Introduction
Spinal cord injury (SCI) is a devastating
complication of thoracic or thoracoabdominal aortic
reconstruction/repair that leads to disabilities, particularly
paraplegia and quadriplegia. It has been estimated that 15–80% of
cases of acute traumatic neuronal damage (primary spinal injury) or
aortic surgery, which are primary spinal injury, may result in
paraplegia or quadriplegia (1,2).
Paraplegia and quadriplegia caused by traumatic primary spinal cord
injury are irreversible and hence, several studies have focused on
treating secondary spinal injury due to ischemia/reperfusion,
hypoxia and glutamate excitotoxicity that are reversible (3,4). The
present study used a rat model of spinal cord ischemia/reperfusion
injury (SCII) in order to elucidate the pathophysiology of
secondary SCII, which has remained to be fully elucidated (5). However, various studies have proposed
that inflammation, apoptosis and oxidative stress are the major
contributors to the development of SCII and associated conditions
(6,7). Hence, a drug that subsides inflammation
and apoptosis along with an improved anti-oxidant capacity would be
the best option for treating SCII conditions.
A large amount of evidence indicated that soy
extracts possess neuroprotective activity against brain
injury/trauma owing to the estrogenic effects of phytoestrogens
(isoflavones), such as daidzein (DZ; 4′,7-dihydroxyisoflavone) and
genistein (8,9). DZ is found in soybeans, soy-based
products (tofu), red clover (Trifolium pratense) as well as
in Chinese herbs such as Kwao Krua (Pueraria mirifica) and
Kudzu (Pueraria lobate) at higher concentrations (10). DZ may be metabolized by intestinal
bacteria to S-equol, which has a broad spectrum of health-promoting
benefits, including antioxidant, anti-inflammatory, anticancer,
anti-obesity as well as cardio- and hepatoprotective properties
(10–12). All these beneficial effects of DZ are
directly linked to its structural similarity to estrogen
(17β-estradiol). Furthermore, DZ/S-equol have been found to act as
agonists of G-protein-coupled estrogen receptor, thus regulating
various signaling pathways (13). In
addition, DZ and genistein also influence the cell survival rate by
desactivating the caspase cascade by positively regulating the
phosphoinositide-3 kinase (PI3K) signaling pathway (14,15).
Methylprednisolone (MP) is a well-known neuroprotective agent that
possesses antioxidant, anti-inflammatory and anti-apoptotic
properties (16), and was hence used
as a standard in the present study.
Apoptosis (programmed cell death) is the major
contributor to neuronal death, which is activated by a
caspase-dependent and -independent pathway (17). In most tissues, cell survival and
proliferation are positively regulated (coordinated) by the
PI3K/Akt signaling pathway via modulating several downstream
molecules, such as mammalian target of rapamycin (mTOR), nuclear
factor erythroid 2-related factor (Nrf2) and glycogen synthase
kinase-3β (GSK-3β) (18,19). Furthermore, apoptosis is one of the
crucial factors for neuronal loss, particularly that associated
with SCII (17). Hence, the present
study was designed to assess whether DZ exerts its neuroprotective
effect by positively upregulating the PI3K/Akt pathway to inhibit
neuronal apoptosis and reduce damage in the rat model of SCII.
Materials and methods
Drugs and reagents
DZ, MP, SDS and Tris buffer were procured from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Isoflurane,
ketamine, physiological saline (0.89%), formaldehyde, DAPI stain,
PBS and xylene were purchased from Beijing Zhongshan Goldenbridge
Biotechnology Co. Ltd. (Beijing, China). All of the other chemicals
were of analytical grade.
Experimental rats
Male Sprague Dawley (SD) rats (n=48; weight, 320–350
g; age, 2 months) were procured from The First People's Hospital of
Yichang Animal House (Hubei, China). Rats were maintained in an
animal facility center in a steel cage, (4 rats in each cage),
under a 12-h light/dark cycle with ad libitum access to food
and water. The study protocol was approved by the Institutional
Review Board of China Three Gorges University (RHTGU.930482), and
was according to the National Institutes of Health Guide for the
Care and Use of Laboratory Animals.
SCI insult
SCI injury was performed according to the method
used by Hwang et al (20)
with a slight modification. In brief, all rats were anesthetized
with ketamine (50 mg/kg) intraperitoneally (i.p.) under fasting
conditions (12 h). Body temperature was stabilized at 37°C by
warming/heating pads and heating lamps. The rats were placed in a
supine position, making sure that neck region was well exposed. The
hair on the neck region (carotid-inguinal) was shaved with
trimmer/shaver. A polyethylene catheter (PE-50) was inserted into
the tail artery to inject heparin as well as to monitor the mean
distal arterial pressure (MDAP). A Fogarty balloon catheter
(Edwards Life Science, Shanghai, China) was inserted via the left
femoral artery into the proximal descending thoracic aorta (~11 cm
from the insertion site), and the balloon was inflated to induce
spinal cord ischemia. During aortic occlusion, the proximal artery
pressure was maintained at 80 mmHg by draining the blood from the
carotid artery into the external blood reservoir filled with
heparinized saline. Aortic occlusion (ischemia) was confirmed by
immediate loss of pulse with decreased MDAP. The balloon was
deflated after 25 min of ischemia to initiate reperfusion by
restoring blood flow. The catheters were then slowly withdrawn, and
the rats were allowed to recover from anesthesia. All of these
procedures, except for aortic occlusion, were performed in
sham-operated control rats, which served as negative controls. Mean
arterial pressure, body temperature and heart beat (vitals) were
continuously monitored throughout the study.
Experimental design
After one week of assimilation, the 48 SD rats were
divided into four treatment groups (n=12 per group): Group I, rats
receiving saline (sham-operated control) without occlusion; Group
II, rats subjected to ischemic insult (SCII); Groups III, and IV;
rats treated with DZ (20 mg/kg) or MP (50 mg/kg) i.p. in saline for
7 days prior to SCII (pre-treatment) and 7 days after SCII
(post-treatment), serving as treatment groups (DZ+SCII;
MP+SCII).
Neurological assessment
Locomotor function (hind limb movement) was
determined on days 1, 3, 5 and 7 after SCII using the Basso,
Beattie and Bresnahan (BBB) motor rating scale. The rating scale
comprises 21 items: 0, no detectable hind limb movement; 1–8,
slight or higher movements of hindlimb joints; 9, dorsal stepping;
10–20, progressive improvement of walking ability; and 21, normal
movement.
Tissue processing
On day 7, after the last neurological assessment,
animals in each group were divided into two sub-groups for
biochemical/bolt analyses (n=6) and terminal deoxynucleotidyl
transferase-mediated deoxyuridine triphosphate nick-end labeling
(TUNEL) assay (n=6). All rats were euthanized on day 7 by i.p.
injection of pentobarbital sodium to remove spinal tissue, which
was immediately fixed with 4% paraformaldehyde for 3 h at 37°C,
followed by dehydration (overnight), embedding in paraffin and
slicing at 5 µm using a microtome for the TUNEL assay (n=6). Spinal
tissue was homogenized in 10% (w/v) Tris-HCl lysis buffer and
centrifuged at 2,500 × g for 20 min at 4°C for use in biochemical
analyses (n=6).
Evaluation of edema (water
content)
Edema was detected using procedures previously
published by Mdzinarishvili et al (21) with certain modifications where the
spinal tissue were super infused with 0.5/min instead of 0.7/min.
The wet and dry weights of spinal samples were measured to
calculate the water content. Spinal edema was defined as the
percentage difference between the two weights.
