Introduction
Mesenchymal stem cells (MSCs) are a type of adult
stem cell, distinguished by their characteristics of
self-replication, low immunogenicity and easy isolation (1–3).
Previous studies have demonstrated that MSCs have the potential to
differentiate into a number of different cell lineages under
suitable conditions, including osteocytes (4), male germ-like cells (5) and diverse neuronal lineages (1,6,7). Due to their distinctive plasticity,
MSCs have been widely applied in tissue engineering to regenerate
cells or tissues, and to repair damaged tissues. For instance, the
intravenous administration of green fluorescent protein-labeled
MSCs in vivo significantly improved neurological function in
a rat stroke model via differentiation into neuron- and glial-like
cells (8,9). However, the therapeutic potential of
MSCs is impeded by their poor survival rate following
transplantation into lesion regions, where they are inevitably
subjected to harsh microenvironments, including hypoxia,
oligotrophy, peroxidation and inflammation (10). These environments eventually lead to
a reduction or depletion of endogenous peroxidases and antioxidants
(11). As a result, there is an
abnormal increase in free radicals that cannot be scavenged, which
induces irreversible cell apoptosis (11). Previous studies have demonstrated
that the majority of transplanted MSCs die within a short time
period and that the survival rate of human MSCs is <0.44% at 4
days following transplantation into immunodeficient mice (12,13).
Furthermore, cellular damage induced by continual hypoxia weakens
the regenerative capacity of MSCs (14). For instance, Deschepper et al
(15) revealed that exposure to
severe hypoxia (1% O2) for 12 consecutive days led to
the widespread death of MSCs; the number of viable MSCs was
markedly reduced when MSCs were cultured in an environment
containing 1% O2 compared with 21% O2
(2.5±1.2×104 vs. 16±3×104 cells). In
addition, Zhu et al (10)
demonstrated that hypoxia and serum deprivation (H/SD) treatment
significantly upregulated the expression of caspase-3 in MSCs,
which subsequently resulted in the initiation of the
caspase-dependent apoptosis signaling pathway. These results
demonstrate that serious abiotic stresses, such as H/SD, lead to
MSC apoptosis. Therefore, if the tolerance of MSCs to adverse
physiological and biochemical factors could be enhanced under
hypoxic and ischemic conditions, this would be of great
significance to their clinical application for the repair or
regeneration of damaged tissues in vivo.
Currently, a number of researchers are aiming to
develop reliable strategies that increase the survival of MSCs
under adverse conditions in vitro. For instance, Lv et
al (16) demonstrated that
hypoxia-inducible factor (HIF)-α overexpression effectively
promoted the viability of MSCs and suppressed apoptosis under
hypoxic conditions. Chinese scholars have demonstrated that Chinese
herbs and their active ingredients, including salidroside (17) and hydroxysafflor yellow A (HSYA)
(18), may protect stem cells
against apoptosis induced by toxic chemicals, including Cytarabine
and β-mercaptoethanol. HSYA is a flavonoid extracted from the herb,
Carthamus tinctorius L., which has been demonstrated to
exhibit potent antioxidative effects on neurons (19) and endothelial cells (20) in vitro. To date, it has been
extensively applied in the treatment of traumatic brain injury
(21) and cardiac-cerebral vascular
disease (22). A previous study
demonstrated that the protective effects of HSYA against brain
injury in a stroke animal model was achieved by attenuating the
elevation of malondialdehyde, decreasing glutathione content and
increasing superoxide dismutase activity (23).
To enhance the survival of MSCs in disease regions
and promote the clinical application of these cells in tissue
repair and regeneration, the authors of present study hypothesized
that HSYA may protect MSCs against H/SD-induced apoptosis, thus
improving the survival rate of transplanted MSCs. To investigate
this hypothesis, H/SD treatment was utilized to mimic the
unfavorable microenvironment of ischemic injury and the
anti-apoptotic effects of HSYA on MSCs under H/SD conditions, and
the associated signaling pathways were analyzed.
Materials and methods
MSC culture
A total of 6 male Sprague Dawley rats (3 weeks old;
45–50 g) were purchased from the Academy of Military Medical
Science of China (Beijing, China), raised under the conditions of
20–25°C, 60±10% humidity and a 12:12-h light/dark cycle, and fed
standard diet and provided with filtered water ad libitum.
All animal experiments were approved by the Animal Ethics Committee
of Hebei North University (Hebei, China; reference number:
2016-1-9-05). The isolation of MSCs from the 6 rats was performed
in accordance with the National Institutes of Health Guide for the
Care and Use of Laboratory Animals. Rats were anesthetized with an
intraperitoneal injection of 400 mg/kg chloral hydrate before the
rats were sacrificed and their femurs were dismembered. The bone
marrow MSCs were then harvested from the femurs using complete
Dulbecco's modified Eagle's medium containing 1 g/l glutamine
(L-DMEM; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
plus 10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific,
Inc.) and seeded into 25 cm2 culture flasks. The cells
were maintained at 37°C in a humidified atmosphere with 95% air and
5% CO2. When confluence had reached ~90%, cells were
harvested with trypsin and subcultured until the third or fourth
generation.
In order to observe the morphology of MSCs,
Diff-Quick kit (Beijing Propbs Biotechnoloty Co., Ltd., Beijing,
China) was applied to stain MSCs for three generations. In brief,
MSCs were seeded in 24-well plate at a density of 1×104
and cultured under the aforementioned conditions. When the
confluence reached ~90%, MSCs were washed three times using PBS
with the pH at 7.2. Subsequently, at room temperature MSCs were
fixed with Reagent A for 30 sec and stained with Reagent B for 30
sec. Then MSCs were observed under a multifunctional fluorescence
microscope (Eclipse 90i; Nikon Corporation, Tokyo, Japan;
magnification, ×200) and images were captured.
Cell authentication
To confirm the mesenchymal properties of the cells
cultured in the present study, cell-surface antigen profiles were
determined by flow cytometry (FCM) analysis (BD Biosciences,
Franklin Lakes, NJ, USA). MSCs express cluster of differentiation
(CD)13, CD44, CD73, CD90 and CD105, but are negative for the
expression of CD45, CD34, CD14 and CD19 (24). Therefore, third generation MSCs
(1×106 cells/ml) obtained in the present study were
incubated with the following antibodies (10 µl): Phycoerythrin
(PE)-CD29 (2) (dilution, 1:1,000;
cat. no. 562154; BD Pharmingen; BD Biosciences), allophycocyanin
(APC)-CD90 (dilution, 1:1,000; cat. no. 561409; BD Pharmingen; BD
Biosciences), fluorescein isothiocyanate (FITC)-CD34 (dilution,
1:1,000; cat. no. ab192547; Abcam, Cambridge, UK) and FITC-CD45
(dilution, 1:1,000; cat. no. 561867; BD Pharmingen; BD Biosciences)
for 30 min at room temperature in the dark. This was followed by
washing with PBS, and fluorescence was detected by FCM equipped
with FACSDiva analytical software (version 8.0.1; BD
Bioscience).
Cell treatments
In order to assess whether HSYA protects MSCs
against H/SD-induced injury, H/SD treatment and HSYA protection
were conducted on fourth generation MSCs. MSCs were first divided
into normal, H/SD and protective groups. In the normal group, MSCs
were cultured under the aforementioned conditions and regarded as
the negative control. In the H/SD group, MSCs were cultured in
L-DMEM (FBS-free) under hypoxic conditions (5% O2)
(25) for 6–48 h. In the protective
group, MSCs were cultured for 6–48 h using L-DMEM (FBS-free)
containing 160 mg/l HSYA (18) under
hypoxic conditions.
