Introduction
Aging is one of the most basic natural laws in the
biological world. Skin aging is an important indicator of human
aging that results from a combination of internal and external
environmental factors (1). Aging
caused by internal factors is called natural aging. Notably, the
exogenous environment may also contribute to human skin aging. Skin
aging may also be caused by ultraviolet (UV) radiation, which is
called photoaging (2). Furthermore,
ultraviolet A (UVA) is the ultraviolet light with the longest
wavelength that can reach the dermal layer of the skin and is a
major contributor of photoaging (3).
Typical characteristics of photoaging include deep
and rough wrinkles, pigmentation spots, dry and loose skin atrophy,
telangiectasia, photoaging purpura and precancerous lesions
(4). Notably, photoaging may result
in more severe outcomes than natural aging; for example, long-term
ultraviolet radiation may trigger skin cancer (5). In general, molecular changes in
photoaging are thought to be an enhancement and amplification of
molecular changes associated with age-related skin aging (6).
Dermal fibroblasts are crucial cellular components
involved in the structural integrity of the skin (7). They serve as the primary producer of
the extracellular matrix scaffold within the dermis, including
collagen, elastin and glycosaminoglycans (8). Studies have indicated that UV exposure
from the sun causes apoptosis of dermal fibroblasts (photodamage)
and contributes to the development of photoaging (9). Hyaluronic acid (HA), an extracellular
matrix molecule synthesized by hyaluronic acid synthase enzymes
(HAS), serves a role in regulating apoptosis in fibroblasts
(10). Injection of HA fillers
provides enrichment of one of the primary ECM compounds, deep
hydration of the skin and strongly stimulates fibroblasts, which
act on specific receptors cluster of differentiation (CD)44,
HA-mediated cell motility and intercellular adhesion molecule-1 to
synthesize novel scaffold compounds (11,12).
UV has been demonstrated to activate the MAPK
signaling pathway and increase the expression of matrix
metalloproteinases (MMPs), which promote degradation of collagen
(13). Large-scale decomposition of
collagen is the basis of photoaging (14,15).
UV light is absorbed by the skin molecules, and may
produce a large number of harmful compounds, including hydrogen
peroxide and superoxide anions, which are called reactive oxygen
species (ROS). These promote oxidative damage to cell components,
including cell walls, lipid membranes, mitochondria and DNA, and
promote apoptosis, cellular aging and inflammation (2). ROS may also influence the photoaging
process indirectly by affecting MAPK and transforming growth
factor-β signaling (9). The
mechanism of photoaging is complex and various regulatory signaling
pathways have been associated with this process.
Polycomb group genes (PcG) encode chromatin
modification proteins that control development and have key roles
in the regulation mechanisms of cell proliferation,
differentiation, aging and tumorigenesis (16). Because PcG relies on multiple protein
complexes, it is called multiple comb inhibition complexes (PRCs).
Polycomb repressive complex (PRC)1 and PRC2 are the active
components of PcG (17).
In the PcG gene family, the subunit E(z) or EZH2 of
the PRC2 complex serves a central role in gene regulation (18). EZH2 is involved in the formation of
the chromatin structure, regulation of gene expression and growth
control, and thus has pluripotency (19). Previous studies have indicated that
the appearance of senescence-associated heterochromatic foci is
associated with the increase of H3K9 methylation level in aging
human fibroblasts (20).
The PRC1 complex subunit BMI-1 is a negative
regulator of the inhibitor of CDK4/alternative reading frame
(Ink4a/Arf) locus, which encodes cell cycle regulatory proteins and
tumor suppressors, including P16Ink4a and P19Arf (21). However, little research exists on PcG
epigenetic regulation in the photoaging process in human skin.
Therefore, the present study aimed to investigate whether EZH2 and
BMI-1 are involved in the regulation of UVA radiation-induced
photoaging in human skin fibroblasts (HSFs). The effects of
inhibitors on UVA radiation-induced photoaging in fibroblasts was
investigated using the EZH2 inhibitor, GSK126. The results of the
present study may provide a novel theoretical basis for the
research and treatment of skin photoaging.
Materials and methods
Cell culture
HSFs were purchased from CoBioer Biosciences Co.,
Ltd. (Nanjing, China). The cells were cultured in Dulbecco's
modified Eagle medium (DMEM) (Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum
(ScienCell Research Laboratories, Inc., Carlsbad, CA USA), 1%
penicillin and 1% streptomycin. The cells were incubated under 5%
CO2 at 37°C.
Chemicals
GSK126 was purchased from Selleck Chemicals
Scientific, Inc., Houston, TX, USA and dissolved in dimethyl
sulfoxide. GSK126 was stored at −20°C in a freezer and diluted with
medium to a final concentration (2 µM) for experiments.
HSFs (1×105 cells/well) were seeded in
6-well plates with DMEM supplemented with 10% fetal bovine serum,
1% penicillin and 1% streptomycin. Cells were incubated in an
atmosphere containing 5% CO2 at 37°C for 24 h. The
medium was replaced with different concentrations (0, 2, 4 and 8
µM) of GSK126. Once the cells were further incubated for 24 h at
37°C, the cells were collected for RNA extraction and further
experiments.
HSFs (1×105 cells/well) were seeded in
6-well with DMEM containing 10% fetal bovine serum, 1% penicillin
and 1% streptomycin. The cells were incubated in an atmosphere
containing 5% CO2 at 37°C for 24 h. Cells were divided
into four groups: (−), cells without UVA and GSK126; GSK126 (2 µM);
UVA (10 J/cm2); and UVA (10 J/cm2)+GSK126 (2
µM). The cell culture medium was replaced with an equal volume of
sterile phosphate-buffered saline (PBS) in all groups (prior to
radiation exposure in the irradiated groups). The radiation dose
used was 10 J/cm2 in the UVA (10 J/cm2) and
UVA (10 J/cm2)+GSK126 (2 µM) groups. The PBS was
discarded at the end of UVA irradiation, media containing serum and
antibiotics were added to the (−) and UVA (10
J/cm2)+GSK126 (2 µM) groups, media containing serum and
antibiotics and 2 µM GSK126 was added to GSK126 (2 µM) and UVA (10
J/cm2)+GSK126 (2 µM) groups, and cells were further
incubated at 37°C for 24 h. Images of the cells were then captured
with an optical microscope (magnification, ×40) to observe the cell
growth and collected for RNA extraction and further
experiments.
UVA irradiation
The ZF-1 Three Ultraviolet analyzer (Shanghai HQ
Instruments Co., Ltd., Shanghai, China) was used as a radiation
source, the wavelength was set to 365 nm and an UV radiation meter
(Kühnast Strahlungstechnik, Dresden, Germany) was used to measure
the radiation dose. According to the experiment, the appropriate
number of HSFs were inoculated onto the corresponding orifice
plates and, 24 h later, the desired radiation dose
(J/cm2) was selected for UV irradiation. The cell
culture medium was replaced with an equal volume of sterile PBS
prior to radiation exposure. The cells were placed under the UV
analyzer for irradiation under sterile conditions and the
irradiation height was <2 cm. Following irradiation, PBS was
replaced with fresh medium.
Cell viability assay
A cell counting kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) assay was used to determine
cell viability, according to the manufacturer's instructions. It is
based on dehydrogenase activity detection in viable cells. The
formazan dye generated by dehydrogenases absorbs light at a
wavelength of 450 nm. The amount of formazan dye in cells is
directly proportional to the number of living cells.
HSFs were seeded into 96-well plates at a density of
0.5×104 cells/well, with 200 µl of culture medium per
well in triplicate wells for each experimental group. When cells
were adherent (24 h post seeding), they were divided into four
different UVA treatment groups: 0, 2.5, 5 and 10 J/cm2.
At specific time points after irradiation (0, 12 and 24 h), 20 µl
of CCK-8 reagent was added to each well. After incubation for 1, 2,
or 3 h at 37°C, absorbance was detected at 450 nm.
Hoechst staining
A Hoechst apoptosis staining kit (Beyotime Institute
of Biotechnology, Haimen, China) was used to determine cellular
apoptosis, according to the manufacturer's instructions. Under this
staining, apoptotic nuclei demonstrate a dense white staining with
fragmentation. Briefly, sterile cover slips were placed at the
bottom of 6-well plates, and 0.3×105 cells in 2 ml of
DMEM supplemented with 10% fetal bovine serum, 1% penicillin and 1%
streptomycin were added to each well. A total of 24 h after cell
irradiation, the medium was removed and 0.5 ml of fixative solution
was added. Cells were fixed for 10 min at room temperature. The
cells were then washed twice with PBS for 3 min each time and 0.5
ml Hoechst 33258 staining solution was added. The cells were
stained in a shaker for 5 min at room temperature and subsequently
washed twice with PBS for 3 min each time. The cells were then
carefully mounted onto a glass slide with a drop of fluorescence
quenching mounting liquid. The cells were then observed under a
fluorescence microscope.
