Introduction
Alcoholic liver disease (ALD), characterized by
liver metabolic disorder, is an acute, chronic liver injury caused
by long-term heavy drinking (1). The
incidence of ALD has steadily increased in the last 10 years and it
is now the second major cause of liver damage, secondary only to
viral hepatitis (2). Acetaldehyde,
the toxic product of alcohol metabolism, is the primary cause of
ALD (3). According to previous
studies, the interleukin-1β (IL-1β) signaling pathway serves a role
in lipid hepatitis, oxidative stress injury and apoptosis caused by
chronic alcohol intake (4); its role
is correlated with activation of the nucleotide-binding
oligomerization domain, leucine rich repeat containing family,
pyrin domain containing 3 (NLRP3) inflammasome (5).
NLRP3 has previously been reported as a key mediator
of pro-inflammatory cytokine release (6). A number of studies have demonstrated
that NLRP3 may also activate hepatic stellate cells and increase
the severity of liver fibrosis; these mechanisms may be associated
with the upregulation of transforming growth factor-β1 (TGF-β1)
expression (7). Previous reports
have confirmed that NLRP3 upregulates the expression of caspase-1,
which mediates the transformation of pro-IL-1β into IL-1β (8,9). It has
also been reported that triiodothyronine (T3) alleviates oxidative
stress, inflammation and apoptosis caused by hepatic
ischemia-reperfusion injury, and its mechanism of action is
associated with negative regulation of the NLRP3 signaling pathway
(10). Furthermore, Lycium
barbarum has been reported to inhibit the NLRP3 inflammasome
signaling pathway, thereby reducing alcohol-induced liver cell
damage (11). Previous studies have
confirmed that changes in thyroxine levels are associated with the
severity of liver damage (10,12).
Serum and free T3 concentrations gradually decrease as the severity
of liver disease increases, while their concentrations gradually
increase as liver function improves following treatment (10). Serum thyroxine levels are generally
low in patients with non-alcoholic hepatitis and are improved
following the application of thyroxine receptor agonists (1). However, the mechanism of action remains
unclear. It was revealed that liver function was significantly
improved in patients with alcoholic cirrhosis following T3
treatment (2,12). To study the underlying mechanism, the
following animal experimental study was performed.
Materials and methods
Patients and treatments
A total of 10 patients (male: female, 7:3; mean age,
50±4.5 years) with liver cirrhosis admitted to Jining First
People's Hospital (Jining, China) were randomly selected from March
to September 2016. The inclusion criteria were as follows: i)
Compliance with the diagnostic criteria of alcoholic cirrhosis
(13); ii) good medication
compliance; iii) no serious complications. Exclusion criteria: i)
liver cirrhosis caused by hepatitis B virus (HBV) or lithiase; ii)
patients with contraindication of T3; iii) patients with thyroid
diseases that affect this experiment. Blood samples were collected
from median vein of elbow prior the treatment and alanine
transaminase (ALT) and aspartate transaminase (AST) levels was
measured. Patients were treated with T3 (0.1 mg/kg/day, orally
before eating) for 2 weeks. Further blood samples were collected
and the ALT/AST content was measured again.
Animals and treatments
A total of 40 male C57/BL6 (age, 8 weeks; weight,
20–25 g) mice were purchased from Beijing Vital River Laboratory
Animal Technology Co., Ltd. (Beijing, China), housed in a
temperature controlled room (21±2°C), relative humidity 40–60%, on
a 12 h light/dark cycle and provided with standard mice chow and
water. Mice had free access to water and food. Mice were randomly
divided into four groups: Control (n=10), ALD (n=10), ALD+T3 (n=10)
and ALD+T3+ AMP-activated protein kinase inhibitor (CC; n=10). CC
is a commonly used inhibitor of the AMP-associated pathway
(9). With the exception of the
control group, all mice were treated with alcohol by gavage (4
g/kg/day) for 4 weeks. Mice in the control group were treated with
saline of the same volume. Afterwards, the ALD+T3 group was also
treated with T3 by gavage (0.1 mg/kg/day) for 1 week, while the
ALD+T3+CC group was treated with T3 (0.1 mg/kg/day) and CC (10
mg/kg/day) by gavage for 1 week. Mice in the control and ALD groups
were administered with the same amount of normal saline by gavage
for 1 week. Following the successful establishment of the ALD
model, mice were anesthetized with 10% chloral hydrate (0.01 ml/g)
and euthanized by asphyxiation. In brief, anesthetized mice were
placed in a sealed chamber and a CO2 input device was
connected. CO2 was continuously injected at a flow rate
of 30% volume per minute until the CO2 concentration in
the chamber reached 100%. Following sacrifice, blood samples were
collected from the tail and centrifuged at 2,500 × g, 4°C for 10
min to separate the serum, which was stored at −80°C for subsequent
analysis. Liver tissues were harvested via thoracotomy and the left
lobes were fixed in 4% formaldehyde solution for standby
application at room temperature for 24 h. The remaining liver
tissues were cut into small pieces (2×2 mm) and stored at −80°C
prior to subsequent analysis. The present study was approved by the
Medical Ethics Committee of Jining First People's Hospital and was
performed in accordance with the regulations and procedures
regarding human and animal subject protection. Patients or their
guardians have provided written informed consent for
publication.
Detection of serum ALT, AST and total
bilirubin (TBIL)
Serum transaminases (including ALT and AST) and TBIL
levels may be used as indices for measuring liver damage (14). Serum samples from mice in each
experimental group were analyzed using a fully automatic
biochemical analyzer (Mindray, Shenzhen, China). The serum samples
were diluted (1:10) using normal saline to the same ratio to detect
the ALT, AST and TBIL content and the results were analyzed.
