Introduction
Schizophrenia is a severe mental disorder, which
affects ~1% of the world's population (1). Approximately 70% of schizophrenia cases
are inheritable and the condition has a major impact on the quality
of life (2). Currently, the
pathogenesis of schizophrenia remains unclear (2). Glutamate is an excitatory
neurotransmitter, which interacts with the
N-methyl-D-aspartate receptor (NMDAR), a subtype of
glutamate receptors. NMDAR is associated with learning, memory,
cognition and synaptic plasticity (3,4). NMDAR
subunit 1 (NR1) is a major functional subunit of the NMDAR family
(5). NMDAR is regarded as a major
contributing factor in the development of schizophrenia (6) and is associated with specific symptoms
induced by changes in the glutamatergic system (7).
Several studies have reported that non-competitive
NMDAR antagonists, including dizocilpine (MK-801), phencyclidine
and ketamine, impair spatial-delayed alternation performance and
produce similar behavioral responses in psychosis (6,8,9). Animal models with psychosis have been
established based on these agents (10). Furthermore, studies have demonstrated
that NMDAR is dysfunctional in schizophrenia, particularly in the
hippocampus (11,12). It is well-known that the hippocampus
is the region of the brain associated with emotions, learning and
memory and synaptic plasticity, which suggests that NMDARs,
specifically a dysregulation or hypofunction of NMDARs, may serve a
key role in the pathogenic process of schizophrenia (4). Previous studies have indicated that
adult neurogenesis is presented within the hippocampus (13,14).
However, studies are yet to determine associations between NMDARs
and hippocampal neural stem cell (NSC) proliferation and
apoptosis.
In a previous study, NMDAR antagonist MK-801 was
evaluated to establish a schizophrenia-like mouse model, which
produced behavioral responses that closely resembled those observed
in hippocampal neurogenesis (14–16).
Other studies focused on MK-801 to establish schizophrenia-like
symptoms and models of behavior (6,17).
In the present study, MK-801 was employed to
establish a cell model of schizophrenia, which was used to
investigate effects of NMDA on proliferation and apoptosis of
hippocampal NSCs. To confirm an association between NMDARs and
hippocampal NSCs proliferation and apoptosis induced by MK-801, the
mechanism of NMDAR in the pathogenesis of schizophrenia and
regulation of hippocampal NSCs was evaluated. Overall, the present
study contributed to a better understanding of the mechanisms of
schizophrenia and provided an experimental basis for further
research in this area.
Materials and methods
Materials
Dulbecco's modified Eagle's medium (DMEM)/nutrient
mixture F-12 (F12), StemPro® Accutase® Cell
Dissociation reagent, fetal bovine serum (FBS) and B27 were
purchased from Life Technologies (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). Epidermal growth factor (EGF) and basic
fibroblast growth factor (bFGF) were purchased from PeproTech, Inc.
(Rocky Hill, NJ, USA). Anti-nestin antibody (cat. no. N5413),
poly-L-lysine hydrobromide (PLL), MK-801 and NMDA were obtained
from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). CellTiter
96® Aqueous One Solution Cell Proliferation assay (MTS)
was purchased from Promega Corporation (Madison, WI, USA). Annexin
V-fluorescein isothiocyanate (FITC) apoptosis assay kit was
purchased from BestBio Company (Shanghai, China).
Animals
A total of 50 neonatal C57/BL mice pups <24 h old
(weight, ~1.2 g) were provided by the Experimental Animal Centre of
Ningxia Medical University (Yinchuan, China). The male:female ratio
was 1:1. Animals were housed in an animal room (temperature,
22–26°C; relative humidity, 40–60%) with a 12 h light/dark cycle.
Maternal animals were individually housed in cages and had free
access to standard laboratory chow and water. The newborn offspring
were used for the present study within 24 h of birth. The mice were
provided by the Experimental Animal Center of Ningxia Medical
University (Yinchuan, China). All protocols were preapproved by the
Ethics Committee of Ningxia Medical University (2014–014).
Primary culture of NSCs
The bodies of newborn C57/BL mice were disinfected
with 75% ethanol. Mice were decapitated to obtain the brain tissue,
which was kept on ice. Hippocampal tissues were dissected under an
anatomic microscope and washed in cold Dulbecco's PBS (D-PBS).
Accutase treatment was combined with mechanical separation to
digest the hippocampal tissues and the isolated cells were
suspended in DMEM/F12, then centrifuged at 200 × g at room
temperature (RT) for 5 min and the supernatant was discarded. The
cell suspension was collected following passing through a 400-mesh
sieve and seeded in culture bottles (25 cm3). The
conditional culture medium was composed of DMEM/F12 (1:1), B27
(2%), bFGF (20 ng/ml), EGF (20 ng/ml) and penicillin-streptomycin
(100 U/ml). The density of cells was 0.5–1.0×106/ml. The
original volume of conditioned medium was 4 ml following culturing
at 37°C in an atmosphere of 5% CO2 for 3–4 days an
additional 1 ml conditioned medium was added to the culture bottle.
At 5–7 days post-culture the diameter of the neurospheres were ~100
µm and subculturing was performed. NSCs were digested by accutase
for ~5 min. The NSC suspension was collected and centrifuged at 200
× g at RT for 5 min. The supernatant was discarded and the cells
were resuspended in conditional culture medium. The cellular
density was 0.5–1.0×106/ml in a culture bottle. The
cells were incubated in an incubator containing 5% CO2
at 37°C.
