Introduction
Stroke is one of the most common causes of mortality
and a leading cause of disability worldwide (1). Survivors are typically afflicted by
certain levels of functional impairment, including motor, sensory
and cognitive dysfunction (2–4).
Evidence suggests that up to 64% of patients exhibit a certain
degree of cognitive impairment, with 20–30% of patients
demonstrating dementia at 3 months following stroke (5–7).
Learning and memory deficits are among the most common cognitive
impairments and severely affect patients' daily activities and
quality of life, which leads to them becoming a significant burden
on their families and society (8,9).
Acupuncture is one of the most commonly used and
important therapies in traditional Chinese medicine and has been
applied clinically for over a millennium (10). Electroacupuncture (EA), a combination
of electrical stimulation with acupuncture, has demonstrated
efficacy in alleviating cognitive impairments and improving
learning and memory in patients and animal models post-stroke
(11–14). However, the exact mechanism of its
benefits on impaired cognition remains unclear.
Neuroinflammation is a prime pathological factor in
stroke, involving a number of complex cellular processes and
activities, including astrocyte and microglial proliferation and
the production of inflammatory cytokines, including tumor necrosis
factor (TNF)-α and interleukin (IL)-1β (15,16).
Previous animal studies suggested a mechanistic link between
neuroinflammation and cognitive function following traumatic brain
injury (TBI) and also in Alzheimer's disease (AD) models (17–19). In
addition, hippocampal neuroinflammation is responsible for
mediating cognitive dysfunction in the aging brain, in
post-cerebral ischemia, and also in AD (20–22).
Cholinergic anti-inflammatory pathways are critical
regulators of inflammation. These pathways primarily involve the
interaction of vagus nerve cholinergic signaling with α7 nicotinic
acetylcholine receptors (α7nAChRs) on immune cells, which leads to
inhibition of pro-inflammatory cytokine production, thereby
preventing excessive inflammatory responses (23). The α7nAChR serves as an important
signaling receptor in cholinergic anti-inflammatory pathways and is
closely associated with learning and memory (24–26).
Counteracting neuroinflammatory processes via cholinergic
anti-inflammatory signaling prevents progressive tissue damage in
the brain following stroke (27). EA
is known to act in a similar manner by inhibiting neuroinflammation
and preventing development of cognitive dysfunction in patients
post-stroke (14,28). In addition, a previous study
demonstrated that treatment with EA prior to ischemia/reperfusion
(I/R) injury is neuroprotective because EA prevents the
downregulation of α7nAChR in neurons within the ischemic penumbra
(29). The present study aimed to
elucidate whether EA ameliorates learning and memory through
α7nAChR-mediated inhibition of neuroinflammation in a rat model of
focal cerebral I/R injury.
Materials and methods
Animals
A total of 65 male Sprague-Dawley (SD) rats (250–280
g, ages 10–12 weeks) were provided by Shanghai SLAC Laboratory
Animal Co., Ltd. [Laboratory Animal Use Certificate no. SCXK (SH)
2012–0002] and housed under controlled conditions with a 12-h
light/dark cycle, 22±2°C temperature and 55±15% humidity for at
least 1 week prior to surgery and treatment. All animals were
allowed ad libitum access to standard rodent food and water.
All animal treatments and experiments were conducted in strict
accordance with the National Institutes of Health Guide for the
Care and Use of Laboratory Animals, and were approved by the
Institutional Animal Care and Use Committee of Fujian University of
Traditional Chinese Medicine (FUTCM; Fujian, China).
Cerebral I/R injury model
Transient focal cerebral ischemia was established
via middle cerebral artery occlusion (MCAO) as previously described
(30) in 55 rats. Briefly, rats were
anesthetized with an intraperitoneal injection of 10% chloral
hydrate (3 ml/kg body weight; catalogue no. 30037516; Changzhou
Haituo Experimental Instrument., Co., Ltd., Changzhou, China)
diluted with 0.9% saline in a sterilized surgical site. Following a
midline incision in the neck, the left common carotid artery (CCA),
left external carotid artery (ECA) and the internal carotid artery
(ICA) were carefully exposed and dissected. The left middle
cerebral artery (MCA) was occluded by introducing an embolus
through the ICA. To occlude the origin of the left MCA, a nylon
filament (0.36±0.02 tip diameter; Guangzhou Jialing Co. Ltd.,
Guangzhou, China) was inserted (~20 mm) from the ECA and ICA into
the MCA. Reperfusion was accomplished by withdrawing the filament
after 2 h of occlusion-mediated ischemia. The incision was sutured
and sterilized. During and following surgery, the internal
temperature of the animals was maintained at 37°C using a heating
pad. For rats in the control group (n=10), the arteries were
similarly exposed, but not immobilized in a blinded manner as
previously described (30). The
neurological deficits of the rats were scored as follows: A score
of 0 represented no neurological deficit, 1 indicated mild deficits
(failure to fully extend the right forepaw), 2 (circling to the
right) and 3 (falling to the right) indicated moderate deficits,
and a score of 4 represented severe deficits (complete loss of
walking ability). Rats that received MCAO and a score of 0 or 4
were excluded from the experiment.
Groups
When the MCAO model was established, the rats were
randomly assigned into four groups according to neurological
deficit scores: i) MCAO group (n=13); ii) MCAO + EA group (EA
group, n=13); iii) MCAO + EA + normal saline (EA + NS group, n=12)
and iv) MCAO + EA + methyllycaconitine group (EA + MLA group,
n=12). Control rats received surgery without artery ligation
(n=10). Therefore, there were five groups in total in the present
study.
EA treatment
Rats were administered EA for 30 min daily for 7
days, starting 2 days after I/R surgery. The acupuncture needles
(0.3 mm diameter) were inserted at a depth of 2–3 mm into the
Baihui (DU 20) and Shenting (DU 24) acupoints, which are commonly
used to treat post-stroke cognitive impairment in China (31). Electrical stimulation was then
generated using the EA apparatus (Model G6805; Shanghai Medical
Instrument Factory, Shanghai, China) with disperse-dense waves of a
frequency of 2–10 Hz and an intensity of 2–4 mA. The rats in the
control and MCAO groups remained in their cages without special
intervention.