Anti-oxidant enzymes and
myeloperoxidase (MPO) activity assays
Superoxide dismutase (SOD) and catalase (CAT)
activity was quantified in spinal tissue (homogenate) using
commercial kits (SOD, cat. no. A001; CAT, cat. no. A007; Nanjing
Jiancheng Bioengineering Institute, Nanjing, China) based on the
manufacturer's protocols. One unit (U) of SOD activity is equal to
the amount of enzyme that inhibits the auto-oxidation reaction by
50%. One U of CAT activity in 1 mg of tissue protein was defined as
the amount that consumed 1 µmol of H2O2 at
405 nm during 1 sec. MPO activity in spinal homogenate was measured
using an MPO assay kit (cat. no. A044; Nanjing Jiancheng
Bioengineering Institute) according to the supplier's instructions.
One U of MPO activity was defined as the amount of enzyme degrading
1 mmol peroxidase/min at 25°C and was expressed as U/g of wet
tissue.
Inflammatory and apoptotic
markers
The nuclear factor (NF)-κB free p65 subunit in the
nuclear fraction isolated from the spinal tissue using the
Nuclear/Cytosolic Fractionation Kit (cat. no. AKR171; Cell Biolabs
Inc., San Diego, CA, USA) was evaluated using an ActivELISA kit
(cat. no. IMK503; Imgenex; Novus Biologicals, LLC, Littleton, CO,
USA). The levels of tumor necrosis factor-α (TNF-α) and caspase-3
in spinal tissue (cytosolic fraction) were measured by ELISA using
the Quantikine Rat TNF-α and Quantikine Active Caspase-3 kits
(TNF-α, cat. no. MTA00B; caspase-3, cat. no. KM300/DYC; R&D
Systems Inc., Minneapolis, MN, USA) according to the manufacturer's
protocols.
TUNEL assay
Apoptosis (cell death) in spinal tissue was assessed
using the In Situ BrdUTP-Red DNA fragmentation kit/TUNEL assay kit
from Abcam (cat. no. ab66110; Cambridge, MA, USA), according to
manufacturer's instructions. In brief, spinal tissue was fixed in
4% formaldehyde and washed with PBS for 5 min. The slides were
incubated with protease K (20 µg/ml) and 0.5% Triton X-100 for 20
min at 37°C. Subsequent to washing with PBS, slices were incubated
with 50 µl TUNEL reaction mixture (red) for 30 min at 37°C.
Finally, samples were counterstained with DAPI to visualize all
nuclei (blue) and apoptotic cells (red/pink) by confocal
microscopy. Quantification of positive cells per mm2 was
performed at a magnification of ×400 in a double-blinded manner in
each section of spinal tissue.
Western blot analysis
For analysis of PI3K, total Akt, phosphorylated
(p)-Akt, B-cell lymphoma 2 (Bcl-2) and Bcl-2-associated X protein
(Bax) proteins in the spinal tissue, were extracted using RIPA
lysis buffer (Sigma-Aldrich; Merck KGaA) and the total cellular
protein was estimated using a BCA Protein assay reagent kit
(BioVision, Inc., Milpitas, CA, USA). Subseuently nuclear/cytosolic
proteins were isolated using a Nuclear/Cytosol Extraction kit (cat.
no. AKR171; Cell Biolabs Inc., San Diego, CA, USA) based on the
supplier's instructions. Equal quantities of protein (50 µg/lane)
was separated by 8% SDS-PAGE and then electrotransferred onto a
nitrocellulose membrane by a semi-dry blotting system (GE
Healthcare, Little Chalfont, UK). The membrane was blocked with
Tris-buffered saline (TBS) containing Tween-20 and 5% skimmed milk
and probed with primary antibody at 4°C overnight. The following
antibodies were used in the present study: Rabbit polyclonal
anti-PI3K (cat. no. sc-67306; 1:1,000 dilution), anti-Akt and p-Akt
antibody (cat. nos. sc-5298 and sc-135650; 1:2,000 dilution), mouse
anti-Bcl-2 antibody and anti-Bax antibody (cat. no. sc-509 and
sc-20067; 1:500 dilution) as well as mouse monoclonal anti-rat
β-actin antibody (cat. no. sc-47778; 1:500 dilution) (all from
Santa Cruz Biotechnology, Inc., Dallas, TX, USA), which served as
an internal control (housekeeping protein). Subsequently, samples
were incubated with the secondary antibodies, horseradish
peroxidase-conjugated anti-mouse antibody (cat. no. sc-2380/2371;
1:2,000 and 1:5,000 dilution, respectively; Santa Cruz
Biotechnology, Inc) in TBS at room temperature for 1 h and washed
with TBS. The bound antibodies were visualized using an enhanced
chemiluminescence system (ChemiDoc-17001401; Image Lab-5.2.1;
Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
Values are expressed as the mean ± standard
deviation. SPSS version 23 (International Business Machines, Inc.,
Armonk, NY, USA) was used for statistical analysis. Variations
between groups of experimental rats were evaluated by one-way
analysis of variance and the least significant difference was
determined using a post-hoc multiple comparison test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effect of DZ on locomotor
function
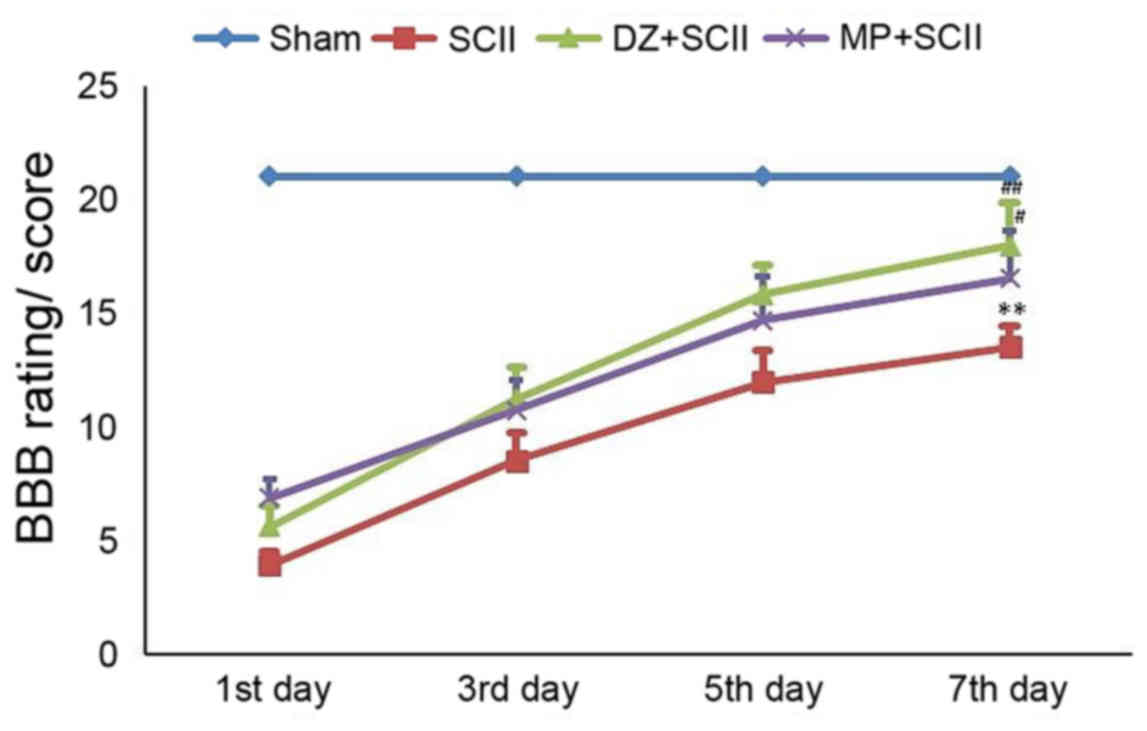
Fig. 1 portrays the
hind limb movement (locomotor function) of sham-operated control
and experimental rats. Sham-operated control rats showed normal
hind limb movement with an average BBB score of 21 without any
neurological deficits. By contrast, SCII rats displayed a
concomitant decline in hind limb movement evidenced by paraplegia
with the lowest BBB score at all time-points (days 1, 3, 5 and 7;
P<0.01). Treatment with DZ and MP greatly ameliorated the
impairment of hind limb movement by effectively alleviating
neuronal damage, which was evidenced by an elevated BBB score
compared with that in SCI-insulted rats (P<0.01 and
<0.05).