Detection of IL-1β, TGF-β1 and
α-smooth muscle actin (α-SMA) in liver tissues
Levels of IL-1β, TGF-β1 and α-SMA were detected in
liver tissues from mice in each group using the classical
immunohistochemical method (MLB00C, MB100B, MAB1420; R&D
Systems, Inc., Minneapolis, MN, USA) as previously described
(15). The results were
statistically analyzed using SPSS version 18.0 (SPSS, Inc.,
Chicago, IL, USA).
Pathological changes in liver
tissues
Liver tissues were fixed with 10% formaldehyde
solution at room temperature for 24 h, embedded in paraffin and
sectioned into 5 µm thickness according to previous methods
(16), followed by hematoxylin and
eosin staining as previously described (16). Pathological changes in liver tissues
were observed under an optical microscope (magnification, ×400) and
the number of hepatic pseudo-lobules in each visual field selected
a random was counted and compared by eye, followed by relevant
statistical analysis using SPSS version 18.0 (SPSS, Inc.).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Frozen liver tissue samples from mice in each group
were thawed at room temperature and lysed using TRIzol (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) for RNA
detection. RNA was extracted using the RNeasy Plus Mini kit
(Qiagen, Inc., Valencia, CA, USA). Relevant primer sequences of
IL-1β (9), TGF-β1 and α-SMA
(17) were as used in previous
studies (Table I). Extracted RNA was
reverse transcribed into cDNA by RT kit (K1622, Thermo Fisher
Scientific, Inc., Waltham, MA, USA) and amplified by PCR using the
following protocol: 95°C for 30 sec, followed by 40 cycles of 95°C
for 5 sec and 60°C for 31 sec. mRNA expression in each group was
calculated using the 2−∆∆Cq method (18).
 | Table I.Primer sequences for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primer sequences for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Direction | Sequence
(5′-3′) |
|---|
| Interleukin-1β | Forward |
AAAGCTCTCCACCTCAATTGG |
|
| Reverse |
TCGTTGCTTGTCTCTCCTTA |
| Transforming growth
factor-β1 | Forward |
GCGGACTACTATGCTAAAGAGG |
|
| Reverse |
GTAGAGTTCCACATGTTGCTCC |
| α-smooth muscle
actin | Forward |
TGACCCAGATTATGTTTGAGACC |
|
| Reverse |
CCAGAGTCCAGCACAATACCA |
| GAPDH | Forward |
GCGGGCGCTGGAGGAGAA |
|
| Reverse |
GGATCTTCATGAGGTAGTCAGTC |
Western blotting
Frozen liver tissue samples from mice in each
experimental group were thawed and protein was extracted using a
radioimmunoprecipitation lysis buffer [PBS, 0.5% NP-40, 0.5% sodium
deoxycholate, 0.1% sodium dodecyl sulfate, 5.5% β-glycerophosphate,
1 mM dithiothreitol and complete protease and phosphatase
inhibitors (Roche Diagnostics, Basel, Switzerland)]. BCA was used
to quantify the protein content. Protein lysate (50 µg) was
separated using 10% SDS-PAGE gels. Following, proteins were
transferred to polyvinylidene difluoride membranes. Membranes were
blocked with 5% milk at room temperature for 2 h. Membranes were
incubated overnight at 4°C with primary antibodies: Pro-IL-1β
(1:500; ab14374; Abcam, Cambridge, MA, USA), IL-1β (1:500;
ab150777; Abcam), TGF-β1 (1:500; ab64715; Abcam), α-SMA (1:500;
ab124964; Abcam), NLRP3 (1:500; ab214185; Abcam), caspase-1 (1:500;
ab1872; Abcam) and GAPDH (1:500; ab37168; Abcam). Followed by 1 h
incubation at room temperature with goat-anti rabbit (1:500;
sc2004; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) or goat
anti-mouse (1:500; sc2005; Santa Cruz Biotechnology, Inc.)
horseradish peroxidase-conjugated secondary antibodies. Bands were
quantified by densitometry using a Biosens SC-750 Gel Documentation
system (Shanghai Bio-Tech Co., Ltd., Shanghai, China) and the
relative expression of pro-IL-1β, IL-1β, TGF-β1, α-SMA, NLRP3 and
caspase-1 were determined. GAPDH was used as the internal
reference.
Statistical analysis
SPSS software version 18.0 (SPSS, Inc.) was used to
analyze data, and all data are presented as the mean ± standard
deviation. The difference between two experimental groups was
analyzed using a t-test and multiple group comparisons were
performed using one-way analysis of variance followed by a least
significant difference post hoc test. P<0.05 was considered to
indicate a statistically significant difference. Each experiment
was repeated for three times.