Identification of NSCs
Immunofluorescence staining of nestin (a maker of
NSCs) was performed to confirm the successful culture of NSCs. In
brief, 0.01% PLL was coated on the sterile cover slips in 24-well
plates for 30 min. Then the 24-well plates were washed three times
with PBS. On day 4, the 2nd generation of NSCs were seeded at a
density of 1.0×105/ml in the 24-well plates coated with
PLL for 24 h in an incubator containing 5% CO2 at 37°C.
The NSCs were washed with 0.01 M PBS and fixed with 4%
paraformaldehyde for 30 min at RT, rinsed with 0.01 M PBS and
incubated with 10% BSA for 1 h at 37°C. The cells were incubated
overnight at 4°C with the primary antibodies against nestin
(1:200). In the control group, the primary antibodies were
substituted with 0.01 M PBS. The glass plates were subsequently
washed with 0.01 M PBS three times and incubated with the
FITC-labeled goat anti-rabbit IgG (H+L) (1:200; cat. no. A0562;
Beyotime Institute of Biotechnology, Haimen, China) secondary
antibodies at RT for 2 h. Then slides were washed three times with
0.01 M PBS. DAPI was used to counterstain the nuclei at room
temperature for 10 min and then washed three times with 0.01 M PBS.
Images were acquired using an Olympus FV1000 confocal microscope.
The positive cells were quantified using Image-Pro Plus 6.0
software (Media Cybernetics, Inc., Rockville, MD, USA).
Preparation of MK-801 and NMDA
MK-801 was dissolved in DMEM/F12 at a concentration
of 5 mM and stored at −20°C. Aliquots were further diluted in
DMEM/F12 to a final concentration of 50, 100, 200, 400 and 800 µM
in wells. NMDA was dissolved in DMEM/F12 at a concentration of 50
mM and stored at −20°C. Aliquots were further diluted in DMEM/F12
to a concentration of 5 mM and then to a final concentration of 50,
100, 200, 400 and 800 µM in wells.
MTS assay
To assess effects of MK-801 and NMDA on hippocampal
NSCs proliferation, four experimental groups were defined: Control
group; MK-801 (M) group, with cells treated with 200 µM MK-801 for
24 h in an incubator at 37°C; NMDA (N) group, with cells treated
with 100 µM NMDA for 2 h in an incubator at 37°C; M+N group, with
cells treated with 200 µM MK-801 for 24 h prior to adding 100 µM
NMDA for 2 h in an incubator at 37°C. The 96-well plates contained
1.0×104 cells per well and the NSCs were incubated in an
incubator containing 5% CO2 at 37°C. Following the above
treatment, 20 µl of MTS was pipetted into each well of the 96-well
plate containing cells in 100 µl culture medium. Plates were
incubated at 37°C for 4 h in a humidified, 5% CO2
atmosphere. Absorbance at 490 nm was recorded using a plate
reader.
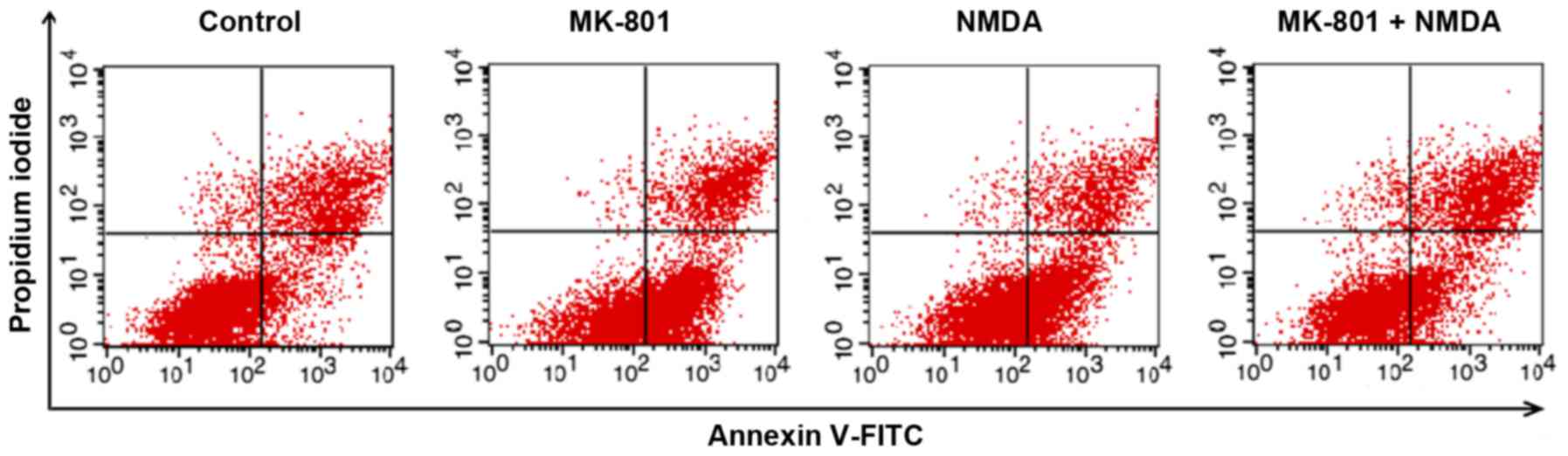
Flow cytometry
Apoptosis in NSCs was measured by flow cytometry
following staining with Annexin V-FITC and propidium iodide (PI).
Cells were treated as described previously and resuspended in
binding buffer at 1.0×105 cells/ml. Cell sample solution
(100 µl) was incubated with FITC-conjugated annexin V (5 µl) and PI
(5 µl) for 15 min at room temperature in the dark. Cells were
analyzed by flow cytometry with FlowJo software 7.6.5 (FlowJo LLC,
Ashland, OR, USA). Annexin V is a sensitive indicator for detecting
early apoptosis in cells. The nucleus of cells in middle and late
stages of apoptosis is stained by PI.
