Introduction
Acinetobacter baumannii is an important
opportunistic pathogen that causes various types of human
infections and has become a primary cause of nosocomial infections
because of its broad antimicrobial resistance (1–3).
Aminoglycosides, a type of broad-spectrum antibiotics, continue to
serve an important role in treating serious infections caused by
gram-negative bacteria (4). However,
aminoglycoside resistance of A. baumannii has rapidly
increased and given rise to more challenges in the clinical
treatment of infections (5).
A. baumannii shows resistance to
aminoglycosides since functional aminoglycosides can be modified by
various aminoglycoside-modifying enzymes, including
acetyltransferases, phosphotransferases and
nucleotidyltransferases, into non-functional forms in the bacteria
(6). In addition, aminoglycoside
antibiotics bind specifically to the A-site of 16S ribosomal (r)RNA
in the 30S small subunit and interfere with the decoding of mRNA to
inhibit protein synthesis (7). In
addition, at least ten 16S rRNA methylase genes (armA,
rmtA, rmtB, rmtC, rmtD, rmtE, rmtF, rmtG, rmtH and
npmA) have been identified (8–12). These
16S rRNA methylases, which lead to the high-level resistance of
various aminoglycosides, can easily transfer to other bacteria
since their genes are typically present on plasmids (13). Therefore, the emergence and spread of
such bacteria should be carefully monitored. Since the 16S rRNA
methylases are key factors in the aminoglycoside resistance of
A. baumannii, the investigation of the acquisition of 16S
rRNA methylase genes by clinical isolates is important for the
prevention and treatment of their infections (14).
Aminoglycosides and carbapenems represent the class
of antimicrobials that are used to treat A. baumannii
infections. Aminoglycoside antibiotics are frequently ineffective
against strains of A. baumannii; however, combinations of
aminoglycosides and carbapenems can produce synergistic effects to
treat infected patients (15,16).
Previously, it has become evident that the outgrowth of
carbapenem-resistant isolates has resulted in it being difficult to
treat A. baumannii infections. One of the most important
mechanisms underlying the resistance of carbapenems is the
production of carbapenemases in A. baumannii (17). Class D oxacillinases (OXA type) are
the primary cause of prevalence in A. baumannii strains
(18). In addition, causes stem from
class B β-lactamases (metallo-β-lactamases) and Klebsiella
pneumoniae Carbapenemase (KPC) producers. These carbapenemases
are a diverse group of β-lactamases that are active against the
carbapenems, resulting in their limited clinical use.
Several studies have documented the co-existence of
blaOXA-23 and armA in multidrug resistant
A. baumannii isolates (19–22). For
example, Doi et al (19)
first discovered that two of five A. baumannii isolates
coproduced OXA-23 β-lactamase and ArmA in North America in 2007. In
addition, further cases were reported in Korea (20,23),
India (24), France (25), Bulgaria (26), Italy (27), Latvia (28), East Africa (29), Yemen (30), Japan (31), Brunei (32), Egypt (33) and China (21,34,35). The
authors of the present study previously determined that
extended-spectrum β-lactamase and 16S rRNA methylase are coproduced
in A. baumannii (36).
However, the high-level resistance to aminoglycosides, coupled with
carbapenem resistance in A. baumannii, were not reported
over the 4-year period in China, particularly in western China.
The aim of the present study was to explore the
high-level resistance mechanisms against aminoglycosides, and to
investigate the presence of carbapenemases among strains of A.
baumannii. In addition, the relatedness of aminoglycoside- and
carbapenem-resistant strains, determined through epidemiologic
examination, is described. To the best of our knowledge, the
present study is the first to document the emergence of A.
baumannii producing blaOXA-23 and
blaOXA-51 carbapenemase-encoding genes among
armA 16S rRNA methylases at a university hospital in western
China. Furthermore, the results aim to emphasize that the dearth of
appropriate treatments remains a primary concern regarding
multidrug-resistant infections.
Materials and methods
Clinical isolates
A total of 118 strains of A. baumannii were
consecutively collected in a university hospital of western China
between February 2012 and July 2013. Rapid species identification
was performed by polymerase chain reaction (PCR), as reported
within ‘Resistance gene amplification’ and previously described
(37). A. baumannii was
identified and confirmed if the following two PCR products were
yielded: A 425-bp internal control amplicon corresponding to the
recA gene of Acinetobacter spp., and a 208-bp
fragment of the 16S rRNA intergenic spacer region of A.
baumannii (38).
Non-baumannii species of Acinetobacter, which yielded
the 425-bp product alone, were excluded in this study. Isolates
were obtained from specimens including sputum, secretion, lavage
fluids, blood and other specimens. All strains were stored at
−80°C. Bacteria were grown on tryptose agar or Mueller-Hinton
broth. No amplicons were obtained with bacteria belonging to other
genera.
Antimicrobial susceptibility
testing
The minimum inhibitory concentrations (MICs) of
amikacin and gentamicin (Sangon Biotech Co., Ltd., Shanghai, China)
for A. baumannii were determined on Mueller-Hinton agar
plates by agar dilution according to the protocol recommended by
the Clinical and Laboratory Standards Institute (39). MICs of meropenem and imipenem (Sangon
Biotech Co., Ltd.) were tested in high-level
aminoglycoside-resistant isolates. The results were interpreted
according to the CLSI guidelines. Escherichia coli [American
Type Culture Collection (ATCC) 25922] and A. baumannii (ATCC
19606) (ATCC, Manassas, VA, USA) were used as quality control
strains.
Resistance gene amplification
The aminoglycoside-modifying enzyme genes and the
16S rRNA methylase genes were detected by PCR. Total DNA was
extracted by boiling at 95°C for 15 min. After centrifugation at
13,000 × g for 10 min to pellet the debris, the supernatant
was stored at −20°C for further assays. PCR was performed in a
total volume of 50 µl containing 0.2 mM of each deoxynucleotide,
0.5 µM of each primer (Table I), 2.5
units Taq polymerase (Takara Bio, Inc., Dalian, China) and 5
µl 10X buffer (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Primers listed in Table I were
synthesized by Sangon Biotech Co., Ltd.. The PCR thermal cycling
conditions were as follows: Initial denaturation at 94°C for 5 min
in order to obtain partial activation of Taq polymerase;
then, the number of cycles was increased to 30, each consisting of
a denaturation step for 30 sec (at 94°C), an annealing step for 30
sec (at 55°C for armA, rmtA, rmtB,
rmtC, rmtD, rmtE, rmtF, rmtG,
rmtH and npmA, at 53°C for ant(2″)-Ia,
aph(3′)-1a, aac(3)-Ia
and aac(3)-IIa, and at 56°C
for aac(6′)-Ib) and an extension step for 30 sec (at 72°C).
Each amplification experiment included a blank containing the
reagent except for target DNA. The products were electrophoresed in
1% agarose gels and visualized under ultra-violet light (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). All aac(6′)-Ib PCR
products were directly sequenced and compared with the published
nucleotide (NC_005327.1).
 | Table I.Primers used in the present study for
polymerase chain reaction detection. |
Table I.