Determination of cell apoptosis and
reactive oxygen species (ROS) levels
Following the exposure of cells in each group to
different culture conditions for 48 h at room temperature, double
fluorescence staining using 0.5 µg/ml Hoechst 33342 and 50 µg/ml
propidium iodide (PI) was employed to stain the nuclei for 10 min
at room temperature to determine cell apoptosis. Hoechst
33342-positive cells are viable and PI-positive cells are
apoptotic. Using a multifunctional fluorescence microscope (Nikon
90-I; Nikon Corporation, Tokyo, Japan), the percentage of apoptotic
cells in each group was determined by counting Hoechst 33342- and
PI-positive cells in 10 random fields of view.
The ROS level in MSCs was measured using an ROS
assay kit (Beyotime Institute of Biotechnology, Haimen, China) in
accordance with the manufacturer's protocol. After the exposure of
MSCs in three groups to different culture conditions for 6, 12, 24
and 48 h, MSCs were incubated with 2′,7′-dichlorodihydrofluorescein
diacetate (DCF; Xiamen Bioluminor Bio-Tech Co., Ltd., Xiamen,
China) for 30 min at 37°C. Following washing with PBS three times,
DCF fluorescence intensity was measured using FCM equipped with
FACSDiva analytical software. Hoechst 33342 (0.5 µg/ml) was used to
stain the nuclei after 24 h of treatment; Hoechst 33342 was
incubated with the MSCs for 10 min at room temperature and images
were obtained under a fluorescence microscope (magnification,
×200).
Cell viability and proliferation
assay
Cell viability was assessed using an MTT assay kit
(Shanghai Gefan Biotechnology Co., Ltd., Shanghai, China). Briefly,
fourth generation MSCs (8×103 cells/ml) were seeded into
96-well plates. Once the cells had adhered to the culture vessel,
they were divided into the aforementioned groups and exposed to the
associated conditions for 6, 12, 24 and 48 h. MTT reagent (5 mg/ml)
was then added and the cells were incubated for 4 h at 37°C.
Ice-cold PBS was used to wash the cells, and 150 µl DMSO was then
added. The absorbance of each well was measured at 570 nm using a
microplate reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Each cell treatment was assayed >3 times.
For the determination of cell proliferation, a
5-ethynyl-2′-deoxyuridine (EdU) kit (KeyGen Biotech, Co., Ltd.,
Nanjing, China,) was used. Briefly, 0.6 ml/well MSCs at a density
of 5×104 cells/ml were seeded in 6-well plates and when
the MSCs had adhered to the surface of the culture vessel, MSCs
were exposed to their respective culture conditions. EdU (20 µM)
was then added and the cells were incubated for 24 h at 37°C.
Thereafter, at room temperature the cells were washed with PBS,
fixed in 4% polyformaldehyde for 10 min at room temperature and
stained with kFluor488-azide [dilution, 3:1,000 (v/v); KeyGen
Biotech, Co., Ltd.] for 30 min at room temperature. The cells were
then stained with 0.5 µg/ml Hoechst for 10 min at room temperature.
Images were acquired using a multifunctional fluorescence
microscope. Three fields were randomly selected from each dish and
at least three dishes from each group were counted to determine the
number of EdU-positive cells.
Western blotting
Considering that HIF-1/vascular endothelial growth
factor (VEGF) signaling contributes to protection against ischemic
brain injury (26) and that
caspase-3 and cytochrome c (cytC) promote cell apoptosis (27), a western blot analysis was performed
to measure the expression of these proteins. MSCs (0.6 ml/well)
were seeded in 6-well plates at density of 5×104
cells/ml. When the cells were attached to the surface of culture
plates, MSCs were exposed to the different treatment conditions for
24 h. Total protein was extracted from MSCs on ice using RIPA Lysis
buffer (Applygen Biotech, Co., Ltd., Beijing, China). Meanwhile, to
detect the content of cytC in the cytoplasm, cytoplasmic protein
was extracted from abovementioned MSCs using nuclear/cytoplasmic
protein extraction kit (Sangon Biotech Co., Ltd., Shanghai, China).
MSCs were harvested and treated with Buffer A for 15 min at room
temperature, Buffer B was added for 1 min on ice and vortexed for
15 sec. The cells were then kept on ice for 1 min and centrifuged
at 14,000 × g for 5 min at 4°C. The resulting supernatant only
contained cytoplasmic protein. The protein concentration was
determined using a bicinchoninic acid assay kit (BestBio Company,
Shanghai, China). Protein (25 µg) from each sample was mixed with
loading buffer and resolved by 11% SDS-PAGE.
Following electrophoresis, protein bands were
electrotransferred onto polyvinylidene fluoride (PVDF) membranes.
The membranes were blocked using 10% non-fat milk at room
temperature for 1 h to prevent non-specific binding. The membranes
were subsequently incubated with the following rabbit anti-rat
antibodies obtained from Bioss Antibodies Inc. (Beijing, China):
HIF-1α (dilution, 1:2,000; cat. no. bs-0737R), VEGF (dilution,
1:2,000; cat. no. bs-1665R), cleaved caspase-3 (dilution, 1:2,000;
cat. no. bs-0081R), cytC (dilution, 1:2,000; cat. no. bs-0013R) and
β-actin antibody (dilution, 1:2,000; cat. no. bs-0061R) at 37°C for
2 h. Following washing with Tris-buffered saline and 0.1% Tween 20,
the membranes were incubated with horseradish peroxidase-conjugated
goat anti-rabbit secondary antibody (dilution, 1:3,000; cat. no.
bs-0295G; Bioss Antibodies Inc.) for 1 h at room temperature. An
enhanced chemiluminescence detection reagent (Beyotime Institute of
Biotechnology) was incubated with the PVDF membranes for 3–5 min at
room temperature in the dark. The protein bands were visualized
using the Aplegen Omega Lum W gel imaging system (Gel Company,
Inc., San Francisco, CA, USA). The average area and band densities
from 3–5 independent blots were used to represent the expression of
all proteins. All experimental groups used a β-actin antibody as
the loading control.
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR analysis)
To further assess the potential protective effect of
HSYA against H/SD-induced injury in MSCs, RT-qPCR was performed to
assess the expression of HIF-1α and VEGF following H/SD treatment.
Total RNA was extracted from MSCs using TRNzol reagent (Tiangen
Biotech Co., Ltd., Beijing, China) in accordance with the
manufacturer's protocol. The purity of RNA was determined using
spectrophotometry (wavelength range, 190–1,100 nm). The quality of
extracted RNA was considered acceptable when the optical density
(OD) at 260/280 nm was between 1.9 and 2.0. Total RNA (2 µg) was
then reverse transcribed into cDNA using the First-Strand cDNA
Synthesis kit (Tiangen Biotech Co., Ltd.) containing oligo dT18
primers. The specific primers for RT-qPCR were designed using the
online Primer-Blast tool (www.ncbi.nlm.nih.gov/tools/primer-blast/).
Amplification was performed using the Premix Taq PCR kit (Takara
Biotechnology Co., Ltd., Dalian, China), and the following
thermocycling conditions: An initial denaturation step at 94°C for
5 min, followed by 30 cycles of 94°C for 40 sec, 55°C for 40 sec
and 72°C for 2 min, and then one cycle at 72°C for 10 min. The
primers, amplification fragment lengths and annealing temperatures
are presented in Table I. β-actin
was used as a control housekeeping gene. The relative expression
level of target genes was plotted as a fold change compared with
the control using the 2−ΔΔCq method (28).
 | Table I.Reverse transcription-quantitative
polymerase chain reaction primer sequences. |
Table I.