Apoptosis detection by flow
cytometry
A total of 1.0×105 HSFs in 2 ml of DMEM
supplemented with 10% fetal bovine serum, 1% penicillin and 1%
streptomycin were added to each well of 6-well plates in triplicate
wells for each experimental group. When cells were adherent for 24
h, they were exposed to different doses of irradiation (0, 2.5, 5
and 10 J/cm2). HSFs were transferred into 5-ml tubes and
washed with PBS. An annexin V-FITC/PI apoptosis detection kit
(Nanjing KGI Biological Technology Development Co., Ltd., Nanjing,
China) was used to stain the nucleus according to the
maunfacturer's instructions and the cells were resuspended in
binding buffer (100 µl). Subsequently, cells were filtered into
tubes through filter paper to remove cell clumps. Following this,
50 µl Annexin V-fluorescein isothiocyanate solution and 50 µl
propidium iodide were added to each tube. The tubes containing the
cells were then incubated in the dark at room temperature for 15
min. The cells were then analyzed by BD FACS Aria II SORP sorting
flow cytometer (BD Biosciences, Franklin lake, NJ, USA). The
excitation and emission wavelengths were 488 and 530 nm,
respectively.
Senescence-associated β-galactosidase
(SA-β-gal) staining
SA-β-gal activity was determined 24 h after UVA
irradiation. A Senescence β-Galactosidase Staining Kit was
performed, according to the manufacturer's instructions (Beyotime
Institute of Biotechnology). A total of 0.3×105 cells in
2 ml of culture medium were added to each well of 6-well plates in
triplicate for each experimental group. A total of 24 h after cell
irradiation, cells were washed once with PBS at room temperature,
and 1 ml β-gal staining fixative solution was added to each well.
The cells were fixed for 15 min at room temperature. The fixative
solution was discarded and the cells were washed three times with
PBS, 3 min for each wash. Staining fluid (1 ml) was subsequently
added to each well. A plastic wrap was used to seal the 6-well
plates and cells were incubated overnight at 37°C. The following
day, the cells were observed under an optical microscope.
Erythrocyte exclusion experiment (a
particle exclusion assay)
Human peripheral blood (6 ml) was collected and the
plasma and leukocyte layer were separated following centrifugation
at 566 × g at 4°C for 5 min. Following this, erythrocytes were
washed three times with PBS, 3 min for each wash at 4°C. The
erythrocytes were fixed with 10% formaldehyde solution (5:1;
Nanjing KeyGen Biotech Co., Ltd., Nanjing, China) for 10 min at
room temperature, then erythrocytes were washed three times with
PBS, 3 min for each wash at 4°C and diluted to an erythrocyte
suspension at a final concentration of 107 cells/ml.
Cells were seeded in 6-well plates at 0.5×104
cells/well. Once the cells were irradiated for 24 h, 100 µl
eythrocyte suspension was added to each well and the samples were
incubated in an upright position for 5 min at 37°C. Fibroblasts
were observed using a particle exclusion assay and images were
captured using a Zeiss Axiovert phase-contrast microscope (22). Hyaluronidase (200 µl; 1 mg/ml;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was added to the 0
J/cm2 dose group, mixed gently, and incubated for 5 min
at 37°C before fibroblasts were observed under a microscope and
images were captured.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from HSFs using TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's instructions, and quantified spectrophotometrically.
Total RNA (1 µg) from each sample was subjected to first-strand
cDNA synthesis using a PrimeScript™ RT reagent kit with gDNA Eraser
(Takara Biotechnology Co., Ltd., Dalian, China), according to the
kit's instructions. cDNA was generated after the removal of genomic
DNA by treatment at 42°C for 2 min and subjected to the following
thermocycling conditions: 37°C for 15 min, 85°C for 5 sec; and
maintenance at 4°C.
A Takara PCR amplification kit (Takara Biotechnology
Co., Ltd.) was used according to the manufacturer's protocol with a
CFX Real-Time PCR Detection system (Bio-Rad Laboratories, Inc.) for
qPCR. The fluorescent dye used was SYBR Green (Takara PCR
amplification kit; Takara Biotechnology Co., Ltd.). The reaction
conditions were as follows: Initial denaturation at 95°C for 3 min;
and 45 cycles of denaturation at 95°C for 15 sec, annealing at 54°C
for 20 sec and extension at 72°C for 30 sec. The primers used are
demonstrated in Table I. Gene
expression was normalized to the level of GAPDH in a given sample.
The expression level of genes and gene alternative splicing
products were calculated and analyzed using the 2−ΔΔCq
relative quantification method (23). Each experimental treatment was
conducted in triplicate.
 | Table I.Primers and conditions for polymerase
chain reaction analysis. |
Table I.
Primers and conditions for polymerase
chain reaction analysis.
| Gene | Direction | Primer
sequences |
|---|
| EZH2 | Forward |
5′-ATGCGACTGAGACAGCTCAA-3′ |
|
| Reverse |
5′-TGGGATGACTTGTGTTGGAA-3′ |
| BMI-1 | Forward |
5′-TGGATCGGAAAGTAAACAAAGAC-3′ |
|
| Reverse |
5′-TGCATCACAGTCATTGCTGCT-3′ |
| HAS1 | Forward |
5′-TACAACCAGAAGTTCCTGGG-3′ |
|
| Reverse |
5′-CTGGAGGTGTACTTGGTAGC-3′ |
| HAS2 | Forward |
5′-GTGGATTATGTACAGGTTTGTGA-3′ |
|
| Reverse |
5′-TCCAACCATGGGATCTTCTT-3′ |
| HAS3 | Forward |
5′-GAGATGTCCAGATCCTCAACAA-3′ |
|
| Reverse |
5′-CCCACTAATACACTGCACAC-3′ |
| Hyal-1 | Forward |
5′-CCAAGGAATCATGTCAGGCCATCAA-3′ |
|
| Reverse |
5′-CCCACTGGTCACGTTCAGG-3′ |
| Hyal-2 | Forward |
5′-GGCTTAGTGAGATGGACCTC-3′ |
|
| Reverse |
5′-CCGTGTCAGGTAATCTTTGAG-3′ |
| CD44 | Forward |
GCTATTGAAAGCCTTGCAGAG |
|
| Reverse |
CGCAGATCGATTTGAATATAACC |
| EGF | Forward |
AGTTTTTCTGAATGGGTCAAGG |
|
| Reverse |
TCCAATTTATTGCCATTCCAG |
| EGFR | Forward |
ATGAGATGGAGGAAGACGG |
|
| Reverse |
CGGCAGGATGTGGAGAT |
| Smad2 | Forward |
GGAGCAGAATACCGAAGGCA |
|
| Reverse |
CTTGAGCAACGCACTGAAGG |
| Smad4 | Forward |
ATGACTTTGAGGGACAGC |
|
| Reverse |
GGAAGCCACAGGAATG |
| P16 | Forward |
GGAGCAGCATGGAGCCTTC |
|
| Reverse |
CATCATCATGACCTGGATC |
| MMP-1 | Forward |
TGTGGTGTCTCACAGCTTCC |
|
| Reverse |
CTTGCCTCCCATCATTCTTC |
| GAPDH | Forward |
GTGAAGGTCGGAGTCAACG |
|
| Reverse |
TGAGGTCAATGAAGGGGTC |
Statistical analysis
All data are presented as mean ± standard deviation.
Data was analyzed using SPSS software (version 22.0; IBM Corp.,
Armonk, NY, USA). Analysis of variance followed by Dunnett's test
was performed. P<0.05 was considered to indicate a statistically
significant difference.