Results
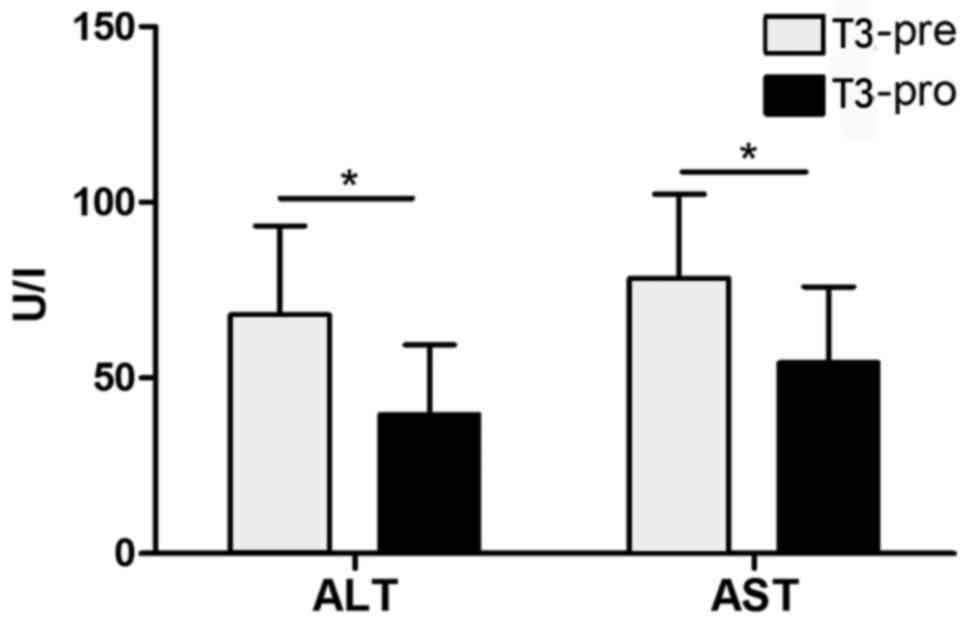
T3 improves liver function and reduces
serum transaminase (ALT/AST) content in patients with ALD
It was revealed that patients' ALT/AST content
following T3 application was significantly decreased compared with
prior to the oral administration of T3 (P<0.05; Fig. 1).
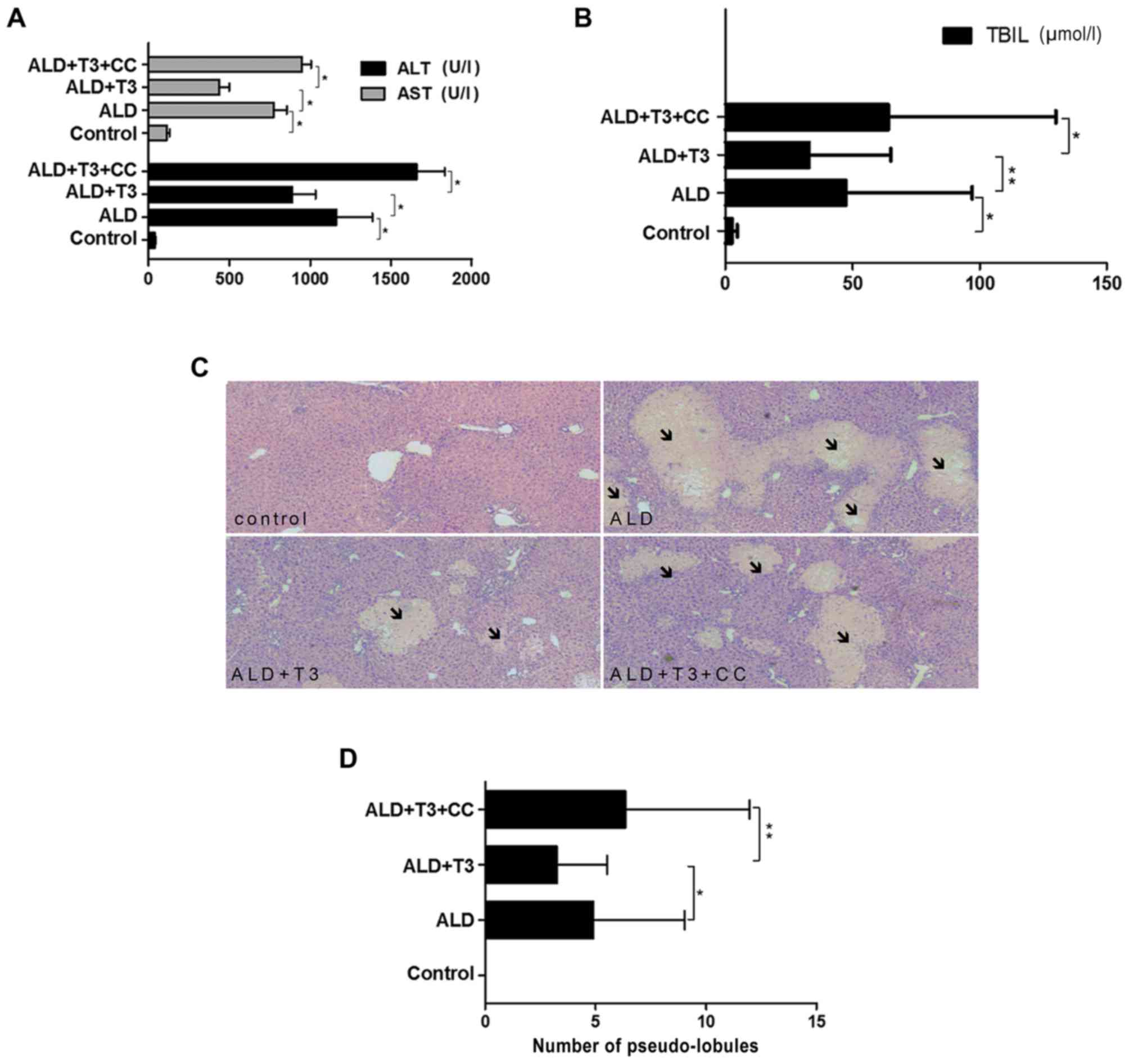
T3 improves liver function and
ameliorates liver damage in mice with alcoholic cirrhosis
The results demonstrated that serum ALT/AST
(Fig. 2A) and TBIL (Fig. 2B) levels in the ALD+T3 group were
significantly reduced compared with those in the ALD group
(P<0.05), whereas ALT/AST levels in the ALD+T3+CC group were
significantly increased compared with those in the ALD+T3 group
(P<0.05). These results suggest that T3 may effectively reduce
the levels of liver enzymes and bilirubin in mice with alcoholic
cirrhosis and have a significant protective effect on the
liver.
