Introduction
The treatment of implant-related infections has been
a concern for orthopedists for a long period of time (1). The new antibacterial agent, an
endogenous antibiotic peptide known as human β-defensin-3 (HBD-3),
is important in the inhibition of biofilm formation and immune
regulation (2). In a previous study,
it was found that the effect of HBD-3 in killing drug-resistant
Staphylococcus was significantly enhanced inside
biomembranes (3). Additionally,
HBD-3 inhibited the expression of the film-forming gene
icaAD, and of the drug-resistant gene mecA (4). The supernatant of
Staphylococcus aureus can induce stimulated osteoblasts of
rats to secrete a structural and functional congener of HBD-3, the
murine β-defensin 14 (MBD-14) through activation of the p38
mitogen-activated protein kinase (MAPK) (5). Furthermore, in acute suppurative
osteomyelitis models infected with drug-resistant
Staphylococcus aureus, the application of a p38 MAPK agonist
promoted the increased release of MBD-14 in the infection site and
inhibited osteomyelitis (6). These
results strongly suggested that HBD-3 is useful in the prevention
and treatment of implant-related infections in orthopedics.
However, the mechanisms that lead HBD-3 to induce
and regulate immune responses and to inhibit bone infections remain
unknown. In the present study, the effects of HBD-3 on the
lipopolysaccharide/Toll-like receptor-4 (LPS/TLR4)- mediated
signaling pathway and on the maturation process of dendritic cells
were determined and analyzed to clarify the mechanisms of action of
the antimicrobial peptide.
Materials and methods
Soluble expression, purification and
identification of TLR4 extracellular segments in Escherichia coli
(E. coli)
Plasmid was constructed to determine the protein
expression. The pCDNA3-TLR4 plasmid was purified from E.
coli using a plasmid mini kit (Hangzhou Vitagene Biochemical,
Hangzhou, China) according to the manufacturer's instructions.
Polymerase chain reaction (PCR) was used to amplify a fragment from
the purified plasmid that contains a human TLR4 template, using the
forward primer, 5′-CGGGATCCATGATGTCTGCC-3′ (containing a
BamHI restriction site) and reverse primer,
5′-GCAAGCTTCTTATTCATCTGACAGG-3′ (containing a HindIII
restriction site) (Shanghai Boya Biotechnology Co., Ltd., Beijing,
China). The PCR conditions used were: pre-denaturation at 94°C for
4 min, 30 cycles of denaturation at 94°C for 40 sec, followed by
annealing at 53°C for 50 sec, and extension at 72°C for 1 min, and
a final extension at 72°C for 10 min. PCR products were extracted
according to the instructions of the ‘DNA rapid
purification/recycling kit’, and the purified products were
preserved at −20°C.
PCR products and pET32a were subjected to a
double-enzyme digestion by BamHI and HindIII. After
extraction of the appropriate fragments, a ligation was set up
using T4 DNA ligase. Ligations were used to transform E.
coli. and were placed onto LB medium containing ampicillin.
After plasmid purification from single colonies and enzyme
digestion to verify the restriction pattern of the constructs,
sequencing was used to select the right recombinant expression
vector pET32a-TLR4, containing the TLR4 segment fused to a
thioredoxin tag. BL21 (DE3) plysS competence bacteria were
transformed with the purified vector and selected in LB medium with
ampicillin following incubation at 37°C overnight. For the
expression, a new culture was created into the same 1% of the total
type of medium using starter culture, at 37°C in an oscillating
incubator. When the optical density reached A600
nm=0.6–1.0, IPTG was added to a final concentration of 1 mmol/l and
the culture was induced for an additional 4 hours. The induced
cultures were centrifuged, the pellets were resuspended into buffer
solution A (10 mmol/l imidazole, 300 mmol/l NaCl and 50 mmol/l
NaH2PO4, pH 8.0), ultrasonication was
performed and the resulting suspensions were then centrifuged at
3,000 × g for 10 min at 4°C. The supernatants containing the
soluble protein fractions were saved.
Protein purification
Soluble protein fractions were allowed to pass
through an Ni2+-NTA column (Qiagen, Valencia, CA, USA)
that was previously balanced with buffer solution A by gravity
flow. Buffer solution B (20 mmol/l imidazole, 300 mmol/l NaCl and
50 mmol/l NaH2PO4, pH 8.0) was then used to
rinse the column. The target protein was eluted from the column
with buffer solution C (250 mmol/l imidazole, 300 mmol/l NaCl and
50 mmol/l NaH2PO4, pH 8.0). Subsequently, the
target protein was ultrafiltrated and concentrated, and fully
dialyzed in 50 mmol/l phosphate-buffered saline (PBS) (pH 7.2) to
clear away the imidazole and salt contents. The final product was
the purified TLR4-Trx fusion protein.
Identification of TLR4-Trx protein
using western blot analysis
Purified TLR4-Trx protein (300 µg) was separated
with sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE), and transferred to a nitrocellulose filter. The filter
was blocked with 30 g/l dried skimmed milk at 4°C overnight. The
following morning, HRP-anti His antibody (rabbit anti-mouse
monoclonal antibody, catalog no. 201321) was diluted in blocking
buffer (1:1,000; Nanjing Jingmei Chemical Co., Ltd., Nanjing,
China) and incubated with the nitrocellulose membrane at 37°C for 2
h. TBST was used to wash the membrane 5 times for 10 min; and color
was developed using a DAB color development kit for the appropriate
amount of time. The PVDF membrane was dipped into distilled liquid
to terminate color development and air-dried. Images were captured
to confirm the size of the target protein. The protein fraction
from BL21 (DE3)/PET32a was used as a negative control.
Preliminary study of the regulation
mechanism of HBD-3 on dendritic cells
HBD-3 was diluted to 1% in 0.025 mol/l carbonate
buffer at pH 9.0, and placed into a dialysis bag. Fluorescein
isothiocyanate (FITC) was prepared as a 0.1 mg/ml solution in the
carbonate buffer solution, and used as a diluent in a 10 mg/ml
globulin solution. The dialysis bag was immersed into the globulin
FITC solution containing 10-fold its volume. Dialysis occurred at
4°C on a stirring plate with a magnet for 2 h. The dyalized
contents of the bag were immediately filtered using a Sephadex G50
gel to remove the free fluorescein and aliquots were packed and
stored at 4°C. The resulting F/P ratio of the stained FITC-HBD-3
was 2.