Reverse transcription-quantitative
polymerase chain reaction primer sequences.
| Genes | Primers
(5′-3′) | Tm (°C) | Fragment length
(bp) |
|---|
| HIF-1α |
| 55 | 218 |
|
Sense |
AACAAACAGAATCTGTCCTCAAACC |
|
|
|
Antisense |
CAGGTAATGGAGACATTGCCAG |
|
|
| VEGF |
| 55 | 151 |
|
Sense |
CAGCGACAAGGCAGACTATT |
|
|
|
Antisense |
GTTGGCACGATTTAAGAGGG |
|
|
| β-actin |
| 55 | 246 |
|
Sense |
TCACCCACACTGTGCCCATCTATGA |
|
|
|
Antisense |
CATCGGAACCGCTCATTGCCGATAG |
|
|
Statistical analysis
All data were analyzed using SPSS 17.0 software
(SPSS, Inc., Chicago, IL, USA) and expressed as the mean ± standard
deviation. Homoscedasticity of primary data was first detected
using Levene's test, and variance was demonstrated to be
homogeneous. Comparisons among multiple groups were analyzed by
two-way analysis of variance. Pairwise comparisons between
different groups were performed using a Tukey's honest significance
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Morphology and surface-antigen profile
determination of MSCs
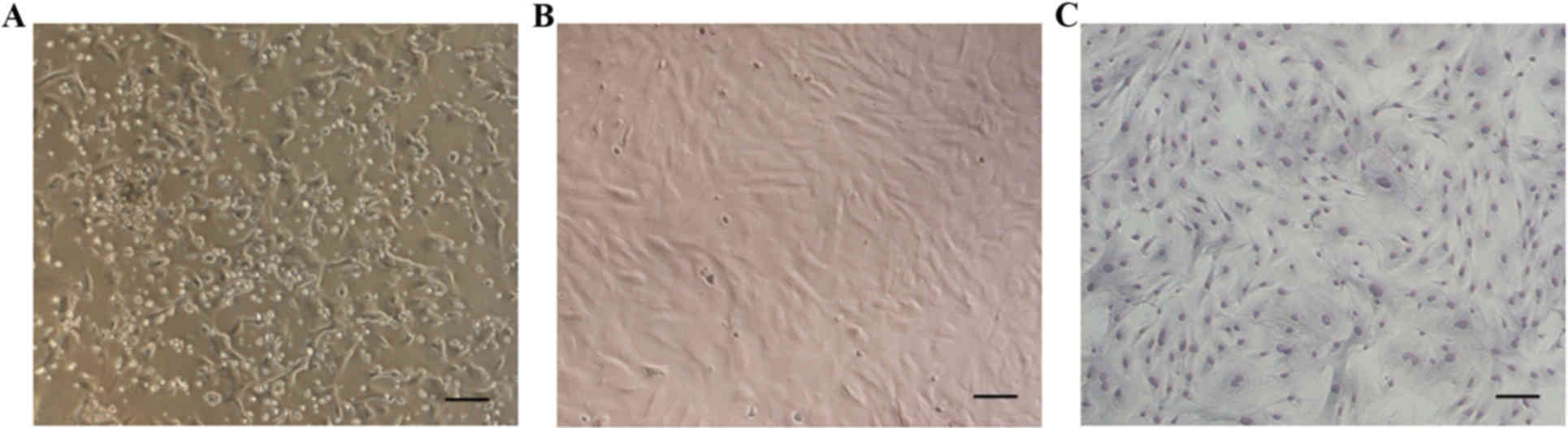
Following 72 h of primary culture, the majority of
adherent cells exhibited an elongated shape (Fig. 1A). Non-adherent cells were gradually
removed by changing the medium. Following 7–8 days of culture,
adherent cells reached 80–90% confluence. Cells were then harvested
with trypsin and subcultured until the third generation. At this
stage, the MSCs exhibited a fibroblast- or spindle-like morphology
when they reached 80–90% confluence (Fig. 1B and C).
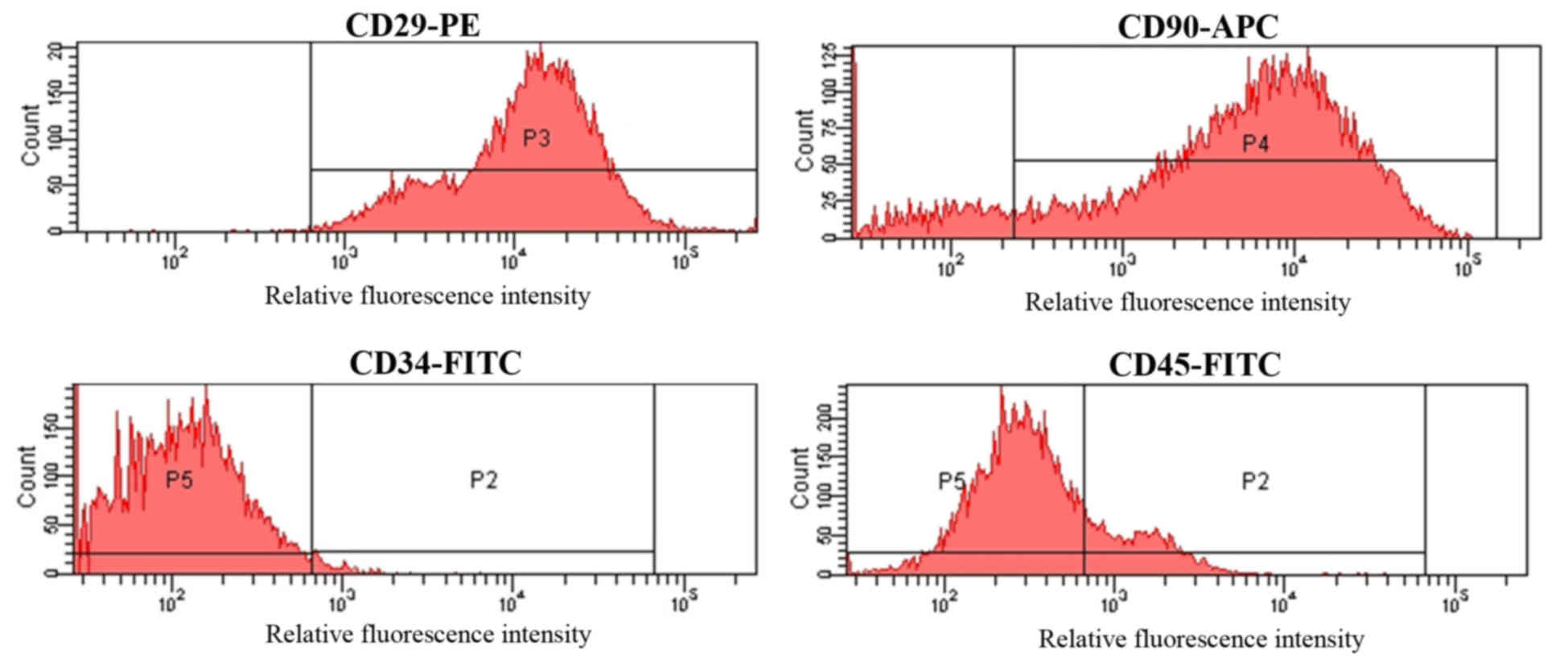
Third generation MSCs were incubated with
FITC-conjugated antibodies targeting CD34 and CD45, a PE-conjugated
antibody against CD29 and an APC-conjugated antibody targeting
CD90. The cell surface-antigen profiles of MSCs were then
determined by FCM. As demonstrated in Fig. 2, MSCs were strongly positive for CD29
and CD90, with positive rates of 97.05±2.38 and 87.13±2.47%,
respectively. By contrast, MSCs were negative for CD34 and CD45,
which were present in 1.94±0.27 and 12.61±2.29% of the cell
population, respectively (Fig.