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical significance was calculated by one-way analysis of
variance followed by the LSD post hoc test. All experiments were
performed in triplicate. P<0.05 was considered to indicate a
statistically significant difference.
Results
Identification of NSCs
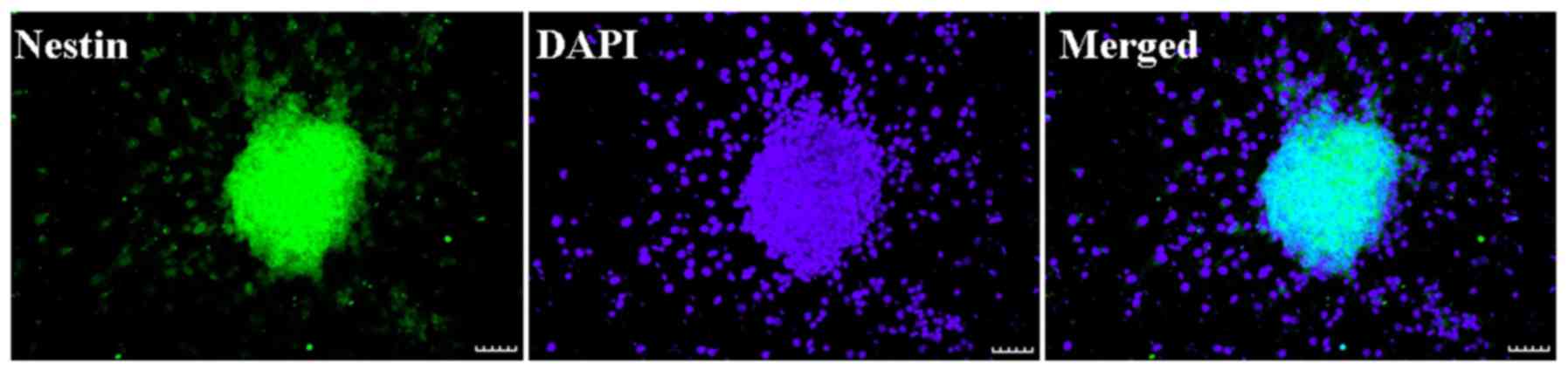
Neurosphere with nestin expression (green) and DAPI
staining (blue) are presented in Fig.
1. Nestin is an identification marker of NSCs or neural
progenitor marker. DAPI is an identification marker for cell
nuclei. The merged image demonstrated that nestin (green) and DAPI
(blue) were co-localized in the cultured cells. The result
indicated that NSCs had a high purity and may be applied in the
proposed study.
Concentration dependence of MK-801 and
NMDA treatments on NSCs viability
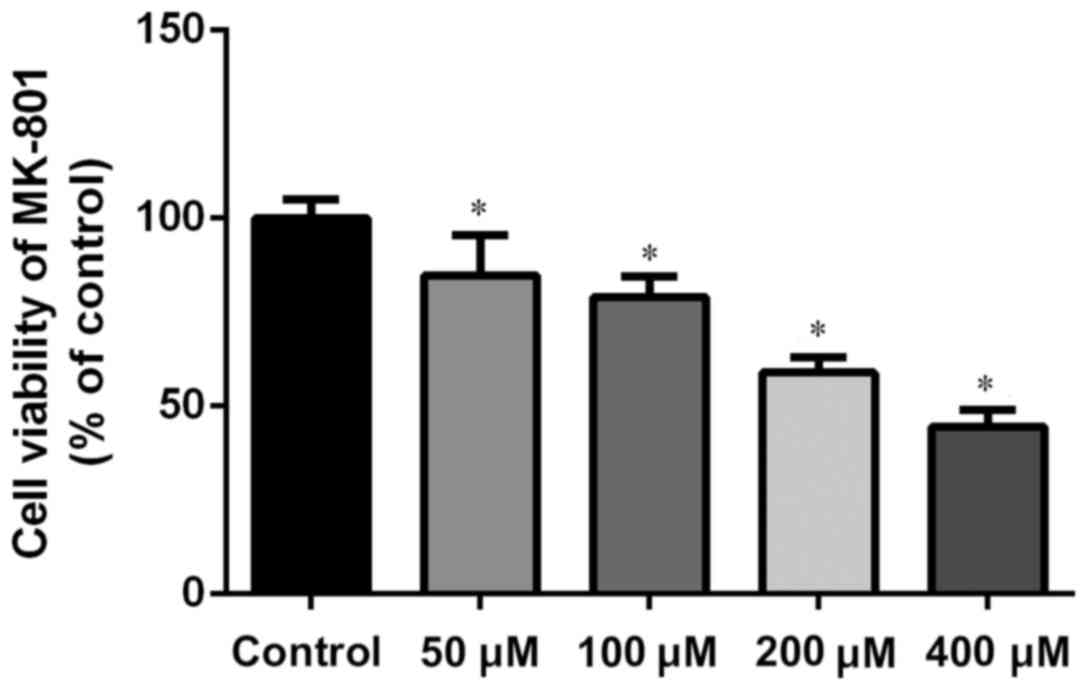
The concentrations of MK-801 and NMDA were confirmed