Morphological analysis was performed on liver
tissues collected from mice in each experimental group. It is
accepted that pseudo-lobules are a pathological feature of liver
cirrhosis (19). A number of hepatic
lobules were observed in the experimental groups (Fig. 2C). The number of pseudo-lobules
observed in the ALD+T3 group was significantly reduced compared
with the ALD group (P<0.05), while the number of pseudo-lobules
observed in the ALD+T3+CC group was significantly increased
compared with the ALD+T3 group (P<0.01). These results suggest
that T3 may alleviate the severity of liver fibrosis in mice with
alcoholic cirrhosis. Taken together, these findings suggest that
triiodothyronine improves liver function and alleviates the
severity of alcoholic liver cirrhosis in mice, while its effect may
be blocked by CC.
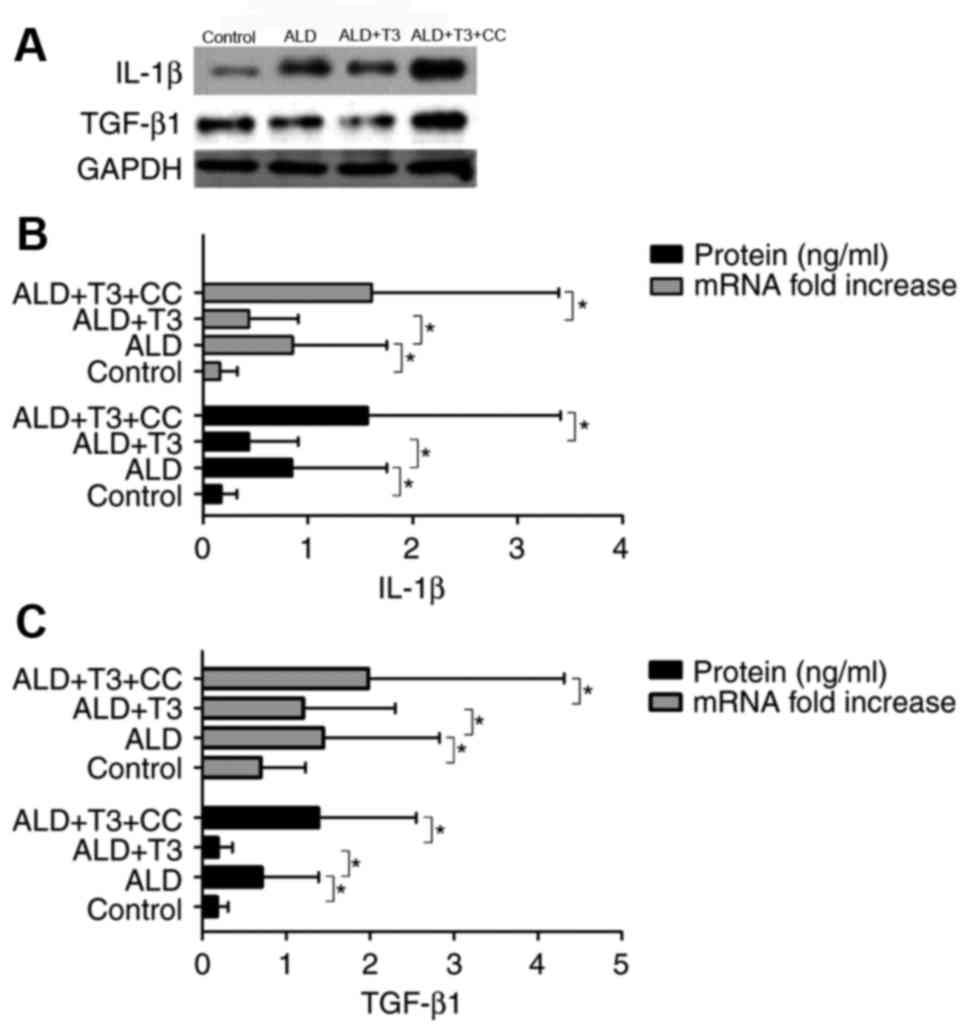
T3 downregulates IL-1β and TGF-β1 in
liver tissues
IL-1β (4) and TGF-β1
(20) are important inflammatory
mediators associated with the occurrence and development of
alcoholic cirrhosis. Western blotting (Fig. 3A) and ELISA analysis revealed that
the expression of IL-1β (Fig. 3B)
and TGF-β1 (Fig. 3C) mRNA and
protein was significantly decreased in the ALD+T3 group compared
with the ALD group (P<0.05). However, expression in the
ALD+T3+CC group was significantly increased compared with the
ALD+T3 group (P<0.05). These results suggest that T3 may
effectively ameliorate the inflammatory response in mice with
alcoholic cirrhosis and that this effect may be countered by
CC.