Primers used in the present study for
polymerase chain reaction detection.
| Primer | Target | Oligonucletides (5′
to 3′) | Expected size
(bp) |
|---|
| armA
forward | armA |
AGGTTGTTTCCATTTCTGAG | 591 |
| armA-R |
|
TCTCTTCCATTCCCTTCTCC |
|
| rmtA
forward | rmtA |
CTAGCGTCCATCCTTTCCTC | 635 |
| rmtA-R |
|
TTTGCTTCCATGCCCTTGCC |
|
| rmtB
forward | rmtB- |
CCCAAACAGACCGTAGAGGC | 585 |
| rmtB-R |
|
CTCAAACTCGGCGGGCAAGC |
|
| rmtC
forward | rmtC |
CGAAGAAGTAACAGCCAAAG | 711 |
| rmtC-R |
|
ATCCCAACATCTCTCCCACT |
|
| rmtD
forward | rmtD |
CGGCACGCGATTGGGAAGC | 401 |
| rmtD-R |
|
CGGAAACGATGCGACGAT |
|
| rmtE
forward | rmtE |
ATGAATATTGATGAAATGGTTGC | 823 |
| rmtE-R |
|
TGATTGATTTCCTCCGTTTTTG |
|
| rmtF
forward | rmtF |
GCGATACAGAAAACCGAAGG | 589 |
| rmtF-R |
|
ACCAGTCGGCATAGTGCTTT |
|
| rmtG
forward | rmtG |
AAATACCGCGATGTGTGTCC | 250 |
| rmtG
reverse |
|
ACACGGCATCTGTTTCTTCC |
|
| rmtH
forward | rmtH |
GCTTAAACCCGCTGATGCT | 332 |
| rmtH
reverse |
|
AAACCAGGTGGCGTAGTGC |
|
| npmA
forward | npmA |
GGAGGGCTATCTAATGTGGT | 371 |
| npmA
reverse |
|
GCCCAAAGAGAATTAAACTG |
|
| ant(2″)-Ia
forward |
ant(2″)-Ia |
GCTTACGTTGTCCCGCATTT | 215 |
| ant(2″)-Ia
reverse |
|
CCTTGGTGATCTCGCCTTTC |
|
| aph(3′)-Ia
forward |
aph(3′)-Ia |
CGAGCATCAAATGAAACTGC | 623 |
| aph(3′)-Ia
reverse |
|
GCGTTGCCAATGATGTTACAG |
|
| aac(3)-Ia
forward |
aac(3)-Ia |
GACATAAGCCTGTTCGGTT | 372 |
| aac(3)-Ia
reverse |
|
CTCCGAACTCACGACCGA |
|
| aac(3)-IIa
forward |
aac(3)-IIa |
ATGCATACGCGGAAGGC | 822 |
| aac(3)-IIa
reverse |
|
TGCTGGCACGATCGGAG |
|
| aac(6′)-Ib
forward |
aac(6′)-Ib |
AAGCGTTTTAGCGCAAGAGT | 366 |
| aac(6′)-Ib
reverse |
|
GCGTGTTTGAACCATGTACA |
|
| OXA-23
forward | OXA-23 |
GATCGGATTGGAGAACCAGA | 501 |
| OXA-23
reverse |
|
ATTTCTGACCGCATTTCCAT |
|
| OXA-24
forward | OXA-24 |
CAAGAGCTTGCAAGACGGACT | 420 |
| OXA-24
reverse |
|
TCCAAGATTTTCTAGCRACTTATA |
|
| OXA-51
forward | OXA-51 |
TAATGCTTTGATCGGCCTTG | 353 |
| OXA-51
reverse |
|
TGGATTGCACTTCATCTTGG |
|
| OXA-58
forward | OXA-58 |
TCGATCAGAATGTTCAAGCGC | 530 |
| OXA-58
reverse |
|
ACGATTCTCCCCTCTGCGC |
|
| NDM-1
forward | NDM-1 |
TCTCGACATGCCGGGTTTCGG | 475 |
| NDM-1
reverse |
|
ACCGAGATTGCCGAGCGACTT |
|
| KPC
forward | KPC |
GCTCAGGCGCAACTGTAAGT | 823 |
| KPC
reverse |
|
GTCCAGACGGAACGTGGTAT |
|
| IMP
forward | IMP |
CTACCGCAGAGTCTTTG | 587 |
| IMP
reverse |
|
AACCAGTTTTGCCTTACCAT |
|
| SIM
forward | SIM |
TACAAGGGATTCGGCATCG | 570 |
| SIM
reverse |
|
TAATGGCCTGTTCCCATGTG |
|
Genes coding for classes A, B and D carbapenemases
were investigated among high-level aminoglycoside-resistant
isolates by PCR. The genes encoding class A, such as Klebsiella
pneumoniae carbapenemase gene (blaKPC)
(40), class B, such as the
metallo-β-lactamase genes [blaIMP (41), blaVIM-1 (42), blaSIM (43) and blaNDM-1
(44)] and class D, such as CHDLs
[blaOXA-23 (45),
blaOXA-24 (45),
blaOXA-51 (46)
and blaOXA-58 (47)], were also analyzed using PCR.
Reaction conditions of PCR were as follows: 94°C for 5 min; and 30
cycles of 94°C for 30 sec, 56°C for 30 sec and 72°C for 30 sec;
followed by a final extension at 72°C for 5 min.
Multilocus sequence typing (MLST)
MLST was performed according a the previously
described A. baumannii MLST study (48). Briefly, internal fragments of seven
housekeeping genes (gltA, gyrB, gdhB,
recA, cpn60, gpi and rpoD) were
amplified by PCR (49). The
sequences of the seven housekeeping genes were compared with
existing sequences in the MLST database (50) for the assignment of allelic numbers.
Sequence types (STs) were assigned according to their allelic
profiles. New allele sequences and STs were assigned in accordance
with the PubMLST database (50). The
eBURST program (http://eburst.mlst.net) was used to cluster STs into
clonal complex (CC) and infer evolutionary descent (51).
Results
Antimicrobial susceptibility of
aminoglycosides
All 118 clinical strains were identified as A.
baumannii by 16S rRNA and recA amplification. Among
these isolates, 73 (61.86%) and 72 (61.02%) strains were resistant
to gentamicin and amikacin, respectively (Tables II and III). Thus, the resistance to amikacin and
gentamicin was observed in 66 (55.93%, 66/118) A. baumannii
isolates. A total of 78 (66.1%, 78/118) isolates were resistant to
amikacin and gentamicin, and 75 (96.15%, 75/78) of the strains
showed a high level of resistance (MIC, ≥256 µg/ml; Table III).
| Table II.Susceptibilities to two types of
aminoglycosides of A. baumannii isolates. |
Table II.