In vitro immature dendritic cell
(imDC) induction
Peripheral blood from healthy adult volunteers was
processed with heparin as an anticoagulation agent. Mononuclear
cells (MNCs) were isolated via density gradient centrifugation
(Ficoll). PBS was used to wash away platelets, MNCs were allowed to
adhere to the vessel wall for 3 h at 37°C and the supernatant was
discarded. The remaining non-adherent cells were gently rinsed with
pre-warmed (37°C) RPMI-1640 (Gibco, Grand Island, NY, USA).
Adherent MNCs were scraped off the wall, and suspended in a
1×109 cells/l solution in RPMI-1640 supplemented with
100 ml/l new-born calf serum (Hangzhou Sijiqing Biological
Engineering Material Co., Ltd., Hangzhou, China). Susequently, 100
µg/l rhGM-CSF and 100 µg/l rhIL-4 (both from Peprotech, Inc., Rocky
Hill, NJ, USA) were added into the MNC suspension and the cells
were cultured in an incubator at 37°C. Half the amount of the
medium was exchanged every 3 days and supplementing cytokines were
added when necessary. Cell morphology was observed under a confocal
microscope. After seven days, cells were collected, the phenotype
and function of imDC were analyzed, and the supernatant of each
group collected for measuring the cytokines present in the
supernatant.
Cell binding tests
HBD-3 was bound to imDC. Briefly, imDCs were
suspended in buffer solution A (5 mmol/l Ca2+) and the
cell density was adjusted to 5×109/l. FITC-HBD-3 was
then added to 200 µl of cell suspension to a final concentration of
15 mg/l, and the reaction was allowed to proceed for 30 min at 37°C
in the dark. The potential inhibitor sTLR4 was added to another
group of cells to a final concentration of 30 mg/l, and FITC-HBD-3
was added 10 min later. The reaction was allowed to proceed for 30
min at 37°C in the dark. The cells were then washed 3 times using
PBS prior to flow cytometry (FCM). Unlabelled FITC-HBD-3 was used
as a negative control.
To identify HBD-3 as a competitive inhibitor of the
binding between LPS and imDC, imDCs were resuspended in buffer
solution A to an adjusted cell density of 5×109/l. HBD-3
(15 mg/l) was added into 200 µl cell suspension for the reaction. A
group of cells with no added HBD-3 served as a negative control.
The specificity of HBD-3 was assessed using anti-HBD-3 pAb. The
reaction was allowed to proceed for 30 min at 37°C in the dark, and
then PBS was used to wash the cells 3 times prior to FCM
analysis.
Western blot analysis
Western blot analysis of the binding of HBD-3 and
sTLR4 was performed. Briefly, 2 µg sTLR4 extracellular proteins
were processed by 2-mercaptoethanol loading buffer. After
separation, a 13% SDS-PAGE was stained by CBB R250. The proteins
were then transferred to a PVDF membrane at 60 V for 3 h under
constant pressure. The PVDF membrane was immersed into buffer
solution B for 2 h, followed by buffer solution B containing 5
µg/ml HBD-3, for 3 h at room temperature. The membrane was washed 5
times (1:2,000). Mouse anti-human HBD-3 mAb HYP131-11 (R&D
Systems, Inc., Minneapolis, MN, USA) was added to the washed
membrane, and the binding was allowed to proceed for 60 min at room
temperature. The 1:2,000 sheep anti-rat IgG (Sigma, St. Louis, MO,
USA) was incubated with the membrane for 60 min at room
temperature. The membrane was then washed and color was developed
using a DAB color development kit prior to capturing the image.
Nuclear proteins were extracted strictly in
accordance with the instruction of a kit (Pierce Biotechnology,
Inc., Rockford, IL, USA) and the concentration of the extraction
was measured using the Bradford method. The activity of nuclear
factor-κB (NF-κB) was detected using western blot analysis. Nuclear
protein extracts (5 µg) were separated with 12% SDS-PAGE. The
proteins were then transferred to a PVDF membrane for 3 h. The
transferred membrane was blocked in PBS-T that contained 3% dried
skimmed milk for 2 h. Subsequently, 1:1,500 HRP-sheep anti-rat IgG
was added and the solution was incubated with the membrane for 60
min at room temperature. The membrane was then washed as described
above, color was developed using a DAB color kit and images were
captured.
Statistical analysis
SPSS 19.0 software was used for statistical
analysis. Quantitative data were presented as mean ± standard
deviation. The Student's t-test was used in group comparisons.
Enumeration data were presented as a percentage (%) and the
χ2 test was applied in group comparisons. P<0.05 was
considered statistically significant.
