Introduction
Staphylococcus aureus is a Gram-positive
bacterium that typically colonizes the surface of the skin and, as
an important pathogen of nosocomial cross-infection, it is also a
common pathogenic bacterium of biofilm infections (1,2). More
than 80% of human infections are caused by biofilms, including
endocarditis, osteomyelitis and implant-associated infections
(3). S. aureus-associated
infections lead to an increase in hospital stays, in addition to
hospital-associated mortality (4).
S. aureus biofilms in humans cause chronic persistent
infections that are difficult to cure and represent a great
challenge for clinical treatment (5).
Biofilms may adhere to surfaces and are enclosed in
a self-produced polymeric matrix, which enhances antibiotic
resistance and is shielded from the immune system (6). Biofilm-associated infections are
difficult to treat and these chronic or relapsing infections
typically require increased drug doses or prolonged antibiotic
treatment, which is associated with development of drug resistances
(7). Biofilm resistance is commonly
10–1,000 times stronger than that of planktonic bacteria (8). At present, vancomycin (VCM) is the most
commonly administered drug for S. aureus biofilm-associated
infections (9). However, there is
cause for concern due to recent developments of VCM-intermediate
S. aureus and vancomycin-resistant S. aureus (VRSA)
strains (10). Combination of VCM
with rifampin, oxacillin, linezolid and tigecycline has been
considered in order to improve treatment effectiveness and to
reduce resistance (11).
Nevertheless, studies (12–14) have indicated that although this
combination may be effective against methicillin-sensitive S.
aureus, it may not hold promise for use in treating
methicillin-resistant S. aureus biofilm infections.
Therefore, there is a requirement to identify innovative and novel
approaches for the prevention of biofilm formation.
Several studies (15–17) have
reported that the use of traditional herbal medicine in the
prevention and treatment of biofilm infections is advantageous.
Traditional herbal medicine may not only inhibit the formation of
biofilms and destroy biofilms, but it may further enhance
resistance to infection when combined with antibiotics (18). At the same time, these treatments
exhibit little side effects, strong pharmacological action, wide
availability and do not easily lead to bacterial drug resistance
(19–20). This has become another notable focus
of research, distinct from antibiotics. A recent study screened 40
types of traditional herbal medicines common in the Guizhou
province of China to test for minimum inhibitory concentration
(MIC) for S. aureus ATCC25923 biofilms (21). The results demonstrated that
Caesalpinia sappan L. has bacteriostatic properties. These
findings were further analyzed in the present study.
Natural products may be utilized as raw materials in
the development of novel antibacterial substances (18). Brazilin (BN) is isolated from the
traditional herbal medicine Caesalpinia sappan L and is a
principal active component of it (20). Evidence has indicated that BN
exhibits multiple biological properties, including immune system
modulatory, antioxidant, anti-inflammatory, antiplatelet,
antihepatotoxicity and antitumor activities (19,20).
Furthermore, a number of studies have demonstrated that BN is
associated with antimicrobial activity (22,23).
The current study aimed to determine whether, and
the degree to which, BN may inhibit S. aureus biofilm
formation and increase antibiotic susceptibility. Another aim was
to demonstrate whether BN was able to reduce secretion of
extracellular polysaccharides and interfere with the quorum-sensing
(QS) system. To the best of our knowledge, this has not yet been
reported and requires further research.
Materials and methods
Bacterial strains
A total of 50 clinical strains of S. aureus were
prepared. Samples were collected from The Affiliated Hospital of
Zunyi Medical University (Guizhou, China) in January 2017 (Table I) and patients provided written
informed consent prior to collection of blood samples and swabs.
The present study was approved by the Ethics Committee of the
Affiliated Hospital of Zunyi Medical University. Isolated S. aureus
C-4-4 was selected as preliminary studies revealed it had the
strongest ability to form biofilms, as determined by Congo red and
Crystal violet staining (data not shown) and is the strain
discussed subsequently. The S. aureus ATCC25923 standard strain
(American Type Culture Collection, Manassas, VA, USA) was used as
quality control. S. aureus strains were routinely cultured on blood
agar (BA) plates at 37°C and 5% CO2 for 24 h. Single
colonies were selected from a culture plate and the bacterial
suspension was diluted in Mueller Hinton (M-H) broth (BD Difco™; BD
Biosciences, Franklin Lakes, NJ, USA) to an optical density at 600
nm (OD600) of 0.1 prior to use.
 | Table I.Samples collected from The Affiliated
Hospital of Zunyi Medical University, Zunyi, China. |
Table I.
Samples collected from The Affiliated
Hospital of Zunyi Medical University, Zunyi, China.
| No | Strain_ID | Age | Sex | Infection site | Month and year |
|---|
| 1 | C-3–077 | 2 | Male | Wound | Jan-17 |
| 2 | C-3–078 | 1 | Female | Wound | Jan-17 |
| 3 | C-3–079 | 54 | Male | Abscess | Jan-17 |
| 4 | C-4–013 | 23 | Female | Respiratory | Jan-17 |
| 5 | C-3–080 | 8 | Male | Respiratory | Jan-17 |
| 6 | C-3–081 | 23 | Female | Abscess | Jan-17 |
| 7 | C-3–082 | 55 | Female | Respiratory | Jan-17 |
| 8 | C-3–083 | 41 | Male | Wound | Jan-17 |
| 9 | C-3–084 | 33 | Male | Wound | Jan-17 |
| 10 | C-3–085 | 37 | Male | Respiratory | Jan-17 |
| 11 | C-3–086 | 3 | Female | Wound | Jan-17 |
| 12 | C-3–087 | 39 | Female | Abscess | Jan-17 |
| 13 | C-3–088 | 21 | Female | Abscess | Jan-17 |
| 14 | C-3–089 | 2 | Male | Wound | Jan-17 |
| 15 | C-3–090 | 0 | Male | Respiratory | Jan-17 |
| 16 | C-3–091 | 64 | Female | Respiratory | Jan-17 |
| 17 | C-3–092 | 50 | Male | Wound | Jan-17 |
| 18 | C-3–093 | 57 | Female | Respiratory | Jan-17 |
| 19 | C-3–094 | 75 | Male | Respiratory | Jan-17 |
| 20 | C-3–095 | 0 | Female | Abscess | Jan-17 |
| 21 | C-3–096 | 52 | Male | Respiratory | Jan-17 |
| 22 | C-3–097 | 63 | Male | Respiratory | Jan-17 |
| 23 | C-3–098 | 3 | Female | Wound | Jan-17 |
| 24 | C-3–099 | 23 | Female | Abscess | Jan-17 |
| 25 | C-3–100 | 2 | Female | Wound | Jan-17 |
| 26 | C-4–001 | 0 | Male | Respiratory | Jan-17 |
| 27 | C-4–020 | 71 | Female | Respiratory | Jan-17 |
| 28 | C-4–003 | 0 | Female | Respiratory | Jan-17 |
| 29 | C-4–004 | 46 | Female | Abscess | Jan-17 |
| 30 | C-4–005 | 4 | Female | Wound | Jan-17 |
| 31 | C-4–006 | 47 | Male | Wound | Jan-17 |
| 32 | C-4–007 | 50 | Male | Wound | Jan-17 |
| 33 | C-4–008 | 39 | Male | Wound | Jan-17 |
| 34 | C-4–009 | 60 | Female | Wound | Jan-17 |
| 35 | C-4–010 | 0 | Female | Respiratory | Jan-17 |
| 36 | C-4–011 | 1 | Male | Wound | Jan-17 |
| 37 | C-4–012 | 64 | Female | Wound | Jan-17 |
| 38 | D-1–078 | 45 | Male | Wound | Jan-17 |
| 39 | D-1–079 | 19 | Male | Abscess | Jan-17 |
| 40 | D-1–080 | 5 | Female | Bacteremia | Jan-17 |
| 41 | D-1–081 | 48 | Male | Respiratory | Jan-17 |
| 42 | D-1–082 | 37 | Male | Abscess | Jan-17 |
| 43 | D-1–083 | 51 | Male | Wound | Jan-17 |
| 44 | D-1–084 | 51 | Male | Abscess | Jan-17 |
| 45 | D-1–085 | 5 | Male | Bacteremia | Jan-17 |
| 46 | D-1–086 | 51 | Male | Wound | Jan-17 |
| 47 | D-1–087 | 75 | Male | Respiratory | Jan-17 |
| 48 | D-1–088 | 41 | Male | Cerebrospinal
fluid | Jan-17 |
| 49 | D-1–089 | 49 | Male | Bacteremia | Jan-17 |
| 50 | D-1–090 | 59 | Female | Cerebrospinal
fluid | Jan-17 |
Determination of the effect of BN on
S. aureus
BN (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
purity was confirmed to >98% using high-performance liquid
chromatography. BN (1,024 µg/ml) was dissolved in deionized water
and stored at 4°C until use. The pH of the solution was maintained
at 7.2–7.4. Antimicrobial susceptibility testing of BN and VCM was
performed using the standardized broth microdilution method in
96-well U-bottom plates (Costar 3590; Corning Incorporated,
Corning, NY, USA) using M-H broth, according to the 2015 Clinical
and Laboratory Standards Institute guidelines (24). VCM (0.5 µg/ml; Sigma-Aldrich; Merck
KGaA) was used as a positive control.