2).
Detection of cell apoptosis and ROS
levels
Staining with Hoechst 33342 and PI was used to
detect the level of cell apoptosis. Compared with the normal group,
a significant and time-dependent increase in the proportion of MSCs
was observed following H/SD treatment, and the proportion of
apoptotic cells was >35% in the H/SD group following 48 h of
treatment (Fig. 3A and B). By
contrast, the percentage of apoptotic cells was ~15% in the
protective group, which was significantly lower than the H/SD
group. This indicated that HSYA treatment reduced the level of
apoptosis.
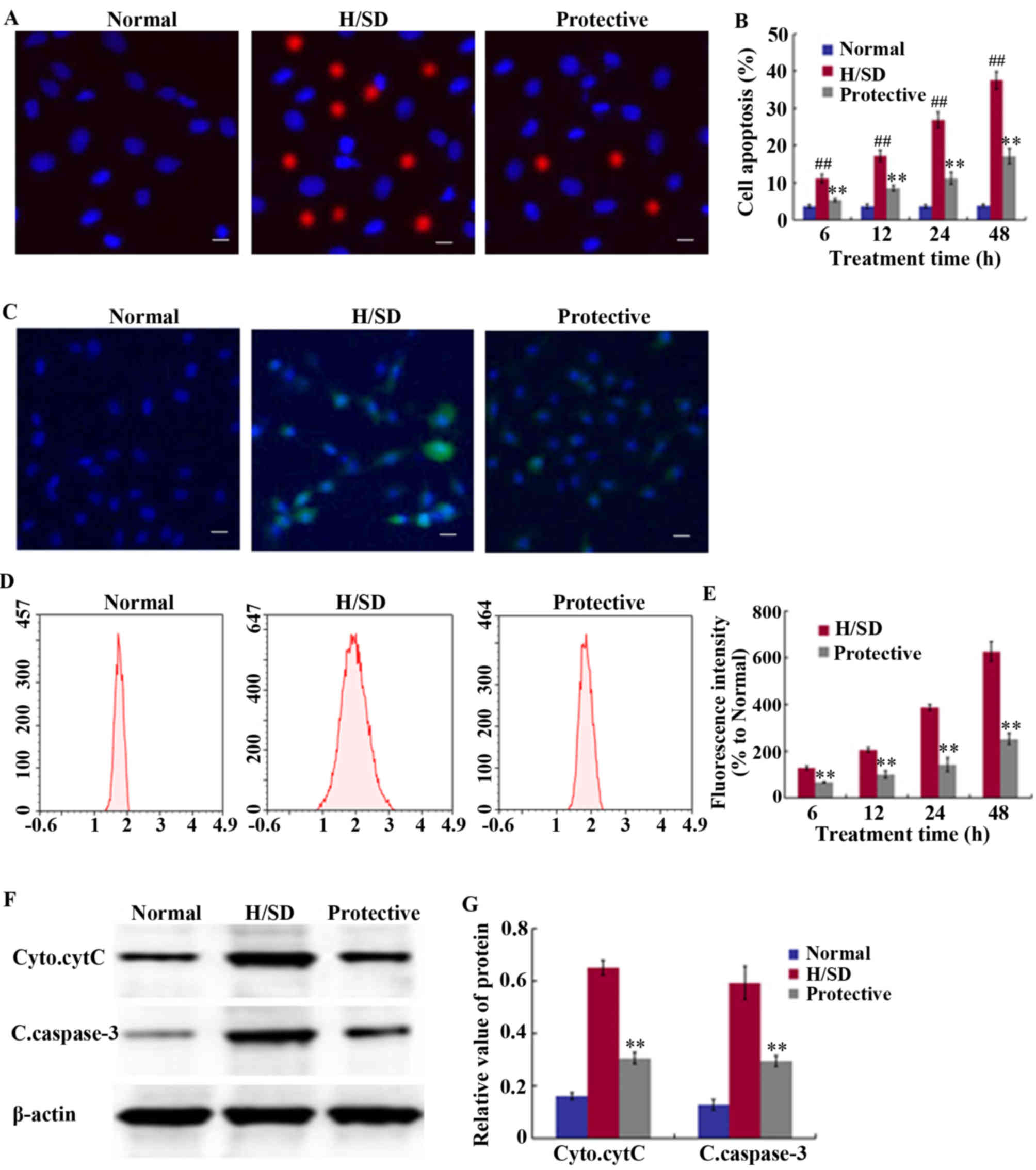 | Figure 3.Effect of HSYA on apoptosis levels
and ROS generation in MSCs. (A) Determination of cell apoptosis
through fluorescence staining procedures following exposure to H/SD
treatment for 48 h (scale bar, 50 µm). (B) Quantitative MSC
apoptosis levels at 6, 12, 24 and 48 h. (C) Determination of ROS
accumulation through fluorescence staining following exposure to
H/SD treatment for 48 h (scale bar, 50 µm), and (D) determination
of DCF fluorescence intensity by FCM expressed relative to the
normal control group following 6–48 h of H/SD treatment. (E)
Quantitative ROS levels at 6, 12, 24 and 48 h. (F) Western blotting
analysis of cytC and caspase-3 following 24 h of H/SD culture. (G)
Quantification of relative band intensities. ##P<0.01
vs. the normal. **P<0.01 vs. the H/SD group. HSYA,
hydroxysafflor yellow A; ROS, reactive oxygen species; MSCs,
mesenchymal stem cells; H/SD, hypoxia and serum deprivation; FCM,
flow cytometry; DCF, 2′,7′-dichlorodihydrofluorescein diacetate;
Cyto.cytC, cytoplasmic cytochrome C; C.caspsase-3, cleaved
caspase-3. |
An anoxic environment induces ROS production, which
is one of the primary factors leading to cell apoptosis and
senescence (11). As indicated in
Fig. 3C-E, MSCs exposed to H/SD
conditions exhibited a notable increase in ROS levels. The FCM
results revealed that at 48 h following H/SD culture, a marked
increase in DCF fluorescence intensity was observed, which was
~6.5-fold higher than the normal control group and ~3-fold higher
than the protective group (Fig.
3C-E). As presented in Fig. 3E,
fluorescence intensity at 48 h in the protective group was
221.33±20.21% and in the H/SD group it was 655.67±20.74%.
To further elucidate the mechanisms underlying the
anti-apoptotic effects exerted by HSYA, western blot analysis was
performed to assess the expression levels of cleaved caspase-3 and
cytoplasmic cytC. As presented in Fig.
3F and G, HSYA significantly suppressed the release of cytC
from the mitochondria, as well as the expression of cleaved
caspase-3 under H/SD conditions. The relative band density of
cytoplasmic cytC was 0.31±0.02 in the protective group and
0.65±0.03 in the H/SD group, while the level of cleaved caspase-3
was 0.29±0.02 in the protective group and 0.59±0.06 in the H/SD
group.