through the MTS test. MK-801 and NMDA inhibited growth of NSCs in a
dose-dependent manner. The research dosage of drugs is always
confirmed using the IC50, which is the dosage at which
50% of the cells die. Typically the dosage of a drug selected for
treatment use will cause 50–80% cell viability. MK-801 at 200 µM is
~60% cell viability (200 µM for 24 h; Fig. 2). Therefore, MK-801 (200 µM for 24 h)
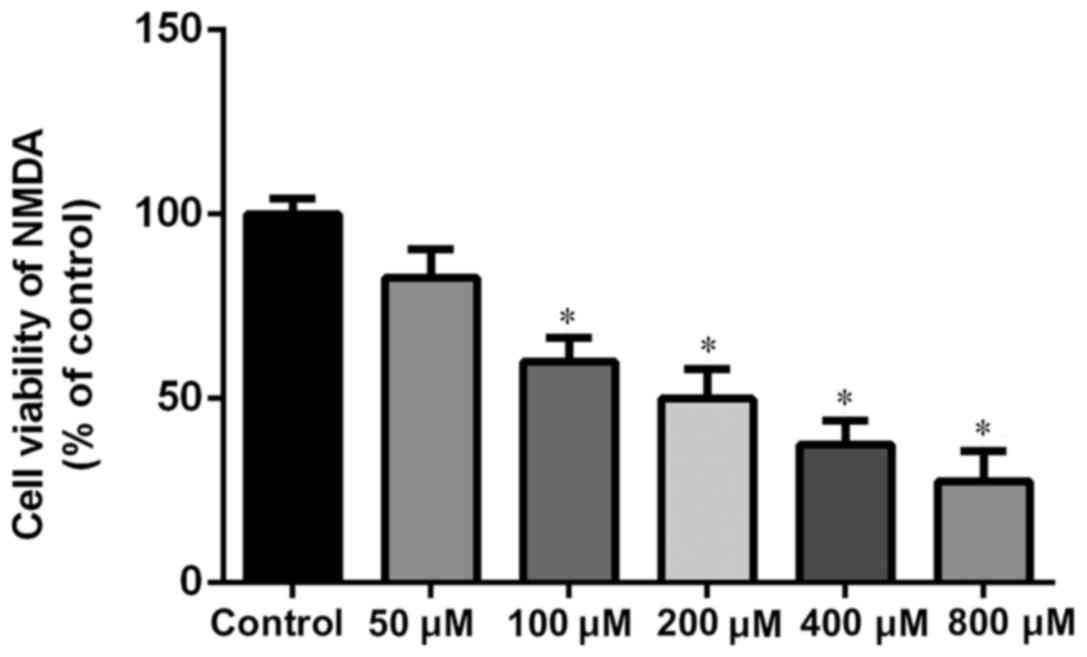
was used in the present experiments. NMDA at 100 µM is ~60% cell
viability (100 µM for 2 h; Fig. 3).
Therefore NMDA (100 µM for 2 h) was selected for use in the present
experiments.
MK-801 and NMDA affect hippocampal
NSCs proliferation
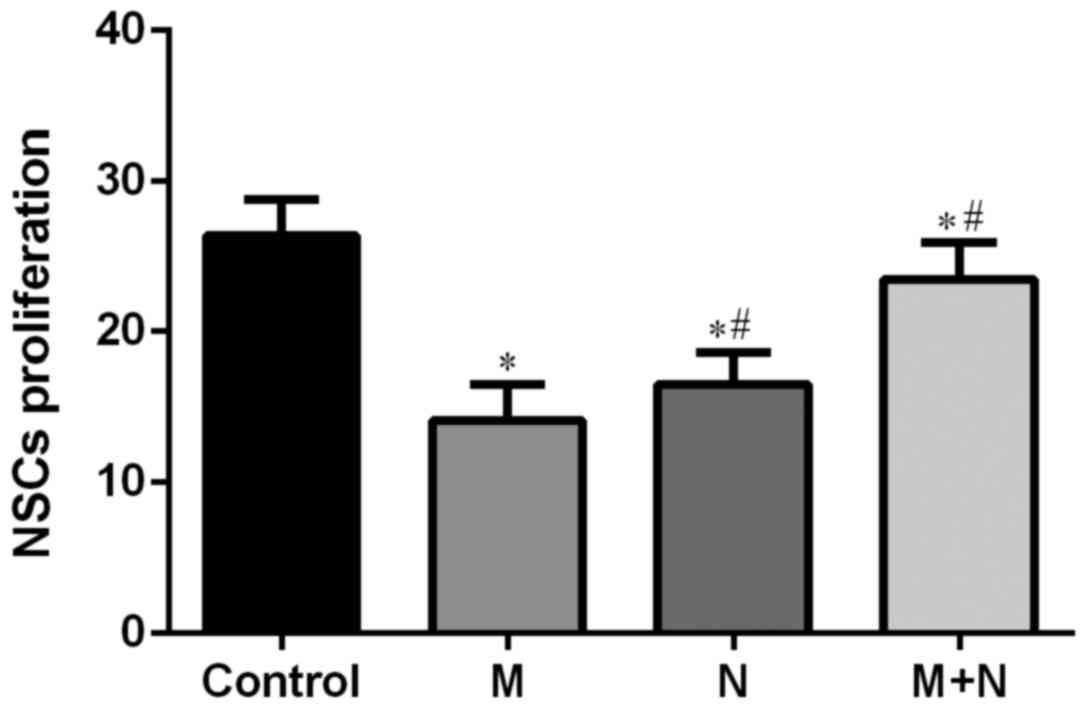
Compared with the control group, cell survival rates
of M, N and M+N groups were significantly decreased (P<0.01).
Compared with the M group, cell survival rates of N and M+N groups
increased significantly (P<0.01; Fig.