Crystal violet staining elaborating
the effect of BN on S. aureus biofilm formation
S. aureus strains were cultured on BA plates at 37°C
and 5% CO2 overnight. The following day, the bacterial
suspension was diluted in M-H broth to OD600=0.1. Bacterial
suspension (100 µl) was applied to sterile polystyrene 96-well
plates and 100 µl BN (1/2, 1 or 2 MIC) were added to final
concentrations of 1/4, 1/2 and 1 MIC, respectively. VCM (100 µl, 2
MIC) was used as positive control and deionized water as blank.
Suspensions were cultivated at 37°C and 5% CO2 for 6,
12, 24 and 48 h. Unbound cells from S. aureus 6-, 12-, 24- and 48-h
incubation plates were removed by washing with PBS (3×). The
biomass of each slice was air-dried for 20 min at 37°C and stained
for 20 min at 25°C with 200 µl filtered 0.25% crystal violet,
washed with PBS (3×) to remove unbound stain, dried at 37°C for 20
min and solubilized in 200 µl 95% ethanol for 15 min at 25°C. A
reading at 570 nm using an ELISA microplate reader (Multiskan FC;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) was obtained.
Experiments were performed six times.
Fluorescence microscopy elaborating
the effect of BN on S. aureus biofilm formation
A polystyrene carrier (10×10 mm) was placed in a
24-well plate and 100 µl bacterial suspension were applied along
with 100 µl BN (1/2, 1 and 2 MIC). VCM (100 µl; 2 MIC) was used as
positive control and deionized water as blank. Plates were
incubated at 37°C and 5% CO2 for 6, 12, 24 and 48 h.
Unbound cells from S. aureus 6-, 12-, 24- and 48-h incubation
plates were removed by washing with PBS (3×). Biofilms were stained
with fluorescein isothiocyanate-conjugated concanavalin A
(FITC-ConA) for 20 min at 25°C and washed with PBS to remove stain.
Fluorescent images were obtained using BX53 fluorescence microscopy
(Olympus Corporation, Tokyo, Japan).
Crystal violet staining elaborating
the combined effect of BN and VCM on S. aureus biofilms
Bacterial suspension (200 µl) was added to 96-well
plates and cultivated at 37°C and 5% CO2 for 48 h to
mature the biofilm. Plates were gently washed with PBS (3×). BN
(100 µl) with or without VCM was added to each well and incubated
at 37°C in 5% CO2 for 6, 12, 24 or 48 h. The final
concentrations of BN were 1/2, 1 and 2 MIC (16, 32 and 64 µg/ml)
and VCM were 1/2, 1 and 2 MIC (0.25, 0.5 and 1 µg/ml). Plates were
cultivated at 37°C for 6, 12, 24 and 48 h. Plates were washed with
PBS to remove planktonic cells. The biomass of each slice was
air-dried at 37°C for 20 min and stained for 20 min at 25°C with
200 µl filtered 0.25% crystal violet, washed with PBS (3×) to
remove unbound stain, dried for 20 min at 37°C and solubilized in
200 µl 95% ethanol for 15 min at 25°C. A reading at 570 nm using an
ELISA microplate reader was obtained. Experiments were performed
six times.
Fluorescence microscopy elaborating
the combined effect of BN and VCM on S. aureus biofilms
Biofilms were prepared in 24-well plates containing
a polystyrene slice as described. Slices were gently washed with
PBS (2×). BN (100 µl) with or without VCM was added to each well.
Concentrations of BN were 1/2, 1 and 2 MIC (16, 32 and 64 µg/ml)
and VCM were 1/2, 1 and 2 MIC (0.25, 0.5 and 1 µg/ml). Plates were
incubated at 37°C for 6, 12, 24 and 48 h. Slices were washed with
PBS to remove planktonic cells. Biofilms were stained with
FITC-ConA for 20 min at 25°C and washed with PBS to remove stain.
Fluorescent images were captured by fluorescence microscopy.
Experiments were performed in triplicate.
Biofilm bacterial colony-forming unit
(CFU) counts
Bacterial suspension (200 µl) was added to 96-well
plates and cultivated at 37°C and 5% CO2 for 48 h to
mature the biofilms. The mature biofilms were treated with 100 µl
1/2, 1 and 2 MIC BN with or without 1/2, 1 and 2 MIC VCM and
incubated at 37°C for 6, 12, 24 and 48 h. Slices were washed with
PBS to remove planktonic cells and sonicated to disrupt cell
clumps. Biofilm bacterial CFU counts were conducted by plating
serial dilutions on TSB agar. Experiments were performed in
triplicate.
Confocal laser scanning microscopy
(CLSM)
The biofilm was determined using a double live/dead
staining kit (L13152 LIVE/DEAD® BacLight™ Bacterial
Viability kits; Thermo Fisher Scientific, Inc.), containing nucleic
acid stains SYTO9 (green) and propidium iodide (PI, red), which
differ in their ability to penetrate healthy bacterial cells. SYTO9
stain labels live bacteria, whereas PI penetrates bacteria with
damaged membranes. Bacterial suspension (4 ml) was added to the
bottom of a 24-well plate. Following 48 h incubation at 37°C,
plates were washed with PBS and 1/2, 1 and 2 MIC BN with or without
1/2, 1 and 2 MIC VCM was added followed by incubation at 37°C for
48 h. Slices were washed with PBS. According to the manufacturer's
protocol, the slices were stained with SYTO9/PI for 20 min at 25°C
in the dark. A TCS SP5 microscope (Leica Microsystems GmbH,
Wetzlar, Germany) was used to investigate S. aureus biofilms.
Scanning electron microscopy
(SEM)
Polystyrene plates (24×24 mm) were sterilized and
placed into 6-well plates for 48 h to form the biofilm. BN (2 MIC,
64 µg/ml) with or without VCM (2 MIC, 1 µg/ml) were added to each
well, followed by incubation at 37°C for 48 h. Slices were washed
with PBS and fixed at 4°C with 2.5% glutaraldehyde for 8 h.
