Introduction
Enterovirus (EV) 71 is a member of the
picornaviridae family of viruses that contain single-stranded
positive-sense RNA (1), which
primarily infects children <5 years old (2). Infants with EV71 infection develop
sores on their hands, feet, buttocks and mouth; therefore, the
infection is referred to as hand, foot and mouth disease (HFMD)
(3). Several types of enterovirus
have been indicated to cause HFMD in children; however, EV71 is
commonly associated with severe complications of the nervous system
in infants with HFMD and is therefore considered to be a major
virulent pathogen and cause of mortalities in HFMD (4). Previous studies have indicated that a
total of 488,955 HFMD cases and 126 HFMD-associated mortalities
were reported in 2008 in China, of which EV71 infection was the
primary cause (5). EV71 is a
non-enveloped virus with a single-stranded positive-sense RNA
genome that is ~7.5 kb in size and belongs to the enterovirus genus
of the picornaviridae family (6).
EV71 is classified into five genotypes as follows: A, B, C, D and
F, and further subdivided into B0-B5 and C1-C5 subgenotypes
(7). HFMD is an important public
health issue and further understanding of the mechanisms underlying
the development of HFMD are required to develop effective
treatments. In mainland China, Taiwan and Singapore, EV71
whole-virus vaccines have been produced by various manufacturers.
Two available inactivated EV71 vaccines have been approved by the
China Food and Drug Administration, one produced by Sinovac Biotech
Ltd. and the other by the Chinese Academy of Medical Science (all
Beijing, China) (8–10). Currently, no effective treatment is
available to treat severe EV71 infections, aside from symptomatic
therapies (11).
Typically, Vero and RD cell lines have been used to
isolate EV71 (12), and suckling
BALB/c mice have been studied to evaluate the virulence and
pathogenicity of the isolated strain (13). A previous study indicated that
neonate mice were able to be fatally infected with EV71 from 0–7
days of age, whereas mice >7 days old hardly did not exhibit any
symptoms (14). A similar situation
possibly exists in adult humans, which raises various questions,
including the following: Whether EV71 may be transmitted from
pregnant women with occult EV71 infection to the fetuses; the
pathogenic damage that occurs to the fetus when the mother is
infected with EV71; and whether the neutralizing antibody for EV71
from a mother may protect her infant.
In the present study, one EV71 strain from a patient
with HFMD exhibiting severe neurologic symptoms was isolated and
characterized. The strain of EV71 was identified as subgenotype
C4a, and was used as the C4 subgenotype has been indicated to be
the most prevalent EV71 subgenotype in mainland China (5,15). The
present study aimed to construct an EV71-infected adult murine
model to evaluate the influence of active EV71 infection in adult
mice. Female adult ICR mice were challenged twice with isolated
EV71; first by hydrodynamic injection (HI) into their tails and
secondly via intraperitoneal injection (IP) as a booster. EV71
maintained a significantly higher titer with HI compared with IP
challenge. Two successive high titer EV71 challenges induced
mortality in the majority of the adult mice. Mice that survived
were identified to have impaired fertility, and their litter size
was significantly reduced compared with the control group. The
antibody directed against the VP1 conserved region of EV71
persisted in female murine sera at a high titer for >2 years
after challenge. The maternal antibody in the sera of the offspring
murine also persisted for ~1 year at a moderate titer and
disappeared after ~2 years.
Materials and methods
Ethics statement
Approval for the present study was obtained from the
Ethics Committee of the Shaoxing People's Hospital (Shaoxing,
China) and informed written consent was obtained from all patients.
All animal protocols were approved by the Ethics Committee of the
Institute of Animal Care of Zhejiang University (Shaoxing, China)
and animals were cared for in accordance with institutional
guidelines of the Shaoxing People's Hospital. Specimens were
confirmed to be positive for EV71 infection by evaluating the VP1
conserved region by polymerase chain reaction (PCR) analysis
(5).
Cell lines and viruses
Vero African green monkey kidney cells and RD cells
were kindly provided by Professor Bo Zhang (Wuhan Institute of
Virology, Chinese Academy of Sciences, Wuhan, China), and were
cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.), 100
U/ml penicillin and 100 µg/ml streptomycin at 37°C in an atmosphere
containing 5% CO2. A total of 300 filtered swab samples from 3- to
5-year-old children (one sample per child; sex ratio was 1:1) with
HFMD were collected in Shaoxing, China, between 2012 and 2013 and
were added to the wells with cultured Vero cells (2 wells per
sample, ~106 cells per well) as previously described
(5,15). EV71 strains in the culture
supernatant were harvested by thawing and refreezing based upon the
appearance of cytopathic effects, such as pyknosis, turning round,
and floating in the medium. Following propagation by cell culture
and identification using the plaque assay (described below), the
EV71 virus stock was adjusted to 1.0×107 pfu and used as
the parental virus in all assays (16).
Serum cytokine and chemokine
detection
Cytokines and chemokines in the sera (10 µl) from
EV71-infected mice were quantitatively measured by FLEXMAP 3D
quantification (Luminex Corporation, Austin, TX, USA) as described
previously (17). A series of
cytokines, including interleukin (IL) 1-α, 1β, 2, 3, 4, 5, 6, 10,
17F and 21, 12p40, 12p70, 17A, 13, 23p19, interferon-γ-inducible
protein 10 (IP-10), keratinocyte chemoattractant (KC)/chemokine
C-X–X moftif ligand 1 (CXCL1), interferon-γ, granulocyte-macrophage
colony-stimulating factor, and tumor necrosis factor (TNF)-α,
eotaxin and monocyte chemoattractant protein (MCP)-1, were detected
using Milliplex MAP kits (cat. no. RCYTO-80K; EMD Millipore,
Billerica, MA, USA), which were used in combination as described
previously (17). The mean of ≥2
independent measurements was calculated using xPONENT software
version 3.1 (Luminex Corporation).