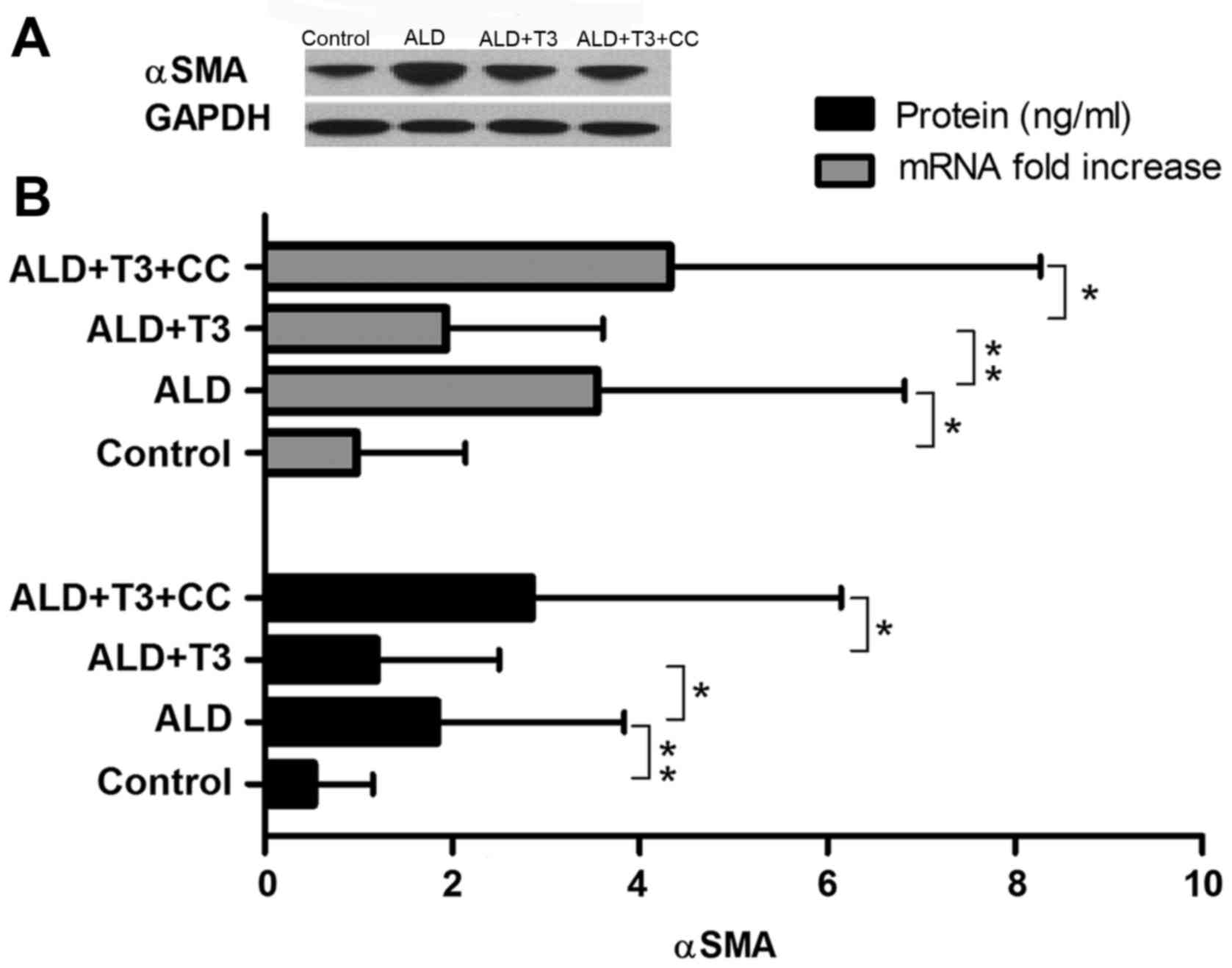
T3 reduces the expression of α-SMA in
liver tissues
Previous studies have demonstrated that the
occurrence and development of liver cirrhosis is associated with
the activation of hepatic stellate cells (21), which results in colloid and
deposition. A key marker of hepatic stellate cell activation is the
production of α-SMA (22). In the
present study, western blotting revealed that the expression of
α-SMA protein in the ALD+T3 group was significantly decreased
compared with the ALD group (P<0.05); however, it was
significantly increased in the ALD+T3+CC group compared with the
ALD+T3 group (P<0.05; Fig. 4).
These results suggest that T3 may effectively reduce the α-SMA
content in the liver tissues of mice with alcoholic cirrhosis. It
was also revealed that the level of α-SMA mRNA was significantly
decreased in the ALD+T3 group compared with the ALD group
(P<0.01), whereas it was significantly increased in the
ALD+T3+CC group compared with the ALD+T3 group (P<0.05). These
results reveal that T3 significantly reduces the expression of
α-SMA mRNA and protein in ALD mice and alleviates the severity of
liver cirrhosis.
T3 negatively regulates the
NLRP3-Caspase-1-pro-IL-1β-IL-1β signaling pathway and the
NLRP3-TGF-β1 signaling pathway
To further study the molecular mechanism by which T3
alleviates liver damage in ALD mice, various associated proteins
were detected. Western blotting (Fig.
5A) revealed that the expression of NLRP3, Caspase-1, IL-1β and
TGF-β1 in the ALD+T3 group was significantly decreased compared
with the ALD group (P<0.05), whereas it was significantly
increased in the ALD+T3+CC group compared with the ALD+T3 group
(P<0.05; Fig. 5B). Conversely,
the expression of pro-IL-1β was significantly increased in the
ALD+T3 group compared with the ALD group and significantly
increased in the ALD+T3+CC group compared with the ALD+T3 group
(both P<0.05). Previous studies have demonstrated that NLRP3
activates pro-caspase-1 into caspase-1, which can mediate
hepatocyte apoptosis (23) and
promote the transformation of pro-IL-1β into IL-1β8,
which aggravates inflammatory injury in liver tissues. These
results suggest that T3 alleviates liver injury in mice with
alcoholic cirrhosis via negative regulation of the NLRP3 signaling
pathway; however, this effect may be blocked by CC.