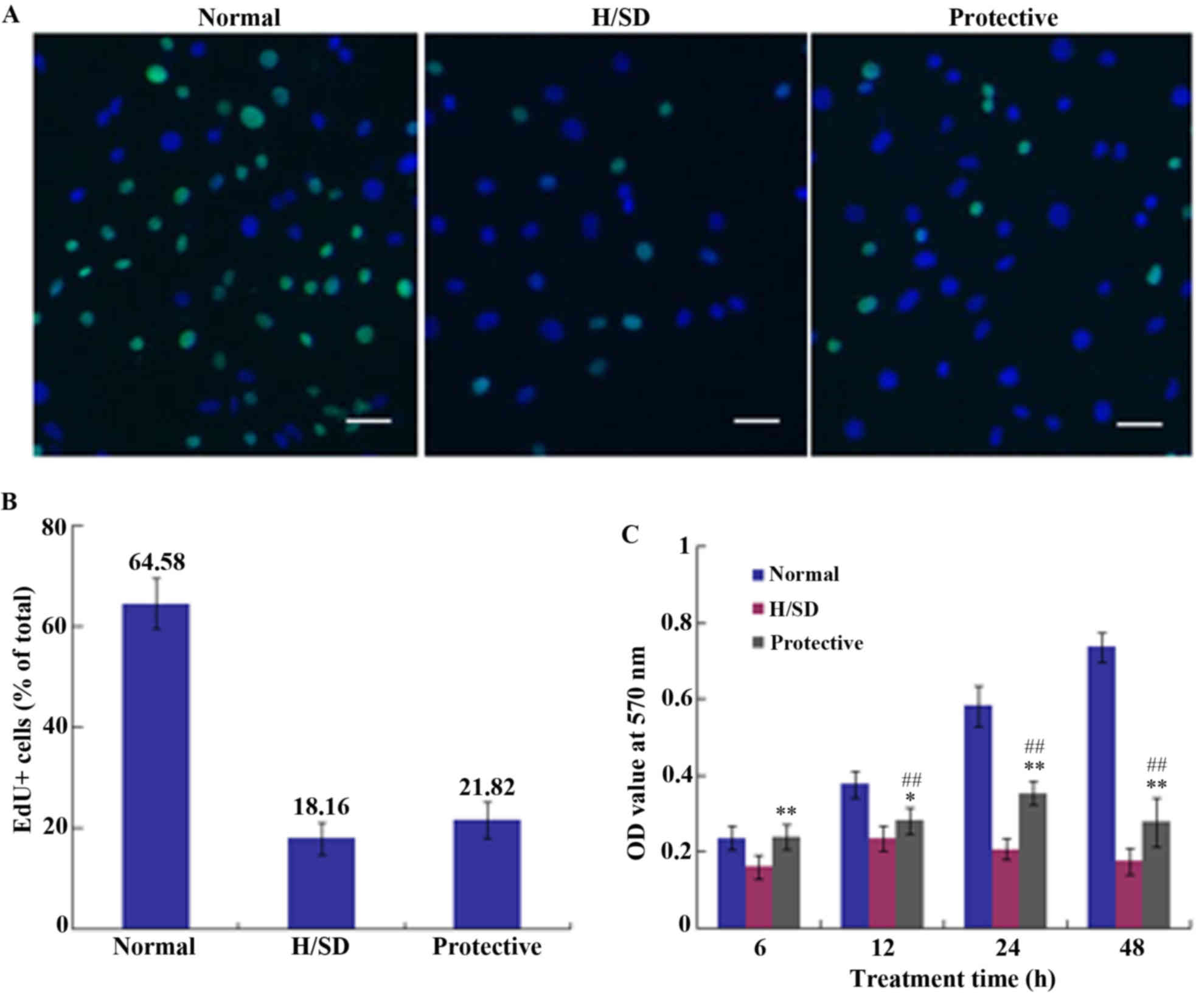
Cell proliferation and viability
assays
To determine cell proliferation, medium containing
an EdU/kFluor488-azide was applied to cell cultures for 24 h. The
percentage of EdU-positive cells in normal, H/SD and protective
groups was 64.58±5.02, 18.16±3.19 and 21.82±3.53%, respectively
(Fig. 4A and B). Thus, there was no
marked difference between the H/SD and protective groups, which
indicated that HSYA exhibited no notable effect on MSC
proliferation under H/SD conditions.
An MTT assay was used to assess cell viability and
the OD value was determined at 570 nm using a microplate reader. As
demonstrated in Fig. 4C, cell
viability in the protective HSYA group was significantly higher
when compared with the H/SD group; however, it was reduced when
compared with the normal group. Continuous H/SD treatment for 48 h
led to a significant decrease in cell viability in the protective
group compared with the normal group. As presented in Fig. 4C, cell viability in the protective
group was <40% of that in the normal group following 48 h of
treatment. Therefore, compared with the H/SD group, HSYA protection
exerted a positive effect on cell viability following a short
period of exposure, which may be associated with ROS scavenging.
The decrease of ROS stabilizes mitochondrial membrane integrity,
which contributes to the maintenance of cell viability.
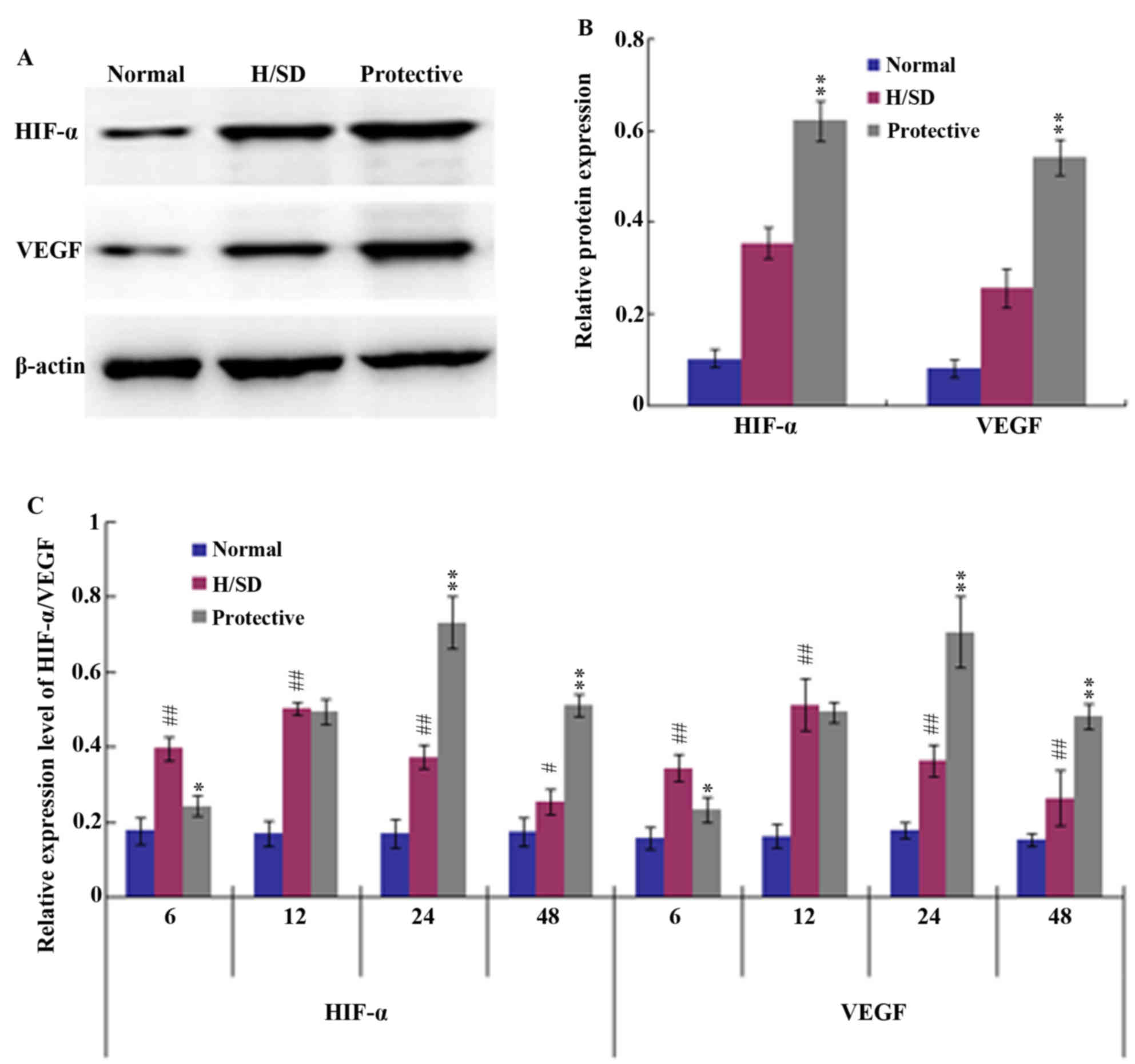
HIF-1/VEGF signaling pathway
To further investigate the anti-apoptotic effect of
HSYA on MSCs under H/SD conditions, the potential underlying
mechanisms were assessed. HIF-1 and its target gene VEGF are known
to be involved in a number of biological functions, such as cell
proliferation and anti-apoptotic mechanisms (29). Therefore, western blot and RT-qPCR
analyses were employed in the present study to determine whether
HSYA activates the HIF-1/VEGF signaling pathway. Western blotting
indicated that the expression levels of HIF-1α and VEGF in the
protective group were significantly higher than those in the H/SD
group following 24 h of H/SD treatment (Fig. 5A and B). Consistent with these
observations, the RT-qPCR results demonstrated that 6–12 h of H/SD
treatment upregulated the expression of HIF-1α and VEGF in the H/SD
group when compared with the normal group (Fig. 5C). H/SD exposure and HSYA protection
for 24 h led to a peak in HIF-1α and VEGF expression, after which
their mRNA levels began to decrease. The decline in HIF-1α and VEGF
expression levels may have been associated with a decrease in cell
viability.