Plaque assay
Viruses in samples were routinely amplified by
passaging the swab-derived virus through the RD cells for one round
(24 h) prior to reinfection with Vero cells for the plaque assay.
Virus stock was produced by harvesting the supernatant of
EV71-infected-Vero cells. Virus titer and morphology were
determined via a bilayer plaque assay (16). A series of 1:10 dilutions were
prepared by diluting 15 ml virus stock with 135 ml DMEM
supplemented with 10% FBS, and 100 ml of each dilution was seeded
into each well of a 6-well plate containing confluent Vero cells
(1×106 cells/well; plated 1 day in advance) as described
previously (18). Plates were
incubated at 37°C for 1 h before the first layer of 1.2% agar was
added. After 72 h of incubation at 37°C, a second layer of 1.2%
agar containing the neutral red pH indicator was added. Following
incubation at 37°C for 12–24 h, images of the plaques were captured
and the number of formed plaques was determined as described
previously (16,18). The viral titer was calculated as
pfu/ml. The limit of detection was 10 pfu/ml.
Determination of viral titers in the
tissue and serum samples of infected mice
A total of 100 female ICR mice (8–10 weeks old;
20.36±1.31 g) were purchased from the Model Animal Research Center
of Nanjing University (Nanjing, China) and maintained under
specific pathogen-free conditions (22–26°C, 40–70% humidity, 10 h
light and 14 h dark per day) in the Central Animal Laboratory of
Shaoxing Hospital of Zhejiang University. Mice were housed in cages
with a bevel allowing free access to food and water. A total of 12
mice (6 per group) were challenged with l ml EV71 (1×107
pfu/ml) via HI or IP. Mice were sacrificed by cervical dislocation
24 or 48 h post injection (19,20).
Blood samples were collected from the orbital vein, and organs and
tissues were harvested after systemic perfusion with 50 ml sterile
PBS. The organs and tissues were weighed and then homogenized in
PBS using a mechanical homogenizer, and disrupted by three
freeze-thaw cycles after sacrifice, as described previously
(21). Subsequently, viral titers in
the supernatants of purified homogenates of the liver, brain,
kidney, heart, spleen and lung, together with blood, were
quantified by reverse transcription-quantitative PCR (RT-qPCR)
analysis.
RNA extraction and RT-qPCR
For RT-qPCR, viral RNA was extracted from fresh
murine tissue homogenates, the blood, and throat and anal samples
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.).
Following digestion with DNase I, extracted RNA was reverse
transcribed into cDNA using a High-Capacity cDNA Reverse
Transcription kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.) and oligo-d(T) primers according to the manufacturer's
instructions. The following primers were designed to target the VP1
conserved region of EV71: Forward, 5′-GCAGCCCAAAAGAACTTCAC-3′ and
reverse, 5′-ATTTCAGCAGCTTGGAGTGC-3′. RT-qPCR was performed for 40
cycles as follows: 95°C for 5 sec, 55°C for 15 sec and 72°C for 9
sec using an Mx3005P machine (Agilent Technologies, Inc., Santa
Clara, CA, USA) as previously described (5,15). The
final reaction volume was 25 µl, including 1 µl each of forward and
reverse primers, 5 µl 5X reaction buffer, 5 µl sample as template,
1 µl DNA polymerase and 12 µl water. An EV71 reporter virus
(GenBank accession no. JN256064; provided by Professor Bo Zhang,
Wuhan Institute of Virology, Chinese Academy of Science, China)
with whole EV71 sequence was used as a standard to calculate the
EV71 copy number as described previously (18).
Mouse EV71 challenge
A total of 1 ml Vero culture supernatant with
isolated EV71 (1.0×107 pfu) was quickly injected into
the tail veins of mice to maintain high pressure, or into the
enterocoelia, as described previously (20). The control group (6 mice) received
the same volume of DMEM. A booster injection was administered 2
weeks following the first injection. Blood samples were harvested
24 and 48 h after the last immunization.
For the lethalality assay, four groups of female ICR
mice (6–8 weeks old, 8 mice per group) were challenged with
isolated EV71 using HI or IP. Group 1: Primer and boost challenge
with active EV71; group 2: Primer challenge with active EV71 and
boost with inactive EV71; group 3: Primer challenge with inactive
EV71 and boost with active EV71; group 4: Primer challenge with
inactive EV71 and boost with inactive EV71. Survival rates were
determined using GraphPad Prism software (version 5; GraphPad Prism
Software, Inc., La Jolla, CA, USA).
For the reproductive damage assay, 2 groups of
female mature ICR mice were challenged with active EV71 (24 mice)
or DMEM as control (6 mice) by HI and then raised with male adult
ICR mice to conceive. Booster challenge with active EV71 or DMEM
was administered to the female mice via IP two weeks post primer
challenge. Control group: Primer and boost challenge both with
DMEM. ‘Once’ group: Primer challenge with active EV71 and boost
with DMEM. ‘Twice’ group: Primer and boost challenge both with
active EV71. The litter size of all surviving female mice after
birth was noted and used for the assessment of reproductive
ability.