Surfaces were rinsed three times and fixed with 0.1% osmium
tetraoxide for 1 h. Samples were dehydrated through a graded
ethanol series (30, 50, 70, 80, 90, 95 and 100%) for 15 min each at
room temperature and coated with gold. Images were obtained using
an SU8010 SEM (Hitachi, Ltd., Tokyo, Japan) (25).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis of genes forming
biofilms
Biofilms were prepared and exposed to BN with or
without VCM in 6-well plates as described previously. Total RNA was
extracted using the MiniBEST Universal RNA Extraction kit (Takara
Biotechnology Co., Ltd., Dalian, China) according to the
manufacturer's protocol. Biofilms were centrifuged at 7,104 × g at
4°C for 2 min, 600 µl Buffer RL (Takara Biotechnology Co., Ltd.)
were added and incubated for 2 min at room temperature. The
pyrolysis liquid was transferred into a gDNA Eraser Spin Column and
centrifuged at 13,400 × g at 4°C for 1 min. To the filtrate an
equal amount of 70% ethanol was added. Immediately, the mixture was
transferred into an RNA Spin Column and centrifuged at 13,400 × g
at 4°C for 1 min. Buffer RWA (500 µl) and Buffer RWB (600 µl) were
added to a RNA Spin Column and spun at 13,400 × g at 4°C for 30
sec. RNA pellets were suspended in 100 µl RNase-free water. cDNA
was synthesized using EasyScript® One-Step gDNA Removal
and cDNA Synthesis SuperMix kit (Generon, Slough, UK). RT reaction
mixtures contained total RNA sample (5 µl,) anchored Oligo(dT)18
Primer (0.5 µg/µl; 1 µl), 2× ES Reaction mix (10 µl),
EasyScript® RT/RI Enzyme mix (1 µl), gDNA Remover (1 µl)
and RNase-free water to a final volume of 20 µl. Reverse
transcriptase was inactivated by incubation at 42°C for 15 min,
then 85°C for 5 sec, followed by cooling at 4°C and storage at
−80°C. Primer sequences were as follows: Intercellular adhesion
(ica)A, (189 bp) forward 5′-GCAGTTGTCGATGTTGGCTA-3′ and reverse
5′-TACTTCGTGTCCCCCTTGAG-3′; icaR (186 bp), forward
5′-TACGCCTGAGGAATTTTCTG-3′ and reverse 5′-CCAAATTTTTGCGAAAAGGA-3′;
accessory gene regulator (agr) A (82 bp), forward
5′-ACGTGGCAGTAATTCAGTGTATGTT-3′ and reverse
5′-GGCAATGAGTCTGTGAGATTTTGT-3′; and 16SrRNA (201 bp), forward
5′-TATTTTTCCGGTTGGTCGTC-3′ and reverse 5′-GGCTTAACCTTGCCATCAGA-3′.
qPCR was performed according to the manufacturer's protocol (Takara
Biotechnology Co., Ltd.), in 20 µl reactions, containing SYBR
Premix Ex TaqII (10 µl), forward and reverse primers (0.8 µl), cDNA
(2 µl), ROX Reference Dye II (0.4 µl) and deionized water (6 µl).
An Applied Biosystems 7500 Fast Real-Time PCR system (Applied
Biosystems, Thermo Fisher Scientific, Inc.) was used. Cycling
parameters were as follows: 95°C for 30 sec; 95°C for 5 sec and
60°C for 34 sec for 40 cycles and one melt curve at 95°C for 15
sec, followed by 60°C for 60 sec and 95°C for 15 sec. Experiments
were performed in duplicate and analyzed in triplicate; 16SrRNA was
used as internal control. RT-qPCR data were analyzed using the
relative quantitative (2−ΔΔCq) method
(26).
Statistical analysis
All data were analyzed using SPSS 19.0 (IBM Corp.,
Armonk, NY, USA) and are expressed as the mean ± standard
deviation. Statistical comparisons between groups were made using
one-way analysis of variance followed by Fisher's LSD test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Inhibition of S. aureus by BN
MICs of BN for C-4-4 and ATCC25923 strains were 32
µg/ml. The MIC of VCM for the two strains was 0.5 µg/ml. ATCC25923
was used as control strain; VCM as positive control.
Effects of BN on S. aureus biofilm
formation in vitro
Biofilms were stained with crystal violet and
FITC-ConA to study effects of BN on S. aureus biofilm formation. It
was identified that compared with the control group, varying
treatment times and concentrations of BN had an effect on biofilm
formation, in addition to 6 and 12 h 1/4 MIC BN (P<0.05;
Table II). With an increase in BN
concentration, the effect became more pronounced. The strongest
decreases in OD measurements were observed with 1 MIC BN following
24 and 48 h (Table II).
| Table II.Effect of BN on S. aureus
biofilm formation measured using crystal violet staining. |
Table II.
Effect of BN on S. aureus
biofilm formation measured using crystal violet staining.
|
|
OD570 |
|---|
|
|
|
|---|
| Group | 6 h | 12 h | 24 h | 48 h |
|---|
| Control | 0.274±0.056 | 0.439±0.059 | 0.726±0.119 | 1.133±0.283 |
| 1 MIC VCM |
0.195±0.030a |
0.274±0.053a |
0.356±0.066a |
0.470±0.125a |
| 1/4 MIC BN | 0.244±0.049 | 0.336±0.012 |
0.535±0.099a |
0.672±0.163a |
| 1/2 MIC BN |
0.221±0.028a |
0.308±0.064a |
0.441±0.095a |
0.510±0.108a |
| 1 MIC BN |
0.202±0.033a |
0.281±0.075a |
0.346±0.127a |
0.450±0.121a |
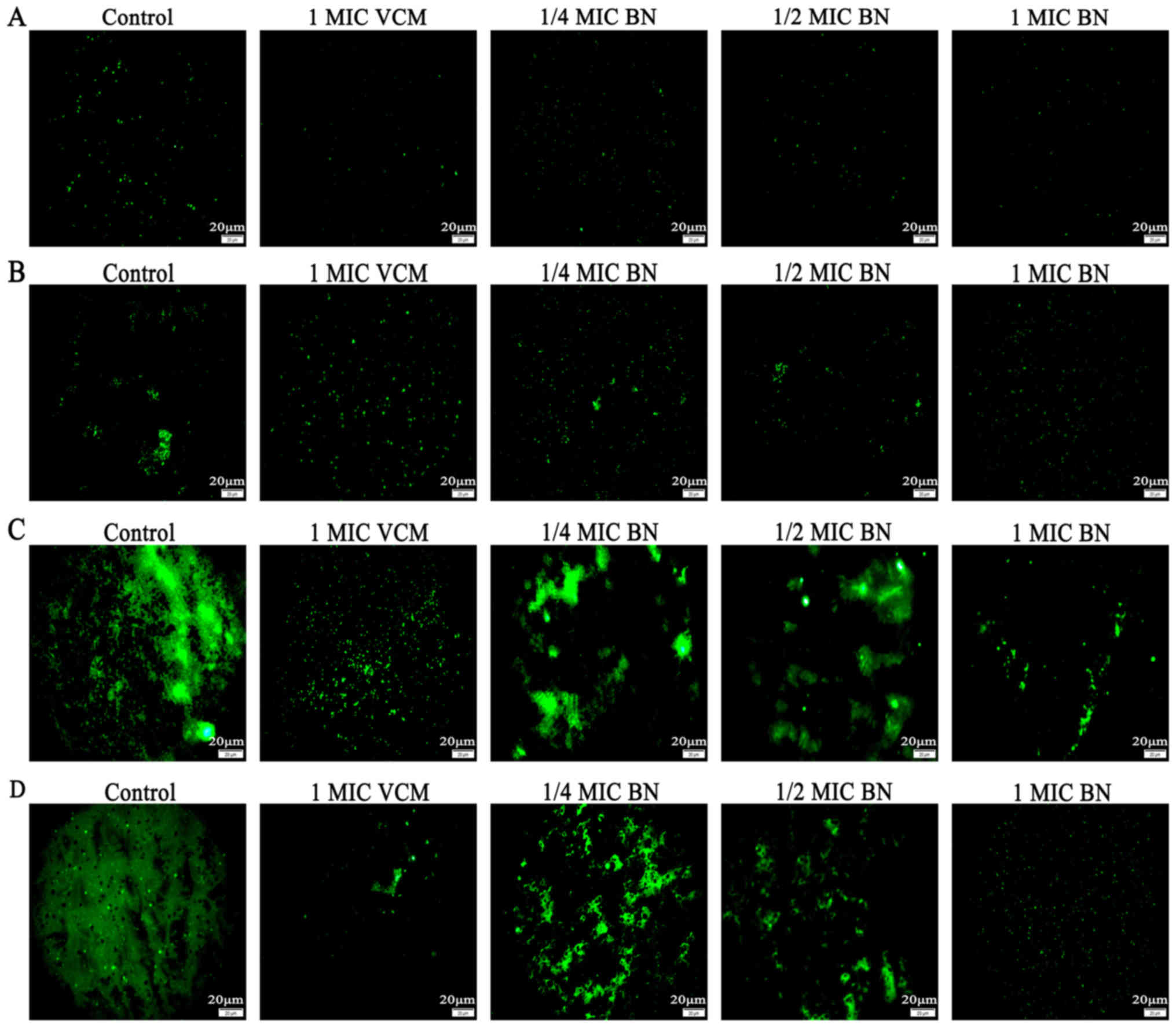
FITC-ConA produces green fluorescence by combining
with polysaccharides in the biofilm. The stronger the biofilm, the
greater the green fluorescence (17). The current study used
FITC-ConA-tagged fluorescence microscopy imaging to evaluate
effects of BN on biofilm formation. The results revealed that 1/4,
1/2 and 1 MIC BN for 12, 24 and 48 h inhibited biofilm formation,
whereas only minor effects were observed at 6 h (Fig. 1). At 48 h, the effect of 1 MIC BN was
most pronounced.