Results
Soluble expression of TLR4
extracellular fragment in E. coli, purification and
identification
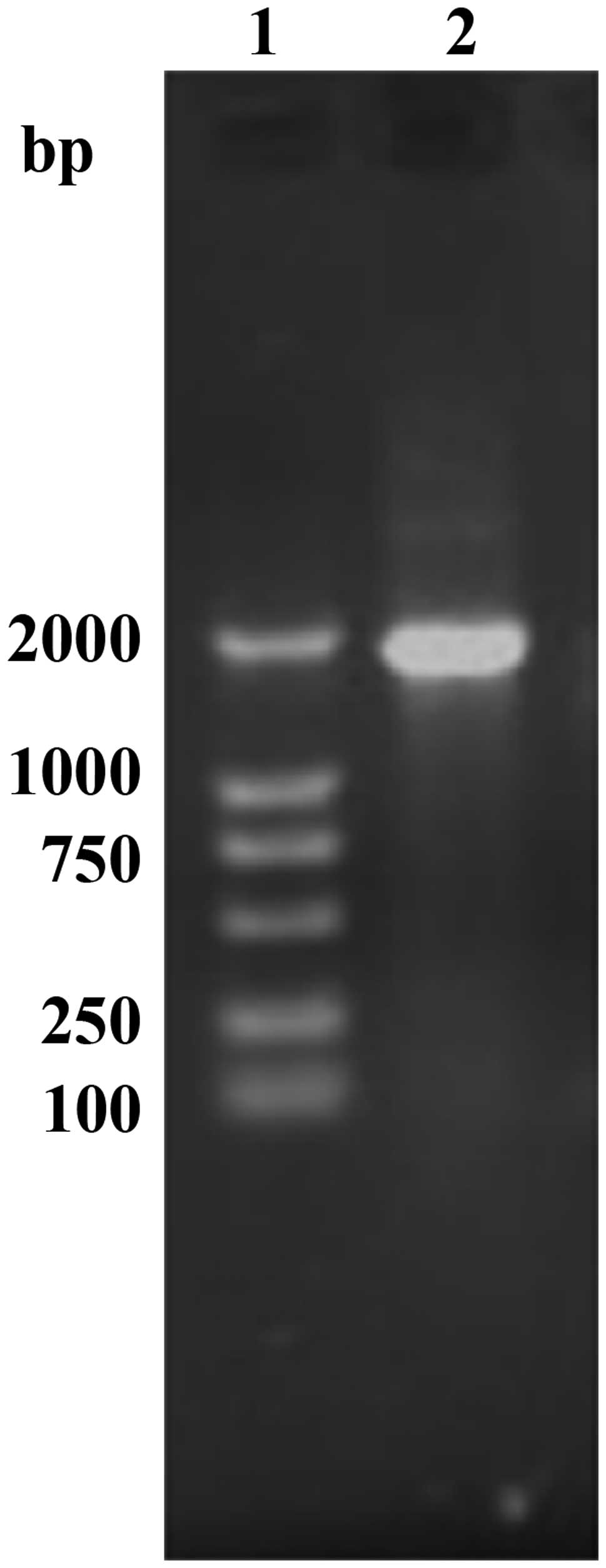
A single amplicon of the expected size (1,911 bp)
was detected after gel electrophoresis and ethidium bromide
staining (Fig. 1). Restriction
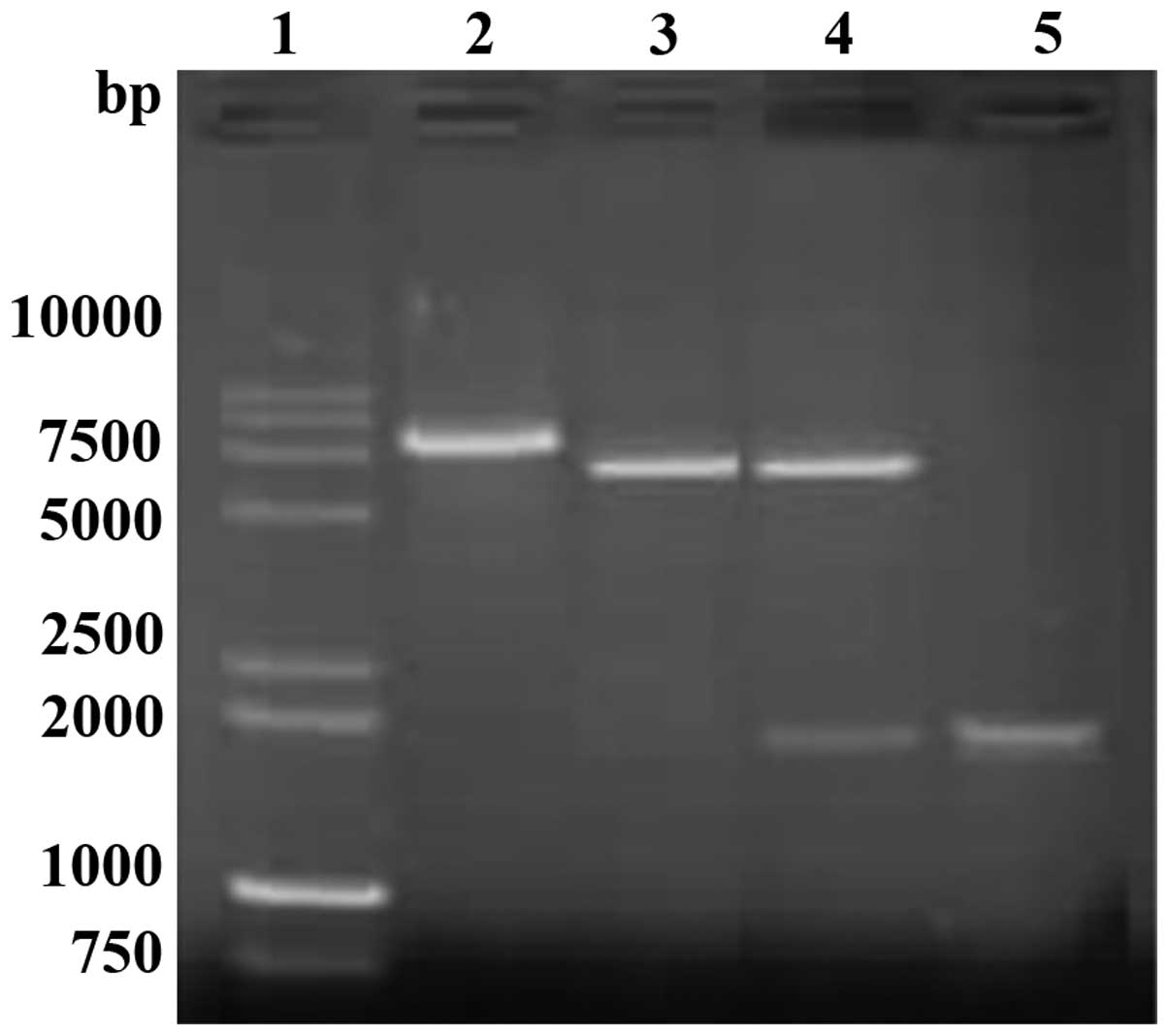
analysis of vector pET32a-TLR with HindIII and BamHI
enzyme digestions showed the expected pattern (5,900 bp for the
vector fragment and 1,911 bp for the amplicon) (Fig. 2). Sequencing results confirmed the
correct exogenous DNA insert pET32a-TLR.