For evaluation of the protectiveness of maternal
antibody, 1-day-old ICR neonatal mice (weight, 1.8–2.0 g) born from
the EV71-challenged and mock mice were challenged with 50 µl active
EV71 or DMEM via intracranial injection as previously described
(22,23). All mice were monitored for clinical
symptoms, paralysis, and death for at least 14 days post-injection.
In addition, for dynamic regularity of EV71-VP1-IgG, the
EV71-challenged maternal mice and their second generation mice were
randomly allocated into 2 groups of 10 mice, and were monitored
daily for clinical signs of infection, including ruffled hair,
hunched back, paralysis and mortality until the end of the assay
(120 weeks), as described previously (22).
Statistical analysis
Statistical analysis was performed using GraphPad
Prism software (version 5; GraphPad Prism Software, Inc.). The
significance of all differences in multiple comparisons were
determined using the Student's t-test. Results are presented as the
mean ± standard deviation of three independent experiments.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Characterization of the isolated EV71
strain
The EV71 strain used as virus stock in the present
study was isolated from clinical samples from children with HFMD.
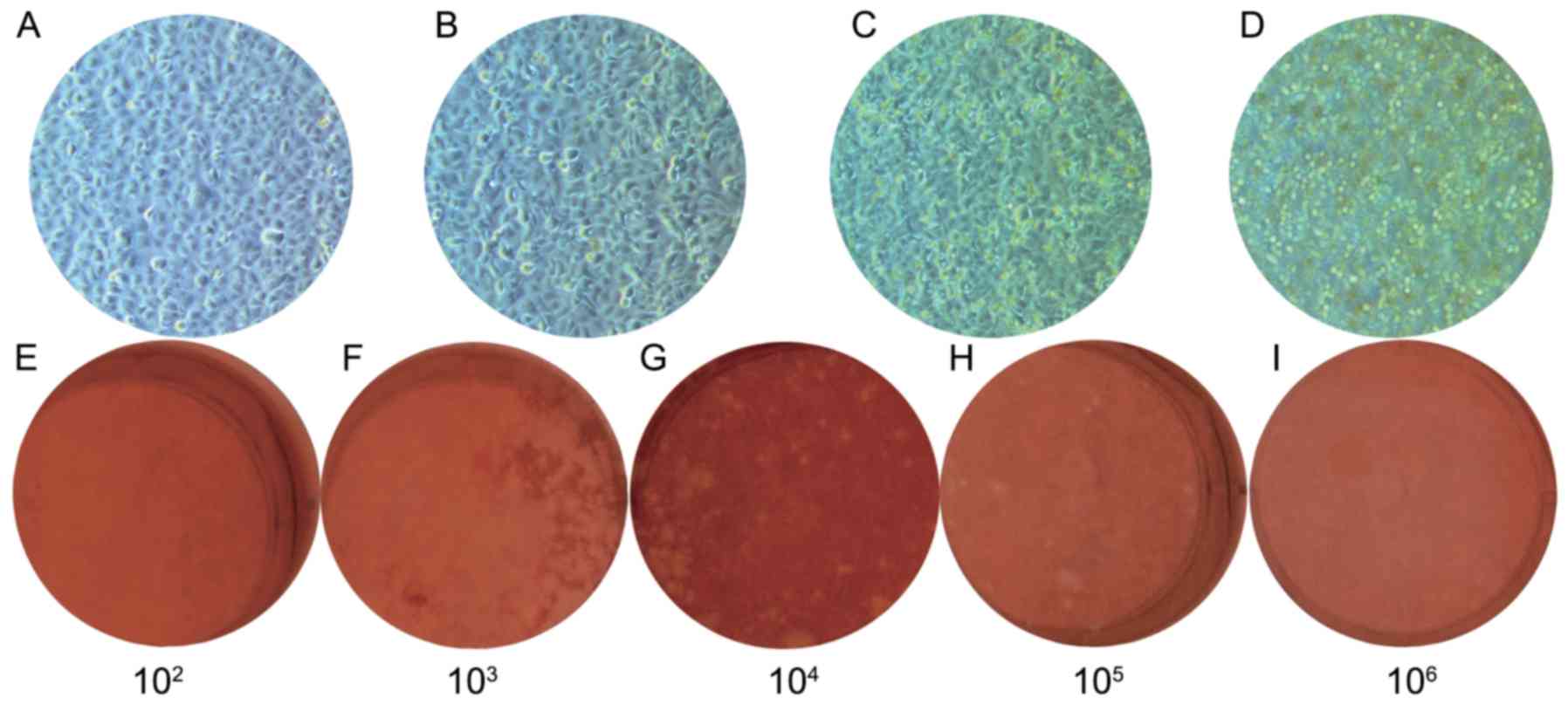
The viruses infected Vero cells, inducing typical CPE as shown in
Fig. 1A-D. In a previous study by
the present authors, the VP1 sequence of this isolated EV71 strain
was amplified, sequenced, aligned, and was identified to be of the
C4a genotype (5,15).
Virus stock was produced by harvesting the
supernatant of EV71-infected Vero cells. Virus titer and morphology
were determined using a bilayer plaque assay (Fig. 1E-I). A total of 21 plaques were
identified in the well with a 1×105 dilution (Fig. 1H). The virus solution with a titer of
2.1×107 was adjusted to 1×107 and used as the
parental virus in all assays.