Results
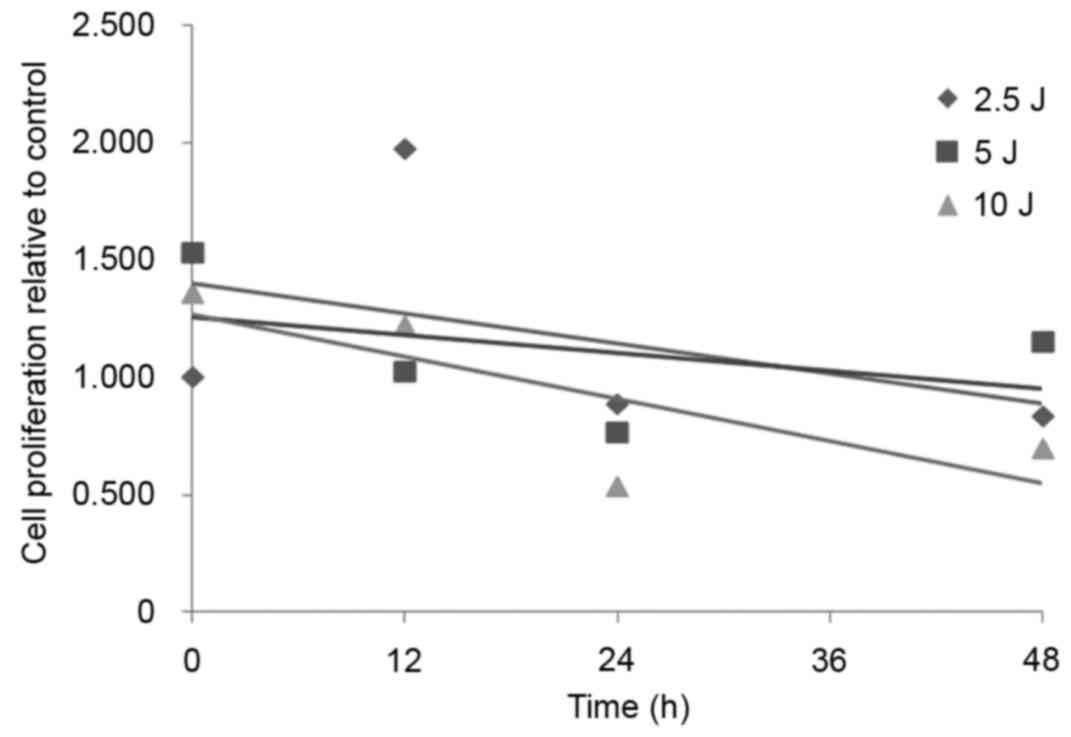
UVA radiation reduces fibroblast
proliferation
As demonstrated in Fig.
1, the fibroblast proliferation activity in the 2.5, 5 and 10
J/cm2 dose groups decreased gradually with the increase
of UVA radiation dose. The 10 J/cm2 UVA radiation dose
had the greatest effect on cell viability, indicating that 10
J/cm2 of UVA radiation is able to markedly inhibit
fibroblast proliferation (Fig.
1).
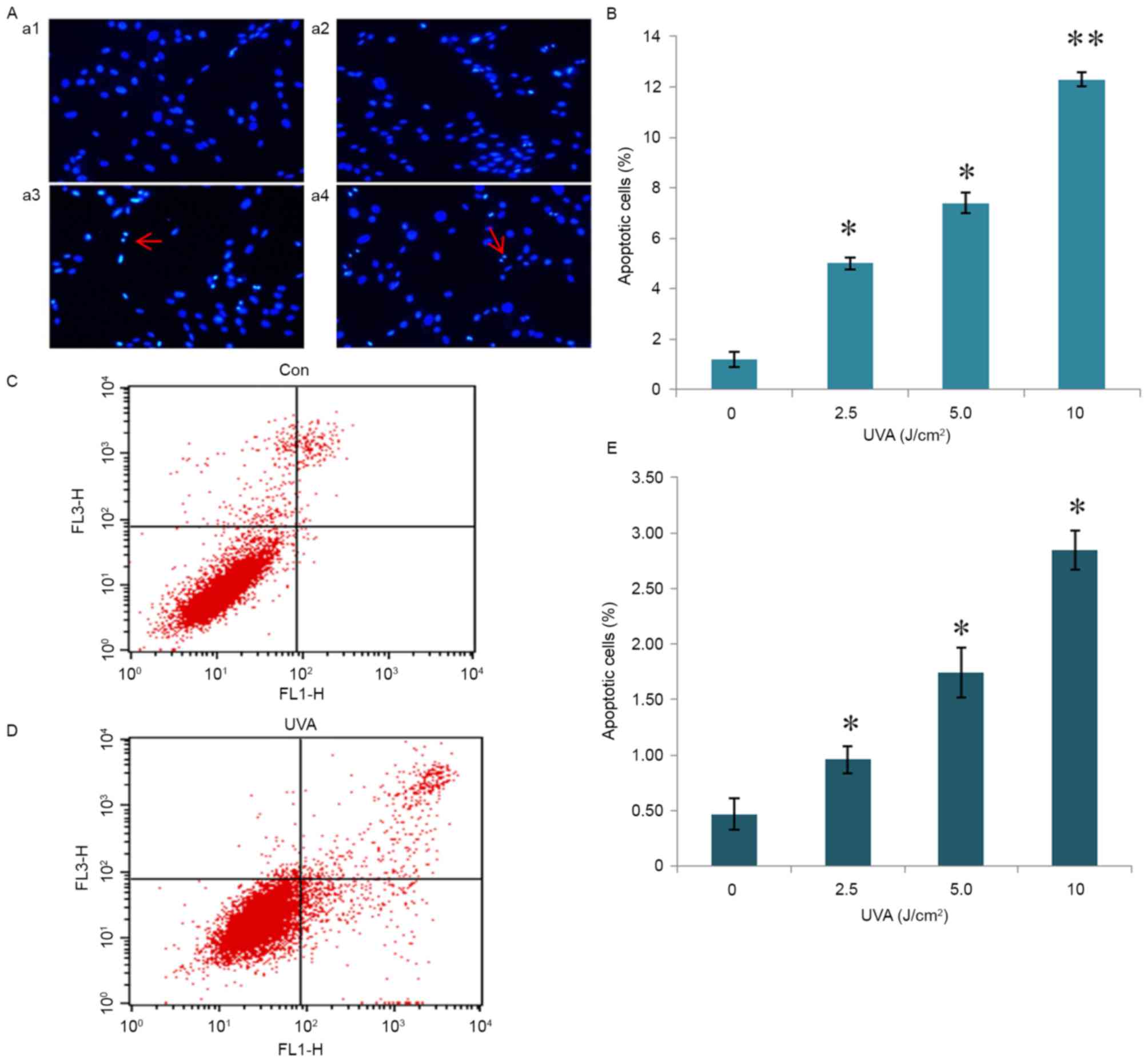
UVA radiation promotes fibroblast
apoptosis
Hoechst staining revealed that cells treated with
UVA radiation demonstrated apoptotic nuclei, with a dense white dye
or crushed nuclei shape (Fig. 2A).
Statistical analysis demonstrated that all doses of UVA radiation
(2.5, 5 and 10 J/cm2) significantly promoted fibroblast
apoptosis (P<0.05) compared with the control group (0
J/cm2) in a dose-dependent manner. The 10
J/cm2 dose promoted fibroblast apoptosis with the most
significant effect (P<0.01; Fig.
2B). The 10 J/cm2 UVA radiation dose demonstrated
the most marked promotion of fibroblast apoptosis compared with the
control group (Fig. 2C and D). The
detection of apoptosis by flow cytometry demonstrated that the
number of all UVA radiation groups significantly increased compared
to the control group (P<0.05; Fig.
2E).
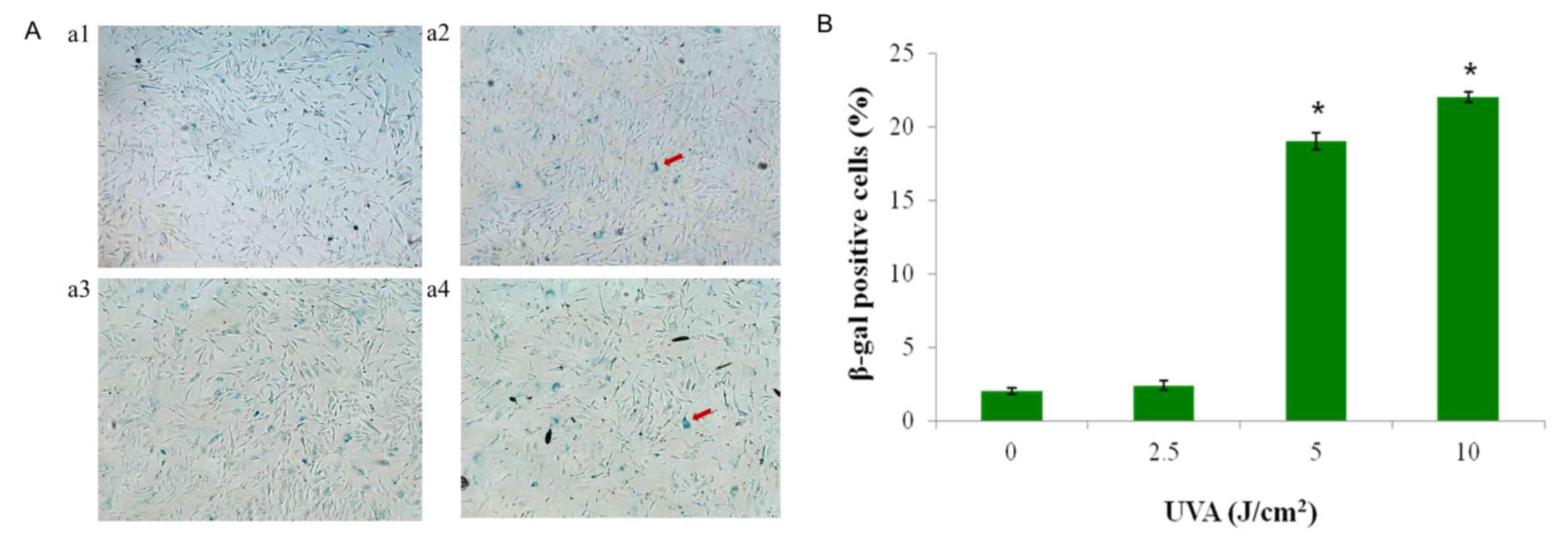
UVA radiation induces fibroblast
senescence
As demonstrated in Fig.
