived Schwann cells.
As shown in Fig. 7C, 35% of Schwann
cells from PBS-treated sciatic nerves were Ki-67-positive, whereas
this proportion was increased to 53% in FA-treated sciatic nerves,
indicating that FA also exerts cell proliferation effects on
Schwann cells in vivo. The expression levels of MAG and MBP
in the sciatic nerve was also detected 4 weeks after crush injury.
Compared with the control, FA treatment induced a 3-fold increase
of MAG and 4-fold increase of MBP expression in sciatic nerves
(P<0.05; Fig. 7D and E).
Discussion
The direct function of FA on the biological
properties of Schwann cells remains largely unknown. Therefore, the
present study investigated the proliferative response of Schwann
cells subsequent to FA exposure. In vitro assays
demonstrated that FA directly induced an increase in Schwann cell
proliferation, which has not been previously reported, to the best
of our knowledge.
While the proliferation of Schwann cells can be
induced by FA, there are currently few data regarding the effect of
FA on Schwann cell differentiation. MAG has been demonstrated to
serve a major role in the formation of myelin, promoting
myelinogenesis and the maintenance of myelin stability through
axo-glial contact in the peripheral nervous system (20). However, during the in vitro
culture of Schwann cells, the biological properties of Schwann
cells have been observed to significantly decline with time,
limiting their application in the construction of artificial nerve
grafts (21). The results of the
current study revealed a marked increase in MAG and MBP expression
levels in FA-treated Schwann cells, suggesting that FA may maintain
and facilitate differentiation properties of cultured Schwann
cells.
The present study also revealed that activation of
the MEK1/ERK1/2 signalling pathway is responsible for the effects
exerted by FA on Schwann cells. A recent study suggested a role for
ERK signalling in promoting myelination due to the finding that
increased ERK activity induces hypermyelination (22). Furthermore, conditional ERK knockout
results have shown that ERK signalling is required for
differentiation and myelination of Schwann cells (23). In the present study, western blot
analysis demonstrated that the protein levels of phosphorylated
MEK1/ERK1/2 in Schwann cells were evidently increased following FA
treatment. In addition, PD98059, an inhibitor of MEK1, was able to
suppress the activation of MEK1/ERK1/2 signalling evoked by FA
treatment. Further results demonstrated that, when the MEK1/ERK1/2
transduction pathway was blocked with PD98059, the Schwann cell
proliferation and differentiation induced by FA was significantly
declined. Therefore, the present results are consistent with the
findings of previous studies (22,23), and
these data suggest that activation of MEK1/ERK1/2 signalling is
responsible for FA-induced cell proliferation and differentiation
of Schwann cells.
In conclusion, the present study demonstrated that
FA treatment was able to promote cell proliferation and expression
of MAG and MBP in Schwann cells through the activation of
MEK1/ERK1/2 signalling. In addition, FA was able to accelerate the
remyelination of peripheral injured nerves by increasing the
proliferating numbers of Schwann cells and promoting myelin
secretion. These findings provide novel information on the direct
effects of FA on Schwann cells and assist in developing further
understanding of the neuroprotective effects produced by FA.
References
|
1
|
Hung HA, Sun G, Keles S and Svaren J:
Dynamic regulation of Schwann cell enhancers after peripheral nerve
injury. J Biol Chem. 290:6937–6950. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chew SY, Mi R, Hoke A and Leong KW: The
effect of the alignment of electrospun fibrous scaffolds on Schwann
cell maturation. Biomaterials. 29:653–661. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ishikawa N, Suzuki Y, Dezawa M, Kataoka K,
Ohta M, Cho H and Ide C: Peripheral nerve regeneration by
transplantation of BMSC-derived Schwann cells as chitosan gel
sponge scaffolds. J Biomed Mater Res A. 89:1118–1124. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Triolo D, Dina G, Lorenzetti I, Malaguti
M, Morana P, Del Carro U, Comi G, Messing A, Quattrini A and
Previtali SC: Loss of glial fibrillary acidic protein (GFAP)
impairs Schwann cell proliferation and delays nerve regeneration
after damage. J Cell Sci. 119:3981–3993. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Heinen A, Beyer F, Tzekova N, Hartung HP
and Küry P: Fingolimod induces the transition to a nerve
regeneration promoting Schwann cell phenotype. Exp Neurol.