Effect of DZ on spinal edema
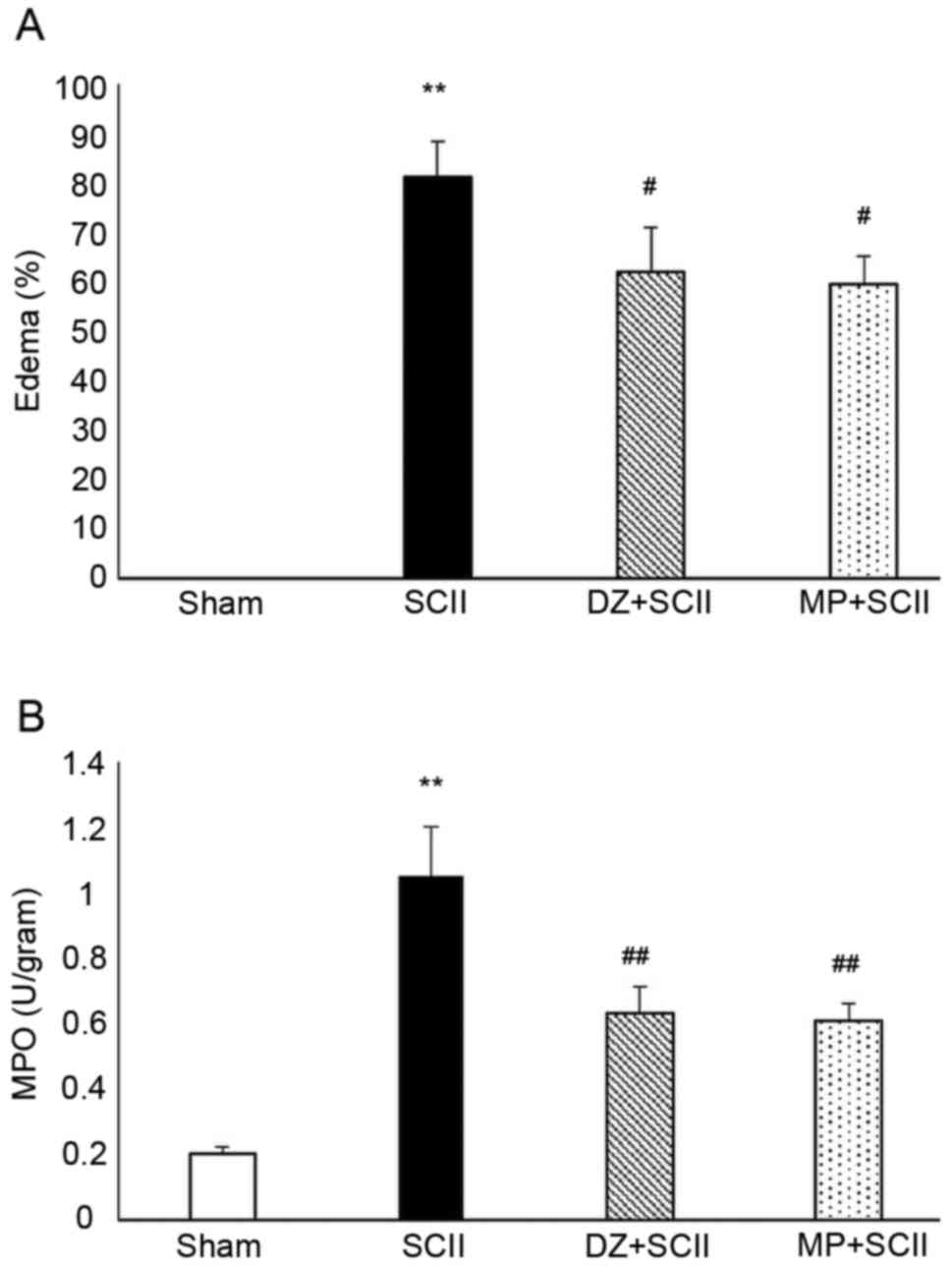
The sham-operated rats did not display any edematous
changes (Fig. 2A). By contrast,
SCI-induced rats had marked edema (81.35%; P<0.01 vs. Sham
group). In comparison with SCI-insulted rats, DZ and MP-treated
rats displayed a significant decline in edema levels at 62.12 and
59.56%, respectively (P<0.05).
Effect of DZ on antioxidant
activity
To determine whether DZ alleviates oxidative stress,
the levels of the endogenous antioxidants SOD and CAT were
determined (Table I). The activities
of SOD and CAT in the spinal cords of SCII rats were markedly
diminished compared with those in the sham-operated control rats
(P<0.05). Treatment with DZ at the dosage of 20 mg/kg
significantly abrogated these decreases (P<0.01), resulting in
near-basal antioxidant levels by abolishing the oxidative stress.
Administration of MP also substantially improved the levels of SOD
and CAT (P<0.05). However, DZ had a better antioxidant activity
when compared with MP.
 | Table I.Effect of DZ on spinal anti-oxidant
activities in experimental rats. |
Table I.
Effect of DZ on spinal anti-oxidant
activities in experimental rats.
| Group | SOD (U/mg
protein) | CAT (U/mg
protein) |
|---|
| Sham (control) |
4.03±0.31 |
73.53±9.51 |
| SCII |
2.89±0.18a |
46.36±5.63a |
| DZ+SCII |
3.86±0.42b |
64.70±6.84b |
| MP+SCII |
3.48±0.51c |
59.85±7.30c |
Effect of DZ on MPO activity
MPO activity was determined to assess the efficacy
of DZ on neutrophil activation and infiltration (Fig. 2B). Compared with that in the
sham-operated control group, the SCII group displayed increased MPO
activity (P<0.01). In comparison, treatment with DZ and MP for 7
days prior to and following SCII significantly restrained MPO
activity by inactivating neutrophils (P<0.01).
Effect of DZ on inflammatory
markers
The anti-inflammatory effect of DZ was evaluated by
assessing various inflammatory cytokines, including TNF-α in the
spinal tissues of the rats and NF-κB subunit p65 in the nuclear
fraction of these tissues. The levels of TNF-α and nuclear NF-κB
p65 in SCI-insulted rats were significantly increased compared with
those in sham-operated rats (P<0.01; Table II). However, DZ and MP substantially
suppressed the levels of TNF-α and nuclear translocation of the
NF-κB p65 subunit, thus indicating its anti-inflammatory
activity.
| Table II.Effect of DZ on spinal inflammatory
and apoptotic markers in experimental rats. |
Table II.