3A, the blue precipitate indicated β-gal activity and the
presence of senescent cells. UVA radiation induced changes in HSF
cellular senescence. The 5 and 10 J/cm2 dose radiation
groups significantly induced cellular senescence compared with the
control group (P<0.05). The 10 J/cm2 dose had the
most significant effect on cellular senescence, whereas the 2.5
J/cm2 radiation group had no significant effect on
senescence compared with the control (0 J/cm2) group
(Fig. 3B).
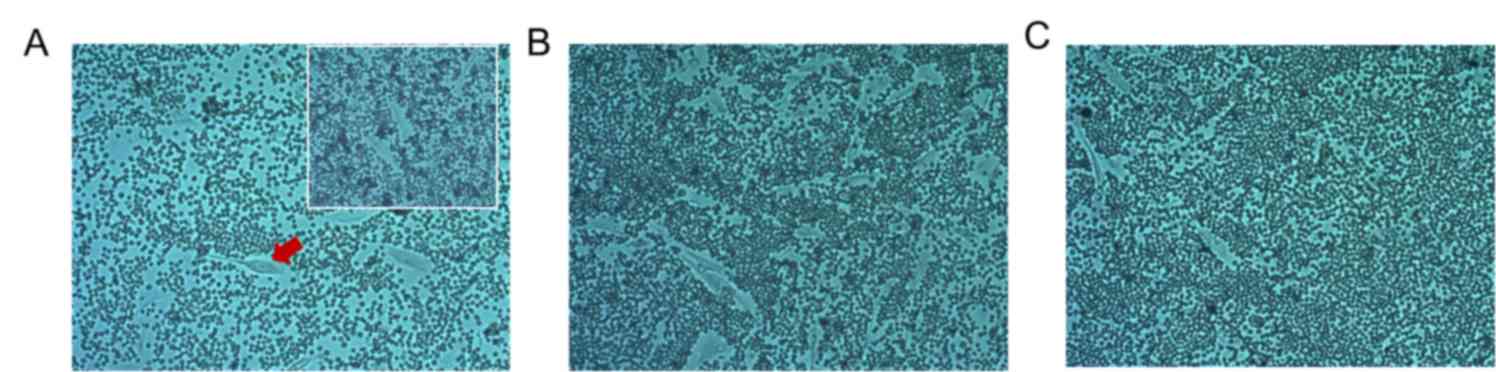
UVA radiation reduces hyaluronic acid
(HA) content
A particle exclusion assay demonstrated that UVA
radiation decreased HA content around HSF cells. Compared with the
control group (0 J/cm2), HSF cell exclusion to
erythrocytes in the 10 J/cm2 dose group decreased. The
exclusion in the 0 J/cm2 group disappeared after adding
100 µl hyaluronidase (1 mg/ml), indicating that the exclusion
effect of red blood cells was caused by HA formation (Fig. 4). As skin aging is predominantly a
result of extracellular matrix reduction, of which, HA is one of
the most important components, the decline of hyaluronan content is
a sign of skin aging (24). Notably,
fibroblasts are the primary generators of HA (8). In the present experiment, UVA radiation
was indicated to reduce the synthesis of HA in fibroblasts, which
demonstrated that a photoaging cell model was successfully
established.
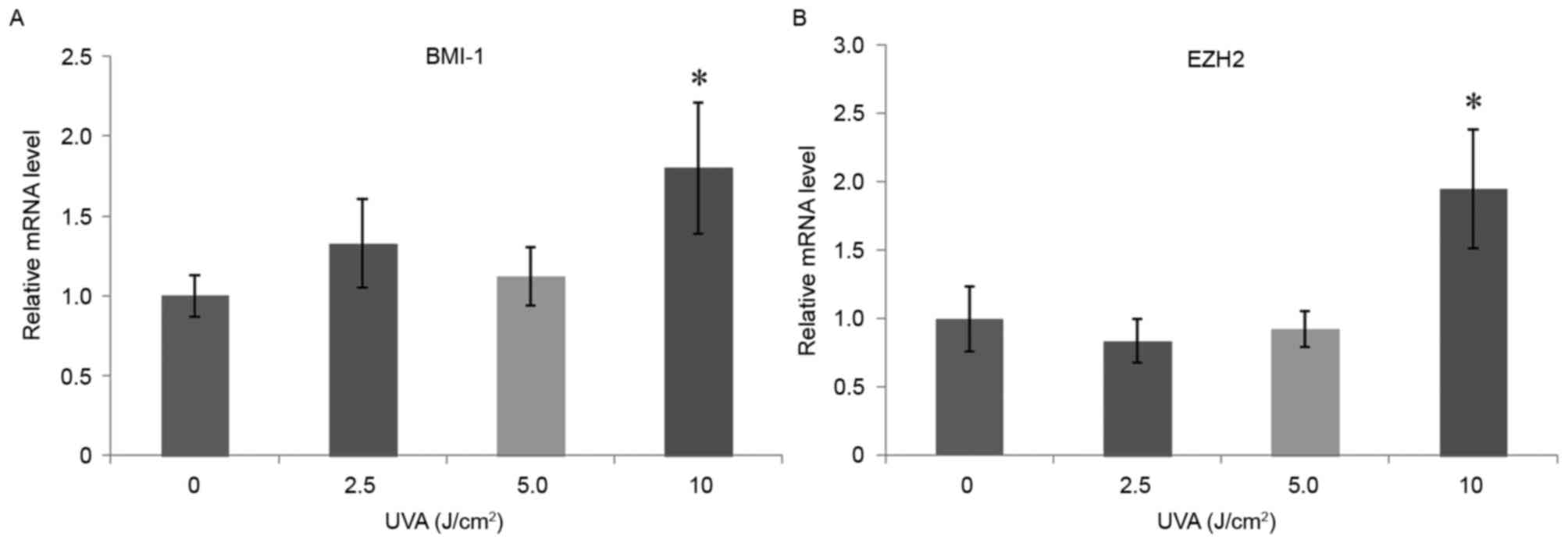
High-dose UVA radiation increases mRNA
expression levels of BMI-1 and EZH2
RT-qPCR results demonstrated that, 24 h after UVA
radiation of HSF cells, BMI-1 and EZH2 mRNA expression levels in
the 10 J/cm2 dose group were significantly upregulated
(P<0.05) compared with the control group (0 J/cm2;
Fig. 5). However, the other dose
groups demonstrated no significant differences compared with the
control group. The 10 J/cm2 dose therefore had the most
significant effect on fibroblast proliferation, promoting
apoptosis, inducing senescence, reducing the surrounding HA content
and upregulating the mRNA expression levels of BMI-1 and EZH2.
Based on these results, the 10 J/cm2 dose was selected
as the UVA treatment dose for the photoaging model.