EV71 challenge with HI produces a
higher viral titer in the tissue compared with IP
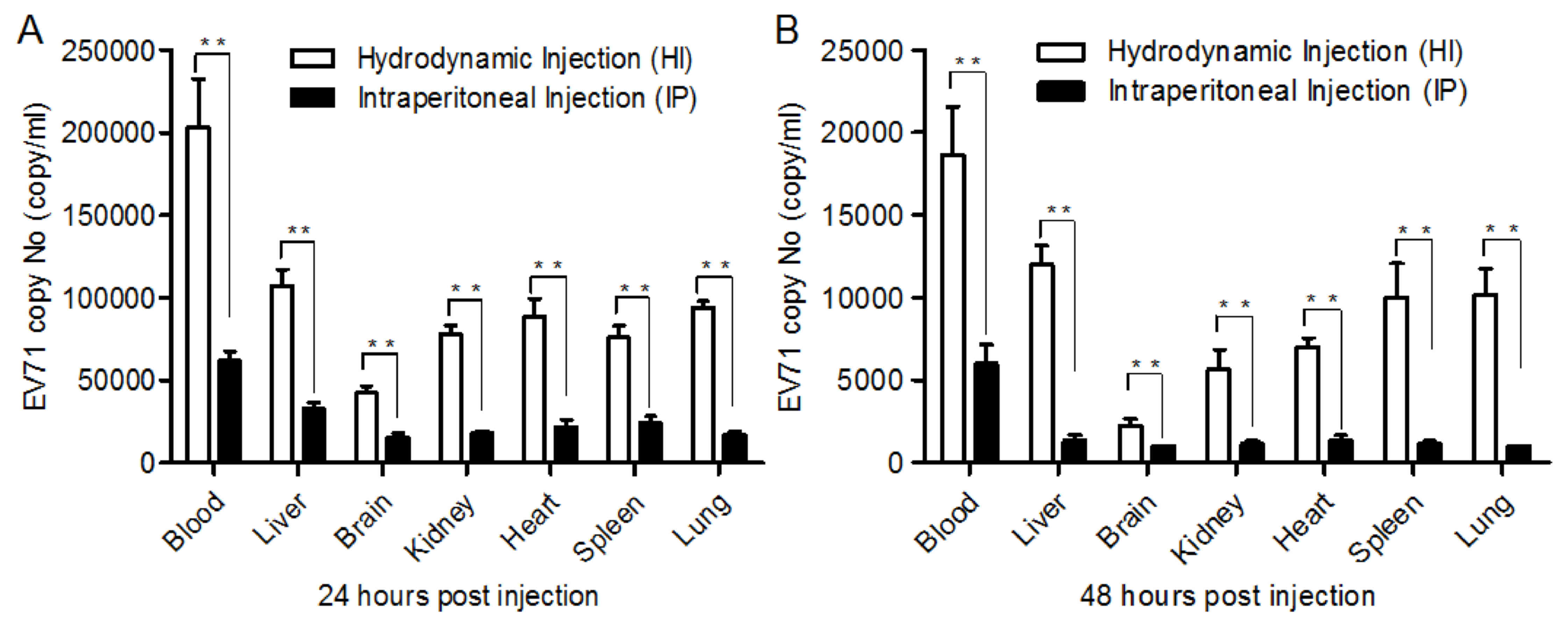
In order to achieve a higher titer EV71 in the
tissue of mice, administration of EV71 by HI and IP was compared.
Tissue samples were homogenized in sterile PBS, disrupted by three
freeze-thaw cycles and centrifuged at 14,000 × g. EV71 copy number
was calculated by RT-qPCR. As indicated in Fig. 2A and B, administration via HI
resulted in a significantly increased EV71 titer compared with IP
at 24 and 48 h after injection (P<0.05), which indicated that HI
was preferable for acquiring a high viral titer in the tissue and
blood.
EV71 challenge is fatal to adult
mice
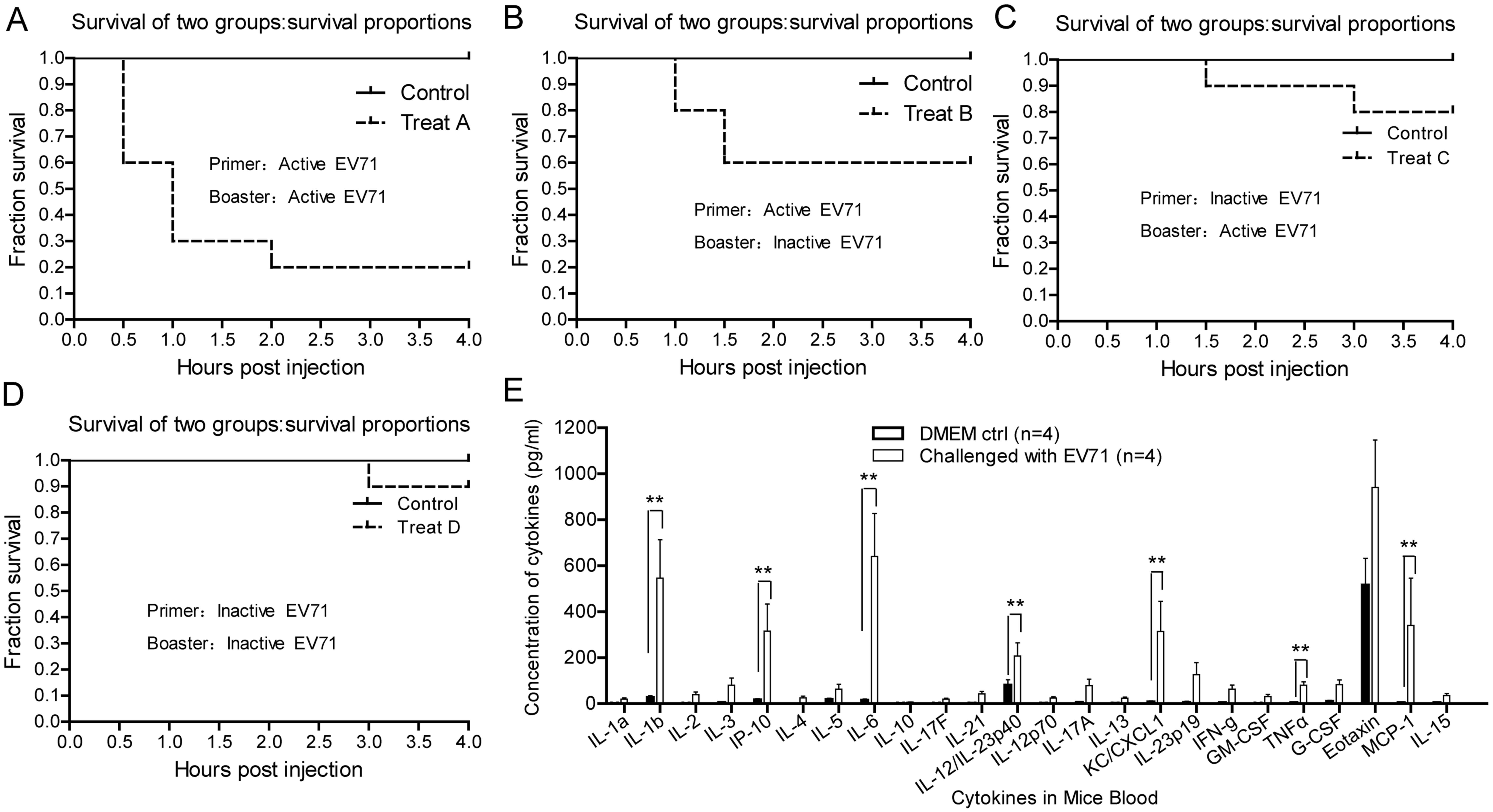
Groups of female ICR mice were challenged with
isolated EV71 via HI or IP. Four h post booster injection, a total
of 80% of mice succumbed to EV71 infection as a result of primer
and booster challenge (Fig. 3A), 40%
of mice succumbed to EV71 when first challenged with active EV71
and then receiving inactive EV71 as a booster (Fig. 3B), 80% of mice survived when first
challenged with inactive EV71 and then receiving active EV71 as a
booster (Fig. 3C) and 90% of mice
survived when treated with inactive EV71 as a primer and booster
(Fig. 3D). It was noted that
mortality was highest half an hour post booster injection.
To further investigate why EV71 challenge induced
sudden fatality the blood samples from mice that survived primer