Effect of DZ on spinal inflammatory
and apoptotic markers in experimental rats.
| Group | TNF-α (ng/mg
ptn) | NF-κB p65 (pg/mg
ptn) | Caspase-3 (ng/mg
ptn) |
|---|
| Sham (control) |
118.25±17.23 |
82.45±7.25 |
9.13±0.51 |
| SCII |
281.46±21.68a |
189.57±17.45a |
35.28±3.52a |
| DZ+SCII |
137.67±15.11b |
91.36±7.24b |
15.63±1.52b |
| MP+SCI |
132.95±12.67b |
99.67±11.46b |
17.57±1.26b |
Effect of DZ on apoptotic markers
Caspase-3 is a crucial apoptotic protease involved
in apoptosis and hence, caspase-3 activity was measured to
investigate the impact of DZ on apoptotic events by ELISA.
Caspase-3 activity in SCII rats was greatly increased compared with
that in sham-operated rats (P<0.01; Table II). Compared with that in the SCII
group, DZ and MP treatment concomitantly attenuated caspase-3
activity (P<0.01).
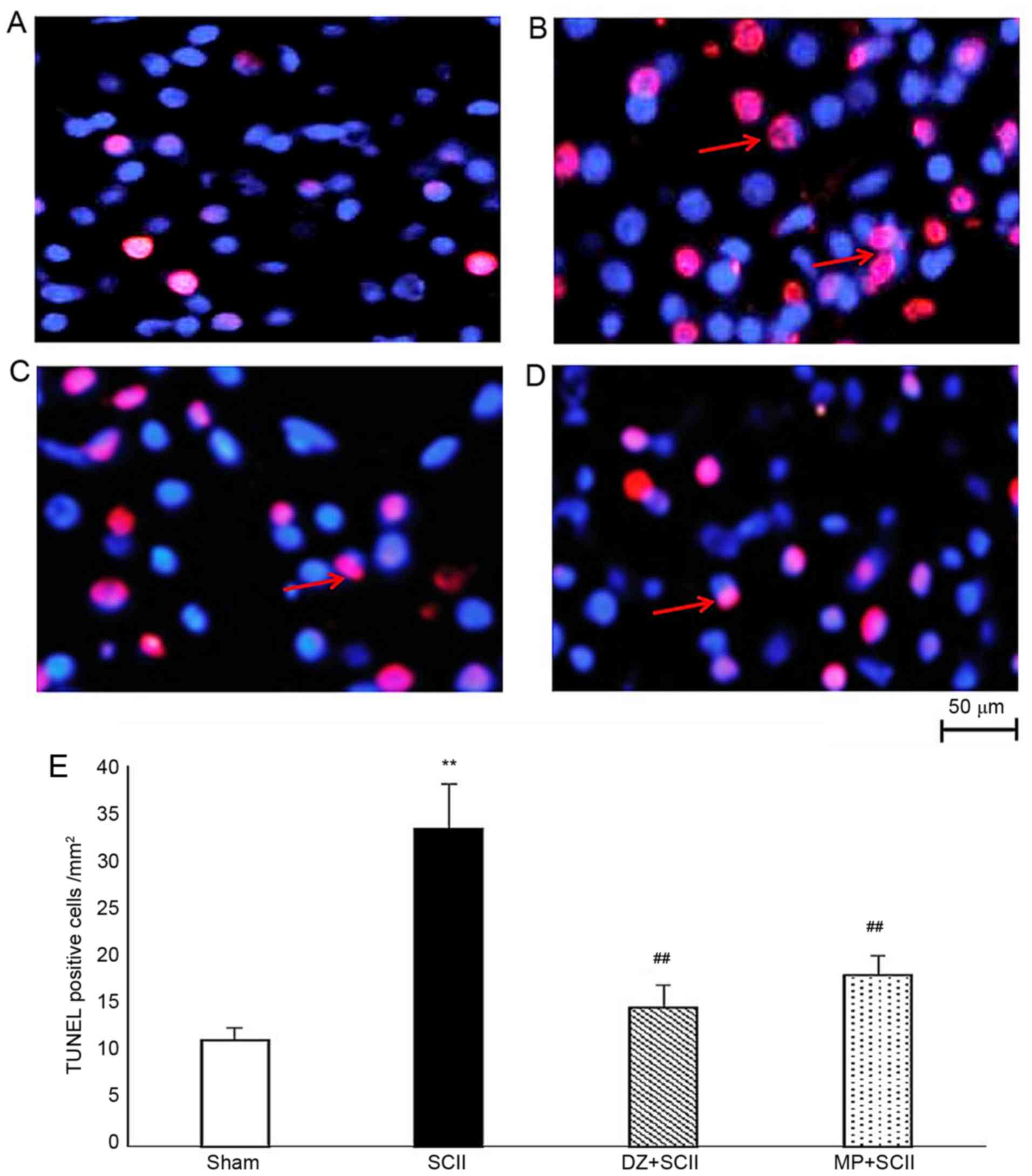
Effect of DZ on apoptotic cells by
TUNEL assay
TUNEL staining was performed to visualize apoptotic
cells in spinal tissues of sham-operated control and experimental
rats (Fig. 3). Histological sections
of spines from SCII rats displayed an increased the number of
TUNEL-positive cells in comparison with those of sham-operated
control rats (P<0.01). However, the DZ and MP groups displayed a
concomitant decline in the number of TUNEL-positive cells, compared
with that in SCII rats (P<0.01).
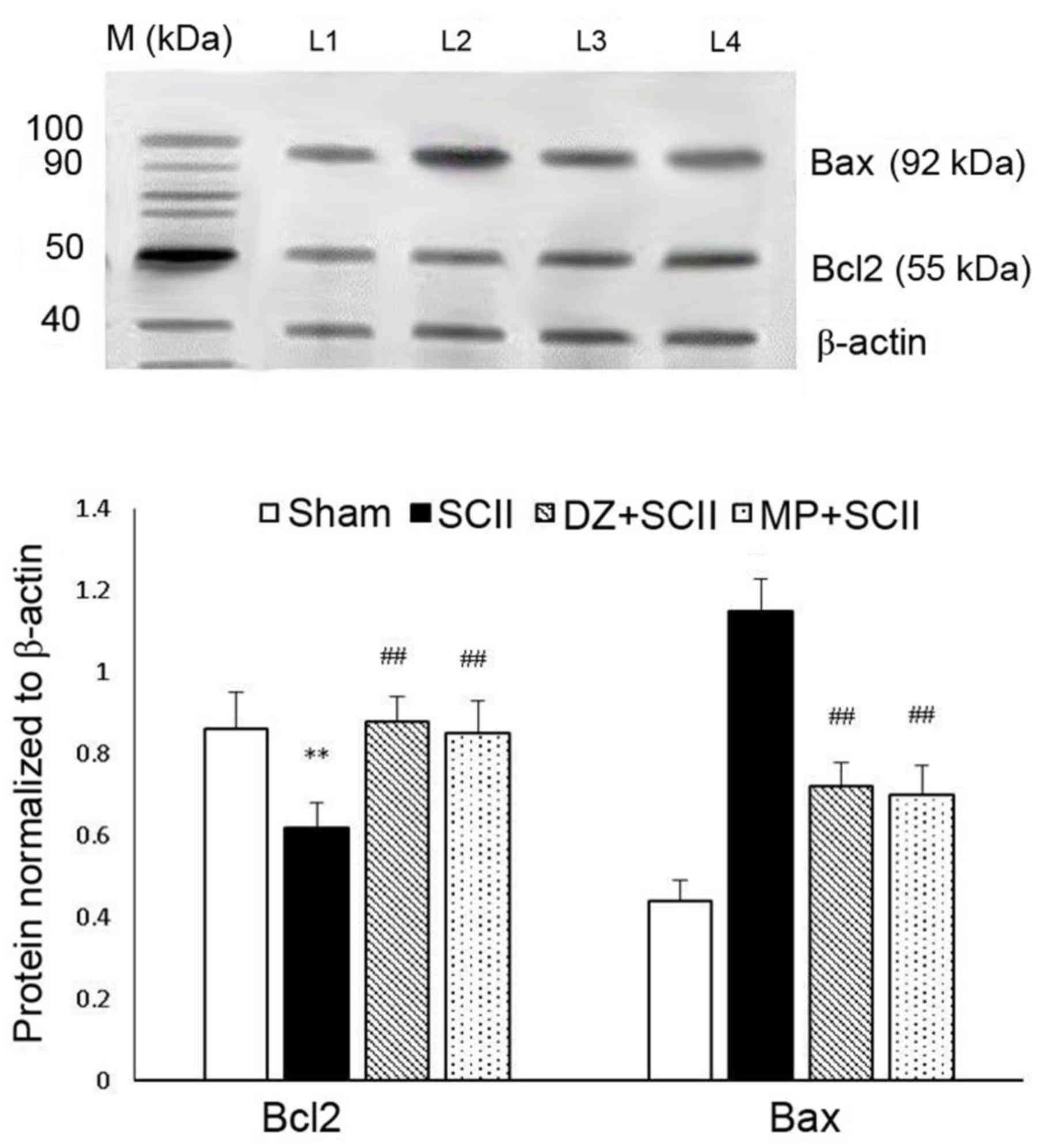
Effect of DZ on the expression of
PI3K/Akt and apoptotic signaling proteins
Western blot analysis was performed to examine the
protein expression of pro-apoptotic protein Bax and anti-apoptotic
protein Bcl-2 levels to determine the anti-apoptotic activity of DZ
(Fig. 4). In addition, the protein
levels of PI3K, Akt and pAkt were also determined to investigate
whether this signaling pathway is associated with the
neuroprotective properties of DZ (Fig.