Discussion
Due to their capacity for self-renewal and
multipotency, MSCs have received attention for their use in
developing treatments for central nervous system diseases,
including cerebral ischemia (30),
stroke (31) and spinal cord injury
(32). MSCs have also been
demonstrated to provide a suitable microenvironment for injured and
damaged tissues through the secretion of cytokines and trophic
factors (33), which serve important
roles in the protection of injured neurons. However, MSCs
transplanted into lesion sites are exposed to hypoxic and/or
ischemic stress; thus, their curative potential is limited.
Therefore, it is important to explore novel strategies for
improving the survival rate of MSCs in vivo. Previous
studies have demonstrated that the short-term exposure of MSCs to
hypoxic conditions may promote cell proliferation and stemness,
thus enriching the pool of cells potentially able to differentiate
into multiple lineages (34–36). However, oxygen-derived free radicals
induced by hypoxia are generally considered to be important
contributors to MSC apoptosis (37).
Recent studies have revealed that HSYA attenuates the breakdown of
the blood brain barrier (26) and
alleviates the imbalance between antioxidants and oxidants, leading
to neuroprotective effects against ischemia-reperfusion injury in
the spinal cord (38), liver
(39) and brain (19) via the scavenging of free
radicals.
In light of the neuroprotective effects conferred on
cells or organs against oxidative damage, HSYA was applied to
protect MSCs against H/SD-induced injury in the present study. The
results demonstrated that when compared with the normal group, the
apoptosis rate of MSCs exposed to H/SD significantly increased and
~40% of MSCs died following 48 h of H/SD treatment. By contrast,
prior protection with 160 mg/l HSYA notably increased the survival
rate of MSCs under H/SD conditions and the apoptosis level in the
protective group was ~15% at the same time point.
As a by-product of oxidative phosphorylation, a
moderate quantity of ROS is necessary for cell survival,
proliferation and promotion of longevity (40). However, in pathological sites,
hypoxia-induced ischemia leads to an imbalance between the
formation and scavenging of free radicals, which results in excess
ROS (41). ROS accumulation leads to
the release of the pro-apoptotic protein cytC into the cytoplasm
from mitochondria and initiates the mitochondrial apoptotic pathway
(42). Once cytC is released into
the cytoplasm, it activates the expression of the downstream
caspase apoptotic family (43). In
this superfamily, cleaved caspase-3, a key mediator of apoptosis,
performs the terminal steps of cell apoptosis and regulates the
upstream induction of cell death (44). The results of the current study
revealed that H/SD induced the production of excess ROS and that
this effect was suppressed by HSYA treatment. Western blot analysis
further confirmed the anti-apoptotic effect of HSYA and revealed
that HSYA notably suppressed the release of cytC into the
cytoplasm, reducing the caspase cascade reaction.
It has been suggested that the ROS scavenging action
of HSYA may contribute to the maintenance and integrity of the
mitochondrial membrane and inhibition of the mitochondrial
apoptosis signaling pathway (22).
The pre-treatment with medicines is an effective measure for
decreasing MSC apoptosis under malignant conditions in vitro
(17,45). For instance, the application of
Nicorandil (45) significantly
protects MSCs against apoptosis induced by H/SD for 9 h.
Comparatively, the results of the present study demonstrated that
the anti-apoptotic effects of HSYA were more considerable, as the
apoptosis rate of MSCs in the protective group was <50% of the
H/SD group.
Further experiments were performed to elucidate the
molecular mechanisms associated with the regulation of apoptosis by
HSYA. Specifically, the involvement of the HIF-1/VEGF signaling
pathway in the anti-apoptotic activity of HSYA in MSCs was
assessed. The HIF-1/VEGF pathway serves a vital role in promoting
the survival of various cell types under hypoxic conditions
(16,29,46).
HIF-1 is a heterodimeric protein composed of two subunits, HIF-1α
and HIF-1β (29). HIF-1α is a
functional subunit and belongs to the helix-loop-helix family of
transcription factors (47). Its
transcriptional activity is regulated by intracellular oxygen
concentrations (46). Under hypoxic
conditions, HIF-1α persists and HIF-1α/β is transported into the
nucleus where it binds to HIF responsive elements and activates the
expression of downstream target genes, such as erythrpoietin and
VEGF, which are involved in energy metabolism and oxygen delivery
(29). A previous study demonstrated
that HIF-1 may serve as a neurotrophic and neuroprotective factor,
which promotes MSC-mediated neurological and functional recovery in
the ischemic brain (48). It is well
established that VEGF, induced by HIF-1α, binds to its
complementary receptors to initiate ERK-1/2 signaling and then
activates the Akt pathway, resulting in cell proliferation,
anti-apoptosis, anti-inflammation and angiogenesis (29,49–51). It
has also been revealed that HIF-1α-induced VEGF expression in MSCs
exerts an essential protective anti-ischemic effect on
cardiomyocytes (52). Consistent
with these observations, Zhong et al (53) observed a significant increase in the
release of VEGF in HIF-1α-modified MSCs, which exerted a protective
effect on neuron-like cells or neurons following ischemia in
vitro and in vivo through the elevated expression of
survivin: A member of the inhibitors of apoptosis protein family.
Furthermore, VEGF has been demonstrated to function as a
neuroprotective factor in motor neurons (54). For instance, VEGF released by MSCs
reduced neuronal apoptosis and promoted neuronal proliferation in a
rat stroke model (9). However,
although HSYA protection was observed to significantly elevate the
expression of HIF-1α and VEGF under H/SD conditions in the present
study, the proliferative ability of MSCs in the protective group
was not enhanced, which may be associated with the lack of growth
factors under serum deprivation. However, in terms of ROS
scavenging activity, HSYA protection enhanced cell viability during
short-term H/SD treatment.