Susceptibilities to two types of
aminoglycosides of A. baumannii isolates.
| Drug name | Resistant isolates,
n (%) | Intermediate
isolates, n (%) | Sensitive isolates,
n (%) | Total, n (%) |
|---|
| Gentamicin | 73 (61.86) | 3 (2.54) | 42 (35.60) | 118 (100) |
| Amikacin | 72 (61.02) | 0 (0.00) | 46 (38.98) | 118 (100) |
| Table III.Molecular resistance characteristics
of 75 high level aminoglycoside resistance isolates. |
Table III.
Molecular resistance characteristics
of 75 high level aminoglycoside resistance isolates.
|
| Susceptibility
patter (MIC in µg/ml) | Resistance
genes |
|---|
|
|
|
|
|---|
| Isolates | Gentamicin | Amikacin | Imipenem | Meropenem | armA |
ant(2″)-Ia |
aac(6′)-Ib |
aph(3′)-Ia |
aac(3)-Ia |
aac(3)-IIa |
blaOXA-23 |
blaOXA-51 |
|---|
| 001 | >1024 | 1024 | 16 | 32 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| 003 | 256 | 1024 | 16 | 32 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| 005 | >1024 | 1024 | 32 | 64 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| 006 | >1024 | 1024 | 32 | 64 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| 007 | >1024 | 1024 | 32 | 64 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 |
| 008 | >1024 | 1024 | 32 | 64 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 1 |
| 011 | >1024 | 1024 | 16 | 32 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| 013 | 256 | 256 | 8 | 16 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 |
| 016 | >1024 | 1024 | 16 | 32 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| 018 | 256 | 256 | 8 | 16 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 1 |
| 020 | 2 | 256 | 8 | 16 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 1 |
| 026 | >1024 | 1024 | 16 | 32 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 1 |
| 027 | >1024 | 1024 | 16 | 32 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 |
| 028 | 1024 | 1024 | 8 | 16 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 1 |
| 030 | >1024 | 1024 | 32 | 32 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 |
| 031 | >1024 | 1024 | 32 | 16 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 1 |
| 034 | >1024 | 1024 | 32 | 64 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 |
| 035 | >1024 | 4 | 32 | 64 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 1 |
| 036 | >1024 | 1024 | 16 | 32 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 |
| 037 | >1024 | 1024 | 32 | 64 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 |
| 039 | >1024 | 1024 | 32 | 64 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 |
| 040 | >1024 | 1024 | 32 | 32 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 |
| 041 | >1024 | 1024 | 32 | 64 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 |
| 042 | >1024 | 512 | 32 | 64 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| 043 | >1024 | 1024 | 32 | 64 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 |
| 044 | >1024 | 1024 | 64 | 128 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 |
| 046 | >1024 | 1024 | 32 | 64 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 |
| 047 | >1024 | 1024 | 64 | 64 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 |
| 048 | >1024 | 1024 | 32 | 64 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 1 |
| 049 | >1024 | 512 | 32 | 64 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| 050 | >1024 | 512 | 32 | 32 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 |
| 051 | >1024 | 1024 | 32 | 32 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 |
| 052 | >1024 | 1024 | 64 | 128 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 |
| 053 | >1024 | 512 | 32 | 32 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 054 | >1024 | 512 | 32 | 32 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 |
| 057 | >1024 | 1024 | 32 | 64 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 |
| 058 | >1024 | 1024 | 32 | 64 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 1 |
| 059 | >1024 | 512 | 32 | 32 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 |
| 060 | >1024 | 1024 | 32 | 32 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 |
| 061 | >1024 | 1024 | 16 | 32 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 1 |
| 062 | >1024 | 512 | 32 | 32 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 1 |
| 063 | >1024 | 1024 | 32 | 32 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 1 |
| 064 | 512 | 8 | 32 | 32 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 |
| 065 | 256 | 8 | 32 | 16 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| 066 | >1024 | 512 | 32 | 64 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 |
| 067 | 256 | 256 | 16 | 32 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| 068 | >1024 | 512 | 32 | 32 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| 069 | >1024 | 512 | 32 | 32 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 |
| 072 | 256 | 256 | 16 | 16 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 |
| 074 | 512 | 2 | 16 | 8 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 1 |
| 075 | 8 | 1024 | 32 | 32 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 1 |
| 076 | >1024 | >1024 | 64 | 64 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 1 |
| 079 | 512 | 512 | 32 | 64 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 |
| 080 | 512 | 512 | 64 | 128 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 1 |
| 082 | 512 | 512 | 32 | 32 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 1 |
| 085 | >1024 | 512 | 16 | 32 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 |
| 087 | 512 | 512 | 32 | 32 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 1 |
| 089 | 4 | 512 | 16 | 32 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 |
| 090 | >1024 | 1024 | 32 | 32 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 1 |
| 093 | >1024 | 1024 | 32 | 128 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 1 |
| 094 | >1024 | >1024 | 16 | 32 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 |
| 095 | 512 | 512 | 16 | 32 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 |
| 096 | >1024 | >1024 | 32 | 32 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 |
| 097 | >1024 | >1024 | 16 | 32 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 |
| 098 | 512 | 512 | 16 | 32 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 1 |
| 099 | 512 | 256 | 32 | 32 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 0 |
| 100 | 8 | 512 | 0.5 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 |
| 101 | >1024 | 512 | 32 | 32 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| 102 | 512 | 256 | 32 | 32 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 |
| 104 | 512 | 256 | 32 | 32 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 |
| 106 | 1024 | 512 | 32 | 32 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 |
| 107 | 512 | 256 | 16 | 32 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 |
| 109 | >1024 | 512 | 8 | 16 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 |
| 113 | >1024 | 1024 | 8 | 0.5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| 120 | >1024 | 2 | 8 | 32 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 |
Co-occurrence of
aminoglycoside-resistant enzymes and carbapenemases
To determine the role of the
aminoglycoside-modifying enzymes in resistance and the 16S rRNA
methylases, PCR was performed to detect the concomitant genes
(Table III). The positive rates of
ant(2″)-Ia, aac(6′)-Ib, aph(3′)-Ia,
aac(3)-Ia and aac(3)-IIa were 66.95 (79/118), 69.49
(82/118), 42.37 (50/118), 39.83 (47/118) and 14.41% (17/118),
respectively (Table IV). Fifty-four
of 118 (45.76%) isolates harboring the 16S rRNA methyalse
armA gene obtained high level of resistance to amikacin and
gentamicin. rmtA, rmtB, rmtC, rmtD,
rmtE, rmtF, rmtG and npmA genes were
not detected in all of the isolates.
| Table IV.Positive rates of genes in A.
baumannii. |
Table IV.
Positive rates of genes in A.
baumannii.
| Gene | Positive rate, %
(n/118) | Gene | Positive rate, %
(n/118) |
|---|
| armA | 45.76 (54/118) |
aph(3′)-Ia | 42.37 (50/118) |
|
aac(6′)-Ib | 69.45 (82/118) |
aac(3)-Ia | 39.83 (47/118) |
|
ant(2″)-la | 66.95 (79/118) |
aac(3)-IIa | 14.41 (17/118) |
There was a marked difference in the distribution of
aminoglycoside-resistant genes among the 75 high-level
aminoglycoside-resistant A. baumannii (Tables III and IV). All 54 (72.0%, 54/75)
armA-positive strains were confirmed to serve a primary role
in high level aminoglycoside resistance. However, 21 (28%, 21/75)
isolates harboring aminoglycoside-modifying enzymes without the
armA gene served the same function (Table V).
| Table V.Distribution of aminoglycoside
resistance genes in 75 high level aminoglycoside resistance
clinical isolates of A. baumannii, expressed as positive (+)
or negative (−). |
Table V.