5). The protein expression of PI3K, Bcl-2 and the pAkt/Akt
ratio in the SCII group displayed a significant downregulation as
compared with that in the sham-operated group (P<0.01), while
the protein expression of Bax was markedly upregulated in SCII rats
(P<0.01). However, compared with those in the SCII group, the
protein levels of PI3K, the pAkt/Akt ratio and Bcl-2 levels in the
DZ and MP groups were significantly increased, while Bax levels
were significantly downregulated (P<0.01 or <0.05).
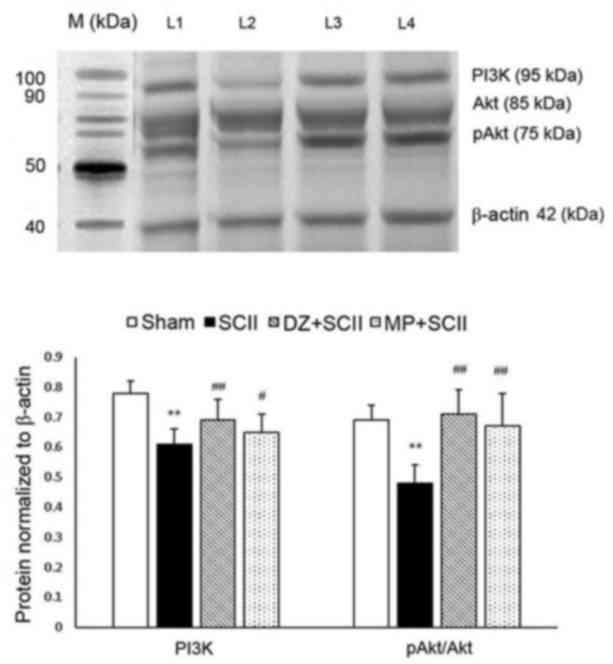 | Figure 5.Effect of DZ on protein expression of
PI3K and pAkt/Akt in spinal tissue homogenate of experimental rats.
Representative western blot images are displayed and quantitative
values are expressed as the mean ± standard deviation for twelve
rats in each group. β-actin was used as internal standard. Lanes:
1, sham control group; 2, SCII group; 3, DZ+SCII group; 4, MP+SCII
group. **P<0.01 vs. sham control group; ##P<0.01,
#P<0.05 vs. SCII group. DZ, daidzein; MP,
methylprednisolone; SCII, spinal cord ischemia/reperfusion injury;
pAkt, phosphorylated Akt; PI3K, phosphoinositide-3 kinase. |
Discussion
The present study was performed to assess the
detailed molecular mechanisms of the neuroprotective effects of DZ
in a rat model of SCII. The PI3K/Akt signaling pathway is crucial
for the maintenance of cell survival (anti-apoptotic effect) and
proliferation in most tissues, particularly neurons and glial cells
(17,19). Hence, the present study postulated
that DZ probably exerts its neuroprotective effects by positively
upregulating various downstream molecules of the PI3K/Akt signal
transduction pathway. To prove this hypothesis, hind limb locomotor
function of experimental rats was evaluated by BBB scoring, and
oxidative stress (anti-oxidants), inflammatory markers (MPO, TNF-α
and NF-p65), apoptotic markers (caspase-3, Bcl-2 and Bax) as well
as the protein expression of signaling molecules such as PI3K and
pAkt were assessed.
The neuroprotective efficacy of pharmaceutical drugs
pertaining to the motor movement was screened through the BBB
scoring system. It is one of the standard methods to measure the
hind limb movement prior to and after injury (7,20). The
results of the present study indicated that SCII group rats showed
a decreased BBB score due to the rapid onset of neurological damage
owing to lack of sufficient blood circulation (hypoxia) under
ischemic conditions. However, supplementation with DZ and MP
significantly improved hind limb movement, which was reflected in a
substantial increase in BBB score. Since DZ and MP act as a potent
antioxidant and anti-inflammatory agents, they suppress oxidative
stress and the subsequent inflammatory response to thereby lower
neuronal damage or deficits. DZ has been reported to lower
neurological deficits by improving the neuronal count and thus
significantly ameliorating hind limb movement (10).
Spinal morphological abnormalities were circuitously
analyzed via quantifying edema based on water content (dry and wet
weight). SCI-induced rats displayed severe edematous conditions
compared with sham-operated rats, likely due to
ischemic/reperfusion causing electrolytic imbalance and resulting
in a high-water content (edema). DZ- and MP-treated rats
demonstrated a decline in edema levels, probably through alteration
of the electrolytic balance through maintenance of the blood-spinal
cord barrier (BSCB). The results are in agreement with those of Ma
et al (11), who reported
that DZ treatment reduced edema in an ischemic rat model.
It is well documented that under ischemic
conditions, free radicals are generated excessively as a cellular
response to ischemic reperfusion, leading to oxidative stress
(6). In the present study, SOD and
CAT activity in SCII animals were significantly decreased owing to
overproduction of free radicals (oxidative stress). The
SCII-induced decreases in antioxidant activity (SOD and CAT) were
reverted to near normal levels by administration of DZ and MP,
likely via their antioxidant effects. Atkinson et al
(22) demonstrated that DZ exhibits
excellent antioxidant activity due to estrogenic activity. In
addition, Xu et al (14)
suggested that the isoflavone DZ may attenuate oxidative stress in
a diabetic cell model owing to its two free hydroxyl groups. Zhang
et al (23) proved that DZ
activated the Nrf2/heme oxygenase-1 signaling pathway through the
PI3K/Akt pathway. Hence, the present study hypothesized that DZ
causes upregulation of various endogenous antioxidants,
particularly SOD and CAT, via PI3K/Akt.
