Introduction
Cirrhosis is the end stage of chronic liver disease
and a major risk factor of hepatocellular carcinoma (1). In recent years, it has been reported
that intestinal barrier disruption and increased intestinal
permeability may lead to bacterial translocation (BT) and
absorption of endotoxemia, which increases susceptibility to
infection (2). The intestinal
barrier includes secreted mucus and the epithelial cell layer
itself. Increased intestinal permeability is the main cause of
intestinal barrier dysfunction (3).
Intestinal epithelial monolayer cells are the main barrier of the
intestinal mucosa, which serves an important role in preventing the
infiltration of a number of harmful substances, allergens, toxins
and luminal microbial pathogens (4,5). The
functional integrity of the epithelial barrier is maintained by
intercellular tight junctions (TJs), which include the
transmembrane protein occludin, claudins and the cytoplasmic
protein zonula occludens 1 (ZO-1) (6). Intercellular TJ protein destruction is
associated with a number of diseases, including cirrhosis (7). Therefore, restoring the integrity of
the intestinal barrier is an important step in preventing
intestinal endotoxemia.
Lipopolysaccharide (LPS), also known as endotoxin,
is the main component of the outer wall of Gram-negative bacteria
and is released from the bacterial cell wall via shedding or
bacterial lysis (8,9). Endotoxin causes an increase in
intestinal permeability by activating proinflammatory cytokines,
including tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6) and
nitric oxide, and injuring the intercellular TJ between epithelial
cells (10,11). It has been reported that patients
with cirrhosis develop intestinal endotoxemia with an incidence of
79–92% (12), while other clinical
and experimental studies suggest that liver cirrhosis is associated
with endotoxemia (13,14).
Bletilla striata is a traditional medicine
that has been widely used to treat ulcers, bleeding, burns, bruises
and many other diseases for >1,000 years (15,16).
B. striata polysaccharide (BSP) is extracted from B.
striata and consists of α-mannose, β-mannose and β-glucose
(17). BSP has a number of
biological functions, including anti-inflammatory (18), anti-tumor (19), anti-fibrosis (20) and antibacterial (21) properties, as well as inducing
endothelial cell proliferation and vascular endothelial
growth-factor expression (16).
However, to the best of our knowledge, no previous studies have
investigated the protective role of BSP in epithelial barrier
disruption.
The aim of the present study was to investigate
whether BSP is able to restore the integrity and function of the
intestinal epithelial barrier via inhibiting the expression of
inflammatory cytokines and improving TJs in the small intestine of
rats with thioacetamide (TAA)-induced liver cirrhosis.
Materials and methods
Chemicals and reagents
BSP (≥95% titration) was purchased from Dalian
MeiLun Biotechnology Co., Ltd. (Dalian, China). TAA was purchased
from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany) and dissolved
in physiological saline (0.9% NaCl) to give a 4% solution for
animal experiments. Rabbit anti-rat zonula occludens (ZO)-1 and
occludin were obtained from ProteinTech Group, Inc. (Chicago, IL,
USA). ELISA kits for serum alanine aminotransferase (ALT; cat. no.
C009-2) and aspartate aminotransferase (AST; cat. no. C0010-2) were
purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing,
China). ELISA kits for IL-6 (cat. no. E-EL-R0015), TNF-α (cat. no.
E-EL-R0019) and endotoxin (cat. no. E-EL-R0589) were purchased from
Elabscience Biotechnology (Wuhan, China). GAPDH and horseradish
peroxidase (HRP)-labeled secondary antibodies were acquired from
Wuhan Boster Biotechnology Co., Ltd. (Wuhan, China). RNAiso Plus,
PrimeScript™ RT reagent and SYBR Premix Ex Taq kits were purchased
from Takara Bio, Inc. (Otsu, Japan).
Animals and experimental
protocols
A total of 50 healthy male Sprague-Dawley rats (age,
6 weeks; weight, 180–220 g) were purchased from Hubei Provincial
Center for Disease Control and Prevention (Wuhan, China). All
animals were housed at 22–25°C, relative humidity of 50±10% with a
12 h light/dark cycle and free access to water and food. All animal
experiment protocols used in the present study followed
internationally accepted principles and were approved by The
Institutional Animal Care and Use Committee of Tongji Medical
College, Huazhong University of Science and Technology (Wuhan,
China). Rats were randomly divided into 5 groups (n=10) as follows:
15 mg/kg BSP, 30 mg/kg BSP, 60 mg/kg BSP, experiment and control
groups. Rats in the BSP and experimental groups were administered
intraperitoneally (IP) with TAA (200 mg/kg, dissolved in 0.9% w/v
saline solution to give 4% solution) twice per week for 14 weeks to
induce liver cirrhosis. The control group received IP injections of
the same volume of saline (0.9% w/v). At 12 weeks, 3 rats from the
experimental and control groups were selected at random and
anesthetized by IP. Injection with 10% chloral hydrate (300 mg/kg).