Distribution of aminoglycoside
resistance genes in 75 high level aminoglycoside resistance
clinical isolates of A. baumannii, expressed as positive (+)
or negative (−).
|
armA-positive aminoglycoside
resistance gene profile (n=54) |
blaOXA-23 (n=58) |
blaOXA-51 (n=75) | No. of
isolates |
|---|
|
ant(2″)-Ia | +2 | + | 2 (2.67%) |
|
aac(6)-Ib | +1 | + | 1 (1.33%) |
|
ant(2)-Ia+aac(6′)-Ib | +4 | + | 4 (5.33%) |
|
ant(2)-Ia+aph(3′)-Ia | +2 | + | 2 (2.67%) |
|
aac(6′)-Ib+aac(3)-Ia | +3 | + | 3 (4.0%) |
|
aac(6′)-Ib+aac(3)-IIa | +2 | + | 2 (2.67%) |
|
ant(2″)-Ia+aac(6′)-Ib+aac(3)-Ia |
+9/−3 | + | 12 (16%) |
|
ant(2″)-Ia+aac(6′)-Ib+aac(3)-IIa | +1/−1 | + | 2 (2.67%) |
|
ant(2″)-Ia+aac(6′)-Ib+aph(3′)-Ia | +9/−1 | + | 10 (13.3%) |
|
ant(2″)-Ia+aph(3′)-Ia+aac(3)-IIa | +1 | + | 1 (1.33%) |
|
ant(2″)-Ia+aac(6′)-Ib+aph(3′)-Ia+aac(3)-Ia | +8/−2 | + | 10 (13.3%) |
|
ant(2″)-Ia+aac(6′)-Ib+aph(3′)-Ia+aac(3)-IIa | −1 | + | 1 (1.33%) |
|
ant(2″)-Ia+aac(6′)-Ib+aac(3)-Ia+aac(3)-IIa | +1 | + | 1 (1.33%) |
|
ant(2″)-Ia+aac(6′)-Ib+aph(3′)-Ia+aac(3)-Ia+aac(3)-IIa | −3 | + | 3 (4.0%) |
| None of armA
genes (21) |
|
|
|
|
ant(2″)-Ia | + | + | 1 (1.33%) |
|
aac(6′)-Ib | − | + | 1 (1.33%) |
|
aac(6′)-Ib+ant(2″)-Ia | + | + | 1 (1.33%) |
|
aac(6′)-Ib+aac(3)-Ia | −2 | + | 2 (2.67%) |
|
aac(6′)-Ib+aac(3)-IIa | + | + | 1 (1.33%) |
|
ant(2″)-Ia+aac(6′)-Ib+aph(3′)-Ia | +5 | + | 5 (6.67%) |
|
ant(2″)-Ia+aac(6′)-Ib+aac(3)-Ia | + | + | 1 (1.33%) |
|
ant(2″)-Ia+aac(6′)-Ib+aac(3)-IIa | +2 | + | 2 (2.67%) |
|
ant(2″)-Ia+aph(3′)-Ia+aac(3)-IIa | − | + | 1 (1.33%) |
|
ant(2″)-Ia+aac(6′)-Ib+aph(3′)-Ia+aac(3)-Ia | +/−2 | + | 3 (4.0%) |
|
ant(2″)-Ia+aac(6′)-Ib+aph(3′)-Ia+aac(3)-IIa | + | + | 1 (1.33%) |
|
aac(6′)-Ib+aph(3′)-Ia+aac(3)-Ia+aac(3)-IIa | + | + | 1 (1.33%) |
|
ant(2″)-Ia+aac(6′)-Ib+aph(3′)-Ia+aac(3)-Ia+aac(3)-IIa | + | + | 1 (1.33%) |
Among the 54 isolates that were
armA-positive, the prevalence of blaOXA-23
and blaOXA-51 gene occurrences were 79.63 (43/54)
and 100% (54/54), respectively. In addition, the prevalence of
ant(2″)-Ia, aac(6′)-Ib, aph(3′)-Ia,
aac(3)-Ia, and
aac(3)-IIa positive rates of
genes was distributed in the aminoglycoside-resistant
and-susceptible strains (Table V).
As described above, the present study demonstrated that
aminoglycoside-modifying enzymes were mostly responsible for
moderate level resistance (16 µg/ml<MIC<256 µg/ml) to
aminoglycosides in A. baumannii, whereas armA was
responsible for high-level resistance to aminoglycosides. All 75
isolates with high-level resistance to aminoglycosides harbored the
carbapenemase genes blaOXA-23 (77.33%) or
blaOXA-51 (100%; Tables III and V), which (except one isolate) showed
resistance to the carbapenems, imipenem and meropenem. These data
suggest that the resistance to carbapenems and aminoglysides poses
a threat following combination treatment of A. baumannii
infection.