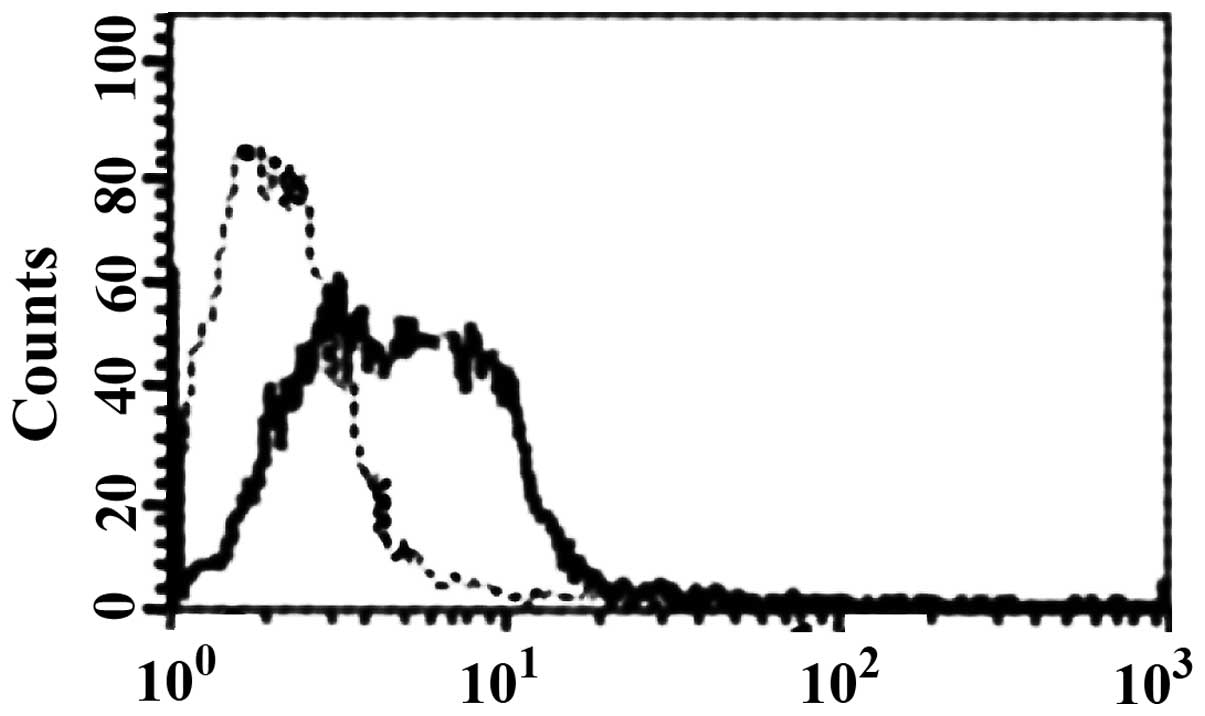
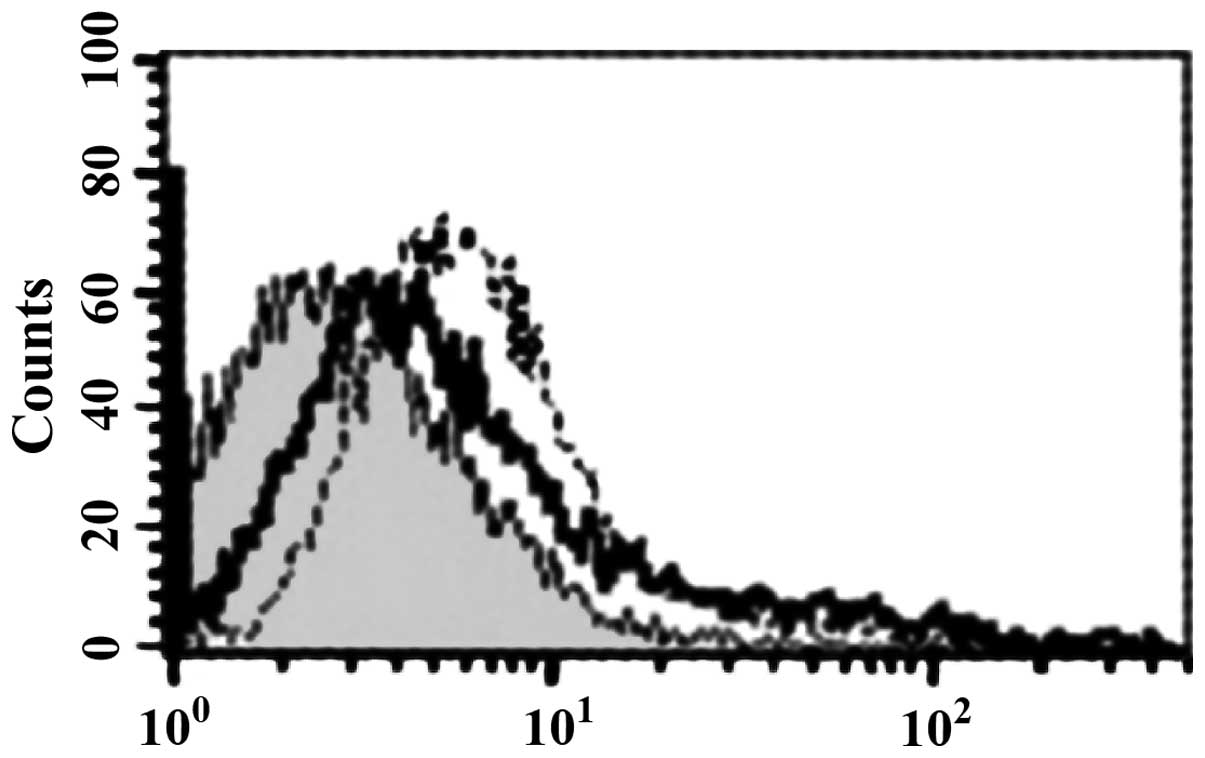
Binding of HBD-3 and imDC
FCM analysis showed that HBD-3 can directly bind to
imDC in 5 mmol/l binding buffer (Fig.