4).
Influence of NMDA and MK-801 treatment
on NSCs apoptosis
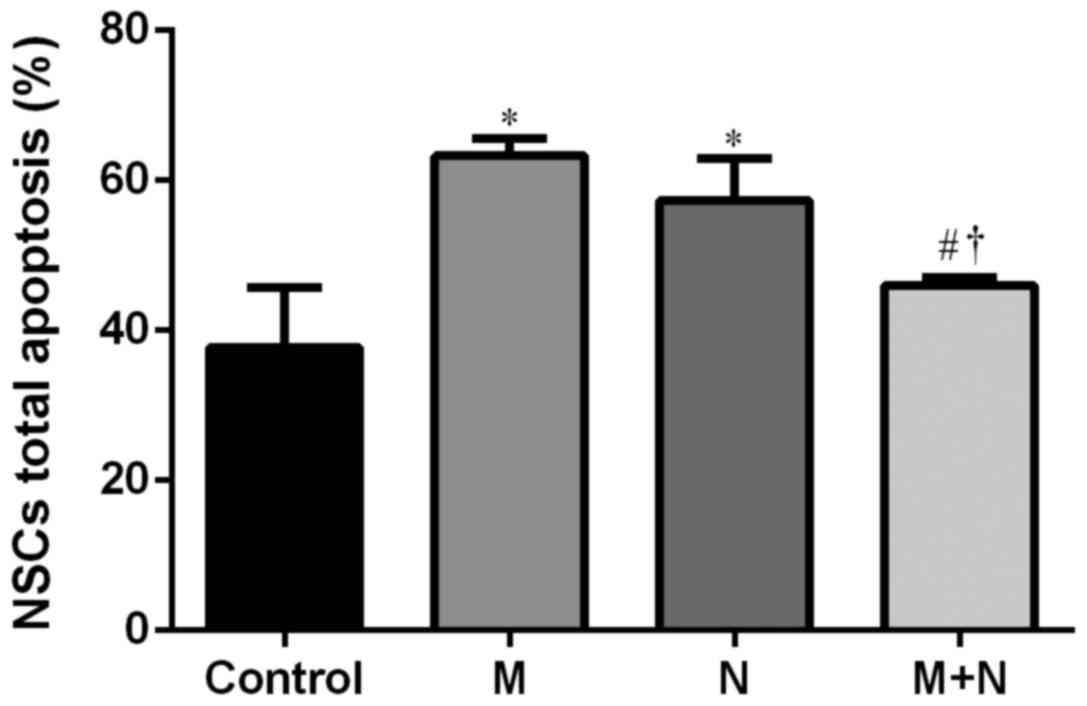
Total apoptosis rates of NSCs in M and N groups
increased compared with the control group (P<0.01). Compared
with M or N group, the total apoptosis rate of the M+N group
decreased (P<0.05). No significant difference was observed
between the M and N groups (Figs. 5
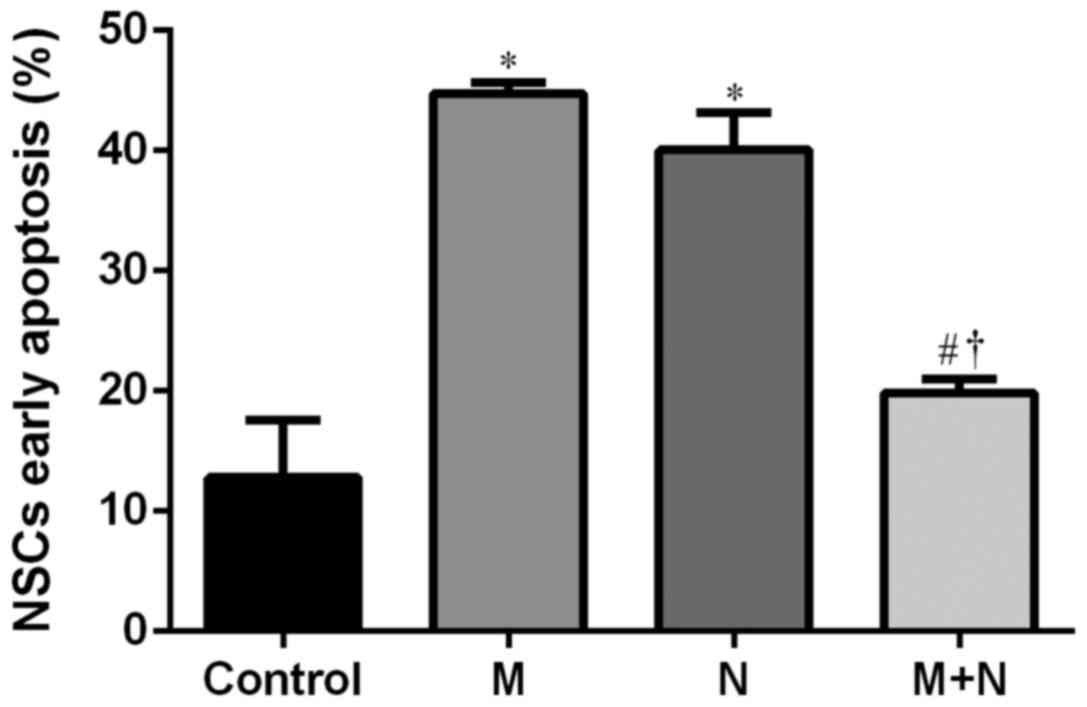
and 6). Early stage apoptotic rates
of NSCs in M and N groups were revealed to increase compared with
the control group (P<0.001). Compared with M and N groups, the
early stage apoptotic rate of the M+N group decreased (P<0.001).