The abdominal cavity was opened with scissors. Livers were
harvested and examined using hematoxylin and eosin (HE) staining to
confirm the development of liver cirrhosis. Rats in the
experimental group were administered with saline at 1 ml/100 g once
daily by gavage for 2 weeks while rats in the BSP groups were
administered with BSP at 15, 30 or 60 mg/kg for 2 weeks at
concentrations of 1.5, 3 or 6 mg/ml at 1 ml/100 g once daily by
gavage. TAA administration was continued until the end of the
14-week experimental period. At 14 weeks, the remaining rats were
injected IP with 10% chloral hydrate (300 mg/kg) and dissected.
Blood samples were collected from portal veins and intestinal
tissues were harvested for further analysis.
Serum alanine aminotransferase (ALT)
and aspartate aminotransferase (AST) analysis
Blood samples were obtained and centrifuged at 1,760
× g for 15 min at 4°C to collect the serum. Serum ALT and AST
levels were determined using assay kits according to the
manufacturer's protocols.
Measurement of serum endotoxin
levels
Endotoxin levels were measured in all groups using
specific ELISA kit according the manufacturer's instructions.
Briefly, portal vein blood samples were collected and centrifuged
at 1,760 × g for 15 min at 4°C to collect plasma, which was
subsequently diluted with endotoxin-free water and incubated at
70°C for 10 min. Samples were then incubated in ice-cold water for
3 min, following which limulus reagents and 200 µl processed mixing
solution were added and 100 µl of mixture was transferred to a
96-well microplate for analysis. The concentration of endotoxin was
calculated using a standard curve and the absorbance of each well
was measured at 450 nm using a microplate reader to calculate the
concentration of endotoxin (EU/ml).
ELISA measurement of IL-6 and TNF-α in
ileal tissues
Harvested ileal tissues were cut into small pieces
and homogenized in pre-cooled PBS containing proteinase inhibitors
(BioSharp, Hefei, China). The supernatant of the homogenate was
collected and ELISA was performed to measure the concentrations of
IL-6 and TNF-α according to the manufacturer's protocol. All
samples were tested in duplicate.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from rat ileal tissues using
RNAiso Plus according to the manufacturer's protocol and stored at
−80°C prior to use. cDNA was produced using a PrimeScript™ RT
reagent kit with the following temperature protocol: 37°C for 15
min and 85°C for 5 sec. qPCR reactions were performed using a
StepOne Plus device (Applied Biosystems; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) at 95°C for 10 sec, followed by 40 cycles
of 95°C for 5 sec and 60°C for 20 sec according to the instructions
of the SYBR Premix Ex Taq kit. Data were quantified using the
2−∆∆Cq method (22).
Primers were synthesized by GenScript (Piscataway, NJ, USA) and
sequences were as follows: ZO-1, forward 5′-GCTCACCAGGGTCAAAATGT-3′
and reverse 5′-GGCTTAAAGCTGGCAGTGTC-3′; occludin, forward
5′-TTACGGCTATGGAGGGTACAC-3′ and reverse
5′-GACGCTGGTAACAAAGATCAC-3′; and GAPDH, forward
5′-GGAAAGCTGTGGCGTGAT-3′ and reverse
5′-AAGGTGGAAGAATGGGAGTT-3′.
Western blotting
Western blotting was performed as previously
described (23). Briefly, total
proteins were extracted from the ileum tissue using
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology, Wuhan, China). A bicinchoninic protein assay kit was
used to measure protein concentrations. A total of 20 µg proteins
in each sample were separated on 10% SDS-PAGE gels and transferred
to polyvinylidene fluoride membranes. Membranes were blocked with
PBS containing 5% nonfat milk powder for 1 h at room temperature
and incubated overnight at 4°C with antibodies against ZO-1
(1:1,000; cat. no. 21773-1-AP), occludin (1:1,000; cat. no.
13409-1-AP) and GAPDH (1:5,000; cat. no. BM1985). Membranes were
washed three times with TBST and incubated at 4°C with HRP-labeled
secondary antibody (1:2,000; cat. no. BA1058) for 1 h. Membranes
were washed with TBST three times and enhanced chemiluminescence
(Millipore; Merck KGaA, Darmstadt, Germany) was used to identify
immunoreactive bands and followed the manufacturer's protocol.
Bands underwent densitometric analysis using the Fuji
ultrasonic-Doppler velocity profile (UVP) system and Image J
software (v.1.50; National Institutes of Health, Bethesda, MD,
USA).
Histopathology
Terminal ileal samples were collected, fixed in 4%
paraformaldehyde solution at room temperature for 24 h, embedded in
paraffin and cut into 4–5 µm sections. Sections were deparaffinized
using dimethyl benzene, dehydrated using alcohol for 2–5 min at
room temperature, washed with distilled water and stained with
hematoxylin at room temperature for 10–30 min. Excess stain was
removed and sections were incubated with 1% acid alcohol for 30 sec
at room temperature and dehydrated using gradient alcohol (70, 85,
95 and 100%; 2–3 min, respectively) at room temperature. Sections
were stained with 0.5% eosin at room temperature for 2–5 min,
incubated with 95% alcohol at room temperature for 2 min and with
xylene at room temperature for 10 min and finally sealed with a
neutral balsam. Sections were mounted and histological changes were
observed using light microscopy (magnification, ×200).