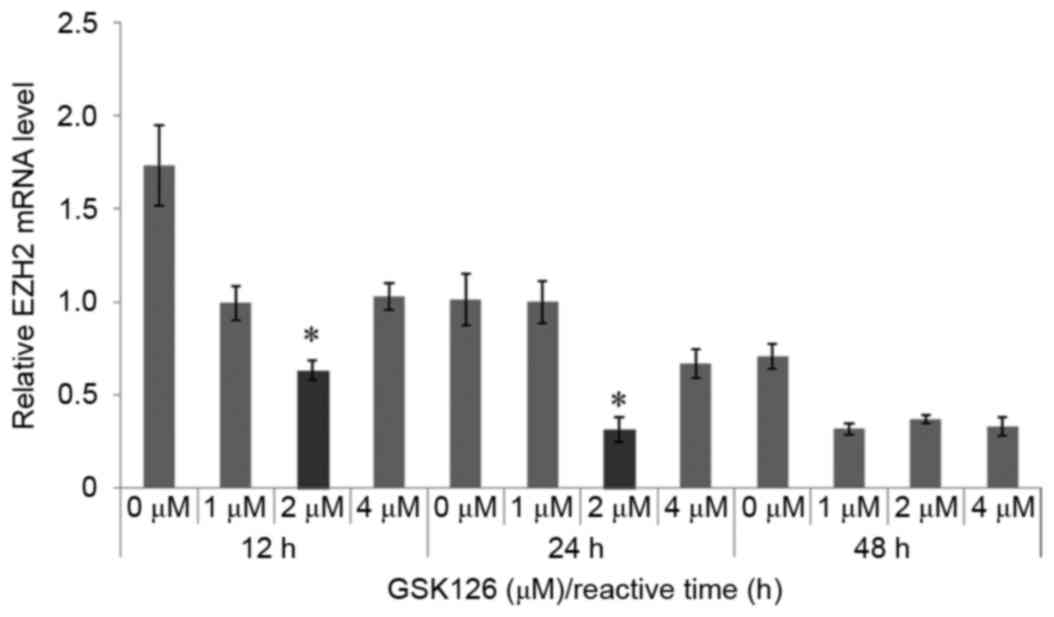
GSK126 inhibits EZH2 mRNA
expression
GSK126 is a methyl transferase activity inhibitor
that could selectively inhibit EZH2-methyl transferase activity. In
the present study, the results demonstrated that GSK126 also
inhibited the mRNA expression of EZH2. GSK126 at a concentration of
2 µM significantly inhibited EZH2 expression compared with
the control group (0 µM GSK126) at 12 and 24 h (P<0.05). The
mRNA expression of the EZH2 was significantly downregulated
after treatment of HSF cells for 12 and 24 h (P<0.05; Fig. 6). Therefore, 2 µM GSK126 was selected
as the inhibitor concentration for further analyses.
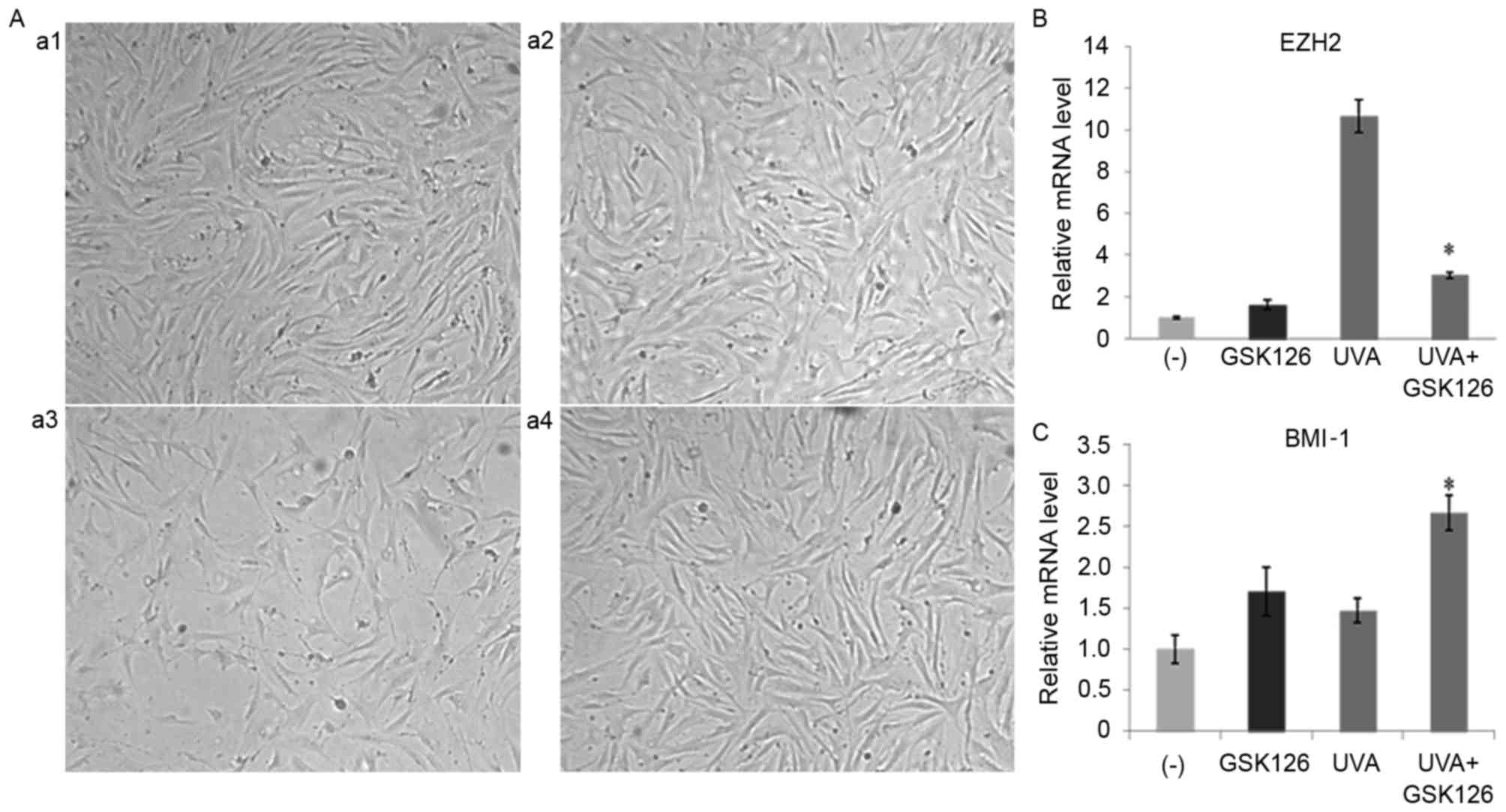
GSK126 inhibits photoaging in
fibroblasts
GSK126 inhibited the photoaging of fibroblasts as
well as expression of the key genes in the PcG family. After
treating cells for 48 h, cell growth state of the cells was
observed under a microscope. It was demonstrated that the addition
of 2 µM GSK126 did not significantly affect cell growth compared
with the control group. However, a decrease in cell number, a poor
growth state and UVA radiation-induced cellular senescence was
observed in fibroblasts in the 10 J/cm2 UVA irradiation
group. Cell growth was largely unaffected in the UVA+GSK126 group,
suggesting that inhibitors may resist UVA radiation-induced
fibroblast senescence (Fig. 7A).
RT-qPCR results demonstrated that the addition of GSK126 after UVA
radiation significantly reduced the increase of EZH2 induced by UVA
radiation (P<0.05; Fig. 7B).
Following radiation, there was a significant increase in BMI-1 mRNA
expression caused by the addition of GSK126 in the UVA+GSK126 group
compared with the UVA radiation group (P<0.05; Fig. 7C). This may be closely linked to
GSK126 resisting fibroblast senescence.
The effect of UVA radiation and GSK126 on HA
anabolic family-related gene expression was analyzed. RT-qPCR
analysis demonstrated that after 10 J/cm2 UVA radiation
on HSF cells, HA synthase 1 (HAS1), HAS2, HAS3, CD44 and
hyaluronidase (Hyal-1) were upregulated, with the highest
expression observed for HAS1, compared with the control group
without any treatment (P<0.05; Fig.
8A-E). After adding GSK126 inhibitors, the upregulation of HAS1
and Hyal-1 in the UVA group was significantly inhibited (P<0.05;
Fig. 8A). However, HAS2, HAS3, CD44
in the UVA+GSK126 group were significantly upregulated (P<0.05;
Fig. 8B-E) compared with the UVA
group, whereas Hyal-2 expression did not change significantly
change (Fig. 8F).