and booster challenge with active EV71 virus alone were collected
and subjected cytokine/chemokine detection. IL-1b, IP-10, IL-6,
IL-12, KC/CXCL1, IL-23p19, TNF-α and MCP-1 were significantly
upregulated in EV71-challenged mice compared with the
non-challenged control group (P<0.01; Fig. 3E). These results suggest that the
fatality of the challenged mice was promoted by potent
immunological stress, not the infectivity and virulence of EV71
Furthermore, these findings suggest that active EV71 possesses high
antigenicity compared with inactive EV71, which triggers
immunoreaction.
EV71 challenge impairs the
reproductive capacity of female mice
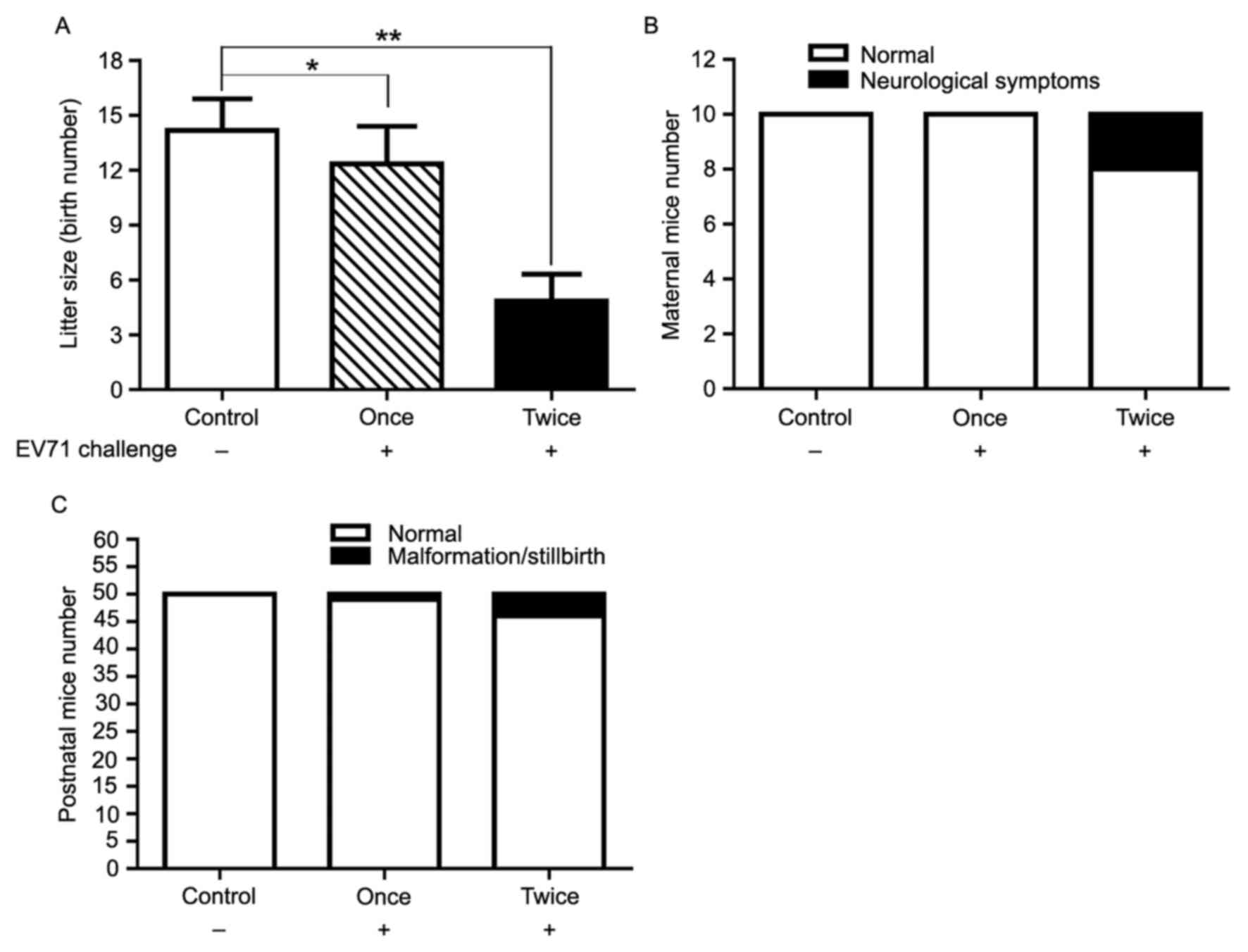
Female mature ICR mice in the ‘Twice’ group were
challenged with active EV71 by HI before and after pregnancy. A
large proportion of the mice succumbed to EV71 infection. Mice that
survived were housed under specific pathogen-free conditions in
individual ventilated cages until parturition. Mice in the ‘Twice’
group gave birth to a mean of 5 suckling mice, whereas litter size
in the ‘Once’ group was 12 and in the control group was 14. Active
EV71 administered as a primer and/or booster was identified to
significantly impair the reproductivity of female mice compared
with the control group (P<0.05; Fig.
4A). A total of 20% of survivors challenged twice with EV71
exhibited neurologic symptoms, including torticollis and walking