Oxidative stress and inflammation have been reported
to be interlinked under spinal or cerebral ischemic/reperfusion
conditions (20). Under SCII
conditions, neutrophils and neuroglia are highly activated and
mobilized (infiltration) due to increased oxidative stress and
disruption of BSCB. Those activated cells may initiate the
inflammatory cascade by triggering NF-κB and subsequently
increasing various pro-inflammatory cytokines such as TNF-α
(24). Hence, the present study
initially measured the activity of MPO, which is the best marker
for neutrophil infiltration, as well as various inflammatory
markers, to determine the inflammatory status. The levels of MPO,
TNF-α and NF-κB p65 were elevated in SCII rats. However, DZ and MP
substantially suppressed the levels of MPO, TNF-α and nuclear
translocation of the NF-κB p65 subunit, thus indicating its
anti-inflammatory activity by inactivating neutrophils and
neuroglial cells. These results are congruent with those of Kim
et al (25), who demonstrated
that DZ administration reduced ischemia/reperfusion-induced
myocardial damage in vivo via inhibition of NF-κB activation
(nuclear translocation of NF-κB p65).
Apoptosis is a programmed cell death, which is an
integral part of various biological events. Apoptosis is
upregulated in various neurodegenerative diseases, including
Alzheimer's, Parkinson's and Huntington's disease as well as in
traumatic injuries, particularly neurological injuries such as SCI
(26,27). The proliferative and apoptotic rate
in each tissue type are maintained in a balance under normal
conditions. Any alteration in either the proliferative or apoptotic
rate may result in pathological conditions such as cancer,
auto-immune diseases or degenerative diseases. Caspases
(cysteine-aspartic proteases) are a family of proteases that has a
critical role in executing apoptosis. Apoptotic caspases are
classified into two types as the initiator (caspase-2, −8 and −9)
and executor (caspase-3, −6 and −7) caspases. The initiator
caspases activate executor caspases to trigger apoptosis (28). Among them, caspase-3 is considered as
the prime executioner protease of the apoptotic cascade through
breaking DNA as well as disassembling its repair system (29). Therefore, the determination of
caspase-3 levels as an apoptotic marker is a crucial criterion to
assess apoptosis, particularly during post-ischemic conditions
(6). In the present study, caspase-3
activity was significantly increased in SCI-insulted rats due to
elevated oxidative stress and inflammatory response, which may
hasten neuronal apoptosis under ischemic conditions. Compared with
that in the SCII group, caspase-3 activity was concomitantly
attenuated in the DZ- and MP-treated groups, probably as a result
of the decreased oxidative stress and the inflammatory cascade.
Rivera et al (9) indicated
that supplementation with daidzein substantially reduced caspase-3
activity and thereby lowered apoptosis. Similarly, Aras et
al (10) demonstrated that
supplementation with daidzein substantially decreased the number of
cells with positive caspase-3 staining by using
immunohistochemistry.
Apoptosis is highly regulated by pro-apoptotic
factors (Bax and Bcl-2-associated death promoter) and
anti-apoptotic factors (Bcl-2 and Bcl extra large protein). Bax and
Bcl-2 have an antagonising interaction (30), and in the present study, the
expression levels of these two proteins we evaluated to determine
apoptosis. The anti-apoptotic factor Bcl-2 usually resides in the
mitochondrial outer membrane and in the nuclear membrane to prevent
the release of cytochrome C from mitochondria into the
cytosol. Thus, Bcl-2 effectively inhibits caspase activation and
thereby inactivates the triggering of the apoptotic caspase
cascade. Furthermore, Bcl-2 inhibits the activation of
pro-apoptotic factor Bax and thus potentially halts apoptotic
events (31,32). In the present study, the protein
expression of Bcl-2 was markedly downregulated, whereas Bax was
notably upregulated in the SCII group compared with that in the
sham-operated group. Upon treatment with DZ and MP, the protein
expression of Bcl-2 was markedly upregulated and Bax was
substantially downregulated. Therefore, it was confirmed that DZ
had effective anti-apoptotic activity by upregulating Bcl-2 and
downregulating Bax to inactivate the apoptotic cascade. Mao et
al (33) indicated that DZ acts
as an anti-apoptotic agent by significantly downregulating the mRNA
expression of the pro-apoptotic protein Bax and upregulating the
anti-apoptotic protein Bcl-2 in to inhibit D-galactose-induced
neurotoxicity. In addition, DZ is an estrogen mimic with the
ability to enhance cell proliferation and prevent neuronal loss
(34). In line with this, the
present study proved that DZ effectively attenuates caspase-3
activity and hence inhibits apoptosis.
A TUNEL assay was performed to visualize the
apoptotic cells, which were activated by pro-apoptotic proteins and
the caspase cascade. The stained spinal sections of SCII rats had a
significantly increased number of TUNEL-positive cells compared
with those in the sham group. However, sections of DZ- or
MP-treated rats had a significantly decreased number of
TUNEL-positive cells compared with those in SCII rats. In a
previous study, the DZ metabolite equol exerted significant
anti-apoptotic activity as reflected by a decreased number of
TUNEL-positive cells in a H2O2-induced
endothelial cell cytotoxicity model (23). The present study proved that DZ
substantially decreased apoptosis by altering the expression of
pro- and anti-apoptotic proteins and thereby attenuating the
concentration of caspase-3.
As mentioned previously, PI3K/Akt pathway has a
pivotal role in neuronal regeneration after acute SCII (19). In addition, numerous studies have
indicated that the neuroprotective activity of soy beans is mainly
bestowed by DZ, geinstein and equol (metabolite of DZ) probably due
to their estrogenic activity via the PI3K/Akt pathway (11,35).
Hence, the present study assessed whether DZ exerts its
neuroprotective activity by upregulating PI3K/Akt signaling
molecules to lower apoptosis and enhance regeneration of neurons.
The protein expression and the pAkt/Akt ratio were significantly
decreased in ischemic/reperfusion-induced animals, which was
inhibited by intervention with DZ. Since DZ is an estrogen mimic,
it activates PI3K/Akt-dependent downstream molecules, which may be
a key mechanism underlying its neuroprotective properties (35,36).
Zhang et al (23)
demonstrated that S-equol exerts its neuroprotective function via
activation of Nrf2 through Akt phosphorylation. A previous study
also demonstrated that DZ downregulated GSK-3β and upregulated mTOR
and thereby inhibited apoptosis through the activation of Bcl-2 in
an ischemic model (37). Fig. 6 presents a schematic of the in-depth
molecular mechanism of the neuroprotective effect of DZ identified
in the present study.