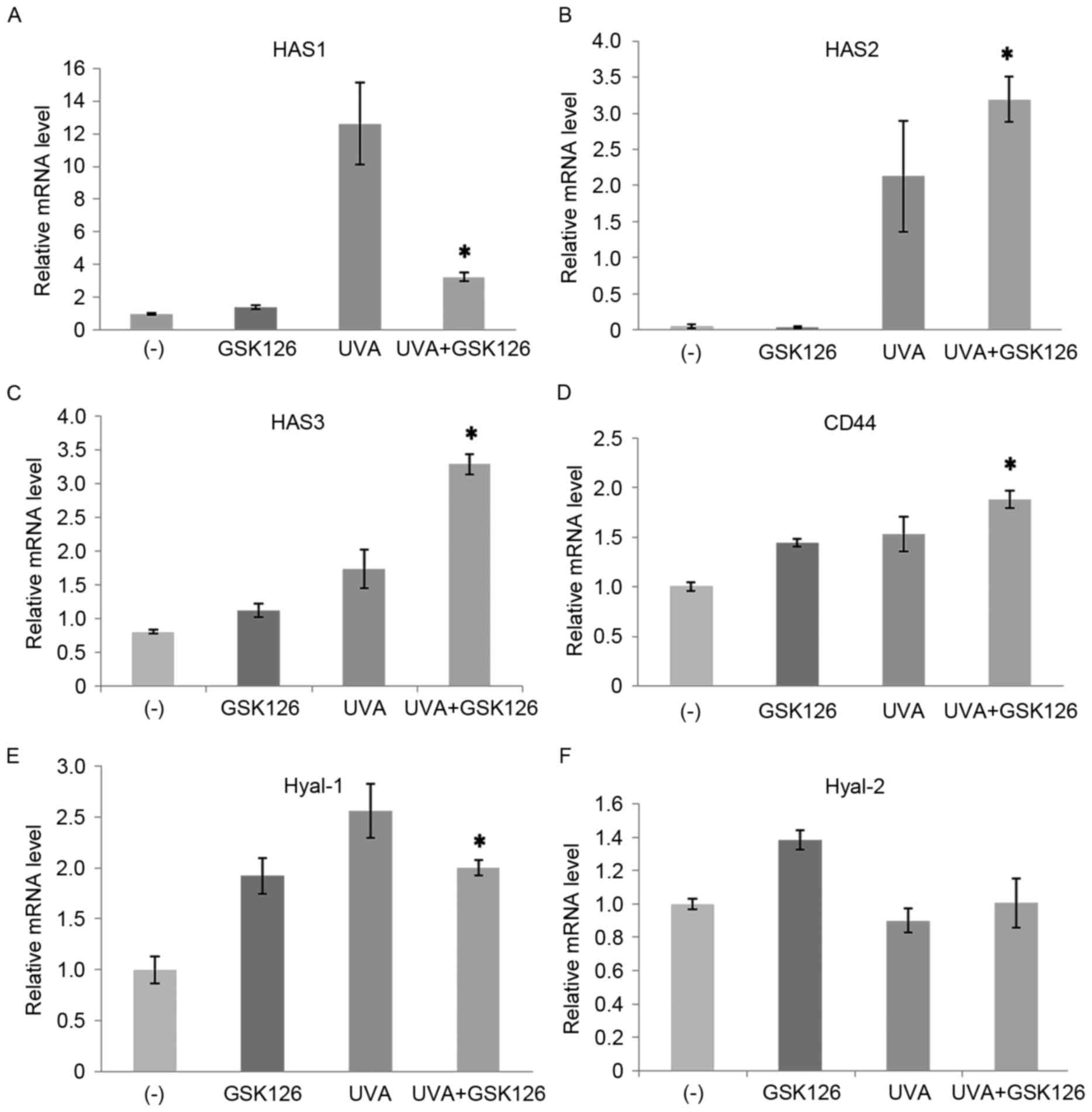 | Figure 8.Effect of UVA radiation and GSK126
inhibitor on fibroblast mRNA expression levels of (A) HAS1, (B)
HAS2, (C) HAS3, (D) CD44, (E) Hyal-1 and (F) Hyal-2. Data are
presented as the mean ± standard deviation. *P<0.05 vs. UVA (10
J/cm2). UVA, ultraviolet A; HAS, HA, hyaluronic acid
synthase; CD, cluster of differentiation; Hyal, hyaluronidase; (−),
control group without any treatment. |
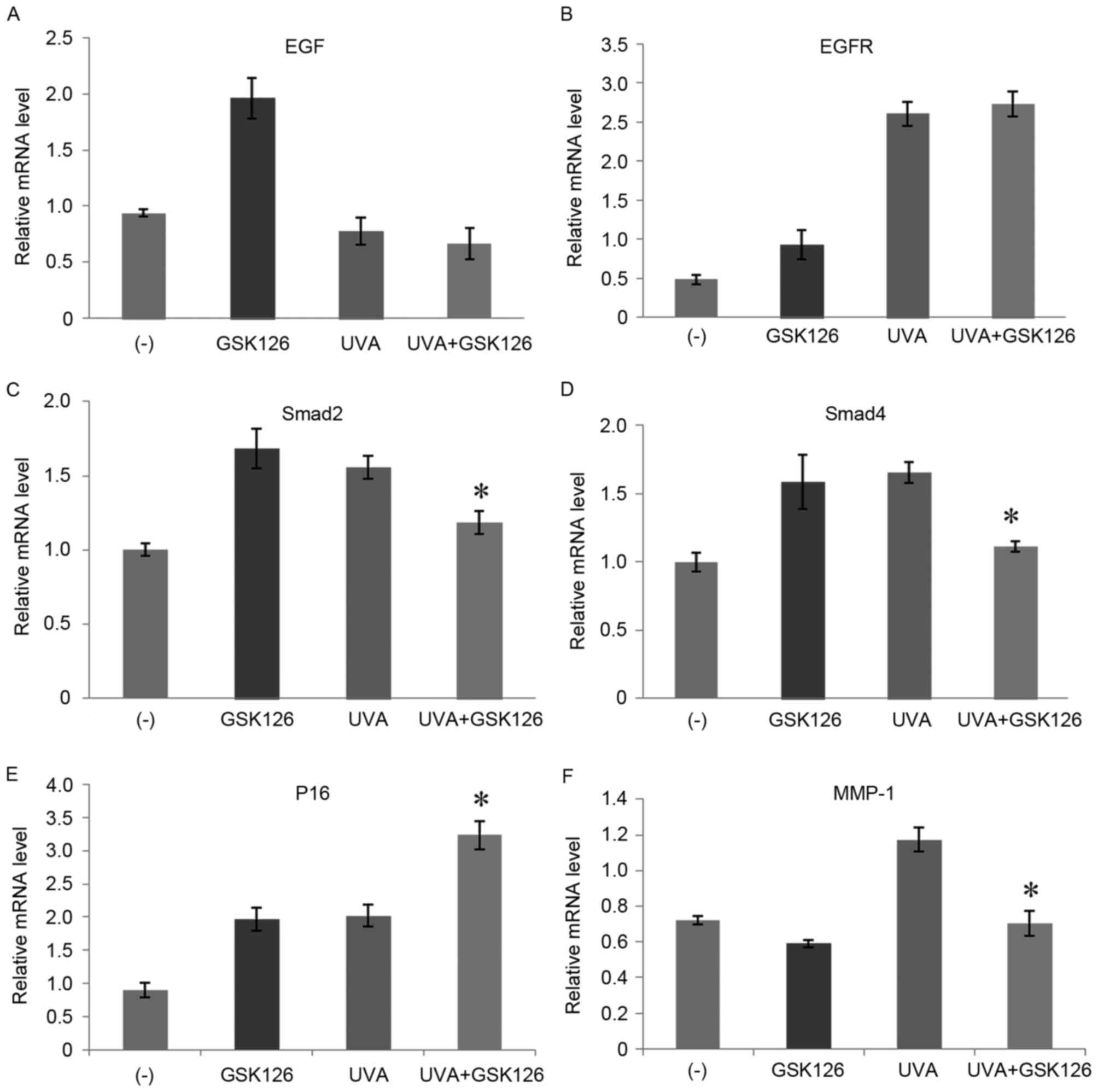
Following this, the effect of UVA radiation and
GSK126 on HSF cell photoaging molecular pathways was investigated
(Fig. 9). RT-qPCR demonstrated that
epidermal growth factor receptor (EGFR), smad2, smad4, P16 and
matrix metalloproteinase-1 (MMP-1) mRNA expression levels were
upregulated after 10 J/cm2 UVA radiation on HSF cells.
After the addition of GSK126, upregulation of smad2, smad4 and
MMP-1 was significantly inhibited (P<0.05; Fig. 9C, D and F). However, the upregulation
of P16 gene expression was further promoted with the addition of
GSK126 (P<0.05; Fig. 9E).
Addition of GSK126 had no significant effect on epidermal growth
factor (EGF) and EGFR expression after UVA radiation (Fig. 9A and B).