BN combined with VCM damages the
biofilm
The current study aimed to compare damaging effects
of BN on S. aureus biofilms in the presence and absence of VCM, in
order to assess synergistic effects. According to the crystal
violet staining results, VCM alone only eradicated biofilms at 2
MIC following 48 h. BN and BN + VCM reduced biofilm formation in a
concentration-dependent manner (Table
III).
| Table III.Effect of BN combined with VCM on
S aureus biofilms analyzed using crystal violet
staining. |
Table III.
Effect of BN combined with VCM on
S aureus biofilms analyzed using crystal violet
staining.
|
|
OD570 |
|---|
|
|
|
|---|
| Group | 6 h | 12 h | 24 h | 48 h |
|---|
| Blank | 1.224±0.246 | 1.291±0253 | 1.226±0.194 | 1.283±0.272 |
| 1/2 MIC VCM | 1.197±0.262 | 1.269±0.182 | 1.168±0.234 | 1.156±0.154 |
| 1 MIC VCM | 1.211±0.287 | 1.264±0.267 | 1.151±0.163 | 1.112±0.116 |
| 2 MIC VCM | 1.140±0.296 | 1.224±0.311 | 1.105±0.121 |
1.041±0.136a |
| 1/2 MIC BN | 1.211±0.291 | 1.021±0.180 |
0.961±0.067a |
0.882±0.121a |
| 1 MIC BN | 1.112±0.287 | 1.048±0.196 |
0.904±0.127a,c |
0.810±0.134a,c |
| 2 MIC BN |
0.965±0.134a |
0.965±0.188a |
0.877±0.176a,c |
0.711±0.201a,c |
| 1/2 MIC VCM+1/2 MIC
BN |
0.796±0.060a |
0.916±0.160a |
0.854±0.121a,c |
0.747±0.146a,c |
| 1/2 MIC VCM+1 MIC
BN |
0.945±0.149a,c |
0.880±0.075a,c |
0.802±0.876a,c |
0.678±0.173a,c |
| 1/2 MIC VCM+2 MIC
BN |
0.903±0.159a,c |
0.857±0.160a,c |
0.772±0.169a,c |
0.644±0.170a,c |
| 1 MIC VCM+1/2 MIC
BN |
0.882±0.148a,c |
0.833±0.211a,c |
0.757±0.120a,c |
0.584±0.214a–c |
| 1 MIC VCM+1 MIC
BN |
0.863±0.219a–c |
0.805±0.177a–c |
0.720±0.170a–c |
0.526±0.222a–c |
| 1 MIC VCM+2 MIC
BN |
0.833±0.199a–c |
0.816±0.070a–c |
0.654±0.182a–c |
0.498±0.144a–c |
| 2 MIC VCM+1/2 MIC
BN |
0.831±0.150a–c |
0.776±0.093a–c |
0.597±0.138a–c |
0.469±0.158a–c |
| 2 MIC VCM+1 MIC
BN |
0.768±0.149a–c |
0.732±0.157a–c |
0.553±0.223a–c |
0.404±0.171a–c |
| 2 MIC VCM+2 MIC
BN |
0.756±0.112a–c |
0.645±0.116a–c |
0.507±0.209a–c |
0.390±0.160a–c |
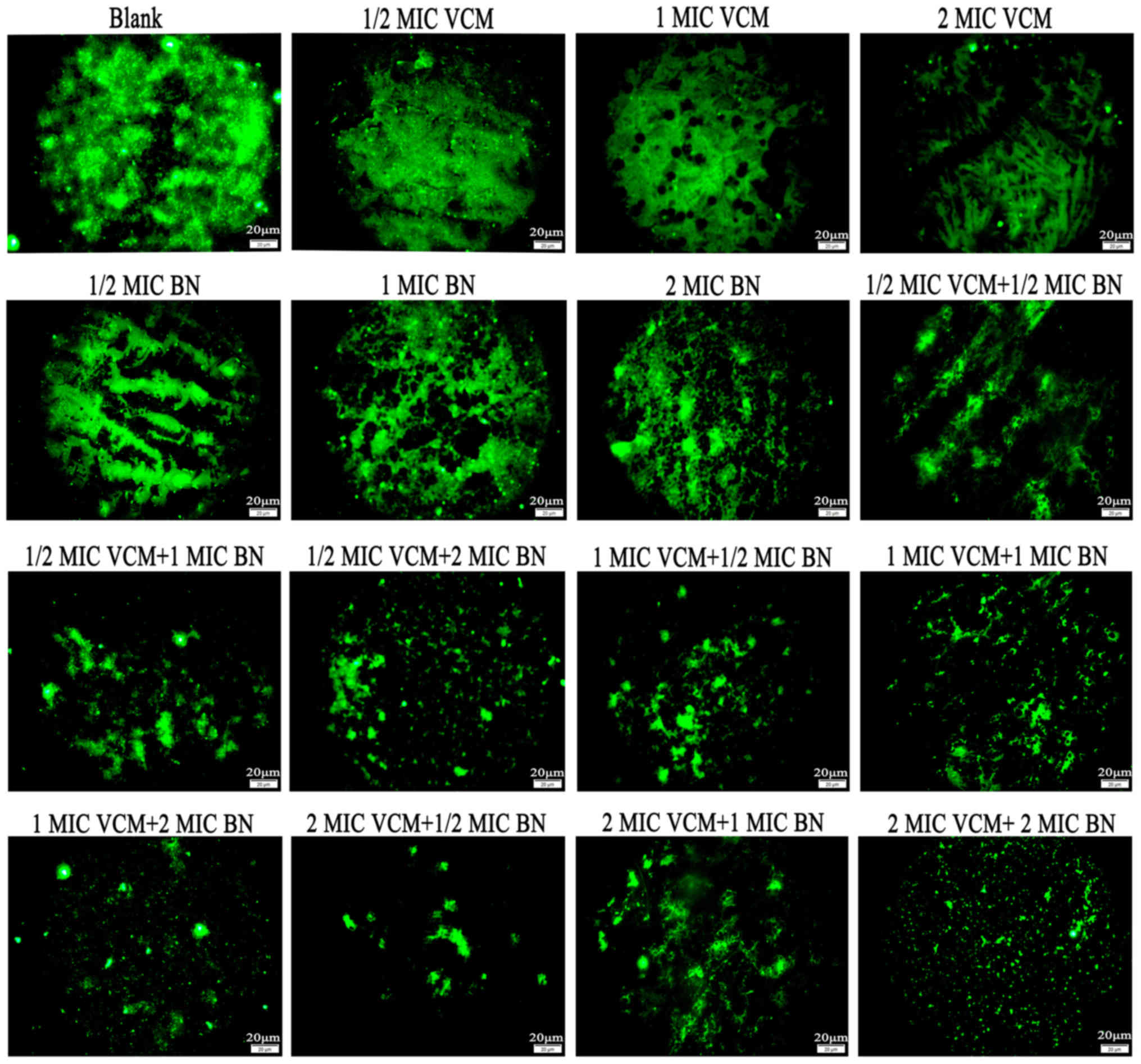
Fluorescence microscopy demonstrated that VCM had no
marked effect on S. aureus biofilms compared with the
control group, which exhibited large areas of green fluorescence.