disorder (Fig. 4B). A number of
neonatal mice produced from female mice challenged with active EV71
presented with abnormalities, including stillbirth and dwarfism.
The proportion of abnormalities were in line with the challenge
number (Fig. 4C).
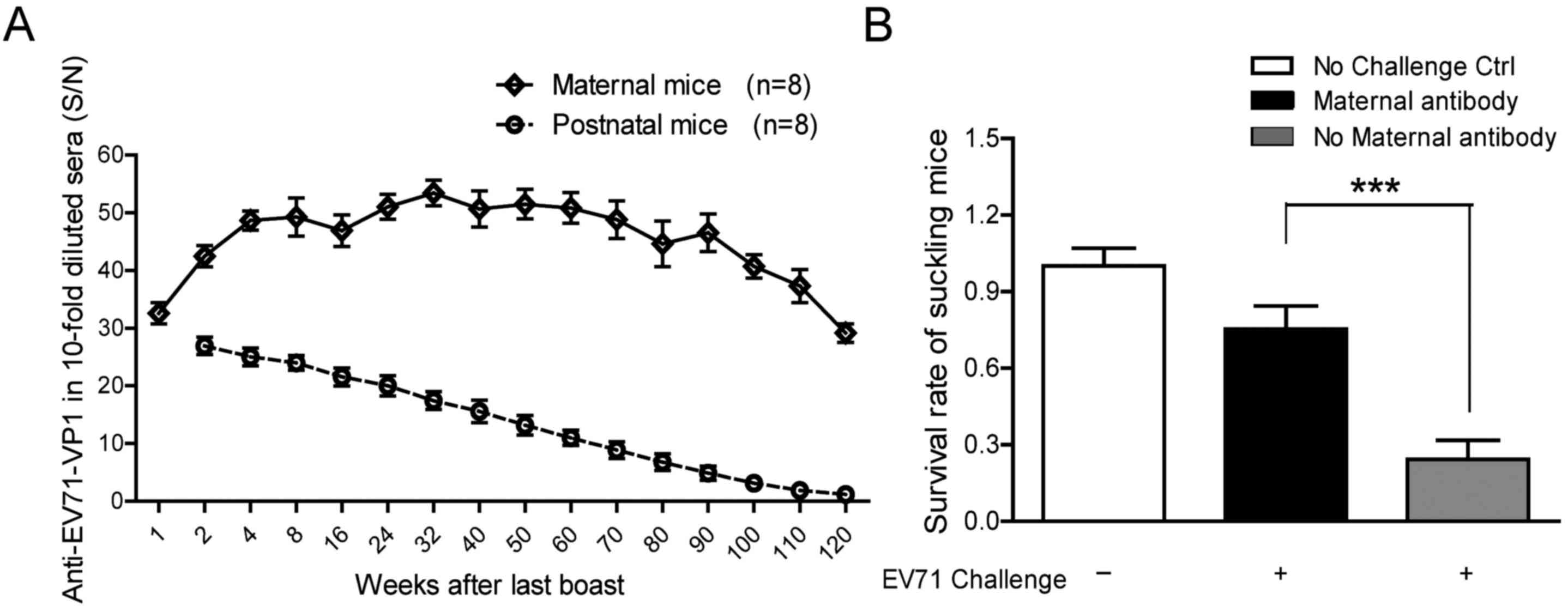
EV71 antibody is maintained at a high
titer for a longer duration after challenge with EV71 twice in
maternal and postnatal mice
Blood samples were collected from 1–120 after the
booster injection and EV71 antibody levels were detected using an
ELISA kit. Maternal EV71-VP1 antibody titer reached a peak ~4 weeks
after the booster injection and persisted at a high level for ~1
year, following which it slowly declined until the end of
monitoring (Fig. 5A). Maternal
antibodies in the blood samples taken from postnatal mice after
birth were also monitored. As illustrated in Fig. 5A, the level of EV71-VP1 antibody
gradually declined after birth and was barely detectable at the end
of monitoring. This maternal antibody was indicated to be
protective, as 3-day-old suckling mice with the maternal antibody
were challenged with active EV71 by intracranial injection. Mice
that were challenged with EV71 and exhibited the maternal antibody
had a significantly increased survival rate compared with mice with
no maternal antibody; >70% of these mice were protected from
EV71-induced fatality compared with 20% of mice without the
maternal antibody (P<0.05; Fig.