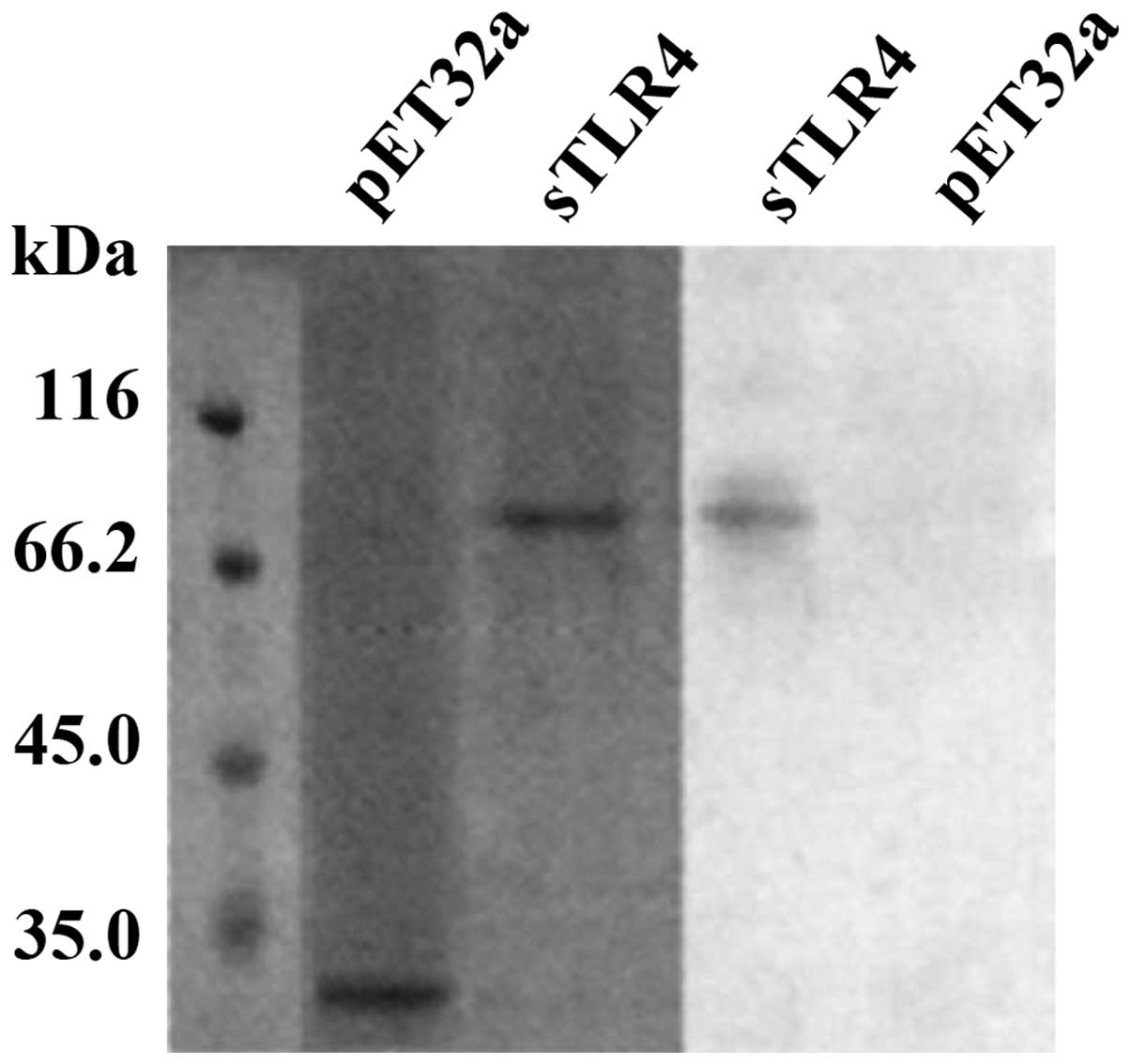
3). Western blot analysis showed that HBD-3 can bind to sTLR4
protein (Fig. 4).
Competitive inhibition tests
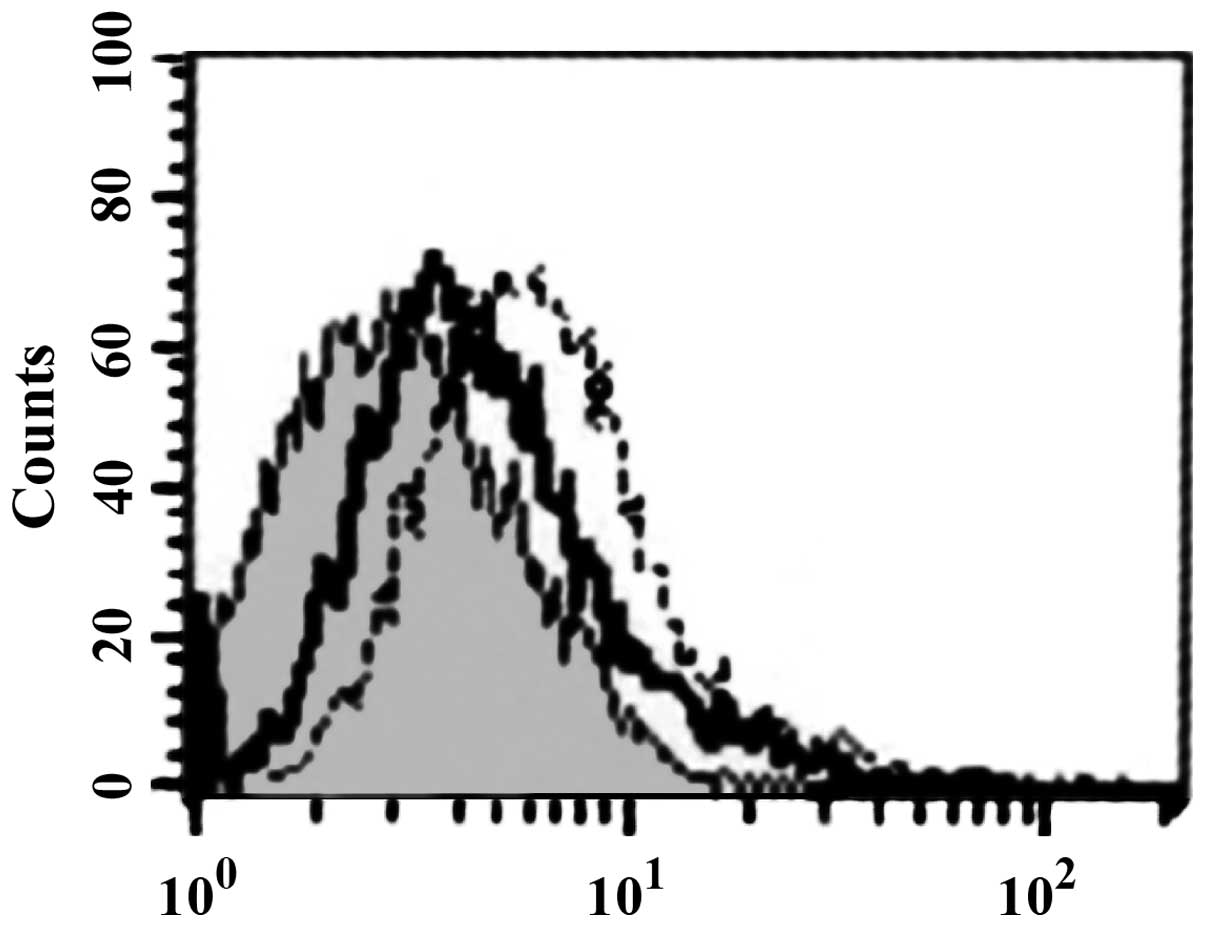
sTLR4 extracellular protein can partially inhibit
the binding of HBD-3 and imDC, indicating that the binding site of
HBD-3 and imDC is associated with TLR4 (Fig. 5). LPS can competitively inhibit the
binding of HBD-3 and imDC, indicating that HBD-3 can bind TLR4,
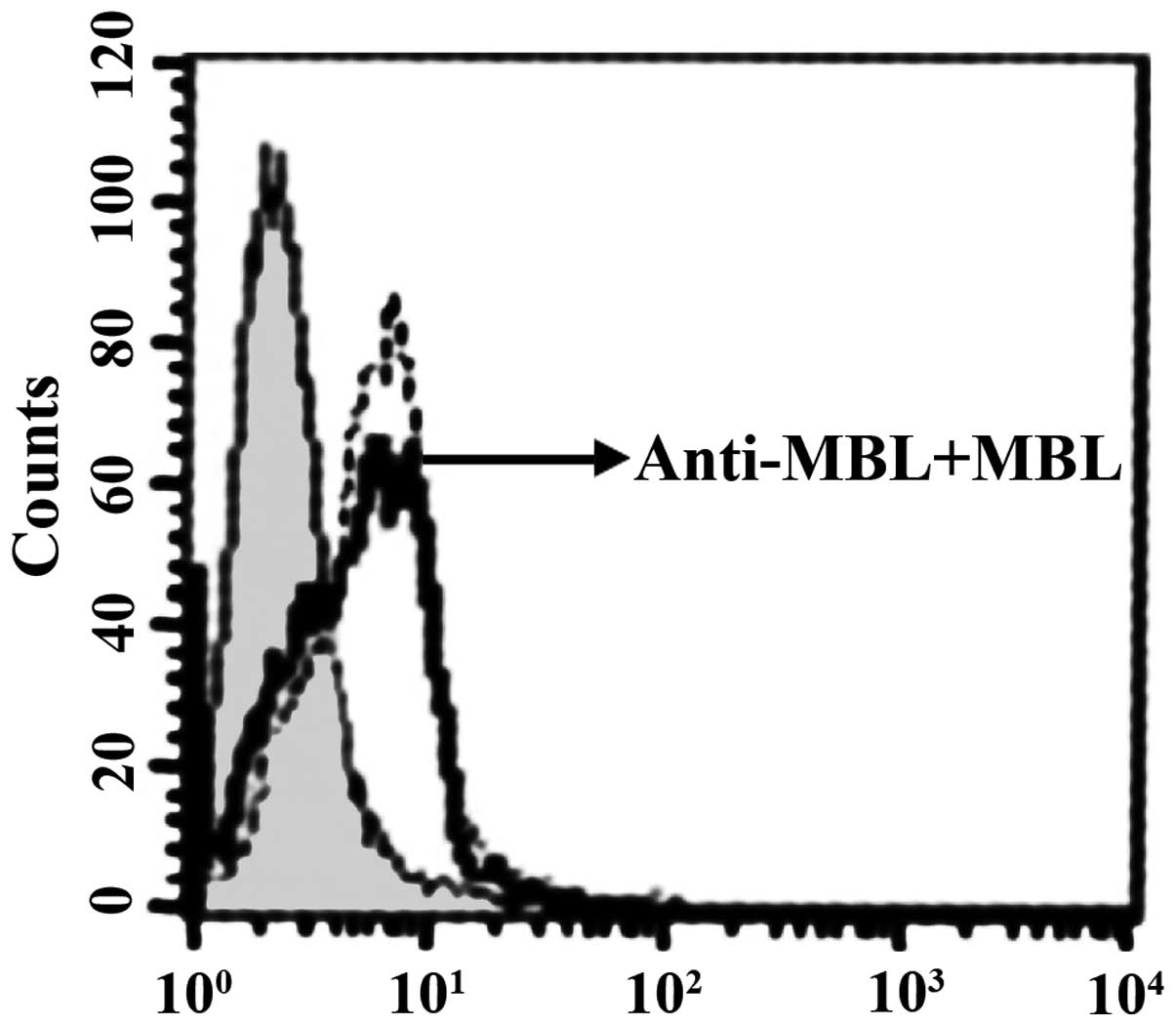
which is found on the surface of imDC (Fig. 6). After the addition of anti-HBD-3
pAb, the inhibitory effect of HBD-3 disappeared, indicating that
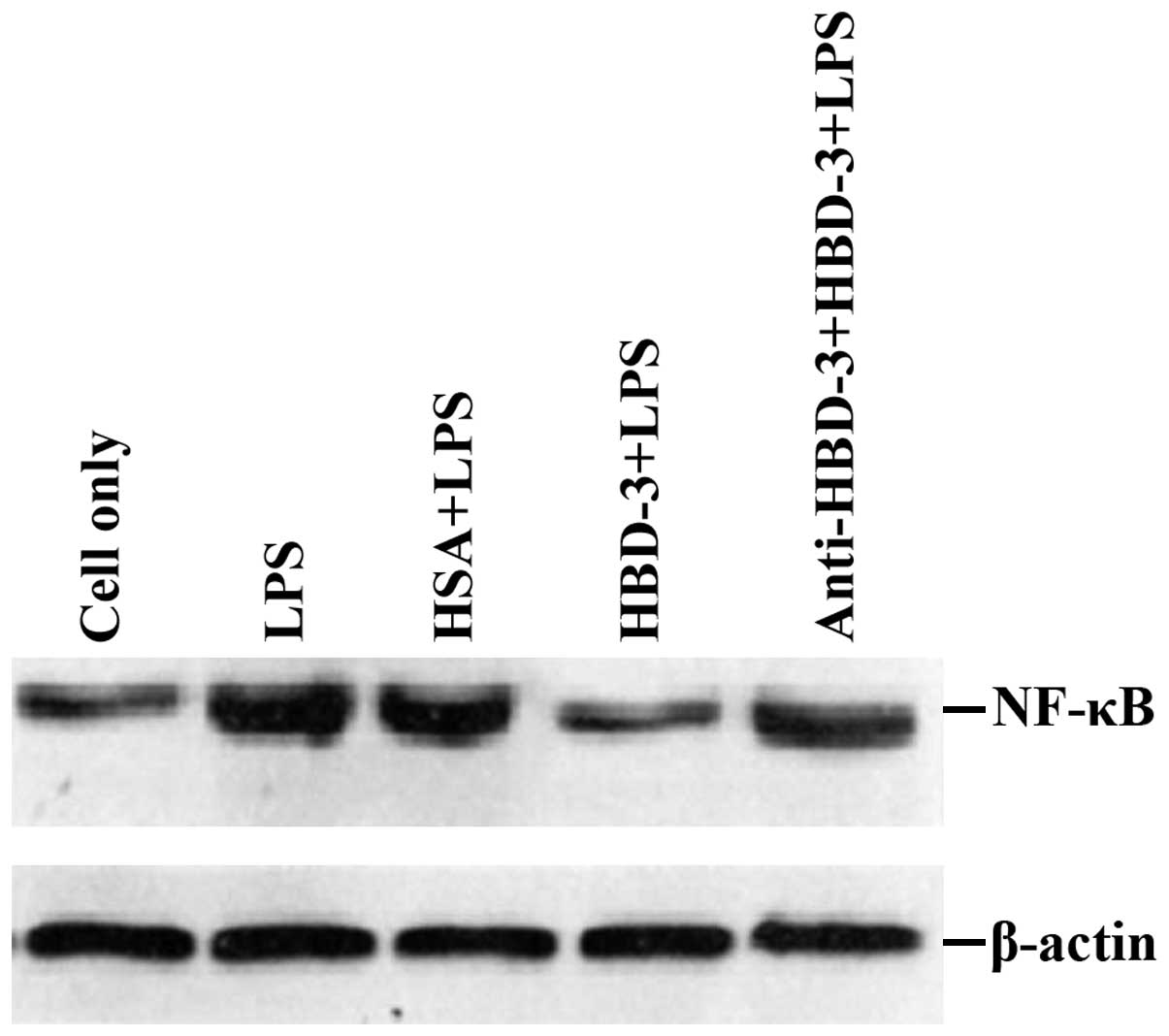
the inhibition of HBD-3 had specificity (Fig. 7). The western blot analysis revealed
that LPS stimulation can significantly increase the nuclear
translocation of NF-κB, which is the nuclear transcription factor
of imDCs, although the translocation was significantly decreased by
high concentrations of HBD-3. After, the addition of anti-HBD-3
pAb, the inhibitory effect of HBD-3 disappeared, indicating that
the inhibition of HBD-3 is specific (Fig. 8).
Discussion
Previous findings revealed that bacterial infection