With an increase in BN concentration, mature biofilms were damaged;
with increasing concentrations of BN + VCM, decreasing green
fluorescence was observed (Fig.
2).
Biofilm bacterial CFU counts
VCM alone only reduced bacterial counts at 2 MIC
following 48 h treatment. BN at 1 or 2 MIC for 6 and 12 h and at
1/2, 1 or 2 MIC following 48 h was able to decrease bacterial
counts. The most distinct effect was observed with 2MIC BN + 2MIC
VCM at 48 h (Table IV).
| Table IV.Influence of BN and VCM of bacteria
counts. |
Table IV.
Influence of BN and VCM of bacteria
counts.
|
| ×106
colony-forming unit/ml |
|---|
|
|
|
|---|
| Group | 6 h | 12 h | 24 h | 48 h |
|---|
| Blank | 256.667±24.502 | 255.000±27.074 | 256.333±14.503 | 256.333±13.204 |
| 1/2 MIC VCM | 239.667±27.502 | 246.333±30.534 | 248.667±18.148 | 241.333±21.221 |
| 1 MIC VCM | 241.667±10.017 | 242.333±30.616 | 239.000±13.454 | 233.333±31.390 |
| 2 MIC VCM | 239.667±23.029 | 238.000±19.519 | 236.333±8.622 |
199.000±19.313a |
| 1/2 MIC BN | 229.333±16.563 | 226.667±23.007 |
215.333±12.897a |
198.333±5.695a |
| 1 MIC BN | 223.000±11.136 |
216.667±8.737a |
206.333±7.506a,c |
168.667±12.583a,c |
| 2 MIC BN |
196.333±16.503a,c |
194.000±31.241a,c |
180.333±14.048a–c |
116.000±14.799a–c |
| 1/2 MIC VCM+1/2 MIC
BN |
200.667±2.517a,c |
192.000±17.692a,c |
182.000±19.313a–c |
113.000±17.692a–c |
| 1/2 MIC VCM+1 MIC
BN |
184.667±26.764a–c |
170.000±33.151a–c |
154.000±13.454a–c |
109.667±9.609a–c |
| 1/2 MIC VCM+2 MIC
BN |
176.667±15.011a–c |
163.000±26.665a–c |
144.667±13.577a–c |
100.667±13.317a–c |
| 1 MIC VCM+1/2 MIC
BN |
177.000±10.583a–c |
156.000±12.530a–c |
137.000±13.454a–c |
95.667±9.074a–c |
| 1 MIC VCM+1 MIC
BN |
168.667±20.550a–c |
149.667±17.474a–c |
128.667±16.258a–c |
84.333±10.408a–c |
| 1 MIC VCM+2 MIC
BN |
146.333±23.029a–c |
143.000±23.516a–c |
111.333±15.275a–c |
70.333±15.308a–c |
| 2 MIC VCM+1/2 MIC
BN |
141.667±35.726a–c |
135.667±11.930a–c |
104.333±13.868a–c |
61.000±10.536a–c |
| 2 MIC VCM+1 MIC
BN |
140.333±23.116a–c |
127.000±5.292a–c |
96.000±7.937a–c |
56.33±9.866a–c |
| 2 MIC VCM+2 MIC
BN |
135.667±29.143a–c |
119.000±17.578a–c |
83.000±16.371a–c |
47.333±11.604a–c |
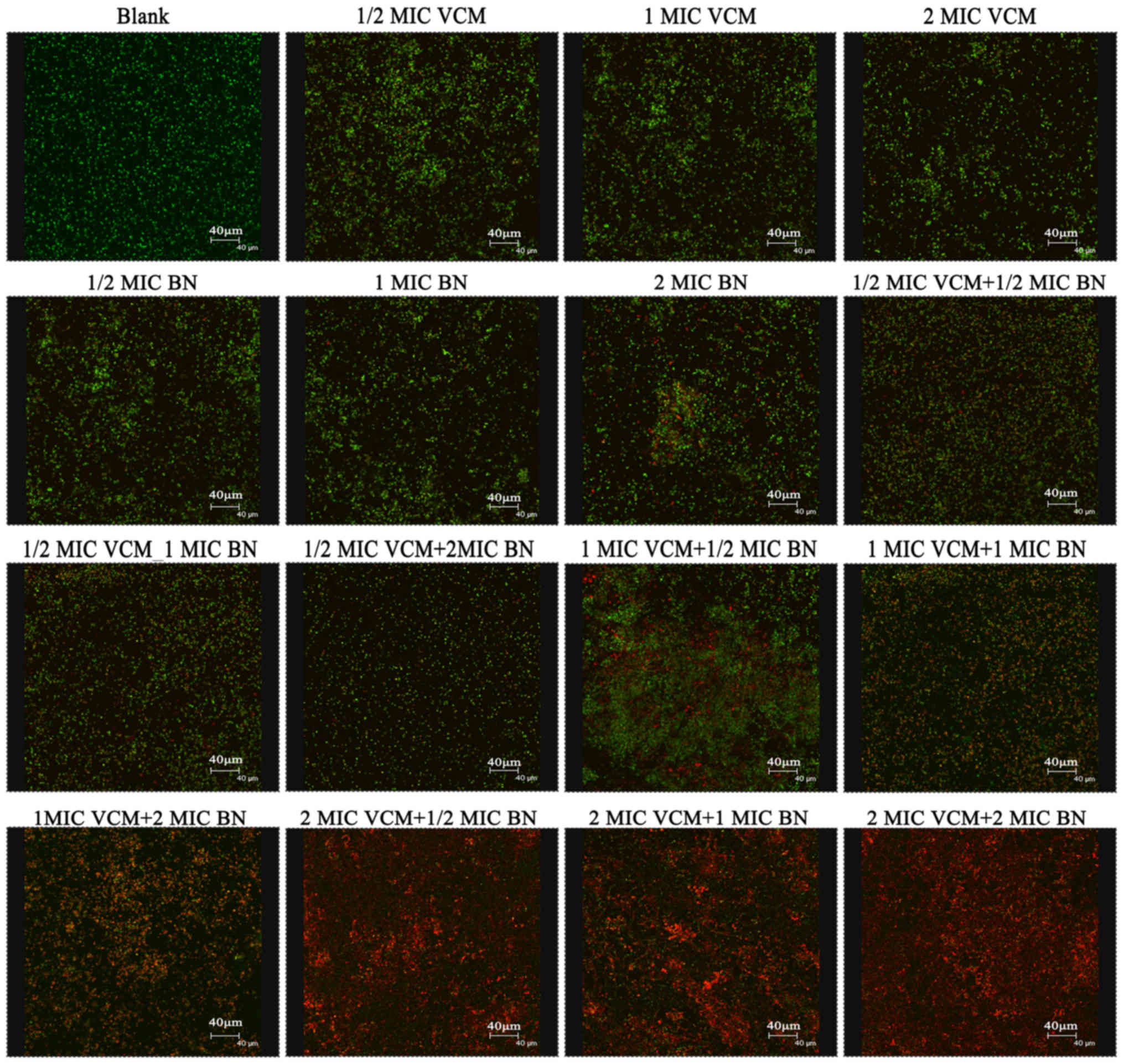
Biofilm analysis using CLSM
SYTO9 stain labels live bacteria, whereas PI
penetrates bacteria with damaged membranes. The results
demonstrated that VCM was unable to damage the biofilm (Fig. 3). VCM groups exhibited mainly green
fluorescence, representing live cells. Increasing concentrations of
BN demonstrated an effect on the biofilm, exhibiting increasing red
fluorescence, representing dead cells. BN + VCM groups exhibited
increased levels of dead cells, represented by red
fluorescence.