Drug administration
Methyllycaconitine (MLA; 5 mg/kg; Sigma-Aldrich,
Merck KGaA, Darmstadt, Germany) was diluted in 0.9% saline and
administered intraperitoneally 30 min prior to each EA treatment
(32). In the EA + NS group, 0.9%
saline was injected, with a volume equivalent to that of the
methyllycaconitine solution.
Morris water maze
Rats were subjected to the Morris water maze 3 days
following surgery to assess spatial learning and memory as
previously described (33). The
water maze apparatus (Chinese Academy of Sciences, Beijing, China)
consisted of a circular, black-painted pool (diameter, 120 cm;
depth, 50 cm) filled with water tinted with black ink (depth, 30
cm; temperature, 26±2°C). The tank was divided into four equal
quadrants and a video camera attached to a computer was placed
above the center of the tank to record the rats. A fixed, 6-cm
platform was submerged 2 cm below the surface of the water. A
number of visual cues were placed in each quadrant. During the
first set of trials, each rat was placed in the water at four
equidistant locations to the platform. When the rats arrived at the
platform and remained on it for 3 sec they were considered to have
found the platform. When the rats were unable to find the platform
within 90 sec, they were placed on the platform for 10 sec and the
time score was 90 sec. The latency time to find the submerged
platform and the total swimming distance were recorded for 4
days.
Rats were subjected to the Morris water maze test
with the platform removed 7 days following surgery. Rats were
placed in the quadrant located diagonally from the target quadrant
and allowed to swim for a maximum of 90 sec. The frequency of
swimming across the former location of the platform in the target
quadrant was recorded.
Immunohistochemistry
Rats were anesthetized with 10% chloral hydrate by
intraperitoneal injection and perfused transcardially with 0.9%
NaCl followed by 4% paraformaldehyde through the left ventricle.
The brain was removed and fixed in 4% paraformaldehyde at 4°C for
24 h. Samples underwent dehydration with an ethanol gradient, 70,
80 and 90% for 1 h each and 100% ethanol for 20 min, 1 h and a
final 20 min. Following washing with xylene, 20 min and 1 h
followed by 20 min, the specimens were embedded in paraffin and cut
into 5-µm sections (RM2235 slice machine; Leica Microsystems GmbH,
Wetzlar, Germany). Following deparaffinization, antigen retrieval
was accomplished by immersing and boiling the sections in a
Tris-EDTA Buffer (10 mM Tris, 1 mM EDTA, 0.05% Tween-20, pH 9.0;
for α7nAChR) or citrate buffer solution [10 mM Tris, 20% citrate,
pH 6.0; for glial fibrillary acidic protein (GFAP) and microglial
marker Iba1] in a microwave oven.
α7nAChR, GFAP and Iba1 levels were analyzed using
immunohistochemistry assay kits [diaminobenzidine (DAB) kit-0017;
Maixin-Bio, Fujian, China] according to the manufacturer's
protocol. Primary antibodies binding to α7nAChR (cat. no. ab24644;
1:100; Abcam, Cambridge, UK), GFAP (cat. no. 3670; 1:500; Cell
Signaling Technologies, Inc., Danvers, MA, USA), and Iba1 (cat. no.
NB100-1028; 1:250; Novus Biologicals, LLC, Littleton, CO, USA) were
incubated with the sections at 4°C overnight and then the sections
were incubated with secondary antibodies, provided in the DAB
kit-0017, incubated at room temperature for 10 min. The positive
cells were stained brown with DAB. Images were captured using a
fluorescence microscope (DFC310 FX; Leica Microsystems GmbH) at
×400 magnification. Positive cells were counted in four randomly
selected microscopic fields using the Motic Med 6.0 CMIAS pathology
image analysis system (Beihang Motic Inc., Beijing, China).
Western blot analysis
Hippocampal tissues were homogenized in
non-denaturing lysis buffer (Beyotime Institute of Biotechnology
Co., Ltd., Beijing, China; no. P0013B). Tissues were then ground on
ice and incubated for 30 min prior to 10 min centrifugation at
1,465 × g at 4°C to separate the supernatant. Total protein in each
sample was measured using the bicinchoninic acid (BCA) assay. A
total of 50 µg protein was separated on a 10% SDS-PAGE gel and
transferred to a polyvinylidene fluoride (PVDF) membrane. The
membranes were blocked for 2 h with blocking buffer (P0023B;
Beyotime Institute of Biotechnology Co., Ltd.) and then incubated
with primary antibodies targeting α7nAChR (cat. no. ab24644;
1:10,000; Abcam), TNF-α (cat. no. 3707; 1:500; Cell Signaling
Technologies, Inc.), IL-1β (1:200; sc-12742; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) or GAPDH (cat. no. ab8245;
1:8,000; Abcam) at 4°C overnight. Following washing with TBS
containing 0.05% Tween-20, the membranes were incubated with
appropriate horseradish peroxidase-conjugated secondary antibody
(cat. no. 7076 for GAPDH; and cat. no. 7074 for α7nAChR, TNF-α and
IL-1β; 1:5,000; Cell Signaling Technologies, Inc.) for 1 h at room
temperature. Blots were developed using enhanced chemiluminescence
(Beyotime Institute of Biotechnology, Co., Ltd.) and images were
obtained and analyzed using a Bio-Image Analysis System, version
1.42q (Bio-Rad Laboratories, Hercules, CA, USA). The optical
densities of the target protein were normalized against the GAPDH
band and the analysis was replicated 3 times.
Statistical analysis
The experimental results for each group are
expressed as the mean ± standard error of the mean. Statistical
analysis was performed with one-way analysis of variance using the
SPSS package for Windows (Version 18.0; SPSS, Inc., Chicago, IL,
USA), homogeneity of variance with a LSD test, and heterogeneity of
variance with Games-Howell (A) test. P<0.05 was considered to
represent a statistically significant difference.
Results
EA reduces learning and memory
impairment following focal cerebral ischemic injury
Following modeling, the rats in the control group
were of good health with no fatalities. In the model group, 3 rats
died due to epilepsy. A further 3 mortalities were observed in the
EA group, 2 in the EA + NS group and 2 in the EA + MLA group.