No significant difference was observed between M and N groups
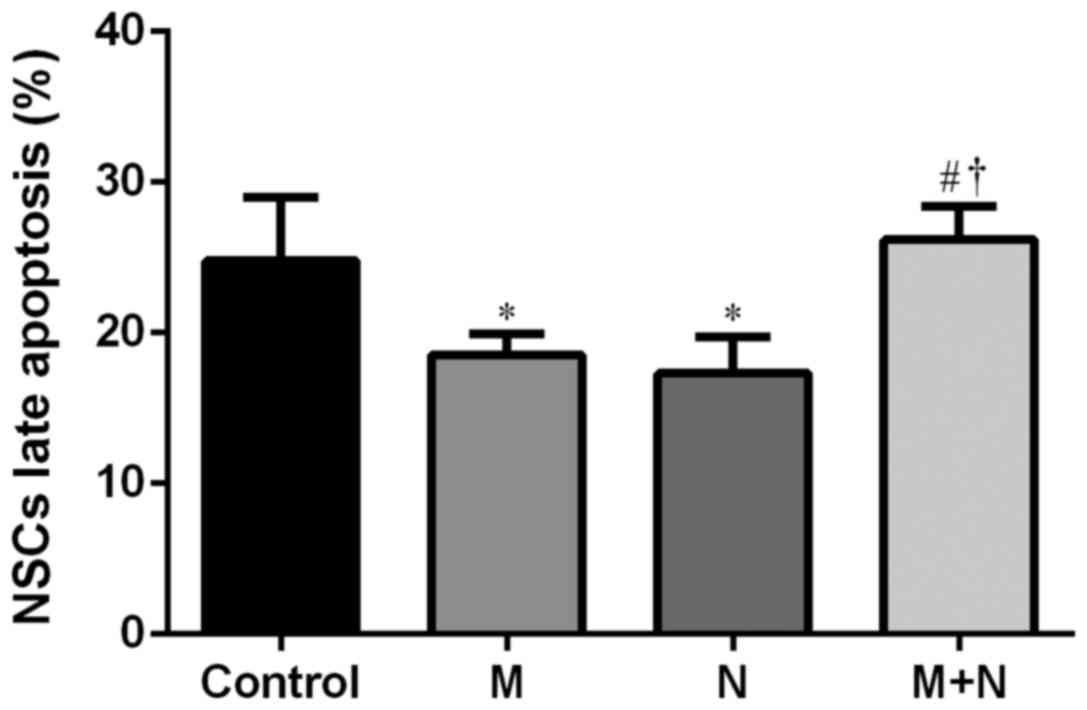
(Fig. 7). Late stage apoptotic rates
of NSCs in M and N groups were significantly reduced compared with
the control group (P<0.01). Compared with the M and N groups,
the late stage apoptotic rate of the M+N group increased
(P<0.01). There was no significant difference between the M and
N groups (Fig. 8).
Discussion
Relapse rates of schizophrenia have become a focus
of attention as its etiology and pathogenesis remain unclear
(2). Glutamatergic dysfunction is
known to be associated with the pathology of schizophrenia
(17). The glutamate receptor
hypothesis postulates that schizophrenia occurs in the hippocampus
through NMDAR glutamate neurotransmitter system exceptions
(5). Glutamic acid caused by
glutamate ester is unusual and may result in symptoms of
schizophrenia (5). Research has
revealed that NMDAR agonists improve symptoms of schizophrenia,
indicating that glutamate NMDAR serves an important role in the
pathogenesis of schizophrenia (10).
NMDAR is an ionic, excitatory glutamate receptor in
the CNS, which is expressed in the hippocampus and cerebral cortex
area (18). NMDAR stimulates nerve
growth and synaptogenesis and the excited mature receptor promotes
learning, memory and other higher order functions (19). Excessive activation of NMDAR causes
Ca2+ accumulation in neurons, inducing neurotoxicity and
leading to cell death (5). Glycine
is an NMDAR agonist, which is used as adjuvant for antipsychotic
drugs, combined with traditional antipsychotic medicines, it
improves negative symptoms of schizophrenia (20). NMDA and its receptor serve a role in
the CNS and affect schizophrenia at least partially mediated by ion
channel activity (2). NR1 is a
functional subunit of the NMDAR family, which is representative of
NMDAR expression levels and is referred to as the core subunit of
NMDAR (21). Further studies have
revealed that low NR1 expression is associated with low spatial
learning and memory function in schizophrenia, and the glutamic
acid system and other specific regions of the brain expressing
NMDAR are closely associated with schizophrenia (11,22). A
previous study using a rat model demonstrated that depression
following cerebral hemorrhage was caused by enhanced expression of
the NMDARs NR1 and NR2B in the hippocampus, accompanied by
decreased bromodeoxyuridine expression (23). The results presented in the present
study demonstrated that NMDAR antagonist MK-801 and NMDAR agonist
NMDA decreased cell viability. NMDARs were significantly inhibited
by MK-801 or activated by NMDA and demonstrated neurotoxicity
through decreased cell viability. MK-801 inhibited the NMDAR
inducing neurotoxicity. Subsequent addition of NMDA activated NMDAR
and neutralized the inhibitory effect of MK-801. NMDA may inhibit
neurotoxicity induced by MK-801.