Molecular genotyping analysis of
drug-resistant isolates
To better assess the A. baumannii clinical
population epidemiology and the genetic background of these
strains, a number of molecular typing systems were applied. By
comparing the ST(s) of 75 high-level aminoglycoside resistant
isolates with all identified ST(s) in A. baumannii in the
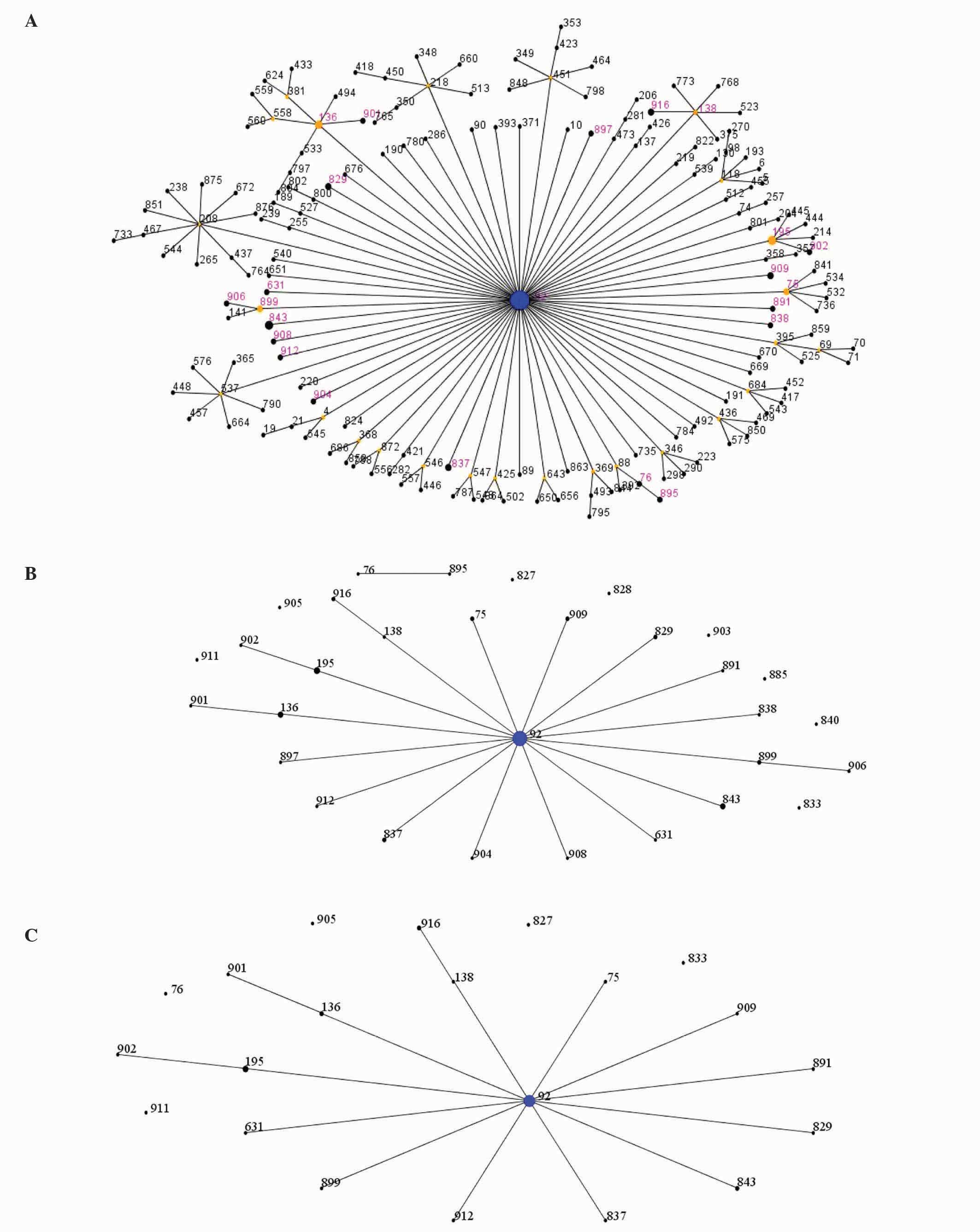
MLST database by eBUSRT analysis, 29 strains were identified that
belonged to ST92, which is a globally distributed strain (Fig. 1A). According to MLST analysis, a
total of 31 different STs were assigned to the 75 high-level
aminoglycoside resistant isolates, of which 21 STs were clustered
into clonal complex 92 (CC92), and the remaining 10 STs were
identified as singletons. The most common ST was ST92, which
accounted for 38.67% (29/75) (Fig. 1A
and B). ST195, followed by ST92, presented in 5 strains, whilst
ST136 and ST843 were detected in 4 strains. ST75, ST829, ST837,
ST899, ST909 and ST916 were represented by 2 isolates. Molecular
analysis revealed that 37 (containing 6 different STs) of the 43
isolates, which produced carbapenemase OXA-23 and 16S rRNA
methylase ArmA, were grouped into CC92, while the remaining 6
isolates, which had 6 different STs, could not be clustered into
any known clonal complex (Fig. 1C).
These data indicate that the prevalence of A. baumannii
isolates was caused by CC92 dissemination.
Discussion
A. baumannii are important hospital-acquired
pathogens that cause various types of human infections (52). The present study demonstrated that 75
(63.56%) strains were high-level resistant to amikacin or
gentamicin, determined by susceptibility testing (Table III), suggesting that these
antibiotics can only be used for treating A. baumannii
infections induced by susceptible strains.
As indicated above, at least one aminoglycoside
resistance gene was detected in aminoglycoside-resistant A.
baumannii strains, and different resistant genes were commonly
present in the same isolates (Tables
III and V). Among these strains,
the dominant aminoglycoside-resistant genotypes are
ant(2″)-Ia, armA and aac(6′)-Ib, which were
present at 66.95, 45.76 and 69.49%, respectively (Table IV). These results indicated that the
presence of armA and aminoglycoside-modifying enzmyes
confers to the high level of aminoglycoside resistance.
The prevalence of armA genes in A.
baumannii isolates has been described in several studies that
showed 50% (52/104) in strains isolated in Lishui, eastern China
(10), 60.4% (61/101) in clinical
strains in Vietnam (53), and 59.54%
(103/173) in hospitals in Beijing, China (54). In the present study, 45.76% (54/118,
Table IV) of isolates harbored the
armA gene, which is similar to the above cases reported in
China. In addition, it was reported that armA was identified
in 90% (97/107) of the multidrug-resistant strains in Shanghai,
eastern China (55). In a previous
study, however, 4 (8.5%) isolates were positive for the methylase
enzyme ArmA in an Algerian hospital (56). In conclusion, armA is highly
prevalent worldwide, particularly in China.
The emergence of high level aminoglycoside
resistance may pose a question for the combination therapy of
aminoglycoside with β-lactams, particularly carbapenems in treating
A. baumannii infections. Previously, A. baumannii
producing OXA-23 have been increasingly described in Shanghai,
eastern China (38). Thus, the
present study identified carbapenemase genes in 75 high-level
aminoglycoside resistance strains. The positive ratios of
blaOXA-51 and blaOXA-23 were
100 (75/75) and 77.33% (58/75), respectively (Table III), further demonstrating that the
intrinsic OXA-51 family and the presence of OXA-23 are the most
prevalent mechanisms for carbapenem resistance in A.
baumannii (57). In addition,
among 54 armA-positive isolates, the prevalence of
blaOXA-23 and blaOXA-51 were
79.63 (43/54) and 100% (54/54) (Table
V), which was similar to a previous study (27,56).
Three hospital disseminations of A. baumannii co-producing
OXA-23 and ArmA were reported in eastern China in 2009 and 2011
(21,34,35). To
the best of our knowledge, the results in the present study are the
first to demonstrate the co-occurrence of carbapenemases OXA-23,
OXA-51 and 16S rRNA methylase ArmA with high level aminoglycoside
resistance among clinical isolates of A. baumannii from
Chengdu, western China.
Previously, it was reported that aminoglycosides
with the aac/aad riboswitch control the expression of
aminoglycoside modification enzymes (58), indicating that bacteria can survive
in an energy saving way. Therefore, these efficient modification
enzymes were responsible for aminoglycoside resistance (Table IV). In addition, it was identified
that the aac(6′)-Ib enzyme is able to modify amikacin, even
in phenotypically amikacin-susceptible isolates (59). Furthermore, the aac(6′)-Ib
(69.49%) A. baumannii isolates were aminoglycoside-positive
(Table IV), which is different from
previous studies (10). The reason
why these differences were observed may be due to the resistance
level caused by aac(6′)-Ib, which was regional-dependent and
host bacterium-dependent (59).