Concerning the Morris water maze performance, as presented in
Fig. 1, no significant differences
were detected in swimming speed on days 3, 4, 5, 6 and 7 following
MCAO among the five groups (Fig. 1A;
P>0.05). This result indicates that the MCAO model did not
affect rat motor function in the Morris water maze. Rats in the
MCAO group demonstrated a longer latency time (Fig. 1B) to reach the hidden platform and
passed the platform position fewer times (Fig. 1C and D) in the water maze tests than
did those in the control group. In EA-treated rats, latency time
was significantly reduced and frequency in passing the platform was
increased compared with that in the MCAO group (Fig. 1B and C; P<0.05). However, rats
treated with EA + MLA demonstrated prolonged latency with a
decreased number of times crossing the platform compared with the
EA + NS group.
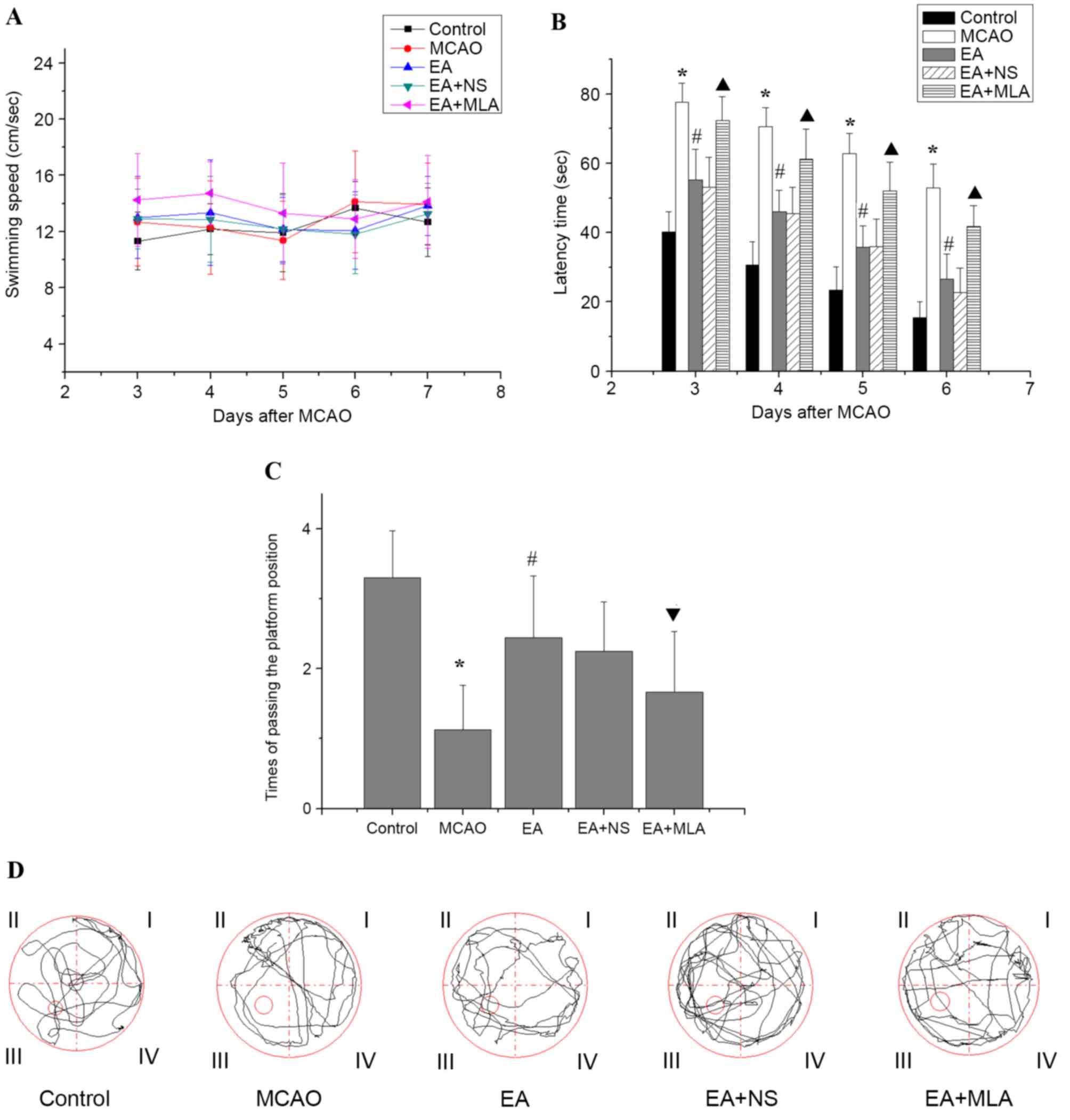 | Figure 1.Effects of EA on the learning and
memory of transient focal cerebral ischemic injured rats in the
water maze. (A) No significant differences in swimming speed were
observed among the control (n=10), MCAO (n=8), EA (n=9), EA + NS
(n=9) and EA+ MLA (n=9) groups up to day 7. (B) The latency time to
reach the hidden platform, (C) the number of times the rats passed
the platform position and (D) typical traces during the test.