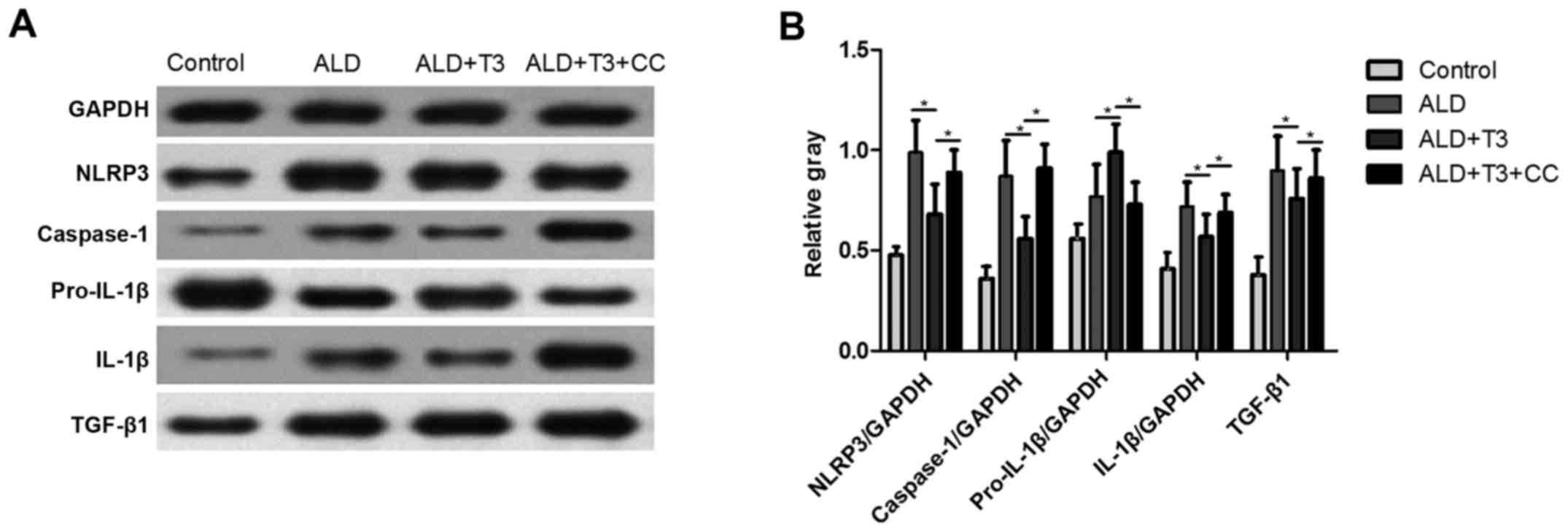 | Figure 5.(A) Protein expression of NLRP3,
caspase-1, pro-IL-1β, IL-1β and TGF-β1 as measured using western
blotting. (B) Data was quantified using densitometry analysis
(n=10). Data are expressed as the mean ± standard deviation.
*P<0.05. IL, interleukin; NLRP3, nucleotide-binding
oligomerization domain, leucine rich repeat containing family,
pyrin domain containing 3; TGF, transforming growth factor; ALD,
alcoholic liver disease; T3, triiodothyronine; CC, AMP-activated
protein kinase inhibitor. |
Discussion
A total of 10 patients with liver cirrhosis were
randomly selected and their ALT/AST levels were detected. Patients
were treated with T3 (0.1 mg/kg/day) orally for 2 weeks. Further
blood samples were taken and ALT/AST content prior to and following
treatment was compared. It was revealed that ALT/AST content was
significantly decreased in patients following T3 application
compared with the baseline. Animal studies were performed to
investigate the potential mechanisms that may be responsible for
this.
It was observed that T3 significantly reduced serum
transaminase and TBIL levels in ALD mice, which indicated that T3
effectively reduced the liver damage caused by ALD. However, the
effect of T3 treatment was significantly blocked by CC. In
addition, morphological observations of liver tissues revealed that
the number of hepatic lobules in the ALD+T3 group was significantly
reduced compared with the ALD group, whereas they were
significantly increased in the ALD+T3+CC group, which suggested
that T3 alleviates the severity of liver fibrosis in ALD mice.
These results indicate that T3 improved liver function and reduced
the severity of liver cirrhosis in ALD mice, while this effect was
blocked by CC. The possible mechanism responsible for the effect of
T3 on ALD mice was then investigated.
Previous studies have demonstrated that chronic
alcohol ingestion causes liver injury and increases the levels of
ALT/AST and TBIL (24,25). Animal experiments have also revealed
that the increase in ALT/AST and TBIL may be associated with
inflammation and apoptosis triggered by endoplasmic reticulum
stress (26,27). It was recently reported that T3
ameliorates ischemia-reperfusion injury by suppressing NLRP3
activation (28,29). Therefore the authors hypothesized
that the protective effects of T3 in ALD may involve inflammatory
mediators and apoptosis factors.
IL-1β (4) and TGF-β1
(20) are important inflammatory
mediators that serve a role in the occurrence and development of
ALD. In the present study, T3 significantly reduced the expression
of IL-1β and TGF-β1 mRNA and protein in the liver tissues from ALD
mice, whereas its effect was blocked by CC. These results suggest
that the initial effect of T3 on ALD was to reduce the expression
of IL-1β and TGF-β1. It has been previously reported that NLRP3 is
a key mediator of proinflammatory cytokine release (28). Previous studies have reported that
caspase-1 promotes the transformation of pro-IL-1β into IL-1β,
which aggravates liver damage (7,8). It has
also been reported that NLRP3 upregulates the expression of
caspase-1 (28). Based on these
previous studies, the authors investigated whether the expression
of IL-1β in ALD was associated with the regulation of the
NLRP3-caspase-1 signaling pathway.