of osteoblasts can stimulate the secretion of cytokines,
chemokines, and endogenous antimicrobial peptide β-defensin
(7). Thus, osteoblasts regulate bone
metabolism and play a role in the regulation of an active immune
defense response and acquired immunity. β-defensin, a new type of
bacterial infection inhibitor characterized by its wide antibiotic
spectrum, rapid action, low drug resistance rate, innate immune
compatibility and its unique role in the immune regulation of bone
infection, has drawn attention in clinical and basic research
settings (8).
Shi et al (9)
demonstrated that Staphylococcus aureus- or Candida
albicans-contaminating maxillofacial silicone elastomers were
significantly inhibited or killed by immersing them into an HBD-3
solution for only 30 min. Other authors found that
Staphylococcus aureus supernatants can induce the release
and expression of HBD-3 through TLR2 and TRL4 on the surface of
osteoblasts (10). Furthermore, the
expression of HBD-3 in bone tissues of acute infection sites and on
the boundary tissues of periprosthetic joint infections was
doubled, indicating that endogenous antimicrobial peptide HBD-3 is
a first defense line after bacterial invasion into bone tissues.
In vitro HBD-3 inhibits TLR4-mediated myeloid
differentiation factor 88 (MyD88) and TIR structural domain adaptor
molecule (TRIF) pathways, and inhibits the activation of NF-κB
(11). In vitro and in
vivo HBD-3 has been shown to inhibit the LPS stimulation of
macrophages to secrete tumor necrosis factor-α (TNF-α) and
interleukin-6 (IL-6). Therefore, it was postulated that HBD-3 is,
not only an inflammatory chemokine, but also a suppressant
immunomodulator, with balancing and bidirectional adjusting effects
in antisepsis and anti-inflammation. Another study confirmed that
in an immune reaction, the structural and functional congener of
HBD-3, MBD-14, initially confronts a microbial invasion through the
innate immune response and then promotes the extensive chemotaxis
of MNC (Mo), macrophages (MФ), neutrophils, imDC and T lymphocytes
in a chemokine receptor 6 (CCR6)- and CCR2 receptor-dependent
manner (12). HBD-3 can also
activate the MAPK extracellular signal-regulated kinase (ERK)
signaling pathway, MФ and antigen-presenting cells (APCs) through
NF-κB-dependent TLR-1 and TLR-2 receptor pathways (13,14).