Neurogenesis involves proliferation and
differentiation of neural precursor cells in the developing and
adult brain, which is associated with NMDAR (19). The dysfunction of adult neurogenesis
is associated with the etiology of schizophrenia (23). At a cellular level, cell
proliferation is a major biological characteristic. The data
presented in the present study were obtained using MTS assays and
flow cytometry to determine effects of MK-801 and NMDA on
proliferation and apoptosis in hippocampal NSCs. MK-801 and NMDA
inhibited cell proliferation when compared with the control cells,
as using MK-801 or NMDA alone significantly inhibited or activated
NMDARs, respectively, leading to neurotoxicity and decreased cell
proliferation. Treatment with MK-801, to inhibit NMDARs, followed
by treatment with NMDA, to reactivate NMDARs, was demonstrated to
neutralize the initial effect of MK-801. NMDA reversed the
inhibitory effect of MK-801 on cell proliferation. MK-801 and NMDA
demonstrated to induce apoptosis when compared with the control
cells. Whereas MK-801 treatment inhibited NMDARs, subsequent
treatment with NMDA suppressed stimulatory effects of MK-801 on
cell apoptosis. Apoptotic rates of hippocampal NSCs significantly
decreased, suggesting that NMDARs may be associated with the
process of hippocampal NSC apoptosis and proliferation.
MK-801 inhibits the function of glutamate in
hippocampal neurons (24). By
blocking Ca2+ channels, it induces neurotoxic effects,
which prevent hippocampal neurogenesis and promote apoptosis of
hippocampal NSCs (25,26). The NMDAR antagonist ketamine alters
neurogenesis by inhibiting proliferation of NSCs (27). In the present study, treatment with
MK-801 for 24 h followed by treatment with NMDA for 2 h revealed
that cell vitality partially recovered and total and early
apoptotic levels significantly decreased. The underlying mechanisms
may be associated with the activation of NMDAR. NMDA activate NMDAR
on hippocampal neurons, change synaptic plasticity and release
excitatory neurotransmitters that condition and inhibit MK-801 in a
non-competitive role to improve cell vitality (28,29). A
further study has revealed that MK-801 treatment [0.2 mg/kg,
intraperitoneal (IP)] increases NSC proliferation in the
hippocampus of a rat model with Parkinson's disease (30). MK-801 treatment results in enhanced
hippocampal neurogenesis in the model. Differences between this
study and the current study were the dose of MK-801 treatment (0.2
mg/kg, IP, rat) compared with 200 µM added to the cell supernatant,
respectively, and alterations in the model type. Observed effects
of MK-801 were therefore different. Effects of low and high dosage
of MK-801 varied on proliferation and neurogenesis. Excitatory
amino acid toxicity exists in Parkinson's or Alzheimer's disease
and MK-801 may be useful in the treatment of these (31). However, effects of MK-801 may be
harmful for normal cells. A cell model using MK-801 was
established.
In conclusion, on a cellular level, NMDA may inhibit
neurotoxic effects of MK-801 on hippocampal NSCs and significantly
improve hippocampal NSCs activity and rate of apoptosis, which
serves an essential role in proliferation and apoptosis.
Limitations of the current study include missing NMDAR knockout or
gene silencing experiments. However, MK-801 and NMDA were used to
evaluate the association between NMDAR and proliferation and
apoptosis of hippocampal NSCs. Exploring regulatory effects of
NMDARs on the functioning of the hippocampus and schizophrenia is
of importance for an increased understanding of the underlying
biology of mental health and it may facilitate identifying novel
treatment targets.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81160169, 81460214,
31660270 and 31460255) and West China Top Class Discipline Project
in Basic Medical Sciences, Ningxia Medical University (grant no.
NXYLXK2017B07).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JD performed the data analyses and drafted the
manuscript. YS performed the experiments. HHZ assisted with the
experiments. QRM, YWZ, YXD and YQH collected and interpreted the
data. JL conceived the study. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
All experimental protocol was approved by the Ethics
Committee of Ningxia Medical University (approval no.
2014-014).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lichtenstein P, Yip BH, Bjork C, Pawitan
Y, Cannon TD, Sullivan PF and Hultman CM: Common genetic
determinants of schizophrenia and bipolar disorder in Swedish
families: A population-based study. Lancet. 373:234–239. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shorter KR and Miller BH: Epigenetic
mechanisms in schizophrenia. Prog Biophys Mol Biol. 118:1–7. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fouad IA, Sharaf NM, Abdelghany RM and El
Sayed NSED: Neuromodulatory effect of thymoquinone in attenuating
glutamate-mediated neurotoxicity targeting the amyloidogenic and
apoptotic pathways. Front Neurol. 9:2362018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dore K, Stein IS, Brock JA, Castillo PE,
Zito K and Sjostrom PJ: Unconventional NMDA receptor signaling. J
Neurosci. 37:10800–10807. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ju P and Cui D: The involvement of
N-methyl-d-aspartate receptor (NMDAR) subunit NR1 in the
pathophysiology of schizophrenia. Acta Biochim Biophys Sin
(Shanghai). 48:209–219. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim TW, Kang HS, Park JK, Lee SJ, Baek SB
and Kim CJ: Voluntary wheel running ameliorates symptoms of
MK-801-induced schizophrenia in mice. Mol Med Rep. 10:2924–2930.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Moghaddam B and Javitt D: From revolution
to evolution: The glutamate hypothesis of schizophrenia and its
implication for treatment. Neuropsychopharmacology. 37:4–15. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Krystal JH, Karper LP, Seibyl JP, Freeman
GK, Delaney R, Bremner JD, Heninger GR, Bowers MB Jr and Charney
DS: Subanesthetic effects of the noncompetitive NMDA antagonist,
ketamine, in humans. Psychotomimetic, perceptual, cognitive, and
neuroendocrine responses. Arch Gen Psychiatry. 51:199–214. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lahti AC, Koffel B, LaPorte D and Tamminga
CA: Subanesthetic doses of ketamine stimulate psychosis in
schizophrenia. Neuropsychopharmacology. 13:9–19. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Javitt DC and Zukin SR: Recent advances in
the phencyclidine model of schizophrenia. Am J Psychiatry.