5B).
Discussion
HFMD typically occurs in children <5 years old,
and EV71 and coxsackie virus A16 have been identified as the
primary causes of HFMD (2). Serious
neurological complications and fatalities are associated with EV71
(24). Due to a lack of effective
treatment options, controlling EV71 epidemics has predominantly
relied upon the formation of EV71 vaccines (11). Therefore, effective vaccines that
protect against EV71-associated HFMD or herpangina are important
(25). Currently, two monovalent
EV71 vaccines have been marketed in mainland China (2). However, clinical data concerning their
effectiveness is not yet available.
Despite the extensive research undertaken since the
discovery of EV71, the following issues require further
investigation: The potential harm to a pregnant woman with EV71
latent infection and her fetus; and the length of time that
anti-EV71 antibody remains effective after vaccination. In the
present study, an isolated C4a EV71 strain was used to challenge
ICR mice. The results indicated that EV71 was maintained a higher
viral titer (calculated by RT-qPCR) in murine tissue and blood when
challenged via HI as displayed in Fig.
2A. Furthermore, >50% of the challenged mice succumbed to
EV71 infection when challenged with EV71 infection twice. In
addition, abnormal symptoms were demonstrated in the survivors and
their progenies. Based on the serum chemokine/cytokine detection
performed, potent immunological stress was likely responsible for
this lethality and teratogenicity, not the infectivity and
virulence of EV71. Active EV71 challenge resulted in more serious
lethality and teratogenicity compared with inactive EV71 challenge.
However, the mechanisms underlying this effect of EV71 require
further research. Furthermore, the neutralizing antibody in
maternal mice and their offspring was present over a long period of
time at a high titer when the mice were challenged twice. These
results indicate that EV71 exhibits notable immunogenicity,
suggesting its suitability as a potential vaccine candidate. In
addition, the presence of the maternal antibody in the fetus was
revealed to be protective against EV71-induced mortality, which
suggests that children <5 years old and women of child-bearing
age may benefit from receiving EV71 vaccination.
The majority of previous studies have reported that
adult mice are not susceptible to EV71 infection, with only
suckling mice able to be infected by EV71. To the best of our
knowledge, the present study is the first to report that active
EV71 challenge inhibits the reproductive capacity of female mice,
and induces neurologic symptoms in adults and offspring. In humans,
occult EV71 infection is common in adults without any symptoms
(2), who may act as a reservoir host
for susceptible infants. Furthermore, if a woman of childbearing
age is infected with EV71, little is known about whether this
infection would harm to her fetus. Thus, further studies warranted
to investigate this effect of EV71 in humans.
In conclusion, a novel challenge method of EV71,
hydrodynamic injection, was established in the present study. Due
to the higher titer of EV71 in the tissue of challenged mice, it
was demonstrated to be more effective than intraperitoneal
injection. Generally speaking, EV71 infection was only fatal to
suckling mice within 7 days of birth. In the present study, a
higher rate of teratogenesis occurred in adult mice following EV71
infection. In particular, the twice-active EV71 challenge may cause
mortality in adult mice, and surviving mice have a significantly
lower reproductive capacity. These results suggest that occult EV71
infection in humans, in particular women of childbearing age, may
pose a serious health risk.
Acknowledgements
The present study was supported in part by the
Medical Scientific Research Foundation of Zhejiang Province (grant
no. 2013KYA208) and the Science Technology Bureau of Shaoxing,
Zhejiang province of China (grant nos. 2013B70063 and
2012B70063).
References
|
1
|
Ye X, Fan C, Ku Z, Zuo T, Kong L, Zhang C,
Shi J, Liu Q, Chen T, Zhang Y, et al: Structural basis for
recognition of human enterovirus 71 by a bivalent broadly
neutralizing monoclonal antibody. PLoS Pathog. 12:e10054542016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gui J, Liu Z, Zhang T, Hua Q, Jiang Z,
Chen B, Gu H, Lv H and Dong C: Epidemiological characteristics and
spatial-temporal clusters of hand, foot and mouth disease in
zhejiang province, China, 2008–2012. PLoS One. 10:e01391092015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ho M, Chen ER, Hsu KH, Twu SJ, Chen KT,
Tsai SF, Wang JR and Shih SR: An epidemic of enterovirus 71
infection in Taiwan. Taiwan Enterovirus Epidemic Working Group. N
Engl J Med. 341:929–935. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xu W, Liu CF, Yan L, Li JJ, Wang LJ, Qi Y,
Cheng RB and Xiong XY: Distribution of enteroviruses in
hospitalized children with hand, foot and mouth disease and
relationship between pathogens and nervous system complications.