*P<0.01 vs. control, #P<0.01 vs. MCAO,
▲P<0.01 vs. EA + NS and ▼P<0.05 vs. EA
+ NS. Data are expressed as mean ± standard error of the mean. EA,
electroacupuncture; MCAO, middle cerebral artery occlusion; NS,
normal saline; MLA, methyllycaconitine. |
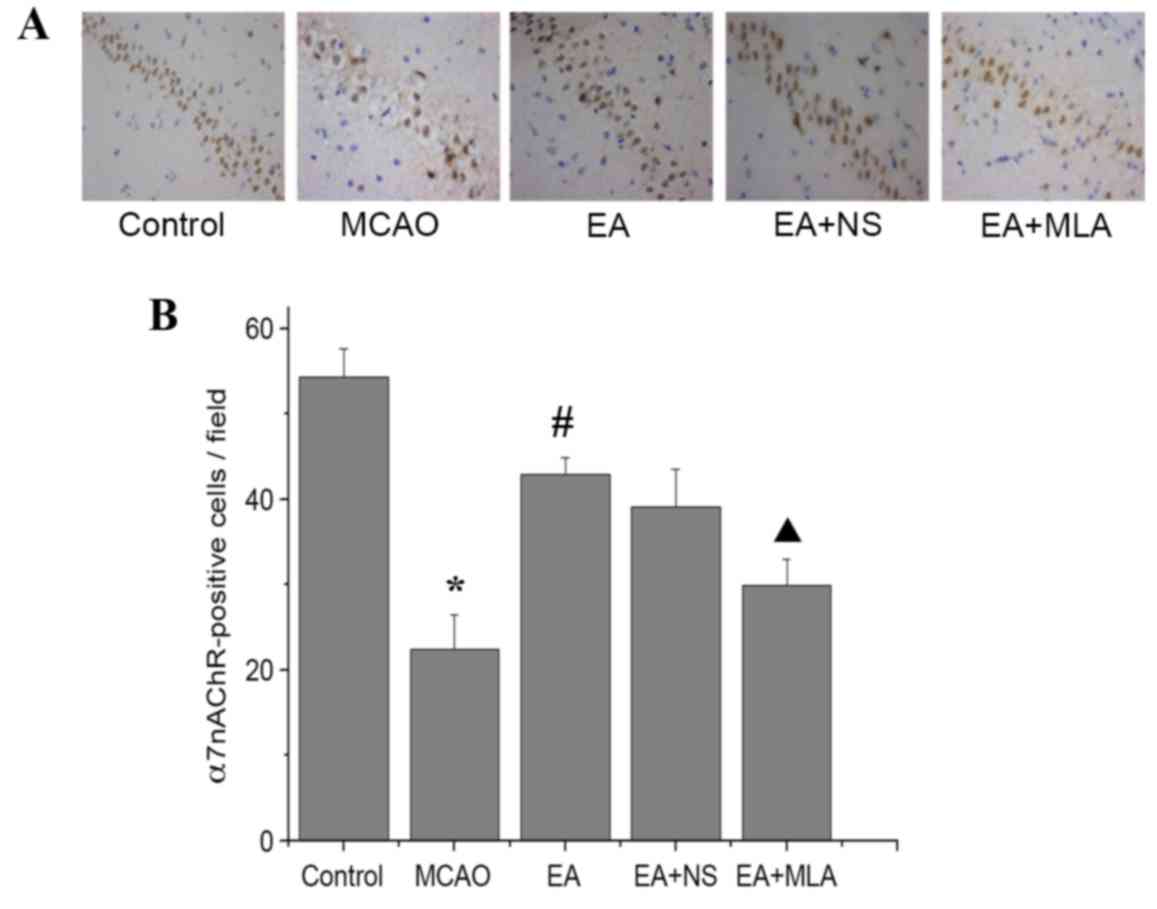
EA activates α7nAChR expression in the
hippocampus
As indicated in Fig.
2, immunohistochemical analysis revealed a significant
reduction in the number of α7nAChR-positive cells in the CA1 region
of the hippocampus in the MCAO group compared with the control
group (P<0.05). However, EA reversed the α7nAChR reduction
caused by I/R injury. Furthermore, MLA decreased α7nAChR expression
levels compared with those in the EA + NS group.
EA suppresses neuroinflammation via
α7-dependent cholinergic pathways in the hippocampus
Cerebral ischemic injury triggers a serious
inflammatory response in the brain. As presented in Fig. 3, elevated expression of the astrocyte
marker GFAP and microglia/macrophage marker Iba1 was detected in
the hippocampus in the MCAO group compared with the control group.
Western blot analysis of α7nAChR was consistent with the
immunohistochemical analysis, and confirmed that the EA-induced
increase in α7nAChR was attenuated by MLA (Fig. 4A and B). Furthermore, elevated
expression of the key inflammatory factors TNF-α and IL-1β
(Fig. 4A and C) was detected in the