It was demonstrated that the expression of NLRP3,
caspase-1 and IL-1β in liver tissues was significantly decreased
following T3 application in ALD mice, whereas the expression of
pro-IL-1β was significantly increased. This suggests that T3
negatively regulates the NLRP3-caspase-1 signaling pathway, thereby
significantly reducing the expression of IL-1β. Previous study has
reported that NLRP3 induces the activation of hepatic stellate
cells, which is essential in the development of liver cirrhosis
caused by ALD (30). The present
study revealed that the mRNA and protein expression of α-SMA in the
ALD group was significantly increased compared with the control
group. Following the application of T3, α-SMA expression in the
liver tissues of the mice was significantly decreased; however, the
effect of T3 was blocked by CC. α-SMA is an important marker of
hepatic stellate cell activation (21), therefore these results suggest that
T3 inhibits the production of TGF-β1 and reduces the activation of
hepatic stellate cells, thereby downregulating the expression of
α-SMA and alleviating the severity of liver cirrhosis.
Evidence suggests that the NLRP3 inflammasome serves
a key role in the occurrence and development of tissue fibrosis
(31). Gasse et al (32) demonstrated that the NLRP3
inflammasome existed in an activated state and was necessary during
bleomycin-induced pulmonary fibrosis. In addition, previous studies
have revealed that the NLRP3 inflammasome mediates the occurrence
of fibrosis in systemic sclerosis24 and this can lead to
cardiac infarction (33). A previous
study using mice with a deleted NLRP3 gene revealed that the NLRP3
inflammasome induced liver fibrosis (33). Therefore, the NLRP3 downregulation
observed in the present study may alleviate the severity of liver
fibrosis in ALD. It has also been reported that the NLRP3
inflammasome regulates the activation of hepatic stellate cells
(34); studies have confirmed that
activation of the NLRP3 inflammasome with monosodium urate crystals
upregulates the expression of TGF-β1 and collagen-1 (6). In the present study, the expression of
NLRP3, TGF-β1 and α-SMA protein was significantly decreased
following the application of T3, which may be blocked by CC. This
indicates that T3 alleviates liver fibrosis in ALD, and that this
is associated, at least in part, with negative regulation of the
NLRP3-TGF-β1 signaling pathway.
In conclusion, T3 alleviates the damage caused by
ALD and its role may be associated with the fact that T3 negatively
regulates the NLRP3 signaling pathway, thereby downregulating IL-1β
and TGF-β1 and reducing the activation of stellate cells. These
findings provide a basis for a novel treatment option; in the
future, hormone therapy may be used to relieve injury caused by
alcoholic cirrhosis. However, there are several limitations in the
present study, most notably the fact that only in vivo
experiments were performed. These results should be supported by
corresponding in vitro experiments. Further investigation is
required to clarify the role of T3 and confirm its mechanism of
action through in vitro experiments.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XD designed the study. HY collected the data. HY and
XD performed the experiments. CL and QB analyzed and QL and ZZ
interpreted the data. All authors read and approved the final
version of the manuscript.
Ethics approval and consent to
participate
This study was approved by the Medical Ethics
Committee of Jining First People's Hospital (Jining, China).
Patients or their guardians have provided written informed consent
for publication.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Federico A, Dallio M, Masarone M, Persico
M and Loguercio C: The epidemiology of non-alcoholic fatty liver
disease and its connection with cardiovascular disease: Role of
endothelial dysfunction. Eur Rev Med Pharmacol Sci. 20:4731–4741.
2016.PubMed/NCBI
|
|
2
|
Arsene D, Farooq O and Bataller R: New
therapeutic targets in alcoholic hepatitis. Hepatol Int.
10:538–552. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schwartz JM and Reinus JF: Prevalence and
natural history of alcoholic liver disease. Clin Liver Dis.
16:659–666. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Petrasek J, Bala S, Csak T, Lippai D,
Kodys K, Menashy V, Barrieau M, Min SY, Kurt-Jones EA and Szabo G:
IL-1 receptor antagonist ameliorates inflammasome-dependent
alcoholic steatohepatitis in mice. J Clin Invest. 122:3476–3489.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim EJ, Park SY, Baek SE, Jang MA, Lee WS,
Bae SS, Kim K and Kim CD: HMGB1 increases IL-1β production in
vascular smooth muscle cells via NLRP3 inflammasome. Front Physiol.
9:3132018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim HY, Kim SJ and Lee SM: Activation of
NLRP3 and AIM2 inflammasomes in Kupffer cells in hepatic
ischemia/reperfusion. Febs J. 282:259–270. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Watanabe A, Sohail MA, Gomes DA, Hashmi A,
Nagata J, Sutterwala FS, Mahmood S, Jhandier MN, Shi Y, Flavell RA
and Mehal WZ: Inflammasome-mediated regulation of hepatic stellate
cells. Am J Physiol Gastrointest Liver Physiol. 296:G1248–G1257.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Esser N, Legrand-Poels S, Piette J, Scheen
AJ and Paquot N: Inflammation as a link between obesity, metabolic
syndrome and type 2 diabetes. Diabetes Res Clin Pract. 105:141–150.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dinarello CA: Immunological and
inflammatory functions of the interleukin-1 family. Annu Rev
Immunol. 27:519–550. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vargas R and Videla LA: Triiodothyronine
suppresses ischemia-reperfusion-induced liver NLRP3 inflammasome
activation: Role of AMP-activated protein kinase. Immunol Lett.