Discussion
Photoaging is the most common form of chronic
sun-induced skin damage due to prolonged exposure. Dermal
connective tissue changes induced by long-term UV irradiation lead
to light skin damage and the appearance of aging (25). As the primary producers of the ECM
scaffold, dermal fibroblasts are the basis of the occurrence of
photoaging (8,26,27). Due
to its long wavelength, UVA may reach the skin up to the dermis,
which is the main cause of photoaging (28). Therefore, the present study utilized
HSF cells, and UVA radiation of fibroblasts was used to establish
an in vitro aging model to observe the effect of UVA
radiation and GSK126 on HSF cell senescence. It was demonstrated
that HSF cell proliferation activity decreased, the percentage of
apoptotic cells increased significantly, cell senescence increased
and HA content around HSF cells decreased (i.e. the cell phenotype
changed considerably) after UVA radiation on fibroblasts at a dose
of 10 J/cm2. Niu et al (29) and Lan et al (30) demonstrated that UVA radiation induced
a significant decrease in fibroblast proliferation activity, which
is consistent with the results of the present study.
In the study of the photoaging mechanism, it was
previously demonstrated that UV radiation may lead to the apoptosis
of a large number of dermal fibroblasts (light damage), and
apoptosis promoted the development of photoaging (9,31).
Hoechst staining and flow cytometry methods were utilized in the
present study to determine whether UVA radiation was able to induce
fibroblast apoptosis, which was more obvious at higher radiation
doses. This demonstrated that UVA radiation may cause direct
fibroblast photodamage, resulting in fibroblast cell apoptosis;
however, determination of the molecular mechanisms involved
requires further investigation.
UVA radiation may reduce fibroblast HA synthesis and
promote the occurrence of aging. An erythrocyte exclusion
experiment was performed in the present study and it was determined
that the particle size exclusion effect of red blood cells
surrounding HSF cells decreased after UVA radiation. The size
exclusion effect to red blood cells in the 10 J/cm2 dose
group almost disappeared, indicating that UVA radiation may lead to
the decrease of HA content surrounding HSF cells.
The increase in β-gal activity levels within
senescent cells is an indicator of cellular senescence (32,33).
Fibroblast senescence is the basis of photoaging and UVA/UVB
radiation are able to induce fibroblast senescence. A study by Wang
et al (34) used β-gal
staining after UVA irradiation of HSFs for 24 h and demonstrated
that the percentage of senescent cells increased from 13.14±1.80 to
56.72±2.04%. In the present study, UVA radiation successfully
induced fibroblast senescence. The results also demonstrated that
UVA radiation induced a greater extent of HSF cell senescence at 10
J/cm2 compared to lower doses.
In the present study, HSFs were subjected to UVA
irradiation in order to induce apoptosis and senescence, thus
decreasing cell proliferation. The most notable effect of UVA
irradiation was observed at a dose of 10 J/cm2. In
addition, HA around the cells at this dose disappeared, and
therefore it was concluded that 10 J/cm2 UVA radiation
may successfully be applied to HSFs to establish an in vitro
model of photoaging. It was also demonstrated that EZH2 and BMI-1
mRNA expression levels were upregulated significantly after 10
J/cm2 of UVA radiation on HSF cells. Accordingly, it was
speculated that the occurrence of HSF cell photoaging was closely
related to the PcG family.
In the present study GSK126 was selected as the
interfering factor of HSF photoaging, and it was demonstrated that
fibroblasts restored growth after the addition of GSK126 following
the UVA radiation-induced aging state. Furthermore, GSK126
significantly inhibited the upregulation of EZH2 mRNA expression
induced by UVA radiation in photoaging HSF cells. It was suspected
that the downregulation of EZH2 by GSK126 resulted in the decrease
of H3K9 methylation levels and inhibited the formation of
aging-associated heterochromatic loci (20,35),
thus preventing fibroblast senescence.
BMI-1 is a negative regulator of the inhibitor of
Ink4a/Arf locus (36). The
overexpression of BMI-1 in BMI-1 deficient primary mouse embryonic
fibroblasts results in downregulation of P16 gene expression, as
well as promotion of cell immortalization and anti-aging (37). As a negative regulator of the cell
cycle, the increase in P16 is closely associated with cellular
aging (36). The present study
demonstrated that both BMI-1 and P16 expression after UVA radiation
of HSF cells in the photoaging model increased, while GSK126
further promoted the expression of BMI-1 and P16. It was
hypothesized that the simultaneous increase of BMI-1 and P16
interfered with the cell aging process. However, further studies
are required to determine the exact mechanism.
Dermis HA has a vital role in strong hydrophilic
interactions, keeping the skin moist and reducing the aging process
(38,39). The present study demonstrated that
the expression of all three HAS and CD44 in the UVA radiation HSF
cell photoaging model were upregulated, with HAS1 demonstrating the
highest expression. GSK126 significantly reduced the upregulation
of HAS1. The inhibitor also downregulated the expression of smad2
and smad4, whereas there was no significant difference in EGF and
EGFR expression compared with UVA-treated cells. Based on these
findings, it was hypothesized that GSK26 inhibits the
CD44-EGFR-extracellular signal-regulated kinase (ERK)
co-localization signal system by downregulating HAS1, smad2 and
smad4, thus inhibiting the differentiation of fibroblasts into
myofibroblasts and impeding scar repair. The upregulation of HAS2,
HAS3 and CD44 promoted the synthesis and secretion of HA, as well
as the proliferation and migration of fibroblasts, However, HAS2,
HAS3 and CD44 upregulation inhibited apoptosis and induced the
regeneration and adhesion of keratinocytes (39). Furthermore, HAS2 upregulation may
enhance the ability of HSF cells to block apoptosis. HAS3
upregulation has been demonstrated to promote the synthesis of a
large number of small molecular weight HA, accelerate blood
circulation and metabolism, and reduce the damage of reactive
oxygen species and other substances to cells (10).
MMPs are a class of enzymes that cause the reduction
of the ECM. The increased activity of MMPs may reduce the
expression of type I collagen, which is one of the causes of skin
aging (40,41). The results of the present study
demonstrated that MMP-1 gene expression was upregulated in UVA
irradiated HSF cells in the photoaging model, indicating that the
decomposition of collagen increased after UVA radiation. However,
GSK126 significantly suppressed the upregulation of MMP-1 gene
expression, thus reducing the degradation of collagen and resisting
the effect of UVA radiation on fibroblast aging.
Sustained oxidative stress and the DNA damage
response are closely related to cellular senescence (42). A previous study indicated that the
nuclear factor (NF1)/smad4 transcriptional repressor protein
complex was able to inhibit adenine nucleotide translocase 2 (ANT2)
transcription-mediated oxidative shock and DNA damage response in
aged fibroblasts (43). The results
of the present study demonstrated that smad4 gene expression was
upregulated in UVA irradiated HSF cells in the photoaging model.
However, GSK126 significantly inhibited this effect, thereby
reducing formation of the NF1/smad4 complex, promoting ANT2
transcription, reducing oxidative stress and the DNA damage
response, and blocking the oxidative stress effects mediated by UVA
radiation.
In conclusion, in the present study, PcG family
control was closely related to UVA radiation-induced fibroblast
photoaging. As a key gene, EZH2 had a vital role in the process of
photoaging. GSK126 inhibited histone methylation catalytic activity
and suppressed EZH2 gene expression. This inhibition may block UVA
radiation-induced cell HSF aging through various mechanisms, such
as reducing H3K9 methylation levels, inhibiting the formation of
aging-associated heterochromatic loci, and interfering with
cellular senescence by upregulating the expression of BMI-1 and P16
genes. Furthermore, GSK126 may differentially regulate the three
kinds of HAS to inhibit the differentiation of fibroblasts into
myofibroblasts, thus inhibiting scar repair, increasing the
synthesis and secretion of HA, promoting the proliferation and
migration of fibroblasts, accelerating blood circulation and
metabolism and enhancing the ability of HSF cells against
apoptosis. GSK1 also inhibited MMP-1 gene expression to reduce the
degradation of collagen, inhibited Hyal-1 gene expression to reduce
the degradation of HA, inhibited the expression of the smad4 gene
to promote ANT2 transcription, reduced oxidative stress and DNA
damage. However, the experimental study only measured gene
expression. Therefore, analysis of the specific regulation modes
and mechanisms of the PcG family is required for future
studies.
Acknowledgements
The present study was supported by the National
Natural Foundation of China (The Correlation Study between Chronic
Delayed Wound Healing in Rats and Pseudomonas Aeruginosa Biofilm;
grant no. 81372068).
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Krutmann J, Bouloc A, Sore G, Bernard BA
and Passeron T: The skin aging exposome. J Dermatol Sci.
85:152–161. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Peres PS, Terra VA, Guarnier FA, Cecchini
R and Cecchini AL: Photoaging and chronological aging profile:
Understanding oxidation of the skin. J Photochem Photobiol B.