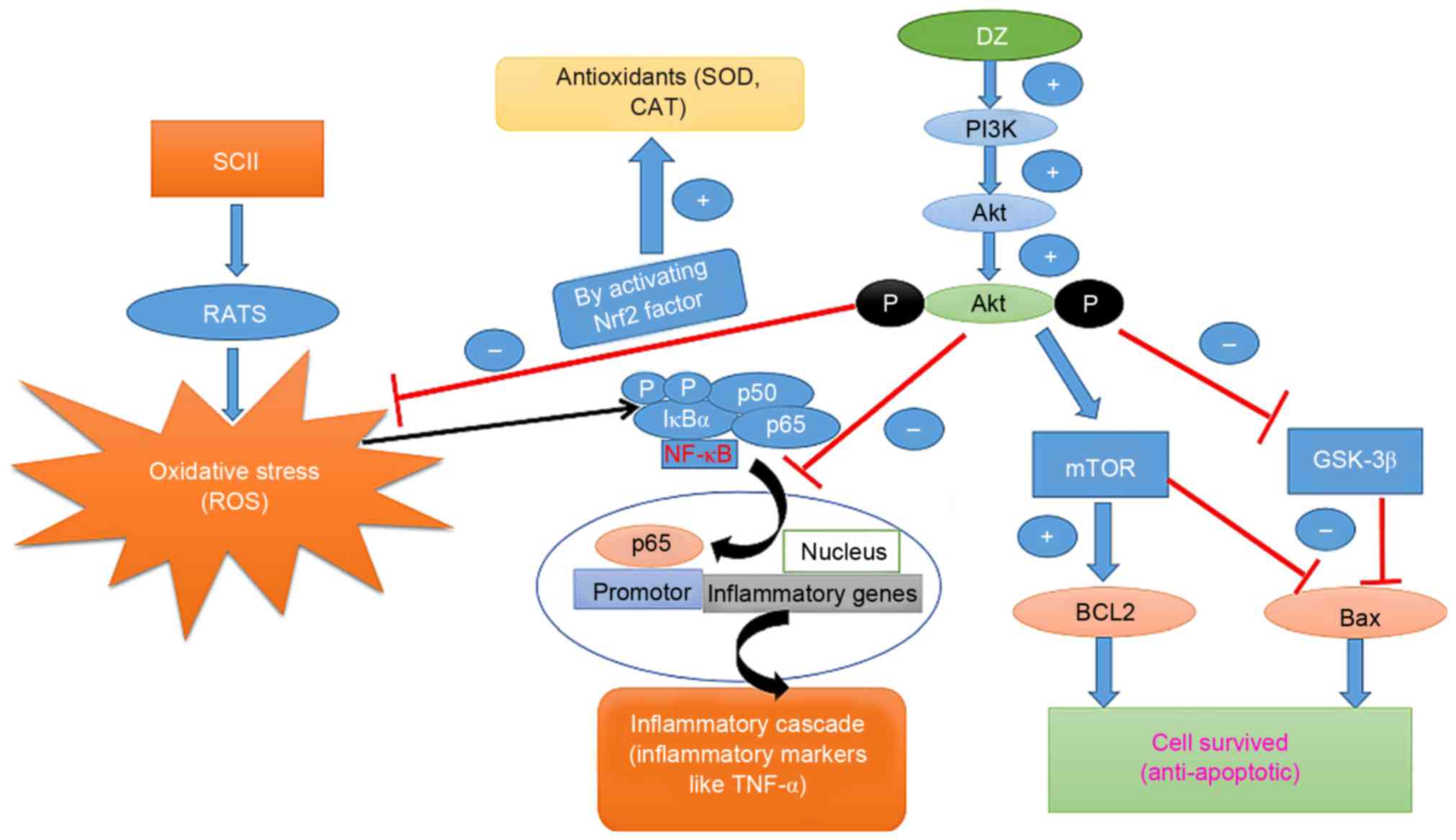 | Figure 6.Schematic representing the in-depth
molecular mechanism of the neuroprotective effect of DZ. DZ,
daidzein; SCII, spinal cord ischemia/reperfusion injury; SOD,
superoxide dismutase; CAT, catalase; PI3K, phosphoinositide-3
kinase; P, phosphate; NF-κB, nuclear factor κB; IκBα, inhibitor of
NF-κB α; TNF, tumor necrosis factor; Bcl-2, B-cell lymphoma 2; Bax,
Bcl-2-associated X protein; mTOR, mammalian target of rapamycin;
GSK, glycogen synthase kinase; Nrf2, nuclear factor erythroid
2-related factor. |
Of note, the present study had certain limitations.
For instance, the BBB staining assay to assess the integrity of
neurons (tight junctions), and detection of markers for
mitochondrial dysfunction (membrane potential) was not performed.
Furthermore, the levels of GSK-3β and mTOR were not measured to
confirm the anti-apoptotic activity of DZ.
In conclusion, the present study suggested that pre-
and post-SCII treatment with DZ for seven consecutive days each
considerably inhibited impairments of hind limb movement (motor
function) via activation of PI3K and Akt, likely through inhibiting
GSK-3β as well as activation (phosphorylation) of mTOR. Activated
mTOR activates anti-apoptotic Bcl-2 protein, which in turn inhibits
pro-apoptotic Bax protein as well as inactivates caspase-3 enzyme
to promote cell survival/proliferation and thereby maintain/restore
neuronal integrity and motor function. Thus, DZ restrained
apoptosis through the PI3K/Akt signaling pathway to exert its
neuroprotective activity. Further study is required to elucidate
the complete mechanism of the neuroprotective effects of DZ.
References
|
1
|
Wong DR, Coselli JS, Amerman K, Bozinovski
J, Carter SA, Vaughn WK and LeMaire SA: Delayed spinal cord
deficits after thoracoabdominal aortic aneurysm repair. Ann Thorac
Surg. 83:1345–1355. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kanellopoulos GK, Xu XM, Hsu CY, Lu X,
Sundt TM and Kouchoukos NT: White matter injury in spinal cord
ischemia protection by AMPA/kainate glutamate receptor antagonism.
Stroke. 31:1945–1952. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang P, Zhang L, Zhu L, Chen F, Zhou S,
Tian T, Zhang Y, Jiang X, Li X, Zhang C and Xu L: The change
tendency of PI3K/Akt pathway after spinal cord injury. Am J Transl
Res. 7:2223–2232. 2015.PubMed/NCBI
|
|
4
|
Maniar HS, Sundt TM III, Prasad SM, Chu
CM, Camillo CJ, Moon MR, Rubin BG and Sicard GA: Delayed paraplegia
after thoracic and thoracoabdominal aneurysm repair: A continuing
risk. Ann Thorac Surg. 75:113–119. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang Z, Zhang C, Hong Z, Chen H, Chen W
and Chen G: C/EBP homologous protein (CHOP) mediates neuronal
apoptosis in rats with spinal cord injury. Exp Ther Med. 5:107–111.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gokce EC, Kahveci R, Gokce A, Sargon MF,
Kisa U, Aksoy N, Cemil B and Erdogan B: Curcumin attenuates
inflammation, oxidative stress, and ultrastructural damage induced
by spinal cord ischemia-reperfusion injury in rats. J Stroke
Cerebrovasc Dis. 25:1196–1207. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lafci G, Gedik HS, Korkmaz K, Erdem H,
Cicek OF, Nacar OA, Yildirim L, Kaya E and Ankarali H: Efficacy of
iloprost and montelukast combination on spinal cord
ischemia/reperfusion injury in a rat model. J Cardiothorac Surg.
8:642013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Burguete MC, Torregrosa G, Pérez-Asensio
FJ, Castelló-Ruiz M, Salom JB, Gil JV and Alborch E: Dietary
phytoestrogens improve stroke outcome after transient focal
cerebral ischemia in rats. Eur J Neurosci. 23:703–710. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rivera P, Pérez-Martín M, Pavón FJ,
Serrano A, Crespillo A, Cifuentes M, López-Ávalos MD, Grondona JM,
Vida M and Fernández-Llebrez P: Pharmacological administration of
the isoflavone daidzein enhances cell proliferation and reduces
high fat diet-induced apoptosis and gliosis in the rat hippocampus.