Virol J. 9:82012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Qin B, Zhang J, Xie W, Liu X, He T, Chen J
and Dong X: Development of a specific latex agglutination test to
detect antibodies of enterovirus 71. Arch Med Res. 46:580–587.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Qing J, Wang Y, Sun Y, Huang J, Yan W,
Wang J, Su D, Ni C, Li J, Rao Z, et al: Cyclophilin A associates
with enterovirus-71 virus capsid and plays an essential role in
viral infection as an uncoating regulator. PLoS Pathog.
10:e10044222014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu CC, Chow YH, Chong P and Klein M:
Prospect and challenges for the development of multivalent vaccines
against hand, foot and mouth diseases. Vaccine. 32:6177–6182. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liang ZL, Mao QY, Wang YP, Zhu FC, Li JX,
Yao X, Gao F, Wu X, Xu M and Wang JZ: Progress on the research and
development of inactivated EV71 whole-virus vaccines. Hum Vaccin
Immunother. 9:1701–1705. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ren J, Wang X, Zhu L, Hu Z, Gao Q, Yang P,
Li X, Wang J, Shen X, Fry EE, et al: Structures of Coxsackievirus
A16 capsids with native antigenicity: Implications for particle
expansion, receptor binding and immunogenicity. J Virology.
89:10500–10511. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chang J, Li J, Wei W, Liu X, Liu G, Yang
J, Zhang W and Yu XF: Determinants of EV71 immunogenicity and
protection against lethal challenge in a mouse model. Immunol Res.
62:306–315. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ng Q, He F and Kwang J: Recent Progress
towards Novel EV71 anti-therapeutics and vaccines. Viruses.
7:6441–6457. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li X, Fan P, Jin J, Su W, An D, Xu L, Sun
S, Zhang Y, Meng X, Gao F, et al: Establishment of cell lines with
increased susceptibility to EV71/CA16 by stable overexpression of
SCARB2. Virol J. 10:2502013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang YF, Chou CT, Lei HY, Liu CC, Wang SM,
Yan JJ, Su IJ, Wang JR, Yeh TM, Chen SH and Yu CK: A mouse-adapted
enterovirus 71 strain causes neurological disease in mice after
oral infection. J Virol. 78:7916–7924. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shih SR, Li YS, Chiou CC, Suen PC, Lin TY,
Chang LY, Huang YC, Tsao KC, Ning HC, Wu TZ and Chan EC: Expression
of capsid [correction of caspid] protein VP1 for use as antigen for
the diagnosis of enterovirus 71 infection. J Med Virol. 61:228–234.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bo HT: tCJkZJhJZjHXnFLjQ: A study on
genetic characteristics of Enterovirus 71 of hand-foot-mouth
disease in Shaoxing city Zhejiang. Prev Med. 26:42014.
|
|
16
|
Li XD, Li XF, Ye HQ, Deng CL, Ye Q, Shan
C, Shang BD, Xu LL, Li SH, Cao SB, et al: Recovery of a chemically
synthesized Japanese encephalitis virus reveals two critical
adaptive mutations in NS2B and NS4A. J Gen Virol. 95:806–815. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen J, Zhao Y, Zhang C, Chen H, Feng J,
Chi X, Pan Y, Du J, Guo M, Cao H, et al: Persistent hepatitis C
virus infections and hepatopathological manifestations in
immune-competent humanized mice. Cell Res. 24:1050–1066. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shang B, Deng C, Ye H, Xu W, Yuan Z, Shi
PY and Zhang B: Development and characterization of a stable eGFP
enterovirus 71 for antiviral screening. Antiviral Res. 97:198–205.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Qin B, Tu C, Zhang B, He T, Fu L and Xu W:
A modified murine model based on hydrodynamic injection for the
analysis of chronic human hepatitis B virus infection. Mol Med Rep.
8:1677–1682. 2013.PubMed/NCBI
|
|
20
|
Qin B, Budeus B, Cao L, Wu C, Wang Y,
Zhang X, Rayner S, Hoffmann D, Lu M and Chen X: The amino acid
substitutions rtP177G and rtF249A in the reverse transcriptase
domain of hepatitis B virus polymerase reduce the susceptibility to
tenofovir. Antiviral Res. 97:93–100. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen IC, Wang SM, Yu CK and Liu CC:
Subneutralizing antibodies to enterovirus 71 induce
antibody-dependent enhancement of infection in newborn mice. Med
Microbiol Immunol. 202:259–265. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li P, Yue YY, Song NN, Li ZH and Meng H:
Study on BALB/c mice with EV71 infection by diverse inoculation
routes. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi.
27:14–17. 2013.(In Chinese). PubMed/NCBI
|
|
23
|
Han JF, Yu N, Pan YX, He SJ, Xu LJ, Cao
RY, Li YX, Zhu SY, Zhang Y and Qin ED: Phenotypic and genomic
characterization of human coxsackievirus A16 strains with distinct
virulence in mice. Virus Res. 179:212–219. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Teng S, Wei Y, Zhao SY, Lin XY, Shao QM
and Wang J: Intestinal detoxification time of hand-foot-and-mouth
disease in children with EV71 infection and the related factors.
World J Pediatr. 11:380–385. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Klein MH: EV71 vaccines: A first step
towards multivalent hand, foot and mouth disease vaccines. Expert
Rev Vaccines. 14:337–340. 2015. View Article : Google Scholar : PubMed/NCBI
|