hippocampus in the MCAO group compared with the control group. In
the EA treatment group, Iba1, GFAP and inflammatory factors were
significantly reduced compared with those in the MCAO group
(Figs. 3C and 4C; P<0.05). However, those inhibitions
were markedly eradicated in the EA + MLA group compared with the EA
+ NS group.
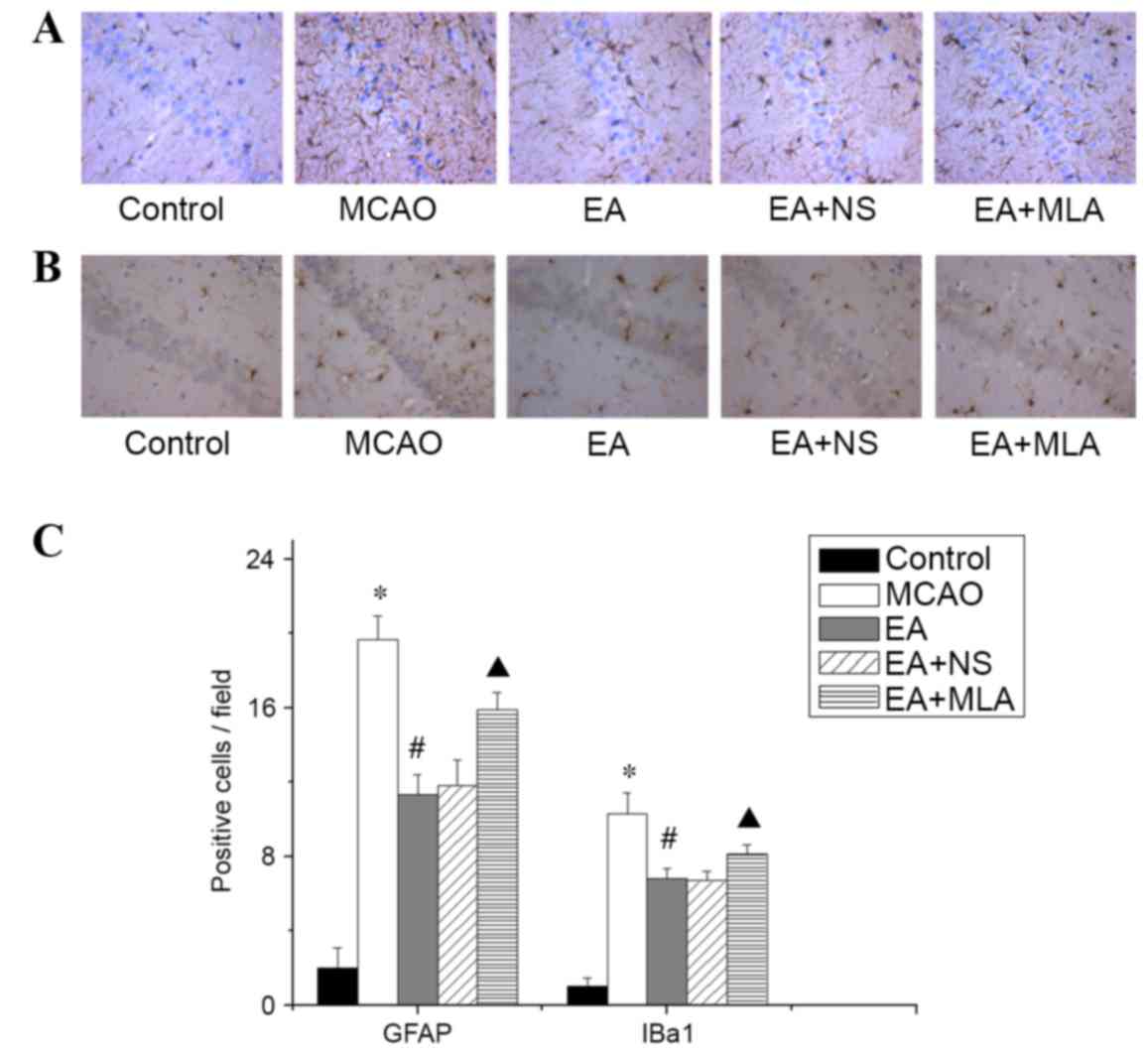 | Figure 3.EA reduces the activity of microglia
and astrocytes in the hippocampus. (A) GFAP-positive astrocytes and
(B) Iba1-positive microglia in the CA1 region of the hippocampus in
the five groups, stained with DAB. (C) Quantified data for the five
groups. *P<0.01 vs. control, #P<0.01 vs. MCAO and
▲P<0.01 vs. EA + NS. Data are expressed as mean ±
standard error of the mean (n=5). Magnification of ×400. EA,
electroacupuncture; GFAP, glial fibrillary acidic protein; Iba1,
microglial marker; MCAO, middle cerebral artery occlusion; NS,
normal saline; MLA, methyllycaconitine; DAB,
3,3′-Diaminobenzidine. |
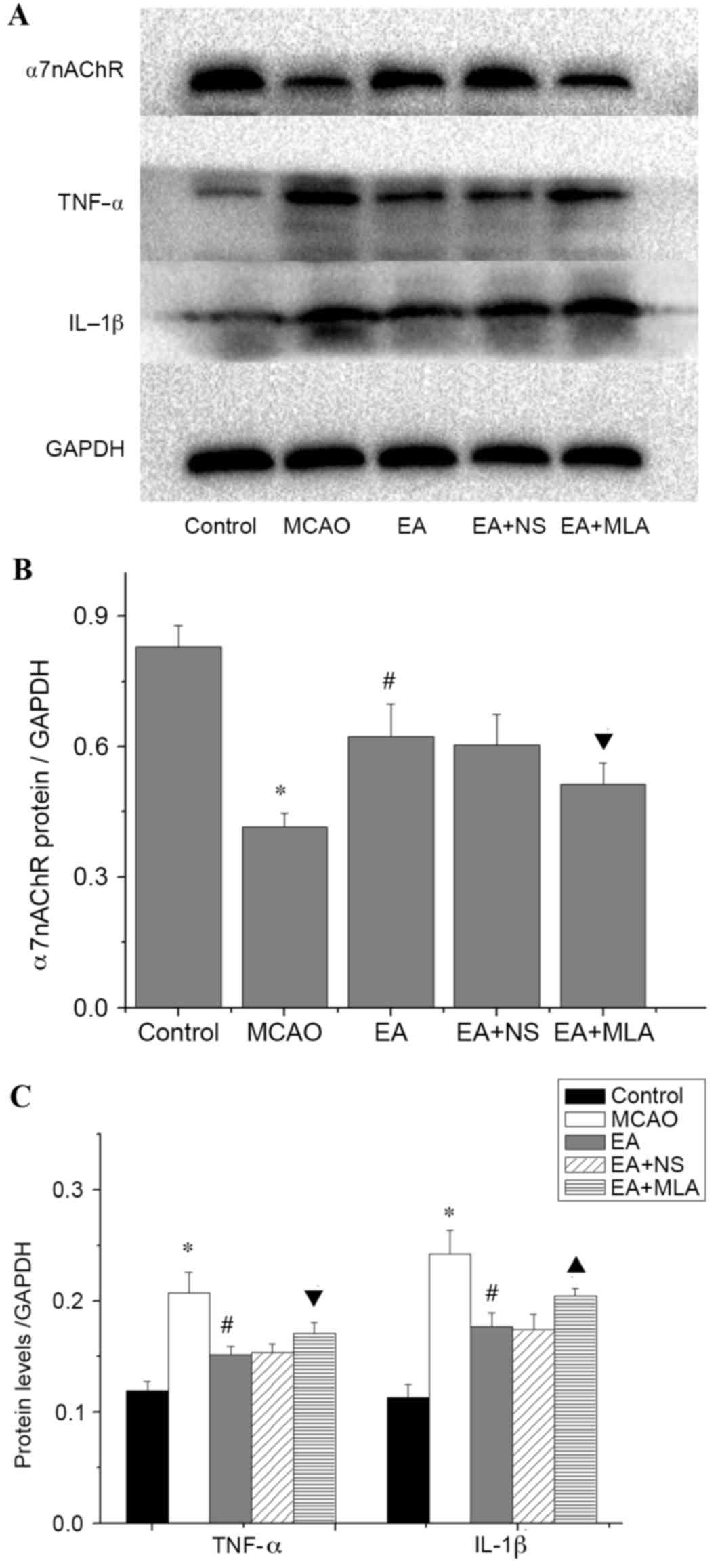 | Figure 4.Western blot analysis of inflammatory
protein expression in the hippocampus. (A) Western blot analysis
demonstrating the concentrations of α7nAChR, TNF-α, IL-1B and
GAPDH. (B) Quantification of the western blots for the levels of
α7nAChR and (C) proinflammatory cytokines TNF-α and IL-1β in the
five groups. *P<0.01 vs. control, #P<0.01 vs.