Biofilm damage assay by SEM
It was observed that biofilms treated with VCM
exhibited large extracellular matrices and a large biofilm mass
compared with the control group. BN was able to reduce the
extracellular matrix and BN + VCM groups exhibited increased
effectiveness, illustrated by damaged biofilms and individual
bacteria (Fig. 4). These findings
indicate that BN may be able to penetrate biofilms.
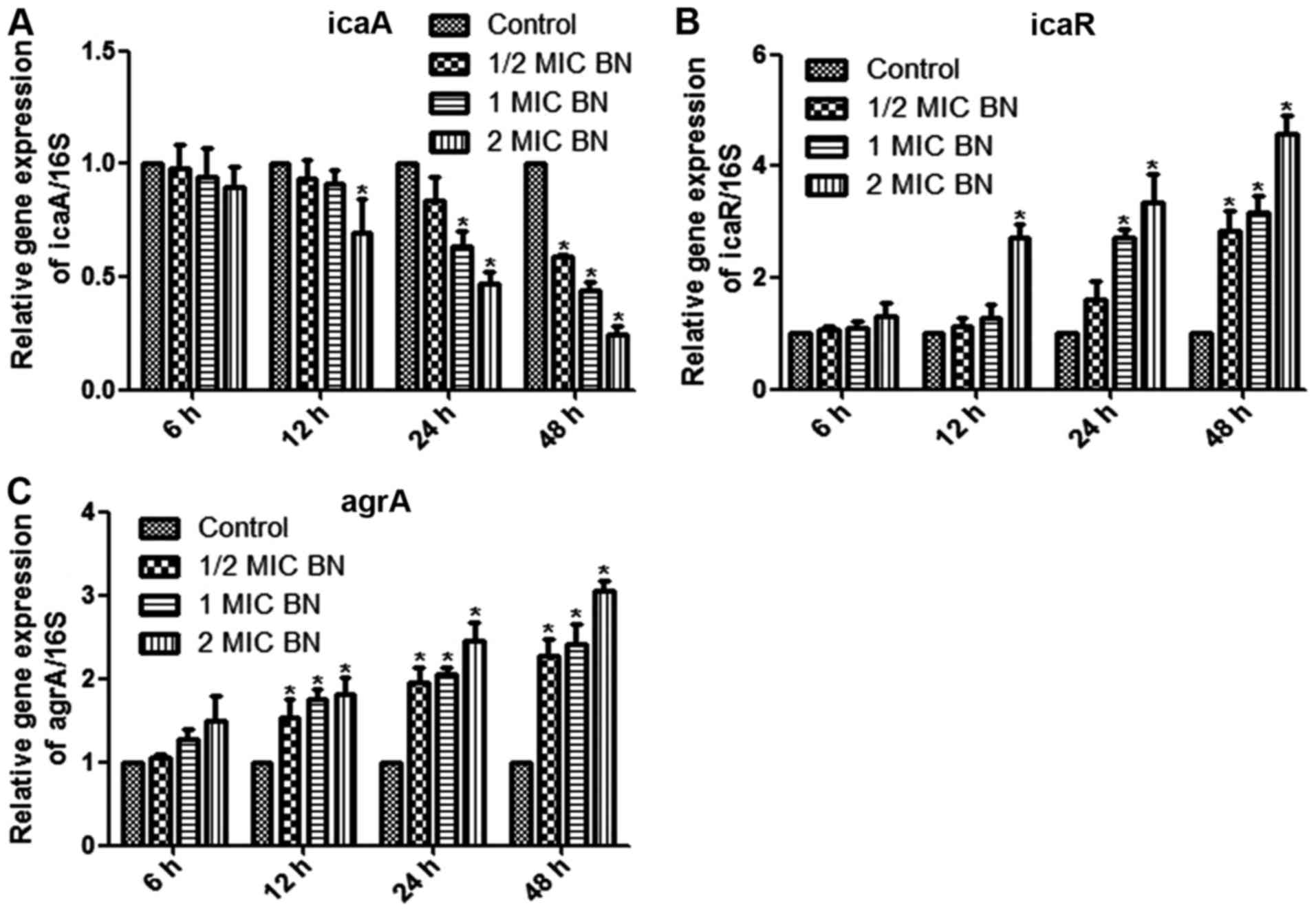
Gene expression analysis
The current study aimed to measure mRNA levels in
different samples using RT-qPCR, with 16SrRNA as internal control.
Mature biofilms were treated with 1/2, 1 and 2 MIC BN for 6, 12, 24
and 48 h. Treatment with 2 MIC BN for 12 h, with 1 or 2 MIC BN for
24 h or with 1/2, 1 or 2 MIC BN for 48 h significantly lowered icaA
expression levels compared with the untreated control (P<0.05;
Fig 5A). Transcription levels of
icaR and agrA were significantly upregulated in BN treatment
groups, compared with the control (P<0.05; Fig. 5B and C). IcaR expression
significantly increased compared with the control using 2 MIC BN
for 12 h, 1 or 2 MIC BN for 24 h or 1/2, 1 or 2 MIC BN for 48 h
(P<0.05; Fig. 5B). BN treatment
significantly upregulated transcription levels of agrA following
treatment times ≥12 h (Fig. 5C). No
significant difference was observed following BN treatment for 6 h
compared with the control.
Discussion
S. aureus biofilm development primarily
includes four stages: Attachment, multiplication, maturation and
dispersal (27). The principal
mechanism of biofilm formation relies on secretion of extracellular
polysaccharides (28). It was
suggested that the matrix of biofilms may be responsible for
increased resistance to antibiotics by acting as a diffusion
barrier, causing chronic persistent infections that are difficult
to cure (29,30). At present, it is of great importance
to identify methods for prevention and treatment of
biofilm-associated infections. Studies have demonstrated that
traditional herbal medicines were widely-sourced, had minimal side
effects, strong pharmacological action and did not lead to
bacterial drug resistance (15–17). The
use of herbal medicines in the prevention and treatment of biofilm
infections has a particular advantage. Traditional herbal medicine
may not only inhibit the formation and destroy biofilms, but may
enhance resistance to infections when combined with antibiotics
(18).
The aim of the current study was to inhibit S.
aureus biofilm formation, as assessed using crystal violet
staining and FITC-ConA fluorescence microscopy. VCM was used as
positive control. The results of the two experiments were similar:
BN inhibited biofilm formation in vitro in a dose-dependent
manner and the greatest effect was obtained with 2 MIC BN following
24 and 48 h. The results were similar to those reported by
Lázaro-Díez et al (6).
At present, VCM is the most commonly administered
drug for S. aureus biofilm-associated infections (31). However, there is cause for concern
due to recent developments of VCM-intermediate S. aureus and
VRSA strains (10). The question of
how to reduce the generation of drug-resistant strains and improve
the effectiveness of VCM in biofilm infections requires further
research. Combining Chinese herbal medicines with VCM is a possible
novel approach to treating biofilm-associated infections.
In the current study, to identify synergistic
effects of BN combined with VCM on S. aureus biofilms,
crystal violet staining, fluorescence microscopy, CFU counting,
CLSM and SEM were used. This array of methods demonstrated that BN
and BN + VCM generally enhanced the destruction of biofilms in a
concentration-dependent manner; 2 MIC BN + 2 MIC VCM exhibited the
most notable effects, whereas VCM alone was ineffective. Chen et
al (32) studied inhibitory
effects of baicalein on S. aureus biofilm formation using
these methods, achieving outcomes similar to experimental results
presented here.