In conclusion, the results of the present study
indicated that HSYA improved the survival rate of MSCs under H/SD
conditions, which mimic the microenvironment of ischemic diseases.
Under these conditions, the pro-survival and anti-apoptotic effects
of HSYA may depend on the activation of the HIF-1/VEGF signaling
pathway, the reduction of ROS production and the maintenance of
mitochondrial membrane integrity.
Acknowledgements
The authors would like to thank all teachers of the
Morphology Key Lab of Hebei North University.
Funding
The present study was funded by the Major Projection
of Hebei North University (grant no. ZD201413).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XS, HY and HW mainly participated in the study
design, writing and critical revision. XS, LS and JD mainly
performed the experiments, and collected and analysed the data. All
authors have read and approved this manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Ethics Committee of Hebei North University (Hebei, China; reference
nο.: 2016-1-9-05).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kim EY, Lee KB, Yu J, Lee JH, Kim KJ, Han
KW, Park KS, Lee DS and Kim MK: Neuronal cell differentiation of
mesenchymal stem cells originating from canine amniotic fluid. Hum
Cell. 27:51–58. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jin W, Xu YP, Yang AH and Xing YQ: In
vitro, induction and differentiation of umbilical cord mesenchymal
stem cells into neuron-like cells by all-trans retinoic acid. Int J
Ophthalmol. 8:250–256. 2015.PubMed/NCBI
|
|
3
|
Han SM, Han SH, Coh YR, Jang G, Chan-Ra J,
Kang SK, Lee HW and Youn HY: Enhanced proliferation and
differentiation of Oct4- and Sox2-overexpressing human adipose
tissue mesenchymal stem cells. Exp Mol Med. 46:e1012014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Heino TJ and Hentunen TA: Differentiation
of osteoblasts and osteocytes from mesenchymal stem cells. Curr
Stem cell Res Ther. 3:131–145. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nejad NA, Amidi F, Hoseini MA, Nia KN,
Habibi M, Kajbafzadeh AM, Mazaheri Z and Yamini N: Male germ-like
cell differentiation potential of human umbilical cord Wharton's
jelly-derived mesenchymal stem cells in co-culture with human
placenta cells in presence of BMP4 and retinoic acid. Iran J Basic
Med Sci. 18:325–333. 2015.PubMed/NCBI
|
|
6
|
Mehranjani Soleimani M and Chian MF:
Cysteine: A novel neural inducer for rat bone marrow mesenchymal
stem cells. Cell J. 16:195–202. 2014.PubMed/NCBI
|
|
7
|
Takeda YS and Xu Q: Neuronal
differentiation of human mesenchymal stem cells using exosomes
derived from differentiating neuronal cells. PLoS One.
10:e01351112015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu J, Sun Z, Sun HS, Weisel RD, Keating A,
Li ZH, Feng ZP and Li RK: Intravenously administered bone marrow
cells migrate to damaged brain tissue and improve neural function
in ischemic rats. Cell Transplant. 16:993–1005. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Deng YB, Ye WB, Hu ZZ, Yan Y, Wang Y,
Takon BF, Zhou GQ and Zhou YF: Intravenously administered BMSCs
reduce neuronal apoptosis and promote neuronal proliferation
through the release of VEGF after stroke in rats. Neurol Res.
32:148–156. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhu W, Chen J, Cong X, Hu S and Chen X:
Hypoxia and serum deprivation-induced apoptosis in mesenchymal stem
cells. Stem Cells. 24:416–425. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nigam S and Schewe T: Phospholipase A(2)s
and lipid peroxidation. Biochim Biophys Acta. 1488:167–181. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tang YL, Tang Y, Zhang YC, Qian K, Shen L
and Phillips MI: Improved graft mesenchymal stem cell survival in
ischemic heart with a hypoxia-regulated heme oxygenase-1 vector. J
Am Coll Cardiol. 46:1339–1350. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Toma C, Pittenger MF, Cahill KS, Byrne BJ
and Kessler PD: Human mesenchymal stem cells differentiate to a
cardiomyocyte phenotype in the adult murine heart. Circulation.
105:93–98. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Buravkova LB, Andreeva ER, Gogvadze V and
Zhivotovsky B: Mesenchymal stem cells and hypoxia: Where are we?
Mitochondrion. 19:105–112. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Deschepper M, Oudina K, David B, Myrtil C,
Collet C, Bensidhoum M, Logeart-Avramogiou D and Petite H: Survival
and function of mesenchymal stem cells (MSCs) depend on glucose to
overcome exposure to long-term, severe and continuous hypoxia. J
Cell Mol Med. 15:1505–1514. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lv B, Li F, Fang J, Xu L, Sun C, Han J,
Hua T, Zhang Z, Feng Z and Jiang X: Hypoxia inducible factor 1α
promotes survival of mesenchymal stem cells under hypoxia. Am J
Transl Res. 9:1521–1529. 2017.PubMed/NCBI
|
|
17
|
Wei YP, Bai H, Sun YQ, Bao S, Qian R and
Liu L: Effects of salidroside on apoptosis of bone marrow
mesenchymal stem cells induced by Ara-C. Zhongguo Shi Yan Xue Ye
Xue Za Zhi. 21:1572–1577. 2013.(In Chinese). PubMed/NCBI
|
|
18
|
Song XQ, Su LN, Wei HP, Liu YH and Yin HF:
Protective effects of hydroxysafflor yellow a against oxidative
damage of β-mercaptoethanol during neural differentiation of
mesenchymal stem cells. Chin Herbal Med. 9:282–288. 2017.
View Article : Google Scholar
|
|
19
|
Wang X, Ma Z, Fu Z, Gao S, Yang L, Jin Y,
Sun H, Wang C, Fan W, Chen L, et al: Hydroxysafflor yellow A
protects neurons from excitotoxic death through inhibition of
NMDARs. ASN Neuro. 8:pii. 2016. View Article : Google Scholar
|
|
20
|
Zhang L, Song Y, Liu CL, Zhu MC, Wang H,
Liu K and Zhu HB: Mechanism of hydroxysaf f lor yellow A induced
endothelial cell proliferation. Chin Traditional Herbal Drugs.
39:403–408. 2008.
|
|
21
|
Wang Y, Zhang C, Peng W, Xia Z, Gan P,
Huang W, Shi Y and Fan R: Hydroxysafflor yellow A exerts
antioxidant effects in a rat model of traumatic brain injury. Mol
Med Rep. 14:3690–3696. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ramagiri S and Taliyan R: Neuroprotective
effect of hydroxy safflor yellow A against cerebral
Ischemi-reperfusion injury in rats: Putative role of mPTP. J Basic
Clin Physiol Pharmacol. 27:1–8. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wei X, Liu H, Sun X, Fu F, Zhang X, Wang
J, An J and Ding H: Hydroxysafflor yellow A protects rat brains
against ischemia-reperfusion injury by antioxidant action. Neurosci
Lett. 386:58–62. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Baksh D, Song L and Tuan RS: Adult
mesenchymal stem cells: Characterization, differentiation, and
application in cell and gene therapy. J Cell Mol Med. 8:301–316.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ito A, Aoyama T, Yoshizawa M, Nagai M,
Tajino J, Yamaguchi S, Lijima H, Zhang X and Kuroki H: The effects
of short-term hypoxia on human mesenchymal stem cell proliferation,
viability and p16(INK4A) mRNA expression: Investigation using a
simple hypoxic culture system with a deoxidizing agent. J Stem
Cells Regen Med. 11:25–31. 2015.PubMed/NCBI
|
|
26
|
Sun L, Yang L, Xu YW, Liang H, Han J, Zhao
RJ and Cheng Y: Neuroprotection of hydroxysafflor yellow A in the
transient focal ischemia: Inhibition of protein
oxidation/nitration, 12/15-lipoxy-genase and blood-brain barrier
disruption. Brain Res. 1473:227–235. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang X, Sun JY, Gao MN and Li Y:
Neuroprotection of overexpression of neuroglobin gene on injury of
SH-SY5Y cells and relevant mechanism. Chin J Biologicals.