148:1301–1308. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Duncan GE, Inada K, Koller BH and Moy SS:
Increased sensitivity to kainic acid in a genetic model of reduced
NMDA receptor function. Brain Res. 1307:166–176. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Harrison PJ, Law AJ and Eastwood SL:
Glutamate receptors and transporters in the hippocampus in
schizophrenia. Ann N Y Acad Sci. 1003:94–101. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Emsley JG, Mitchell BD, Kempermann G and
Macklis JD: Adult neurogenesis and repair of the adult CNS with
neural progenitors, precursors, and stem cells. Prog Neurobiol.
75:321–341. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu J, Suzuki T, Seki T, Namba T, Tanimura
A and Arai H: Effects of repeated phencyclidine administration on
adult hippocampal neurogenesis in the rat. Synapse. 60:56–68. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tanimura A, Liu J, Namba T, Seki T,
Matsubara Y, Itoh M, Suzuki T and Arai H: Prenatal phencyclidine
exposure alters hippocampal cell proliferation in offspring rats.
Synapse. 63:729–736. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yamaguchi M, Suzuki T, Seki T, Namba T,
Juan R, Arai H, Hori T and Asada T: Repetitive cocaine
administration decreases neurogenesis in adult rat hippocampus. Ann
N Y Acad Sci. 1025:351–362. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu J, Qi D, Xing M, Li R, Jiang K, Peng Y
and Cui D: MK-801 induces schizophrenic behaviors through
downregulating Wnt signaling pathways in male mice. Brain Res.
1385:281–292. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bersier MG, Pena C and de Lores Arnaiz
Rodriguez G: The expression of NMDA receptor subunits in cerebral
cortex and hippocampus is differentially increased by
administration of endobain E, a Na+, K+-ATPase inhibitor. Neurochem
Res. 33:66–72. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Baez MV, Cercato MC and Jerusalinsky DA:
NMDA receptor subunits change after synaptic plasticity induction
and learning and memory acquisition. Neural Plast.
2018:50930482018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Evins AE, Fitzgerald SM, Wine L, Rosselli
R and Goff DC: Placebo-controlled trial of glycine added to
clozapine in schizophrenia. Am J Psychiatry. 157:826–828. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Andersson O, Stenqvist A, Attersand A and
von Euler G: Nucleotide sequence, genomic organization, and
chromosomal localization of genes encoding the human NMDA receptor
subunits NR3A and NR3B. Genomics. 78:178–184. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Karlsgodt KH, Robleto K, Trantham-Davidson
H, Jairl C, Cannon TD, Lavin A and Jentsch JD: Reduced dysbindin
expression mediates N-methyl-D-aspartate receptor hypofunction and
impaired working memory performance. Biol Psychiatry. 69:28–34.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Taliaz D, Stall N, Dar DE and Zangen A:
Knockdown of brain-derived neurotrophic factor in specific brain
sites precipitates behaviors associated with depression and reduces
neurogenesis. Mol Psychiatry. 15:80–92. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Glushakov AV, Dennis DM, Morey TE, Sumners
C, Cucchiara RF, Seubert CN and Martynyuk AE: Specific inhibition
of N-methyl-D-aspartate receptor function in rat hippocampal
neurons by L-phenylalanine at concentrations observed during
phenylketonuria. Mol Psychiatry. 7:359–367. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
D'Ascenzo M, Piacentini R, Casalbore P,
Budoni M, Pallini R, Azzena GB and Grassi C: Role of L-type Ca2+
channels in neural stem/progenitor cell differentiation. Eur J
Neurosci. 23:935–944. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ambrosio AF, Silva AP, Malva JO,
Soares-da-Silva P, Carvalho AP and Carvalho CM: Carbamazepine
inhibits L-type Ca2+ channels in cultured rat hippocampal neurons
stimulated with glutamate receptor agonists. Neuropharmacology.
38:1349–1359. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang H, Liu L, Li B, Zhao PP, Xu CM, Zhu
YZ, Zhou CH and Wu YQ: Ketamine interferes with the proliferation
and differentiation of neural stem cells in the subventricular zone
of neonatal rats. Cell Physiol Biochem. 35:315–325. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Simoes AP, Silva CG, Marques JM, Pochmann
D, Porciúncula LO, Ferreira S, Oses JP, Beleza RO, Real JI, Köfalvi
A, et al: Glutamate-induced and NMDA receptor-mediated
neurodegeneration entails P2Y1 receptor activation. Cell Death Dis.
9:2972018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Motin VG and Yasnetsov VV: Effect of NMDA,
a specific agonist to NMDA receptor complex, on rat hippocampus.
Bull Exp Biol Med. 159:704–707. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Singh S, Mishra A, Srivastava N and Shukla
S: MK-801 (Dizocilpine) regulates multiple steps of adult
hippocampal neurogenesis and alters psychological symptoms via
Wnt/beta-catenin signaling in parkinsonian rats. ACS Chem Neurosci.
8:592–605. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kysenius K, Brunello CA and Huttunen HJ:
Mitochondria and NMDA receptor-dependent toxicity of berberine
sensitizes neurons to glutamate and rotenone injury. PLoS One.
9:e1071292014. View Article : Google Scholar : PubMed/NCBI
|