MCAO, ▲P<0.01 vs. EA + NS and ▼P<0.05 vs. EA + NS.
Data are expressed as mean ± standard error of the mean (n=5).
α7nAChR, α7 nicotinic acetylcholine receptors; TNF-α, tumor
necrosis factor-α; IL-1β, interleukin-1β; GAPDH, glyceraldehyde
3-phosphate dehydrogenase; MCAO, middle cerebral artery occlusion;
EA, electroacupuncture; NS, normal saline; MLA,
methyllycaconitine. |
Discussion
In the present study, EA was demonstrated to reduce
learning and memory deficits via activation of α7nAChR-dependent
anti-inflammatory pathways. In the Morris water maze assessment,
the rats demonstrated a prolonged latency to find the hidden
platform and decreased times of passing the platform position
following MCAO treatment. In rats treated with EA, latency was
shortened and the platform crossing time was increased. These
results suggest that EA at the DU 20 and DU 24 acupoints improved
learning and memory ability in cerebral ischemia-injured rats,
which is consistent with previous studies (33,34).
A large hippocampal neuroinflammatory response was
observed following MCAO, and EA reduced this inflammatory response
as demonstrated by the reduction of Iba1 and GFAP expression and
TNF-α and IL-1β production in the injured hippocampus. The
histological evidence further suggests that α7nAChR mediates the
reduction of neuroinflammation. It is known that α7nAChR is
essential for the cholinergic anti-inflammatory response because
loss of the α7 nicotinic receptor subunit fails to inhibit cytokine
synthesis (35). Studies indicate
that two critical signaling pathways, nuclear factor-κB (NF-κB) and
janus kinase/signal transducer and activator of transcription
(Jak/STAT), are required for the α7nAChR-dependent
anti-inflammatory response. Activation of the α7nAChR prevents Iκ-B
breakdown and promotes p65 nuclear translocation to suppress the
transcription of inflammatory cytokines (36,37).
α7nAChR may recruit the tyrosine kinase Jak2, and activate the
transcription factor STAT3 to inhibit pro-inflammatory gene
transcription (38). Previous
research reports that EA may suppress NF-κB activation to induce an
anti-inflammation response (39) and
enhance STAT3 activation (40).
Global cerebral ischemia diminishes the capacity of
the cholinergic anti-inflammatory pathway to control inflammation,
but the application of an α7nAChR agonist protects against
ischemia-induced cell death and inflammation in the hippocampus.
This significantly decreases the mRNA expression of proinflammatory
cytokines and reduces microglial activation, but not astrocyte
activation (41,42). Conversely, treatment with an α7nAChR
antagonist following global cerebral ischemia worsened neuronal
death and led to increased microglial activation (41,42). In
a mouse model of Parkinson's disease, an α7nAChR agonist decreased
the activation of astrocytes and microglia in the substantia nigra,
and these protective effects were abolished by administration of an
α7nAChR-selective antagonist in vitro and in vivo
(43). The current study indicates
that the neuroprotective effects of EA are due to its actions as a
α7nAChR agonist. Future studies may involve investigating an
α7nAChR agonist in comparison with EA to validate this
hypothesis.
Another potential mechanism for the action of
EA-induced improvements in memory function involves effects on
synaptic plasticity. Neuroinflammation has been confirmed to have a
negative effect on learning and memory processes by blocking
long-term potentiation (LTP) in the hippocampus in vitro and
in vivo (44–46). Synaptic plasticity, neurotransmitter
release and fast synaptic transmission are also modulated by the
activation of neuronal α7nAChR (47,48). In
addition, EA pretreatment has been demonstrated to protect the
brain against transient cerebral ischemic injury via increased
α7nAChR expression on neurons (29).
The current study suggests that EA-mediated regulation of
neuroinflammation may enhance LTP, and combined with EA regulation
of α7nAChR expression on neurons, improve spatial learning and
memory as a result.
In the present study, EA-induced expression of
α7nAChR following temporary MCAO may serve the same role as an
α7nAChR agonist in focal cerebral ischemia. Similarly, co-treatment
with MLA, an α7nAChR antagonist, significantly affected the
inhibitory effects of EA on glial activation and expression of
inflammatory factors. In addition, MLA inhibited the improvement of
spatial learning and memory following EA treatment. From these
data, it may be inferred that EA activates an α7nAChR-dependent
anti-inflammatory pathway to control neuroinflammation, which, in
turn, leads to improvement in learning and memory outcomes after
I/R brain injury.
In conclusion, the current study demonstrates that
EA may reduce learning and memory impairment following cerebral I/R
injury. The protective effects of EA appear to be mediated via the
α7nAChR-mediated anti-inflammatory pathway. EA improves cognitive
function by an α7nAChR-mediated mechanism that decreases
neuroinflammation, as demonstrated by reduced glial activation and
inflammatory cytokine production. However, A potential limitation
of the present study is the lack of a positive control using an
a7nAChR agonist group, which should be considered in future studies
to compare the effects of EA and a7nAChR agonists.
Acknowledgements
This study was supported by the Fujian
Rehabilitation Technology of Collaborative Innovation Center
Support Project (grant no. X2012004), the Natural Science
Foundation of China (grant no. 81403462), the Fujian Natural
Science Foundation of China (grant no. 2015J01335) and the Fujian
Key Laboratory of Rehabilitation Technology.
References
|
1
|
Bae CY and Sun HS: Current understanding
of TRPM7 pharmacology and drug development for stroke. Acta
Pharmacol Sin. 34:10–16. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Niemi ML, Laaksonen R, Kotila M and
Waltimo O: Quality of life 4 years after stroke. Stroke.
19:1101–1107. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Carey LM: Somatosensory loss after stroke.