TLR is an important pattern recognition receptor in
the natural immune system, which can, not only mediate the
identification of pathogenic microorganisms and their products, but
also participate in the acquired immune response. It has been
recognized as a bridge that binds natural immunity with acquired
immunity (15). TLR is extensively
expressed in imDC, MФ, and Mo, and plays an important role in
identifying gram-positive teichoic acid, heat shock protein 60, LPS
and the signaling pathways mediated by LPS (16). Based on those and our own findings,
we conclude that HBD-3 binds to imDC in a Ca2+-dependent
manner, and that sTLR4 and LPS can competitively inhibit binding.
HBD-3 can competitively inhibit the binding of LPS and imDC by
binding with imDC. HBD-3 significantly inhibits the translocation
of LPS-induced NF-κB into the nucleus. Thus, HBD-3 can
competitively inhibit the binding of LPS and imDC by binding with
TLR4 molecules expressed in imDCs, thereby preventing LPS from
inducing the maturity of imDC.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81401815), the China
Postdoctoral Science Foundation (grant no. 2015M582900) and the
Jiangsu Postdoctoral Science Foundation (grant no. 1501146C).
References
|
1
|
Montanaro L, Speziale P, Campoccia D,
Ravaioli S, Cangini I, Pietrocola G, Giannini S and Arciola CR:
Scenery of Staphylococcus implant infections in orthopedics. Future
Microbiol. 6:1329–1349. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhu C, Tan H, Cheng T, Shen H, Shao J, Guo
Y, Shi S and Zhang X: Human β-defensin 3 inhibits
antibiotic-resistant Staphylococcus biofilm formation. J Surg Res.
183:204–213. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ning R, Zhang X, Guo X and Li Q:
Staphylococcus aureus regulates secretion of interleukin-6 and
monocyte chemoattractant protein-1 through activation of nuclear
factor kappaB signaling pathway in human osteoblasts. Braz J Infect
Dis. 15:189–194. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jarczak J, Kościuczuk EM, Lisowski P,
Strzałkowska N, Jóźwik A, Horbańczuk J, Krzyżewski J, Zwierzchowski
L and Bagnicka E: Defensins: natural component of human innate
immunity. Hum Immunol. 74:1069–1079. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhu C, He N, Cheng T, Tan H, Guo Y, Chen
D, Cheng M, Yang Z and Zhang X: Ultrasound-targeted microbubble
destruction enhances human β-defensin 3 activity against
antibiotic-resistant Staphylococcus biofilms. Inflammation.
36:983–996. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hinrichsen K, Podschun R, Schubert S,
Schröder JM, Harder J and Proksch E: Mouse beta-defensin-14, an
antimicrobial ortholog of human beta-defensin-3. Antimicrob Agents
Chemother. 52:1876–1879. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Röhrl J, Yang D, Oppenheim JJ and Hehlgans
T: Identification and biological characterization of mouse
beta-defensin 14, the orthologue of human beta-defensin 3. J Biol
Chem. 283:5414–5419. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hancock RE and Sahl HG: Antimicrobial and
host-defense peptides as new anti-infective therapeutic strategies.
Nat Biotechnol. 24:1551–1557. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shi Y, Song W, Feng ZH, Zhao YT, Li F,
Tian Y and Zhao YM: Disinfection of maxillofacial silicone
elastomer using a novel antimicrobial agent: recombinant human
beta-defensin-3. Eur J Clin Microbiol Infect Dis. 28:415–420. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Varoga D, Wruck CJ, Tohidnezhad M,
Brandenburg L, Paulsen F, Mentlein R, Seekamp A, Besch L and Pufe
T: Osteoblasts participate in the innate immunity of the bone by
producing human beta defensin-3. Histochem Cell Biol. 131:207–218.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Semple F, MacPherson H, Webb S, Cox SL,
Mallin LJ, Tyrrell C, Grimes GR, Semple CA, Nix MA, Millhauser GL,
et al: Human β-defensin 3 affects the activity of pro-inflammatory
pathways associated with MyD88 and TRIF. Eur J Immunol.
41:3291–3300. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Röhrl J, Yang D, Oppenheim JJ and Hehlgans
T: Human beta-defensin 2 and 3 and their mouse orthologs induce
chemotaxis through interaction with CCR2. J Immunol. 184:6688–6694.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu C, Qin H, Cheng T, Tan HL, Guo YY, Shi
SF, Chen DS and Zhang XL: Staphylococcus aureus supernatant induces
the release of mouse β-defensin-14 from osteoblasts via the p38
MAPK and NF-κB pathways. Int J Mol Med. 31:1484–1494.
2013.PubMed/NCBI
|
|
14
|
Zhu C, Wang J, Cheng T, Li Q, Shen H, Qin
H, Cheng M and Zhang X: The potential role of increasing the
release of mouse β-defensin-14 in the treatment of osteomyelitis in
mice: a primary study. PLoS One. 9:e868742014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang M, Chen Y, Zhang Y, Zhang L, Lu X and
Chen Z: Mannan-binding lectin directly interacts with Toll-like
receptor 4 and suppresses lipopolysaccharide-induced inflammatory
cytokine secretion from THP-1 cells. Cell Mol Immunol. 8:265–275.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Spadaro M, Montone M, Arigoni M,
Cantarella D, Forni G, Pericle F, Pascolo S, Calogero RA and
Cavallo F: Recombinant human lactoferrin induces human and mouse
dendritic cell maturation via Toll-like receptors 2 and 4. FASEB J.
28:416–429. 2014. View Article : Google Scholar : PubMed/NCBI
|