In the present study, a higher rate of
aac(3)-IIa (14.41%) were
detected. In addition, aac(3)-IIa genes were detected in 47.88% of
E. coli isolated from an Iranian hospital (60). Miro et al (61) found 12.4% of strains possessing
aac(3)-IIa genes. However,
there is a paucity of data regarding the aac(3)-IIa gene distribution in A.
baumannii. It was reported that only 4 strains (3.7%) carried
aac(3)-IIa genes (62); aac(3)-IIa was not identified in any strains
in a study by Nowak et al (63). Previous studies have reported that
aac(3)-IIa modifies
gentamicin, which explains the observed high rate of resistance to
gentamicin in these A. baumannii strains (59). The increasing prevalence of
aminoglycoside resistance is partly associated with the presence of
aac(3′)-IIa.
The PubMLST database assigned A. baumannii
strains to 920 different types. ST92, a globally distributed type,
was the predicted founder of CC92 in the A. baumannii MLST
database. CC92 is the largest and most geographically diverse
clonal complex (64). Combined ST
profiles from MLST and eBURST analyses showed that almost all
isolates were clonally related and CC92 was responsible for the
spread of disease (Fig. 1). The
present study further suggests the possibility that A.
baumannii carrying blaOXA-23 and armA
genes contribute towards CC92 dissemination. In addition, the
present study described the emergence and spread of a clonal strain
of the high-level aminoglycoside-resistant A. baumannii.
These findings support the hypothesis that certain restricted
genetic backgrounds serve an important role in the emergence of
aminoglycoside resistance, since some genetic backgrounds may be
prone to acquire a foreign resistance gene and maintain its
stability and expression (46).
Further analysis of the epidemiology of A. baumannii is
required in order to determine the prevalence of drug-resistant
genes.
In conclusion, the present study demonstrated that
16S rRNA methylase ArmA and modifying enzyme occurrence confer high
level resistance to aminoglycoside in A. baumannii. In
addition, it was identified that the high level aminoglycoside
resistance of A. baumannii strains, harboring high
percentages of positive carbapenemases blaOXA-23
and blaOXA-51, strongly suggest that a better
understanding of the global epidemiology and monitoring for the
presence of resistance genes is urgently required.
Acknowledgments
The authors would like to thank members of the Key
Laboratory of Non Coding RNA and Drug Discovery, the Education
Department of Sichuan Province, Chengdu, China for their input. The
present study was supported by grants from the National Natural
Science Foundation of China (grant nos. 81373454, 31300659,
31470246 and 31401099), Applied Basic Research Programs of Sichuan
Province, China (grant no. 2013jy0065) Scientific Research and
Innovation Team of Sichuan Province, China (grant no. 15TD0025) and
the Preeminent Youth Fund of Sichuan Province, China (grant no.
2015JQO019).
References
|
1
|
Sheikh YA, Marie MA, John J, Krishnappa LG
and Dabwab KH: Prevalence of 16S rRNA methylase genes among
β-lactamase-producing Enterobacteriaceae clinical isolates in Saudi
Arabia. Libyan J Med. 9:244322014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Belbel Z, Chettibi H, Dekhil M, Ladjama A,
Nedjai S and Rolain JM: Outbreak of an armA
Methyltransferase-Producing ST39 Klebsiella pneumoniae clone in a
pediatric Algerian hospital. Microb Drug Resist. 20:310–315. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu Z, Ling B and Zhou L: Prevalence of
16S rRNA methylase, modifying enzyme, and extended-spectrum
beta-lactamase genes among Acinetobacter baumannii isolates. J
Chemother. 27:207–212. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nemec A, Dolzani L, Brisse S, van den
Broek P and Dijkshoorn L: Diversity of aminoglycoside-resistance
genes and their association with class 1 integrons among strains of
pan-European Acinetobacter baumannii clones. J Med Microbiol.
53:1233–1240. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Labby KJ and Garneau-Tsodikova S:
Strategies to overcome the action of aminoglycoside-modifying
enzymes for treating resistant bacterial infections. Future Med
Chem. 5:1285–1309. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ramirez MS and Tolmasky ME: Aminoglycoside
modifying enzymes. Drug Resist Updat. 13:151–171. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cho YJ, Moon DC, Jin JS, Choi CH, Lee YC
and Lee JC: Genetic basis of resistance to aminoglycosides in
Acinetobacter spp. and spread of armA in Acinetobacter baumannii
sequence group 1 in Korean hospitals. Diagn Microbiol Infect Dis.
64:185–190. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bueno MF, Francisco GR, O'Hara JA, de
Oliveira Garcia D and Doi Y: Coproduction of 16S rRNA
methyltransferase RmtD or RmtG with KPC-2 and CTX-M group
extended-spectrum β-lactamases in Klebsiella pneumoniae. Antimicrob
Agents Chemother. 57:2397–2400. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Galimand M, Courvalin P and Lambert T:
RmtF, a new member of the aminoglycoside resistance 16S rRNA N7
G1405 methyltransferase family. Antimicrob Agents Chemother.
56:3960–3962. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang J, Ye M, Jia X, Yu F and Wang M:
Coexistence of armA and genes encoding aminoglycoside-modifying
enzymes in Acinetobacter baumannii. Afr J Microbiol Res.
6:5325–5330. 2012.
|
|
11
|
O'Hara JA, McGann P, Snesrud EC, Clifford
RJ, Waterman PE, Lesho EP and Doi Y: Novel 16S rRNA
methyltransferase RmtH produced by Klebsiella pneumoniae associated
with war-related trauma. Antimicrob Agents Chemother. 57:2413–2416.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wachino J and Arakawa Y: Exogenously
acquired 16S rRNA methyltransferases found in
aminoglycoside-resistant pathogenic Gram-negative bacteria: An
update. Drug Resist Updat. 15:133–148. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wachino J, Shibayama K, Kurokawa H, Kimura
K, Yamane K, Suzuki S, Shibata N, Ike Y and Arakawa Y: Novel
plasmid-mediated 16S rRNA m1A1408 methyltransferase, NpmA, found in
a clinically isolated Escherichia coli strain resistantto
structurally diverse aminoglycosides. Antimicrob Agents Chemother.
51:4401–4409. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nagasawa M, Kaku M, Kamachi K, Shibayama
K, Arakawa Y, Yamaguchi K and Ishii Y: Loop-mediated isothermal
amplification assay for 16S rRNA methylase genes in Gram-negative
bacteria. J Infect Chemother. 20:635–638. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Marques MB, Brookings ES, Moser SA, Sonke
PB and Waites KB: Comparative in vitro antimicrobial
susceptibilities of nosocomial isolates of Acinetobacter baumannii
and synergistic activities of nine antimicrobial combinations.
Antimicrob Agents Chemother. 41:881–885. 1997.PubMed/NCBI
|
|
16
|
Wen JT, Zhou Y, Yang L and Xu Y:
Multidrug-resistantgenes of aminoglycoside-modifying enzymes and
16S rRNA methylases in Acinetobacter baumannii strains. Genet Mol
Res. 13:3842–3849. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chang Y, Luan G, Xu Y, Wang Y, Shen M,
Zhang C, Zheng W, Huang J, Yang J, Jia X and Ling B:
Characterization of carbapenem-resistant Acinetobacter baumannii
isolates in a Chinese teaching hospital. Front Microbiol.