Crit Rev Phys Rehabil Med. 7:51–91. 1995. View Article : Google Scholar
|
|
4
|
Jin YP, Di Legge S, Ostbye T, Feightner JW
and Hachinski V: The reciprocal risks of stroke and cognitive
impairment in an elderly population. Alzheimers Dement. 2:171–178.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pendlebury ST and Rothwell PM: Risk of
recurrent stroke, other vascular events and dementia after
transient ischaemic attack and stroke. Cerebrovasc Dis. 27 Suppl
3:S1–S11. 2009. View Article : Google Scholar
|
|
6
|
Pohjasvaara T, Erkinjuntti T, Vataja R and
Kaste M: Dementia three months after stroke. Baseline frequency and
effect of different definitions of dementia in the Helsinki Stroke
Aging Memory Study (SAM) cohort. Stroke. 28:785–792. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pendlebury ST and Rothwell PM: Prevalence,
incidence, and factors associated with pre-stroke and post-stroke
dementia: A systematic review and meta-analysis. Lancet Neurol.
8:1006–1018. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chan KY, Wang W, Wu JJ, Liu L, Theodoratou
E, Car J, Middleton L, Russ TC, Deary IJ, Campbell H, et al:
Epidemiology of Alzheimer's disease and other forms of dementia in
China, 1990–2010: A systematic review and analysis. Lancet.
381:2016–2023. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Aggarwal NT, Tripathi M, Dodge HH, Alladi
S and Anstey KJ: Trends in Alzheimer's disease and dementia in the
asian-pacific region. Int J Alzheimers Dis.
2012:1713272012.PubMed/NCBI
|
|
10
|
Wu JN: A short history of acupuncture. J
Altern Complement Med. 2:19–21. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu F, Li ZM, Jiang YJ and Chen LD: A
meta-analysis of acupuncture use in the treatment of cognitive
impairment after stroke. J Altern Complement Med. 20:535–544. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen Y, Zhou J, Li J, Yang SB, Mo LQ, Hu
JH and Yuan WL: Electroacupuncture pretreatment prevents cognitive
impairment induced by limb ischemia-reperfusion via inhibition of
microglial activation and attenuation of oxidative stress in rats.
Brain Res. 1432:36–45. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang C, Wen Y, Fan XN, Tian G, Zhou XY,
Deng SZ and Meng ZH: Therapeutic effects of different durations of
acupuncture on rats with middle cerebral artery occlusion. Neural
Regen Res. 10:159–164. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fan XW, Chen F, Chen Y, Chen GH, Liu HH,
Guan SK, Deng Y, Liu Y, Zhang SJ, Peng WJ, et al:
Electroacupuncture prevents cognitive impairments by regulating the
early changes after brain irradiation in rats. PLoS One.
10:e01220872015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Stoll G, Jander S and Schroeter M:
Inflammation and glial responses in ischemic brain lesions. Prog
Neurobiol. 56:149–171. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang Q, Tang XN and Yenari MA: The
inflammatory response in stroke. J Neuroimmunol. 184:53–68. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Webster SJ, Van Eldik LJ, Watterson DM and
Bachstetter AD: Closed head injury in an age-related Alzheimer
mouse model leads to an altered neuroinflammatory response and
persistent cognitive impairment. J Neurosci. 35:6554–6569. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Johnson VE, Stewart W and Smith DH:
Traumatic brain injury and amyloid-β pathology: A link to
Alzheimer's disease? Nat Rev Neurosci. 11:361–370. 2010.PubMed/NCBI
|
|
19
|
Ramlackhansingh AF, Brooks DJ, Greenwood
RJ, Bose SK, Turkheimer FE, Kinnunen KM, Gentleman S, Heckemann RA,
Gunanayagam K, Gelosa G and Sharp DJ: Inflammation after trauma:
Microglial activation and traumatic brain injury. Ann Neurol.
70:374–383. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Buchanan JB, Sparkman NL, Chen J and
Johnson RW: Cognitive and neuroinflammatory consequences of mild
repeated stress are exacerbated in aged mice.
Psychoneuroendocrinology. 33:755–765. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chin Y, Kishi M, Sekino M, Nakajo F, Abe
Y, Terazono Y, Hiroyuki O, Kato F, Koizumi S, Gachet C and
Hisatsune T: Involvement of glial P2Y1 receptors in cognitive
deficit after focal cerebral stroke in a rodent model. J
Neuroinflammation. 10:952013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tweedie D, Ferguson RA, Fishman K,
Frankola KA, Van Praag H, Holloway HW, Luo W, Li Y, Caracciolo L,
Russo I, et al: Tumor necrosis factor-α synthesis inhibitor
3,6′-dithiothalidomide attenuates markers of inflammation,
Alzheimer pathology and behavioral deficits in animal models of
neuroinflammation and Alzheimer's disease. J Neuroinflammation.
9:1062012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Borovikova LV, Ivanova S, Zhang M, Yang H,
Botchkina GI, Watkins LR, Wang H, Abumrad N, Eaton JW and Tracey
KJ: Vagus nerve stimulation attenuates the systemic inflammatory
response to endotoxin. Nature. 405:458–462. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Boccia MM, Blake MG, Krawczyk MC and
Baratti CM: Hippocampal α7 nicotinic receptors modulate memory
reconsolidation of an inhibitory avoidance task in mice.
Neuroscience. 171:531–543. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Acheson DT, Twamley EW and Young JW:
Reward learning as a potential target for pharmacological
augmentation of cognitive remediation for schizophrenia: A roadmap
for preclinical development. Front Neurosci. 7:1032013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kadir A, Almkvist O, Wall A, Langstrom B
and Nordberg A: PET imaging of cortical 11C-nicotine binding
correlates with the cognitive function of attention in Alzheimer's
disease. Psychopharmacology (Berl). 188:509–520. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee ST, Chu K, Jung KH, Kang KM, Kim JH,
Bahn JJ, Jeon D, Kim M, Lee SK and Roh JK: Cholinergic
anti-inflammatory pathway in intracerebral hemorrhage. Brain Res.