103:93–97. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li W, Zhou BR, Hua LJ, Guo Z and Luo D:
Differential miRNA profile on photoaged primary human fibroblasts
irradiated with ultraviolet A. Tumour Biol. 34:3491–500. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gilchrest BA: Photoaging. J Invest
Dermatol. 133:E2–E6. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lavker RM: Cutaneous aging: Chronologic
versus photoaging = Photodamage. Gilchrest B: 1st. Wiley-Blackwell;
Cambridge, MA: pp. 123–135. 1995
|
|
6
|
Helfrich YR, Sachs DL and Voorhees JJ:
Overview of skin aging and photoaging. Dermatol Nurs. 20:177–184.
2008.PubMed/NCBI
|
|
7
|
Thangapazham RL, Darling TN and Meyerle J:
Alteration of skin properties with autologous dermal fibroblasts.
Int J Mol Sci. 15:8407–8427. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wong T, McGrath JA and Navsaria H: The
role of fibroblasts in tissue engineering and regeneration. Br J
Dermatol. 156:1149–1155. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fisher GJ, Kang S, Varani J, Bata-Csorgo
Z, Wan Y, Datta S and Voorhees JJ: Mechanisms of photoaging and
chronological skin aging. Arch Dermatol. 138:1462–1470. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Y, Lauer ME, Anand S, Mack JA and
Maytin EV: Hyaluronan synthase 2 protects skin fibroblasts against
apoptosis induced by environmental stress. J Biol Chem.
289:32253–32265. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Entwistle J, Hall CL and Turley EA: HA
receptors: Regulators of signalling to the cytoskeleton. J Cell
Biochem. 61:569–577. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ghersetich I: Management of aging skin. J
Eur Acad Dermatol Venereol. 9:511997. View Article : Google Scholar
|
|
13
|
Chen T, Hou H, Fan Y, Wang S, Chen Q, Si L
and Li B: Protective effect of gelatin peptides from pacific cod
skin against photoaging by inhibiting the expression of MMPs via
MAPK signaling pathway. J Photochem Photobiol B. 165:34–41. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Quan T, Qin Z, Xia W, Shao Y, Voorhees JJ
and Fisher GJ: Matrix-degrading metalloproteinases in photoaging. J
Investig Dermatol Symp Proc. 14:pp. 20–24. 2009; View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hsu LY, Nien CY, Huang WM, Hsu SC and
Chang TC: Synthesis and protective effects of
bis{4-[N,N-di-(carboxymethyl)amino]phenoxy}alkane derivatives on
UVA-induced production of MMP-1 in human skin fibroblasts. Chem
Pharm Bull (Tokyo). 62:867–874. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Otte AP and Kwaks TH: Gene repression by
polycomb group protein complexes: A distinct complex for every
occasion? Curr Opin Genet Dev. 13:448–454. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Golbabapour S, Majid NA, Hassandarvish P,
Hajrezaie M, Abdulla MA and Hadi AH: Gene silencing and Polycomb
group proteins: An overview of their structure, mechanisms
andphylogenetics. OMICS. 17:283–296. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Koubi M, Chabannon C and Duprez E: The
biological complexity of Polycomb group proteins: The case of EZH2.
Med Sci (Paris). 33:499–505. 2017.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gall Trošelj K, Novak Kujundzic R and
Ugarkovic D: Polycomb repressive complex's evolutionary conserved
function: The role of EZH2 status and cellular background. Clin
Epigenetics. 8:552016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Narita M, Nũnez S, Heard E, Narita M, Lin
AW, Hearn SA, Spector DL, Hannon GJ and Lowe SW: Rb-mediated matin
formation and silencing of E2F target genes during cellular
senescence. Cell. 113:703–716. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jacobs JL and van Lohuizen M: Polycomb
repression: From cellular memory to cellular proliferation and
cancer. Biochim Biophys Acta. 1602:151–161. 2002.PubMed/NCBI
|
|
22
|
Bernert B, Porsch H and Heldin P:
Hyaluronan synthase 2 (HAS2) promotes breast cancer cell invasion
by suppression of tissue metalloproteinase inhibitor 1 (TIMP-1). J
Biol Chem. 286:42349–42359. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nobile V, Buonocore D, Michelotti A and
Marzatico F: Anti-aging and filling efficacy of six types
hyaluronic acid based dermo-cosmetic treatment: Double blind,
randomized clinical trial of efficacy and safety. J Cosmet
Dermatol. 13:277–287. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kurban RS and Bhawan J: Histologic changes
in skin associated with aging. J Dermatol Surg Oncol. 16:908–914.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Quan T, Wang F, Shao Y, Rittié L, Xia W,
Orringer JS, Voorhees JJ and Fisher GJ: Enhancing structural
support of the dermal microenvironment activates fibroblasts,
endothelial cells and keratinocytes in aged human skin in vivo. J
Invest Dermatol. 133:658–667. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Werner S, Krieg T and Smola H:
Keratinocyte-fibroblast interactions in wound healing. J Invest
Dermatol. 127:998–1008. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Philips N, Tuason M, Chang T, Lin Y, Tahir
M and Rodriguez SG: Differential effects of ceramide on cell
via-bility and extracellular matrix remodeling in keratinocytes and
fibroblasts. Skin Pharmacol Physiol. 22:151–157. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Niu T, Tian Y, Cai Q, Ren Q and Wei L: Red
light combined with blue light irradiation regulates proliferation
and apoptosis in skin keratinocytes in combination with low
concentrations of curcumin. PLoS One. 10:e01387542015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lan CC, Ho PY, Wu CS, Yang RC and Yu HS:
LED 590 nm photomodulation reduces UVA-induced metalloproteinase-1
expression via upregulation of antioxidant enzyme catalase. J
Dermatol Sci. 78:125–132. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li DX, Deng TZ, Lv J and Ke J: Advanced
glycation end products (AGEs) and their receptor (RAGE) induce
apoptosis of periodontal ligament fibroblasts. Braz J Med Biol Res.
47:1036–1043. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang JW, Zhang SS, Song JR, Sun K, Zong
C, Zhao QD, Liu WT, Li R, Wu MC and Wei LX: Autophagy inhibition
switches low-dose camptothecin-induced premature senescence to
apoptosis in human colorectal cancer cells. Biochem Pharmacol.
90:265–275. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yu A, Zheng Y, Zhang R, Huang J, Zhu Z,
Zhou R, Jin D and Yang Z: Resistin impairs SIRT1 function and
induces senescence-associated phenotype in hepatocytes. Mol Cell
Endocrinol. 377:23–32. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang YN, Wu W, Chen HC and Fang H:
Genistein protects against UVB-induced senesc-ence-like
characteristics in human dermal fibroblast by p66Shc
down-regulation. J Dermatol Sci. 58:19–27. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Aksoy O, Chicas A, Zeng T, Zhao Z,
McCurrach M, Wang X and Lowe SW: The atypical E2F family member
E2F7 couples the p53 and RB pathways during cellular senescence.
Genes Dev. 26:1546–1557. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dhawan S, Tschen SI and Bhushan A: Bmi-1
regulates the Ink4a/Arf locus to control pancreatic beta-cell
proliferation. Genes Dev. 23:906–911. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jacobs JJ, Kieboom K, Marino S, DePinho RA
and van Lohuizen M: The oncogene and Polycomb-group gene bmi-1
regulates cell proliferation and senescence through the ink4a
locus. Nature. 397:164–168. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lee BM, Han DG and Choi WS: Rejuvenating
effects of facial Hydrofilling using Restylane vital. Arch Plast
Surg. 42:282–287. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Simpson RM, Wells A, Thomas D, Stephens P,
Steadman R and Phillips A: Aging fibroblasts resist phenotypic
maturation because of impaired hyaluronan-dependent CD44/epidermal
growth factor receptor signaling. Am J Pathol. 176:1215–1228. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang Y, Chen H, Wang W, Wang R, Liu ZL,
Zhu W and Lian S: N-terminal5-merpeptideanalog P165 of amyloid
precursor protein inhibits UVA-induced MMP-1expression by
suppressing the MAPK pathway in human dermal fibroblasts. Eur J
Pharmacol. 734:1–8. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chiang HM, Chen HC, Lin TJ, Shih IC and
Wen KC: Michelia alba extract attenuates UVB-induced expression of
matrix metalloproteinases via MAP kinase pathway in human dermal
fibroblasts. Food Chem Toxicol. 50:4260–4269. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hensley K and Floyd RA: Reactive oxygen
species and protein oxidation in aging: A look back, a look ahead.
Arch Biochem Biophys. 397:377–383. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kretova M, Sabova L, Hodny Z, Bartek J,
Kollarovic G, Nelson BD, Hubackova S and Luciakova K:
TGF-β/NF1/Smad4-mediated suppression of ANT2 contributes
tooxidative stress in cellular senescence. Cell Signal.
26:2903–2911. 2014. View Article : Google Scholar : PubMed/NCBI
|