6:9102015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Walther-Rasmussen J and Høiby N: OXA-type
carbapenemases. J Antimicrob Chemother. 57:373–383. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Doi Y, Adams JM, Yamane K and Paterson DL:
Identification of 16S rRNA methylase-producing Acinetobacter
baumannii clinical strains in North America. Antimicrob Agents
Chemother. 51:4209–4210. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim JW, Heo ST, Jin JS, Choi CH, Lee YC,
Jeong YG, Kim SJ and Lee JC: Characterization of Acinetobacter
baumannii carrying bla(OXA-23), bla(PER-1) and armA in a Korean
hospital. Clin Microbiol Infect. 14:716–718. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou H, Du XX, Yang Q, Zhou JY, Yu YS and
Li LJ: Study on carbapenemase and 16S rRNA methylase of
imipenem-resistant Acinetobacter baumannii. Zhonghua Liu Xing Bing
Xue Za Zhi. 30:269–272. 2009.(In Chinese). PubMed/NCBI
|
|
22
|
Adams-Haduch JM, Paterson DL, Sidjabat HE,
Pasculle AW, Potoski BA, Muto CA, Harrison LH and Doi Y: Genetic
basis of multidrug resistance in Acinetobacter baumannii clinical
isolates at a tertiary medical center in Pennsylvania. Antimicrob
Agents Chemother. 52:3837–3843. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sung JY, Kwon KC, Cho HH and Koo SH:
Antimicrobial resistance determinants in imipenem-nonsusceptible
Acinetobacter calcoaceticus-baumannii complex isolated in Daejeon,
Korea. Korean J Lab Med. 31:265–270. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Karthikeyan K, Thirunarayan MA and
Krishnan P: Coexistence of blaOXA-23 with blaNDM-1 and armA in
clinical isolates of Acinetobacter baumannii from India. J
Antimicrob Chemother. 65:2253–2254. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bonnin RA, Potron A, Poirel L, Lecuyer H,
Neri R and Nordmann P: PER-7, an extended-spectrum beta-lactamase
with increased activity toward broad-spectrum cephalosporins in
Acinetobacter baumannii. Antimicrob Agents Chemother. 55:2424–2427.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Strateva T, Markova B, Marteva-Proevska Y,
Ivanova D and Mitov I: Widespread dissemination of
multidrug-resistant Acinetobacter baumannii producing OXA-23
carbapenemase and ArmA 16S ribosomal RNA methylase in a Bulgarian
university hospital. Braz J Infect Dis. 16:307–310. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Brigante G, Migliavacca R, Bramati S,
Motta E, Nucleo E, Manenti M, Migliorino G, Pagani L, Luzzaro F and
Viganò FE: Emergence and spread of a multidrug-resistant
Acinetobacter baumannii clone producing both the carbapenemase
OXA-23 and the 16S rRNA methylase ArmA. J Med Microbiol.
61:653–661. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Saule M, Samuelsen Ø, Dumpis U, Sundsfjord
A, Karlsone A, Balode A, Miklasevics E and Karah N: Dissemination
of a carbapenem-resistant Acinetobacter baumannii strain belonging
to international clone II/sequence type 2 and harboring a novel
AbaR4-like resistance island in Latvia. Antimicrob Agents
Chemother. 57:1069–1072. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Revathi G, Siu LK, Lu PL and Huang LY:
First report of NDM-1-producing Acinetobacter baumannii in East
Africa. Int J Infect Dis. 17:e1255–e1258. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bakour S, Alsharapy SA, Touati A and
Rolain JM: Characterization of Acinetobacter baumannii clinical
isolates carrying bla(OXA-23) carbapenemase and 16S rRNA methylase
armA genes in Yemen. Microb Drug Resist. 20:604–609. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tada T, Miyoshi-Akiyama T, Shimada K,
Shimojima M and Kirikae T: Dissemination of 16S rRNA methylase
ArmA-producing Acinetobacter baumannii and emergence of OXA-72
carbapenemase coproducers in Japan. Antimicrob Agents Chemother.
58:2916–2920. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tojo M, Mawatari M, Hayakawa K, Nagamatsu
M, Shimada K, Mezaki K, Sugiki Y, Kuroda E, Takeshita N, Kutsuna S,
et al: Multidrug-resistant Acinetobacter baumannii isolated from a
traveler returned from Brunei. J Infect Chemothe. 21:212–214. 2015.
View Article : Google Scholar
|
|
33
|
El-Sayed-Ahmed MA, Amin MA, Tawakol WM,
Loucif L, Bakour S and Rolain JM: High prevalence of bla(NDM-1)
carbapenemase-encoding gene and 16S rRNA armA methyltransferase
among Acinetobacter baumannii clinical isolates, Egypt. Antimicrob
Agents Chemother. 59:3602–3605. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhao WS, Liu GY, Mi ZH and Zhang F:
Coexistence of blaOXA-23 with armA and novel gyrA mutation in a
pandrug-resistant Acinetobacter baumannii isolate from the blood of
a patient with haematological disease in China. J Hosp Infect.
77:278–279. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhou H, Zhang T, Yu D, Pi B, Yang Q, Zhou
J, Hu S and Yu Y: Genomic analysis of the multidrug-resistant
Acinetobacter baumannii strain MDR-ZJ06 widely spread in China.
Antimicrob Agents Chemother. 55:4506–4512. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu Z, Ling B and Zhou L: Prevalence of
16S rRNA methylase, modifying enzyme, and extended-spectrum
beta-lactamase genes among Acinetobacter baumannii isolates. J
Chemother. 27:207–212. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen TL, Siu LK, Wu RC, Shaio MF, Huang
LY, Fung CP, Lee CM and Cho WL: Comparison of one-tube multiplex
PCR, automated ribotyping and intergenic spacer (ITS) sequencing
for rapid identification of Acinetobacter baumannii. Clin Microbiol
Infect. 13:801–806. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li Y, Guo Q, Wang P, Zhu D, Ye X, Wu S and
Wang M: Clonal dissemination of extensively drug-resistant
Acinetobacter baumannii producing an OXA-23 β-lactamase at a
teaching hospital in Shanghai, China. J Microbiol Immunol Infect.
48:101–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Clinical and Laboratory Standards
Institute, . Performance Standards for Antimicrobial Susceptibility
Testing. 24th Informational Supplement. CLSI; Wayne, PA, USA: M100,
2014
|
|
40
|
Li Y, Guo Q, Wang P, Zhu D, Ye X, Wu S and
Wang M: Clonal dissemination of extensively drug-resistant
Acinetobacter baumannii producing an OXA-23 β-lactamase at a
teaching hospital in Shanghai, China. J Microbiol Immunol Infect.