In summary, VCM may not disrupt a biofilm when used
alone, whereas BN may be able to do so. BN may reduce the
production of the extracellular polymeric matrix or it may
penetrate the biofilm to increase its permeability to VCM. In the
current study, BN and VCM exhibited a synergy in eradicating the
biofilm.
S. aureus biofilm formation is a complex
dynamic process, involving numerous regulatory mechanisms,
including polysaccharide intercellular adhesion (PIA),
extracellular DNA, global regulatory factors and the QS system
(28). PIA is synthesized by the
ica locus and was first identified in the epidermis S.
aureus biofilm (33). The
ica locus includes icaR (regulatory) and icaADBC
(biosynthetic), which affect biofilm formation (34). PIA is the primary component of the
Gram-positive bacterial biofilm matrix, has the largest influence
on biofilm formation and is a well-studied component (35). Studies have demonstrated that icaR
acts as a repressor gene and PIA generation increases following
knockout of icaR (34,36). In addition, PIA expression is
repressed by tcaR, a transcriptional regulator of the
teicoplanin-associated locus. The protein regulator of biofilm
formation augments ica gene expression, PIA production and
biofilm formation (37–39). In addition, Pamp et al
(40) demonstrated that Spx, a
global regulator of stress response genes, inhibits biofilm
formation, potentially by modulating icaR. When strains were
cultured in anaerobic environments, the S. aureus
respiratory response regulator SrrAB was responsible for PIA,
promoting biofilm formation (41).
Regulation of the ica locus influences production of PIA and
formation of biofilms, making it a novel potential therapeutic
target.
In the current study, mature biofilms were treated
with BN for 6, 12, 24 and 48 h. It was observed that BN was able to
downregulate icaA expression. Transcription levels of icaR were
significantly upregulated. BN may regulate icaA and icaR expression
in order to inhibit the synthesis of PIA and to reduce the
secretion of extracellular polysaccharides.
The S. aureus biofilm QS system includes the
agr system and luxS/AI-2 (42). The
agr QS system has been well-researched and includes agrBDCA and
RNAIII (43). The autoinducer aryl
hydrocarbon receptor-interacting protein (AIP) is generated from
ArgD precursors and is secreted via the AgrB membrane protein
(43). When the extracellular AIP
concentration reaches a threshold, agrC is phosphorylated. AgrC
phosphorylation activates molecular agrA, which in turn activates
RNAIII (44). A previous study
demonstrated that when the agr system is mutated, biofilm formation
is enhanced (45). The agr system
promotes biofilm maturation. The addition of AIP to the mature
biofilm may additionally promote biofilm dispersal (45).
The current study demonstrated that BN was able to
upregulate transcription levels of agrA. BN may inhibit gene
expression of the QS signaling molecule agrA and block RNAIII to
prevent biofilm formation. However, the question of whether BN
inhibits the QS system requires further analysis.
In conclusion, BN was able to damage mature biofilms
potentially by enhancing VCM permeability and through potential
inhibition of the secretion of extracellular polysaccharides and
the QS system. BN is a potential novel treatment against S.
aureus biofilm-associated infections. The mechanisms involved
require further analysis and experimentation in vivo.
Acknowledgements
The authors would like to thank Dr Xun Min, Dr Tao
Zhang and Dr Yingbiao Tian from the Zunyi Medical University
(Zunyi, China) for their expert advice and technical assistance
throughout the study.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZHC, XM, YBT and TZ conceived and designed the
study. DP, ALC, XLC and BS performed the experiments. DP, ZLD, ALC,
XLC and HY analyzed the data. DP, ZLD, XLC and YBT contributed
reagents, materials and analysis tools. DP and ZHC wrote the paper.
TZ revised the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Affiliated Hospital of Zunyi Medical University
and written informed consent was obtained prior to the collection
of all samples.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yu L, Hisatsune J, Hayashi I, Tatsukawa N,
Sato'o Y, Mizumachi E, Kato F, Hirakawa H, Pier GB and Sugaia M: A
Novel Repressor of the ica locus discovered in clinically isolated
super-biofilm-elaborating Staphylococcus aureus. MBio.
8:pii:e02282. –16. 2017. View Article : Google Scholar
|
|
2
|
Yoshii Y, Okuda KI, Yamada S, Nagakura M,
Sugimoto S, Nagano T, Okabe T, Kojima H, Iwamoto T, Kuwano K and
Mizunoe Y: Norgestimate inhibits staphylococcal biofilm formation
and resensitizes methicillin-resistant Staphylococcus aureus
to β-lactam antibiotics. NPJ Biofilms Microbiomes. 3:182017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sollid JU, Furberg AS, Hanssen AM and
Johannessen M: Staphylococcus aureus: Determinants of human
carriage. Infect Genet Evol. 21:531–541. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kong C, Chee CF, Richter K, Thomas N,
Rahman Abd N and Nathan S: Suppression of Staphylococcus
aureus biofilm formation and virulence by a benzimidazole
derivative, UM-C162. Sci Rep. 8:27582018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Waryah CB, Wells K, Ulluwishewa D,
Chen-Tan N, Gogoi-Tiwari J, Ravensdale J, Costantino P, Gökçen A,
Vilcinskas A, Wiesner J and Mukkur T: In vitro antimicrobial
efficacy of tobramycin against Staphylococcus aureus
biofilms in combination with or without DNase I and/or Dispersin B:
A preliminary investigation. Microb Drug Resist. 23:384–390. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lázaro-Díez M, Remuzgo-Martínez S,
Rodríguez-Mirones C, Acosta F, Icardo JM, Martínez-Martínez L and
Ramos-Vivas J: Effects of subinhibitory concentrations of
ceftaroline on methicillin-resistant Staphylococcus aureus
(MRSA) biofilms. PLoS One. 11:e01475692016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Feuillie C, Formosa-Dague C, Hays LM,
Vervaecka O, Derclayea S, Brennanc MP, Fosterb TJ, Geogheganb JA
and Dufrêne YF: Molecular interactions and inhibition of the
staphylococcal biofilm-forming protein SdrC. Proc Natl Acad Sci
USA. 114:3738–3743. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Howlin RP, Brayford MJ, Webb JS, Cooper
JJ, Aiken SS and Stoodley P: Antibiotic-loaded synthetic Calcium
sulfate beads for prevention of bacterial colonization and biofilm
formation in periprosthetic infections. Antimicrob Agents
Chemother. 59:111–120. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Koch G, Yepes A, Förstner KU, Wermser C,
Stengel ST, Modamio J, Ohlsen K, Foster KR and Lopez D: Evolution
of resistance to a last-resort antibiotic in Staphylococcus
aureus via bacterial competition. Cell. 158:1060–1071. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Howden BP, Davies JK, Johnson PD, Stinear
TP and Grayson ML: Reduced vancomycin susceptibility in
Staphylococcus aureus, including vancomycin-intermediate and
heterogeneous vancomycin-intermediate strains: Resistance
mechanisms, laboratory detection, and clinical implications. Clin
Microbiol Rev. 23:99–139. 2016. View Article : Google Scholar
|
|
11
|
Kiedrowski MR and Horswill AR: New
approaches for treating staphylococcal biofilm infections. Ann N Y
Acad Sci. 1241:104–121. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bhattacharya M, Wozniak DJ, Stoodley P and
Hall-Stoodley L: Prevention and treatment of Staphylococcus
aureus biofilms. Expert Rev Anti Infect Ther. 13:1499–1516.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Salem AH, Elkhatib WF and Noreddin AM:
Pharmacodynamic assessment of vancomycin-rifampicin combination
against methicillin resistant Staphylococcus aureus biofilm:
A parametric response surface analysis. J Pharma Pharmacol.