26:1768–1771. 2013.
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cheng L, Yu H, Yan N, Lai K and Xiang M:
Hypoxia-inducible factor-1α target genes contribute to retinal
neuroprotection. Front Cell Neurosci. 11:202017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tian C, Wang X, Wang X, Wang L, Wang X, Wu
S and Wan Z: Autologous bone marrow mesenchymal stem cell therapy
in the subacute stage of traumatic brain injury by lumbar puncture.
Exp Clin Transplant. 11:176–181. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bhasin A, Srivastava MV, Mohanty S, Bhatia
R, Kumaran SS and Bose S: Stem cell therapy: A clinical trial of
stroke. Clin Neurol Neurosurg. 115:1003–1008. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Goni VG, Chhabra R, Gupta A, Marwaha N,
Dhillon MS, Pebam S, Gopinathan NR and Kantharajanna SB: Safety
profile, feasibility and early clinical outcome of
co-transplantation of olfactory mucosa and bone marrow stem cells
in chronic spinal cord injury patients. Asian Spine J. 8:484–490.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Madrigal M, Rao KS and Riordan NH: A
review of therapeutic effects of mesenchymal stem cell secretions
and induction of secretory modification by different culture
methods. J Transl Med. 12:2602014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fotia C, Massa A, Boriani F, Baldini N and
Granchi D: Hypoxia enhances proliferation and stemness of human
adipose-derived mesenchymal stem cells. Cytotechnology.
67:1073–1084. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Estrada JC, Albo C, Benguría A, Dopazo A,
López-Romero P, Carrera-Quintanar L, Roche E, Clemente EP, Enríquez
JA, Bernad A and Samper E: Culture of human mesenchymal stem cells
at low oxygen tension improves growth and genetic stability by
activating glycolysis. Cell Death Difer. 19:743–755. 2012.
View Article : Google Scholar
|
|
36
|
Wang Y, Yang J, Li H and Wang X, Zhu L,
Fan M and Wang X: Hypoxia promotes dopaminergic differentiation of
mesenchymal stem cells and shows benefits for transplantation in a
rat model of Parkinson's disease. PLoS One. 8:e542962013.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chan PH: Role of oxidants in ischemic
brain damage. Stroke. 27:1124–1129. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shan LQ, Ma S, Qiu XC, Zhou Y, Zhang Y,
Zheng LH, Ren PC, Wang YC, Fan QY and Ma BA: Hydroxysafflor yellow
A protects spinal cords from ischemia/reperfusion injury in
rabbits. BMC Neurosci. 11:982010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jiang S, Shi Z, Li C, Ma C, Bai X and Wang
C: Hydroxysafflor yellow A attenuates ischemia/reperfusion-induced
liver injury by suppressing macrophage activation. Int J Clin Exp
Pathol. 7:2595–2608. 2014.PubMed/NCBI
|
|
40
|
Acín-Pérez R, Carrascoso I, Baixauli F,
Roche-Molina M, Latorre-Pellicer A, Fernández-Silva P, Mittelbrunn
M, Sanchez-Madrid F, Pérez-Martos A, Lowell C, et al: ROS-triggered
phosphorylation of complex II by Fgr kinase regulates cellular
adaptation to fuel use. Cell Metab. 19:1020–1033. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gao L, Liu R, Gao F, Wang Y, Jiang X and
Gao X: Plasmon-mediated generation of reactive oxygen species from
near-infrared light excited gold nanocages for photodynamic therapy
in vitro. ACS Nano. 8:7260–7271. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dröse S, Brandt U and Wittig I:
Mitochondrial respiratory chain complexes as sources and targets of
thiol-based redox-regulation. Biochim Biophys Acta. 1844:1344–1354.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang HY, Zhai L, Wang TT, LI S, Zhang YH
and Guo YL: Picroside II plays a neuroprotective effect by
inhibiting cyto C/caspase-9/caspase-3 signal pathway following
ischemia/reperfusion injury in rats. Chin Pharmacol Bull.
33:668–674. 2017.
|
|
44
|
Kanwar JR, Kamalapuram SK and Kanwar RK:
Survivin signaling in clinical oncology: A multifaceted dragon. Med
Res Rev. 33:765–789. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang F, Cui J, Lv B and Yu B: Nicorandil
protects mesenchymal stem cells against hypoxia and serum
deprivation-induced apoptosis. Int J Mol Med. 36:415–423. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gao J, Bai HJ, Wan F, Tian M, Li YY, Li YX
and Si YC: Effects of ginsenoside on proliferation and
differentiation of neural stem cells undergone oxygen-glucose
deprivation/reperfusion. CJTCMP. 32:2291–2297. 2017.
|
|
47
|
Ejtehadifar M, Shamsasenjan K,
Movassaghpour A, Akbarzadehlaleh P, Dehdilani N, Abbasi P,
Molaeipour Z and Saleh M: The effect of hypoxia on mesenchymal stem
cell biology. Adv Pharm Bull. 5:141–149. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Vangeison G, Carr D, Federoff HJ and Rempe
DA: The good, the bad, and the cell type-specific roles of hypoxia
inducible factor-1 alpha in neurons and astrocytes. J Neurosci.
28:1988–1993. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Park IH, Kim KH, Choi HK, Shim JS, Whang
SY, Hahn SJ, Kwon OJ and Oh IH: Constitutive stabilization of
hypoxia-inducible factor alpha selectively promotes the
self-renewal of mesenchy-mal progenitors and maintains mesenchymal
stromal cells in an undifferentiated state. Exp Mol Med.
45:e442013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ball SG, Shuttleworth CA and Kielty CM:
Vascular endothelial growth factor can signal through
platelet-derived growth factor receptors. J Cell Biol. 177:489–500.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Liao FL, Chen RL, Jiang S and Tian C:
Effects of Notch signaling pathway on VEGF promoting Rat
mesenchymal stem cells proliferation. Zhongguo Shi Yan Xue Ye Xue
Za Zhi. 22:1068–1071. 2014.(In Chinese). PubMed/NCBI
|
|
52
|
Dai Y, Xu M, Wang Y, Pasha Z, Li T and
Ashraf M: HIF-1alpha induced-VEGF overexpression in bone marrow
stem cells protects cardiomyocytes against ischemia. J Mol Cell
Cardiol. 42:1036–1044. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhong Q, Zhou Y, Ye W, Cai T, Zhang X and
Deng DY: Hypoxia-inducible factor 1-α-AA-modified bone marrow stem
cells protect PC12 cells from hypoxia-induced apoptosis, partially
through VEGF/PI3K/Akt/FoxO1 pathway. Stem Cells Dev. 21:2703–2717.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Oosthuyse B, Moons L, Storkebaum E, Beck
H, Nuyens D, Brusselmans K, Van-Dorpe J, Hellings P, Gorselink M,
Heymans S, et al: Deletion of the hypoxia-response element in the
vascular endothelial growth factor promoter causes motor neuron
degeneration. Nat Genet. 28:131–138. 2001. View Article : Google Scholar : PubMed/NCBI
|