271:25–35. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nishimoto S, Tanaka H, Okamoto M, Okada K,
Murase T and Yoshikawa H: Methylcobalamin promotes the
differentiation of Schwann cells and remyelination in
lysophosphatidylcholine-induced demyelination of the rat sciatic
nerve. Front Cell Neurosci. 9:2982014.
|
|
7
|
Zhao Z and Moghadasian MH: Chemistry,
natural sources, dietary intake and pharmacokinetic properties of
ferulic acid: A review. Food Chem. 109:691–702. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen XH, Lin ZZ, Liu AM, Ye JT, Luo Y, Luo
YY, Mao XX, Liu PQ and Pi RB: The orally combined neuroprotective
effects of sodium ferulate and borneol against transient global
ischaemia in C57 BL/6J mice. J Pharm Pharmacol. 62:915–923. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cheng CY, Su SY, Tang NY, Ho TY, Chiang SY
and Hsieh CL: Ferulic acid provides neuroprotection against
oxidative stress-related apoptosis after cerebral
ischemia/reperfusion injury by inhibiting ICAM-1 mRNA expression in
rats. Brain Res. 1209:136–150. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yabe T, Hirahara H, Harada N, Ito N, Nagai
T, Sanagi T and Yamada H: Ferulic acid induces neural progenitor
cell proliferation in vitro and in vivo. Neuroscience. 165:515–524.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jiang M, Zhuge X, Yang Y, Gu X and Ding F:
The promotion of peripheral nerve regeneration by
chitooligosaccharides in the rat nerve crush injury model. Neurosci
Lett. 454:239–243. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang H, Liu S, Tian Y, Wu X, He Y, Li C,
Namaka M, Kong J, Li H and Xiao L: Quetiapine inhibits microglial
activation by neutralizing abnormal STIM1-mediated intercellular
calcium homeostasis and promotes myelin repair in a
cuprizone-induced mouse model of demyelination. Front Cell
Neurosci. 9:4922015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fei W, Aixi Y, Danmou X, Wusheng K,
Zhengren P and Ting R: The mood stabilizer valproic acid induces
proliferation and myelination of rat Schwann cells. Neurosci Res.
70:383–390. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
He B, Tao HY and Liu SQ: Neuroprotective
effects of carboxymethylated chitosan on hydrogen peroxide induced
apoptosis in Schwann cells. Eur J Pharmacol. 740:127–134. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sasagasako N, Toda K, Hollis M and Quarles
RH: Myelin gene expression in immortalized Schwann cells:
relationship to cell density and proliferation. J Neurochem.
66:1432–1439. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ju DT, Kuo WW, Ho TJ, Paul CR, Kuo CH,
Viswanadha VP, Lin CC, Chen YS, Chang YM and Huang CY:
Protocatechuic acid from alpinia oxyphylla induces schwann cell
migration via ERK1/2, JNK and p38 activation. Am J Chin Med.
43:653–665. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Varejão AS, Cabrita AM, Meek MF,
Bulas-Cruz J, Melo-Pinto P, Raimondo S, Geuna S and
Giacobini-Robecchi MG: Functional and morphological assessment of a
standardized rat sciatic nerve crush injury with a non-serrated
clamp. J Neurotraum. 21:1652–1670. 2004. View Article : Google Scholar
|
|
18
|
Gupta R, Truong L, Bear D, Chafik D,
Modafferi E and Hung CT: Shear stress alters the expression of
myelin-associated glycoprotein (MAG) and myelin basic protein (MBP)
in Schwann cells. J Orthop Res. 23:1232–1239. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Napoli I, Noon LA, Ribeiro S, Kerai AP,
Parrinello S, Rosenberg LH, Collins MJ, Harrisingh MC, White IJ,
Woodhoo A and Lloyd AC: A central role for the ERK-signaling
pathway in controlling schwann cell plasticity and peripheral nerve
regeneration in vivo. Neuron. 73:729–742. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schnaar RL and Lopez PH: Myelin-associated
glycoprotein and its axonal receptors. J Neurosci Res.
87:3267–3276. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Painter MW, Brosius Lutz A, Cheng YC,
Latremoliere A, Duong K, Miller CM, Posada S, Cobos EJ, Zhang AX,
et al: Diminished Schwann cell repair responses underlie
age-associated impaired axonal regeneration. Neuron. 83:331–343.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ishii A, Furusho M and Bansal R: Sustained
activation of ERK1/2 MAPK in oligodendrocytes and schwann cells
enhances myelin growth and stimulates oligodendrocyte progenitor
expansion. J Neurosci. 33:175–186. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Newbern JM, Li XY, Shoemaker SE, Zhou J,
Zhong J, Wu Y, Bonder D, Hollenback S, Coppola G, Geschwind DH, et
al: Specific functions for ERK/MAPK signaling during PNS
development. Neuron. 69:91–105. 2011. View Article : Google Scholar : PubMed/NCBI
|