63:73–79. 2011. View Article : Google Scholar
|
|
14
|
Olson ME, Slater SR, Rupp ME and Fey PD:
Rifampicin enhances activity of daptomycin and vancomycin against
both a polysaccharide intercellular adhesin (PIA)-dependent and
-independent Staphylococcus epidermidis biofilm. J
Antimicrob Chemother. 65:2164–2171. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fontaine BM, Nelson K, Lyles JT, Jariwala
PB, García-Rodriguez JM, Quave CL and Weinert EE: Identification of
ellagic acid rhamnoside as a bioactive component of a complex
botanical extract with anti-biofilm activity. Front Microbiol.
8:4962017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fu B, Wu Q, Dang M, Bai D, Guo Q, Shen L
and Duan K: Inhibition of pseudomonas aeruginosa biofilm formation
by traditional chinese medicinal herb herba patriniae. Biomed Res
Int. 2017:95847032017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Luo J, Dong B, Wang K, Cai S, Liu T, Cheng
X, Lei D and Chen Y, Li Y, Kong J and Chen Y: Baicalin inhibits
biofilm formation, attenuates the quorum sensing-controlled
virulence and enhances Pseudomonas aeruginosa clearance in a mouse
peritoneal implant infection model. PLoS One. 12:e01768832017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
García-Heredia A, García S,
Merino-Mascorro JÁ, Feng P and Heredia N: Natural plant products
inhibits growth and alters the swarming motility, biofilm
formation, and expression of virulence genes in enteroaggregative
and enterohemorrhagic Escherichia coli. Food Microbiol. 59:124–132.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim J, Lee HK, Chang TS, Kang KS and Hwang
GS: Inhibitory effect of brazilin on osteoclast differentiation and
its mechanism of action. Int Immunopharmacol. 29:628–634. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao XJ, Wang TC, Zhang ZC, Cao YG, Zhang
NS and Guo MY: Brazilin plays an anti-inflammatory role with
regulating Toll-like receptor 2 and TLR 2 downstream pathways in
Staphylococcus aureus-induced mastitis in mice. Int
Immunopharmacol. 27:130–137. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dan P, Zhou X and Chen Z: Research
progress on mechanism and treatment of Staphylococcus aureus
biofilms. Molecular Biomedicine. 23:3745–3749. 2017.
|
|
22
|
Nirmal NP and Panichayupakaranant P:
Antioxidant, antibacterial, and anti-inflammatory activities of
standardized brazilin-rich Caesalpinia sappan extract. Pharm Biol.
53:1339–1343. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yan Y, Chen YC, Lin YH, Guo J, Niu ZR, Li
L, Wang SB, Fang LH and Du GH: Brazilin isolated from the heartwood
of Caesalpinia sappan L induces endothelium-dependent
and-independent relaxation of rat aortic rings. Acta Pharmacol Sin.
36:1318–1326. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Clinical and Laboratory Standards
Institute: Methods for dilution antimicrobial susceptibility tests
for bacteria that grow aerobically; approved standard, 9th ed CLSI
document M7-A9. Clinical and Laboratory Standards Institute; Wayne,
PA: 2012
|
|
25
|
Shi SF, Jia JF, Guo XK, Zhao YP, Chen DS,
Guo YY and Zhang XL: Reduced Staphylococcus aureus biofilm
formation in the presence of chitosan-coated iron oxide
nanoparticles. Int J Nanomedicine. 11:6499–6506. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Balasubramanian S, Othman EM, Kampik D,
Stopper H, Hentschel U, Ziebuhr1 W, Oelschlaeger1 TA and
Abdelmohsen UR: Marine sponge-derived Streptomyces sp. SBT343
extract inhibits Staphylococcal biofilm formation. Front Microbiol.
8:2362017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sugimoto S, Sato F, Miyakawa R, Chiba A,
Onodera S, Hori S and Mizunoe Y: Broad impact of extracellular DNA
on biofilm formation by clinically isolated Methicillin-resistant
and-sensitive strains of Staphylococcus aureus. Sci Rep.
8:22542018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Singh AK, Prakash P, Achra A, Singh GP,
Das A and Singh RK: Standardization and classification of in
vitro biofilm formation by clinical isolates of
Staphylococcus aureus. J Glob Infect Dis. 9:93–101. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Brooks JL and Jefferson KK: Staphylococcal
biofilms: Quest for the magic bullet. Adv Appl Microbiol. 81:63–87.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
van Hal SJ, Lodise TP and Paterson DL: The
clinical significance of vancomycin minimum inhibitory
concentration in Staphylococcus aureus infections: A
systematic review and meta-analysis. Clin Infect Dis. 54:755–771.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen Y, Liu T, Wang K, Hou C, Cai S, Huang
Y, Du Z, Huang H, Kong J and Chen Y: Baicalein inhibits
Staphylococcus aureus biofilm formation and the quorum
sensing system in vitro. PLoS One. 11:e01534682016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
O'gara JP: Ica and beyond: Biofilm
mechanisms and regulation in Staphylococcus epidermidis and
Staphylococcus aureus. FEMS Microbiol Lett. 270:179–188.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Figueiredo AMS, Ferreira FA, Beltrame CO
and Côrtes MF: The role of biofilms in persistent infections and
factors involved in ica-independent biofilm development and gene
regulation in Staphylococcus aureus. Crit Rev Microbiol.
43:602–620. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ma R, Qiu S, Jiang Q, Sun H, Xue T, Cai G
and Sun B: AI-2 quorum sensing negatively regulates rbf expression
and biofilm formation in Staphylococcus aureus. Int J Med
Microbiol. 307:257–267. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gupta A and Mishra S, Singh S and Mishra
S: Prevention of IcaA regulated poly N-acetyl glucosamine formation
in Staphylococcus aureus biofilm through new-drug like
inhibitors: In silico approach and MD simulation study. Microb
Pathog. 110:659–669. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Skurnik D, Cywes-Bentley C and Pier GB:
The exceptionally broad-based potential of active and passive
vaccination targeting the conserved microbial surface
polysaccharide PNAG. Expert Rev Vaccines. 15:1041–1053. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Formosa-Dague C, Feuillie C, Beaussart A,
Derclaye S, Kucharíková S, Lasa I, Van Dijck P and Dufrêne YF:
Sticky matrix: Adhesion mechanism of the staphylococcal
polysaccharide intercellular adhesin. ACS Nano. 10:3443–3452. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Doulgeraki AI, Di Ciccio P, Ianieri A and
Nychas GE: Methicillin-resistant food-related Staphylococcus
aureus: A review of current knowledge and biofilm formation for
future studies and applications. Res Microbiol. 168:1–15. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pamp SJ, Frees D, Engelmann S, Hecker M
and Ingmer H: Spx is a global effector impacting stress tolerance
and biofilm formation in Staphylococcus aureus. J Bacteriol.
188:4861–4870. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kinkel TL, Roux CM, Dunman PM and Fang FC:
The Staphylococcus aureus SrrAB two-component system
promotes resistance to nitrosative stress and hypoxia. MBio.
4:e00696–e13. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Coelho LR, Souza RR, Ferreira FA,
Guimarães MA, Ferreira-Carvalho BT and Figueiredo AM: Agr RNAIII
divergently regulates glucose-induced biofilm formation in clinical
isolates of Staphylococcus aureus. Microbiology.
154:3480–3490. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Atwood DN, Beenken KE, Loughran AJ, Meeker
DG, Lantz TL, Graham JW, Spencer HJ and Smeltzer MS: XerC
contributes to diverse forms of Staphylococcus aureus
infection via agr-dependent and agr-independent pathway. Infect
Immun. 84:1214–1225. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Le KY and Otto M: Quorum-sensing
regulation in staphylococci-an overview. Front Microbiol.
6:11742015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Boles BR and Horswill AR: Agr-mediated
dispersal of Staphylococcus aureus biofilms. PLoS Pathog.
4:e10000522008. View Article : Google Scholar : PubMed/NCBI
|