184:92–97. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xiao J, Zhu Y, Liu Y, Tipoe GL, Xing F and
So KF: Lycium barbarum polysaccharide attenuates alcoholic cellular
injury through TXNIP-NLRP3 inflammasome pathway. Int J Biol
Macromol. 69:73–78. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cable EE, Finn PD, Stebbins JW, Hou J, Ito
BR, van Poelje PD, Linemeyer DL and Erion MD: Reduction of hepatic
steatosis in rats and mice after treatment with a liver-targeted
receptor agonist. Hepatology. 49:407–417. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
O'Shea RS, Dasarathy S and McCullough AJ:
Alcoholic liver disease. AM J Gastroenterol. 105(14–32):
332010.
|
|
14
|
Rej R: Aspartate aminotransferase activity
and isoenzyme proportions in human liver tissues. Clin Chem.
24:1971–1979. 1978.PubMed/NCBI
|
|
15
|
Guesdon JL, Ternynck T and Avrameas S: The
use of avidin-biotin interaction in immunoenzymatic techniques. J
Histochem Cytochem. 27:1131–1139. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kozutsumi Y, Segal M, Normington K,
Gething MJ and Sambrook J: The presence of malfolded proteins in
the endoplasmic reticulum signals the induction of
glucose-regulated proteins. Nature. 332:462–464. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tian X, Zhao C, Guo J, Xie S, Yin F, Huo X
and Zhang X: Carvedilol attenuates the progression of hepatic
fibrosis induced by bile duct ligation. Biomed Res Int.
2017:46127692017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ke B, Shen XD, Zhang Y, Ji H, Gao F, Yue
S, Kamo N, Zhai Y, Yamamoto M, Busuttil RW and Kupiec-Weglinski JW:
KEAP1-NRF2 complex in ischemia-induced hepatocellular damage of
mouse liver transplants. J Hepatol. 59:1200–1207. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee YA, Wallace MC and Friedman SL:
Pathobiology of liver fibrosis: A translational success story. Gut.
64:830–841. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang H, Liu S, Wang Y, Chang B and Wang B:
Nod-like receptor protein 3 inflammasome activation by Escherichia
coli RNA induces transforming growth factor beta 1 secretion in
hepatic stellate cells. Bosn J Basic Med Sci. 16:126–131.
2016.PubMed/NCBI
|
|
21
|
Li J, Li J, Li S, He B, Mi Y, Cao H, Zhang
C and Li L: Ameliorative effect of grape seed proanthocyanidin
extract on thioacetamide-induced mouse hepatic fibrosis. Toxicol
Lett. 213:353–360. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiao J, Friedman SL and Aloman C: Hepatic
fibrosis. Curr Opin Gastroenterol. 25:223–229. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dara L, Ji C and Kaplowitz N: The
contribution of endoplasmic reticulum stress to liver diseases.
Hepatology. 53:1752–1763. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhao S, Li N, Zhen Y, Ge M, Li Y, Yu B, He
H and Shao RG: Protective effect of gastrodin on bile duct
ligation-induced hepatic fibrosis in rats. Food Chem Toxicol.
86:202–207. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu JT, Yang GW, Qi CH, Zhou L, Hu JG and
Wang MS: Anti-inflammatory activity of platycodin d on
alcohol-induced fatty liver rats via tlr4-myd88-nf-kappab signal
path. Afr J Tradit Complement Altern Med. 13:176–183. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang W, Zhong W, Sun Q, Sun X and Zhou Z:
Adipose-specific lipin1 overexpression in mice protects against
alcohol-induced liver injury. Sci Rep. 8:4082018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen X, Bian M, Zhang C, Kai J, Yao Z, Jin
H, Lu C, Shao J, Chen A, Zhang F and Zheng S: Dihydroartemisinin
inhibits ER stress-mediated mitochondrial pathway to attenuate
hepatocyte lipoapoptosis via blocking the activation of the
PI3K/Akt pathway. Biomed Pharmacother. 97:975–984. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vargas R and Videla LA: Triiodothyronine
suppresses ischemia-reperfusion-induced liver NLRP3 inflammasome
activation: Role of AMP-activated protein kinase. Immunol Lett.
184:92–97. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cannito S, Morello E, Bocca C, Foglia B,
Benetti E, Novo E, Chiazza F, Rogazzo M, Fantozzi R, Povero D, et
al: Microvesicles released from fat-laden cells promote activation
of hepatocellular NLRP3 inflammasome: A pro-inflammatory link
between lipotoxicity and non-alcoholic steatohepatitis. PLoS One.
12:e1725752017. View Article : Google Scholar
|
|
30
|
Kawada N: Evolution of hepatic fibrosis
research. Hepatol Res. 41:199–208. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Artlett CM: Inflammasomes in wound healing
and fibrosis. J Pathol. 229:157–167. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gasse P, Mary C, Guenon I, Noulin N,
Charron S, Schnyder-Candrian S, Schnyder B, Akira S, Quesniaux VF,
Lagente V, et al: IL-1R1/MyD88 signaling and the inflammasome are
essential in pulmonary inflammation and fibrosis in mice. J Clin
Invest. 117:3786–3799. 2007.PubMed/NCBI
|
|
33
|
Shinde AV and Frangogiannis NG:
Fibroblasts in myocardial infarction: A role in inflammation and
repair. J Mol Cell Cardiol. 70:74–82. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wree A, Eguchi A, McGeough MD, Pena CA,
Johnson CD, Canbay A, Hoffman HM and Feldstein AE: NLRP3
inflammasome activation results in hepatocyte pyroptosis, liver
inflammation, and fibrosis in mice. Hepatology. 59:898–910. 2014.
View Article : Google Scholar : PubMed/NCBI
|