1309:164–171. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu XY, Zhou HF, Pan YL, Liang XB, Niu DB,
Xue B, Li FQ, He QH, Wang XH and Wang XM: Electro-acupuncture
stimulation protects dopaminergic neurons from
inflammation-mediated damage in medial forebrain bundle-transected
rats. Exp Neurol. 189:189–196. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang Q, Wang F, Li X, Yang Q, Li X, Xu N,
Huang Y, Zhang Q, Gou X, Chen S and Xiong L: Electroacupuncture
pretreatment attenuates cerebral ischemic injury through α7
nicotinic acetylcholine receptor-mediated inhibition of
high-mobility group box 1 release in rats. J Neuroinflammation.
9:242012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen LP, Wang FW, Zuo F, Jia JJ and Jiao
WG: Clinical research on comprehensive treatment of senile vascular
dementia. J Tradit Chin Med. 31:178–181. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tyagi E, Agrawal R, Nath C and Shukla R:
Cholinergic protection via alpha7 nicotinic acetylcholine receptors
and PI3K-Akt pathway in LPS-induced neuroinflammation. Neurochem
Int. 56:135–142. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Feng X, Yang S, Liu J, Huang J, Peng J,
Lin J, Tao J and Chen L: Electroacupuncture ameliorates cognitive
impairment through inhibition of NF-κB-mediated neuronal cell
apoptosis in cerebral ischemia-reperfusion injured rats. Mol Med
Rep. 7:1516–1522. 2013.PubMed/NCBI
|
|
34
|
Han X, Zhao X, Lu M, Liu F, Guo F, Zhang J
and Huang X: Electroacupuncture ameliorates learning and memory via
activation of the CREB signaling pathway in the hippocampus to
attenuate apoptosis after cerebral hypoperfusion. Evid Based
Complement Alternat Med. 2013:1564892013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang H, Yu M, Ochani M, Amella CA, Tanovic
M, Susarla S, Li JH, Wang H, Yang H, Ulloa L, et al: Nicotinic
acetylcholine receptor alpha7 subunit is an essential regulator of
inflammation. Nature. 421:384–388. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yoshikawa H, Kurokawa M, Ozaki N, Nara K,
Atou K, Takada E, Kamochi H and Suzuki N: Nicotine inhibits the
production of proinflammatory mediators in human monocytes by
suppression of I-kappaB phosphorylation and nuclear factor-kappaB
transcriptional activity through nicotinic acetylcholine receptor
alpha7. Clin Exp Immunol. 146:116–123. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Saeed RW, Varma S, Peng-Nemeroff T, Sherry
B, Balakhaneh D, Huston J, Tracey KJ, Al-Abed Y and Metz CN:
Cholinergic stimulation blocks endothelial cell activation and
leukocyte recruitment during inflammation. J Exp Med.
201:1113–1123. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
de Jonge WJ, van der Zanden EP, The FO,
Bijlsma MF, van Westerloo DJ, Bennink RJ, Berthoud HR, Uematsu S,
Akira S, van den Wijngaard RM and Boeckxstaens GE: Stimulation of
the vagus nerve attenuates macrophage activation by activating the
Jak2-STAT3 signaling pathway. Nat Immunol. 6:844–851. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu F, Fang J, Shao X, Liang Y, Wu Y and
Jin Y: Electroacupuncture exerts an anti-inflammatory effect in a
rat tissue chamber model of inflammation via suppression of NF-κB
activation. Acupunct Med. 32:340–345. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhou H, Zhang Z, Wei H, Wang F, Guo F, Gao
Z, Marsicano G, Wang Q and Xiong L: Activation of STAT3 is involved
in neuroprotection by electroacupuncture pretreatment via
cannabinoid CB1 receptors in rats. Brain Res. 1529:154–164. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Norman GJ, Morris JS, Karelina K, Weil ZM,
Zhang N, Al-Abed Y, Brothers HM, Wenk GL, Pavlov VA, Tracey KJ and
Devries AC: Cardiopulmonary arrest and resuscitation disrupts
cholinergic anti-inflammatory processes: A role for cholinergic α7
nicotinic receptors. J Neurosci. 31:3446–3452. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Guan YZ, Jin XD, Guan LX, Yan HC, Wang P,
Gong Z, Li SJ, Cao X, Xing YL and Gao TM: Nicotine inhibits
microglial proliferation and is neuroprotective in global ischemia
rats. Mol Neurobiol. 51:1480–1488. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu Y, Hu J, Wu J, Zhu C, Hui Y, Han Y,
Huang Z, Ellsworth K and Fan W: α7 nicotinic acetylcholine
receptor-mediated neuroprotection against dopaminergic neuron loss
in an MPTP mouse model via inhibition of astrocyte activation. J
Neuroinflammation. 9:982012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bellinger FP, Madamba S and Siggins GR:
Interleukin 1 beta inhibits synaptic strength and long-term
potentiation in the rat CA1 hippocampus. Brain Res. 628:227–234.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kelly A, Vereker E, Nolan Y, Brady M,
Barry C, Loscher CE, Mills KH and Lynch MA: Activation of p38 plays
a pivotal role in the inhibitory effect of lipopolysaccharide and
interleukin-1 beta on long term potentiation in rat dentate gyrus.
J Biol Chem. 278:19453–19462. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tong L, Prieto GA, Kramér EA, Smith ED,
Cribbs DH, Lynch G and Cotman CW: Brain-derived neurotrophic
factor-dependent synaptic plasticity is suppressed by
interleukin-1β via p38 mitogen-activated protein kinase. J
Neurosci. 32:17714–17724. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ondrejcak T, Wang Q, Kew JN, Virley DJ,
Upton N, Anwyl R and Rowan MJ: Activation of α7 nicotinic
acetylcholine receptors persistently enhances hippocampal synaptic
transmission and prevents Ass-mediated inhibition of LTP in the rat
hippocampus. Eur J Pharmacol. 677:63–70. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Huang M, Felix AR, Kwon S, Lowe D, Wallace
T, Santarelli L and Meltzer HY: The alpha-7 nicotinic receptor
partial agonist/5-HT3 antagonist RG3487 enhances cortical and
hippocampal dopamine and acetylcholine release. Psychopharmacology
(Berl). 231:2199–2210. 2014. View Article : Google Scholar : PubMed/NCBI
|