48:101–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Valenzuela JK, Thomas L, Partridge SR, van
der Reijden T, Dijkshoorn L and Iredell J: Horizontal gene transfer
in a polyclonal outbreak of carbapenem-resistant Acinetobacter
baumannii. J Clin Microbiol. 45:453–460. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tsakris A, Pournaras S, Woodford N,
Palepou MF, Babini GS, Douboyas J and Livermore DM: Outbreak of
infections caused by Pseudomonas aeruginosa producing VIM-1
carbapenemase in Greece. J Clin Microbiol. 38:1290–1292.
2000.PubMed/NCBI
|
|
43
|
Ellington MJ, Kistler J, Livermore DM and
Woodford N: Multiplex PCR for rapid detection of genes encoding
acquired metallo-beta-lactamases. J Antimicrob Chemother.
59:321–322. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yong D, Toleman MA, Giske CG, Cho HS,
Sundman K, Lee K and Walsh TR: Characterization of a new
metallo-beta-lactamase gene, bla(NDM-1) and a novel erythromycin
esterase gene carried on a unique genetic structure in Klebsiella
pneumoniae sequence type 14 from India. Antimicrob Agents
Chemother. 53:5046–5054. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Woodford N, Ellington MJ, Coelho JM,
Turton JF, Ward ME, Brown S, Amyes SG and Livermore DM: Multiplex
PCR for genes encoding prevalent OXA carbapenemases in
Acinetobacter spp. Int J Antimicrob Agents. 27:351–353. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fu Y, Zhou J, Zhou H, Yang Q, Wei Z, Yu Y
and Li L: Wide dissemination of OXA-23-producing
carbapenem-resistant Acinetobacter baumannii clonal complex 22 in
multiple cities of China. J Antimicrob Chemother. 65:644–650. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Netsvyetayeva I, Sikora M, Golas M,
Swoboda-Kopec E, de Walthoffen SW, Dembicka O, Fraczek M,
Mlynarczyk A, Pacholczyk M, Chmura A and Mlynarczyk G:
Acinetobacter baumannii multidrug-resistant strain occurrence in
liver recipients with reference to other high-risk groups.
Transplant Proc. 43:3116–3120. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Diancourt L, Passet V, Nemec A, Dijkshoorn
L and Brisse S: The population structure of Acinetobacter
baumannii: Expanding multiresistant clones from an ancestral
susceptible genetic pool. PLoS One. 5:e100342010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Bartual SG, Seifert H, Hippler C, Luzon
MA, Wisplinghoff H and Rodríguez-Valera F: Development of a
multilocus sequence typing scheme for characterization of clinical
isolates of Acinetobacter baumannii. J Clin Microbiol.
43:4382–4390. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
PubMLST, . Acinetobacter baumannii MLST
Databases. http://pubmlst.org/abaumannii/
|
|
51
|
Feil EJ, Li BC, Aanensen DM, Hanage WP and
Spratt BG: eBURST: Inferring patterns of evolutionary descent among
clusters of related bacterial genotypes from multilocus sequence
typing data. J Bacteriol. 186:1518–1530. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Agadzhanian VV, Semenikhin VA, Iu S
Fedorov, Krasulina GP, Gaǐfulina IM and Mironova LA: Experience of
health protection center on organization of medical care for coal
miners in Kuzbass. Med Tr Prom Ekol. 27–30. 2002.(In Russian).
PubMed/NCBI
|
|
53
|
Tada T, Miyoshi-Akiyama T, Kato Y,
Ohmagari N, Takeshita N, Hung NV, Phuong DM, Thu TA, Binh NG, Anh
NQ, et al: Emergence of 16S rRNA methylase-producing Acinetobacter
baumannii and Pseudomonas aeruginosa isolates in hospitals in
Vietnam. BMC Infect Dis. 13:2512013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Nie L, Lv Y, Yuan M, Hu X, Nie T, Yang X,
Li G, Pang J, Zhang J, Li C, et al: Genetic basis of high level
aminoglycoside resistance in Acinetobacter baumannii from Beijing,
China. Acta Pharm Sin B. 4:295–300. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Xiao SZ, Han LZ, Chu HQ, Zhao L, Chen X
and Ni YX: Detection of aminoglycoside resistance related genes in
multidrug-resistant Acinetobacter baumannii isolated from a single
institute of Shanghai, China. Panminerva Med. 57:49–53.
2015.PubMed/NCBI
|
|
56
|
Bakour S, Touati A, Bachiri T, Sahli F,
Tiouit D, Naim M, Azouaou M and Rolain JM: First report of 16S rRNA
methylase ArmA-producing Acinetobacter baumannii and rapid spread
of metallo-β-lactamase NDM-1 in Algerian hospitals. J Infect
Chemother. 20:696–701. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Azimi L, Talebi M, Pourshafie MR, Owlia P
and Lari A Rastegar: Characterization of carbapenemases in
extensively drug resistance Acinetobacter baumannii in a burn care
center in Iran. Int J Mol Cell Med. 4:46–53. 2015.PubMed/NCBI
|
|
58
|
Jia X, Zhang J, Sun W, He W, Jiang H, Chen
D and Murchie AI: Riboswitch control of aminoglycoside antibiotic
resistance. Cell. 152:68–81. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Haldorsen BC, Simonsen GS, Sundsfjord A
and Samuelsen Ø; Norwegian Study Group on Aminoglycoside
Resistance, : Increased prevalence of aminoglycoside resistance in
clinical isolates of Escherichia coli and Klebsiella spp. in Norway
is associated with the acquisition of AAC(3)-II and AAC(6′)-Ib.
Diagn Microbiol Infect Dis. 78:66–69. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Soleimani N, Aganj M, Ali L, Shokoohizadeh
L and Sakinc T: Frequency distribution of genes encoding
aminoglycoside modifying enzymes in uropathogenic E. coli isolated
from Iranian hospital. BMC Res Notes. 7:8422014. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Miro E, Grünbaum F, Gomez L, Rivera A,
Mirelis B, Coll P and Navarro F: Characterization of
aminoglycoside-modifying enzymes in enterobacteriaceae clinical
strains and characterization of the plasmids implicated in their
diffusion. Microb Drug Resist. 19:94–99. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Akers KS, Chaney C, Barsoumian A, Beckius
M, Zera W, Yu X, Guymon C, Keen EF III, Robinson BJ, Mende K and
Murray CK: Aminoglycoside resistance and susceptibility testing
errors in Acinetobacter baumannii-calcoaceticus complex. J Clin
Microbiol. 48:1132–1138. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Nowak P, Paluchowska PM and Budak A:
Co-occurrence of carbapenem and aminoglycoside resistance genes
among multidrug-resistant clinical isolates of Acinetobacter
baumannii from Cracow, Poland. Med Sci Monit Basic Res. 20:9–14.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Hamouda A, Evans BA, Towner KJ and Amyes
SG: Characterization of epidemiologically unrelated Acinetobacter
baumannii isolates from four continents by use of multilocus
sequence typing, pulsed-field gel electrophoresis and
sequence-based typing of bla(OXA-51-like) genes. J Clin Microbiol.
48:2476–2483. 2010. View Article : Google Scholar : PubMed/NCBI
|