PLoS One. 8:e647502013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Aras AB, Guven M, Akman T, Ozkan A, Sen
HM, Duz U, Kalkan Y, Silan C and Cosar M: Neuroprotective effects
of daidzein on focal cerebral ischemia injury in rats. Neural
Regener Res. 10:146–152. 2015. View Article : Google Scholar
|
|
11
|
Ma Y, Sullivan JC and Schreihofer DA:
Dietary genistein and equol (4′,7 isoflavandiol) reduce oxidative
stress and protect rats against focal cerebral ischemia. Am J
Physiol Regul Integr Comp Physiol. 299:R871–R877. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu J, Oka J, Ezaki J, Ohtomo T, Ueno T,
Uchiyama S, Toda T, Uehara M and Ishimi Y: Possible role of equol
status in the effects of isoflavone on bone and fat mass in
postmenopausal Japanese women: A double-blind, randomized,
controlled trial. Menopause. 14:866–874. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Prossnitz ER and Barton M: Estrogen
biology: New insights into GPER function and clinical
opportunities. Mol Cell Endocrinol. 389:71–83. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu SZ, Zhong W, Ghavideldarestani M,
Saurabh R, Lindow SW and Atkin SL: Multiple mechanisms of soy
isoflavones against oxidative stress-induced endothelium injury.
Free Radic Biol Med. 47:167–175. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lovekamp-Swan T, Glendenning M and
Schreihofer DA: A high soy diet reduces programmed cell death and
enhances bcl-xl expression in experimental stroke. Neuroscience.
148:644–652. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pettiford JN, Bikhchandani J, Ostlie DJ,
St Peter SD, Sharp RJ and Juang D: A review: The role of high dose
methylprednisolone in spinal cord trauma in children. Pediatr Surg
Int. 28:287–294. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang J, Deng Z, Liao J, Song C, Liang C,
Xue H, Wang L, Zhang K and Yan G: Leptin attenuates cerebral
ischemia injury through the promotion of energy metabolism via the
PI3K/Akt pathway. J Cereb Blood Flow Metab. 33:567–574. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang P, Zhang L, Zhu L, Chen F, Zhou S,
Tian T, Zhang Y, Jiang X, Li X and Zhang C: The change tendency of
PI3K/Akt pathway after spinal cord injury. Am J Transl Res.
7:2223–2232. 2015.PubMed/NCBI
|
|
19
|
Jung SY, Kim DY, Yune TY, Shin DH, Baek SB
and Kim CJ: Treadmill exercise reduces spinal cord injury-induced
apoptosis by activating the PI3K/Akt pathway in rats. Exp Ther Med.
7:587–593. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hwang JY, Min SW, Jeon YT, Hwang JW, Park
SH, Kim JH and Han SH: Effect of coenzyme Q10 on spinal cord
ischemia-reperfusion injury. J Neurosurg Spine. 22:432–438. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mdzinarishvili A, Kiewert C, Kumar V,
Hillert M and Klein J: Bilobalide prevents ischemia-induced edema
formation in vitro and in vivo. Neuroscience. 144:217–222. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Atkinson C, Frankenfeld CL and Lampe JW:
Gut bacterial metabolism of the soy isoflavone daidzein: Exploring
the relevance to human health. Exp Biol Med. 230:155–170. 2005.
|
|
23
|
Zhang T, Liang X, Shi L, Wang L, Chen J,
Kang C, Zhu J and Mi M: Estrogen receptor and PI3K/Akt signaling
pathway involvement in S-(−)equol-induced activation of Nrf2/ARE in
endothelial cells. PLoS One. 8:e790752013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Palencia G, Medrano JÁ, Ortiz-Plata A,
Farfán DJ, Sotelo J, Sánchez A and Trejo-Solís C: Anti-apoptotic,
anti-oxidant and anti-inflammatory effects of thalidomide on
cerebral ischemia/reperfusion injury in rats. J Neurol Sci.
351:78–87. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim JW, Jin YC, Kim YM, Rhie S, Kim HJ,
Seo HG, Lee JH, Ha YL and Chang KC: Daidzein administration in vivo
reduces myocardial injury in a rat ischemia/reperfusion model by
inhibiting NF-kappaB activation. Life Sci. 84:227–234. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hwang L, Choi IY, Kim SE, Ko IG, Shin MS,
Kim CJ, Kim SH, Jin JJ, Chung JY and Yi JW: Dexmedetomidine
ameliorates intracerebral hemorrhage-induced memory impairment by
inhibiting apoptosis and enhancing brain-derived neurotrophic
factor expression in the rat hippocampus. Int J Mol Med.
31:1047–1056. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sung YH, Kim SC, Hong HP, Park CY, Shin
MS, Kim CJ, Seo JH, Kim DY, Kim DJ and Cho HJ: Treadmill exercise
ameliorates dopaminergic neuronal loss through suppressing
microglial activation in Parkinson's disease mice. Life Sci.
91:1309–1316. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Riedl SJ and Shi Y: Molecular mechanisms
of caspase regulation during apoptosis. Nat Rev Mol Cell Biol.
5:897–907. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rami A: Ischemic neuronal death in the rat
hippocampus: The calpain-calpastatin-caspase hypothesis. Neurobiol
Dis. 13:75–88. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Okuno S, Saito A, Hayashi T and Chan PH:
The c-Jun N-terminal protein kinase signaling pathway mediates Bax
activation and subsequent neuronal apoptosis through interaction
with Bim after transient focal cerebral ischemia. J Neurosci.
24:7879–7887. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen MH, Ren QX, Yang WF, Chen XL, Lu C
and Sun J: Influences of HIF-lα on Bax/Bcl-2 and VEGF expressions
in rats with spinal cord injury. Int J Clin Exp Pathol.
6:2312–2322. 2013.PubMed/NCBI
|
|
32
|
Li Y, Gu J, Liu Y, Long H, Wang G, Yin G
and Fan J: iNOS participates in apoptosis of spinal cord neurons
via p-BAD dephosphorylation following ischemia/reperfusion (I/R)
injury in rat spinal cord. Neurosci Lett. 545:117–122. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mao Z, Zheng YL, Zhang YQ, Han BP, Zhu XW,
Chang Q and Hu XB: The anti-apoptosis effects of daidzein in the
brain of D-galactose treated mice. Molecules. 12:1455–1470. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Han BJ, Li W, Jiang GB, Lai SH, Zhang C,
Zeng CC and Liu YJ: Effects of daidzein in regards to cytotoxicity
in vitro, apoptosis, reactive oxygen species level, cell cycle
arrest and the expression of caspase and Bcl-2 family proteins.
Oncol Rep. 34:1115–1120. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chakrabarti M, Das A, Samantaray S, Smith
JA, Banik NL, Haque A and Ray SK: Molecular mechanisms of estrogen
for neuroprotection in spinal cord injury and traumatic brain
injury. Rev Neurosci. 27:271–281. 2016.PubMed/NCBI
|
|
36
|
Yu F, Sugawara T, Maier CM, Hsieh LB and
Chan PH: Akt/Bad signaling and motor neuron survival after spinal
cord injury. Neurobiol Dis. 20:491–499. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim MH, Park JS, Seo MS, Jung JW, Lee YS
and Kang KS: Genistein and daidzein repress adipogenic
differentiation of human adipose tissue-derived mesenchymal stem
cells via Wnt/β-catenin signalling or lipolysis. Cell Prolif.
43:594–605. 2010. View Article : Google Scholar : PubMed/NCBI
|