Immunohistochemistry (IHC) of ileal
proteins
The ileal tissue specimens were cut into 10 µm
sections following dewaxing and hydrating. Sections were treated
with 3% H2O2-methanol to block endogenous
peroxidase activity, following which they were incubated with 5%
normal goat serum (Wuhan Boster Biological Technology, Ltd.) at
room temperature for 10 min and incubated with ZO-1 and occludin
antibodies (dilution 1:200) overnight at 4°C. Slides were
subsequently washed with PBS and incubated with a biotinylated
secondary antibody (1:500; cat. no. SA00004-2; Wuhan Boster
Biological Technology, Ltd.) for 1 h at room temperature. Slides
were washed with PBS again and incubated with HRP-labeled
streptavidin (1:200; ab214880; Abcam) at 37°C for 1 h. Samples were
developed using diaminobenzene (DAB) at room temperature for 30 sec
and counterstained with hematoxylin at room temperature for 5 min.
Slides were rinsed in distilled water and dehydrated, following
which they were observed under a light microscope (magnification,
×100).
Statistical analysis
Data are presented as the mean ± standard error of
the mean. Data were compared between groups using one-way analysis
of variance with the Student-Newman-Keuls post-hoc test. All
statistical analyses were performed with SPSS software version 12.0
(SPSS, Inc., Chicago, IL, USA). P<0.05 was considered
statistically significant difference. Graphs were created using
GraphPad Prism software (version 6; GraphPad Software, Inc., La
Jolla, CA, USA).
Results
Liver histological changes in rats
following treatment with thioacetamide for 12 weeks
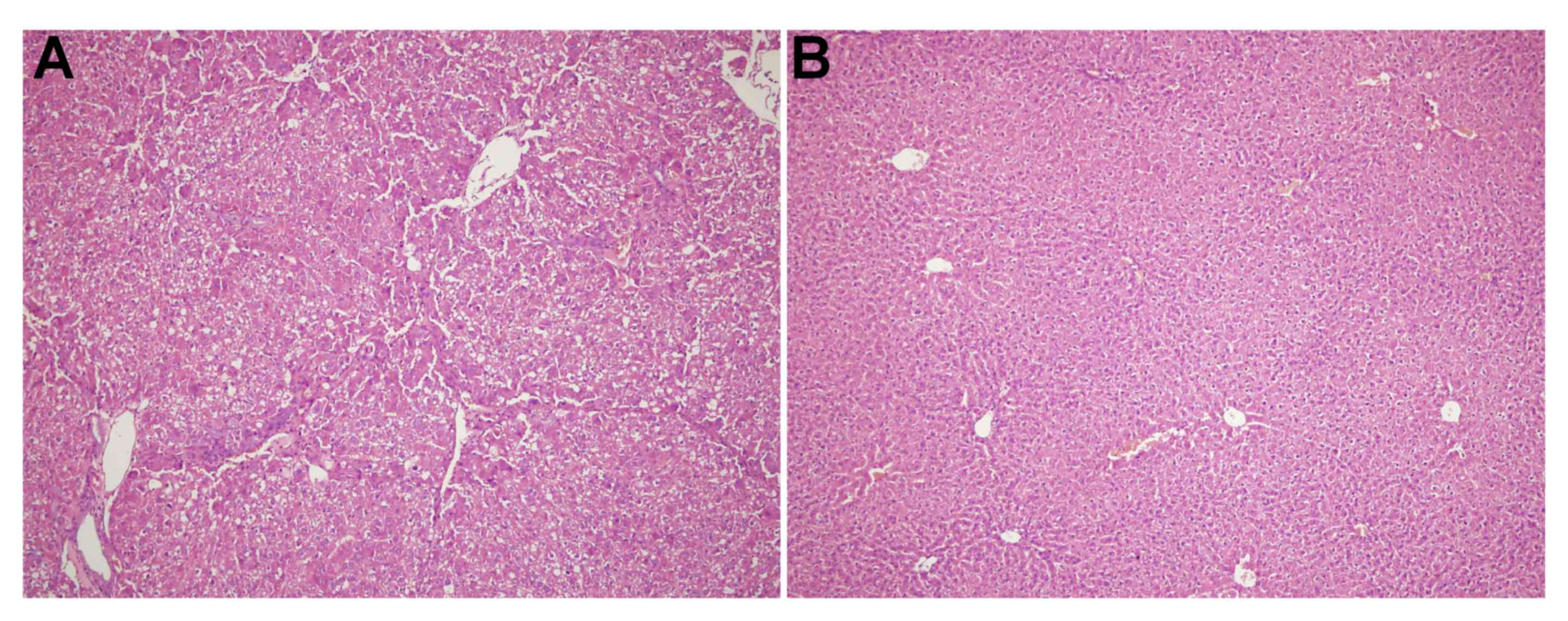
As shown in Fig. 1A,
the liver of the experiment group exhibited a lobular structure
destruction, inflammatory cell infiltration and the loss of
structural integrity. The liver of the control group (Fig. 1B) revealed structural liver integrity
without inflammatory cell infiltration.
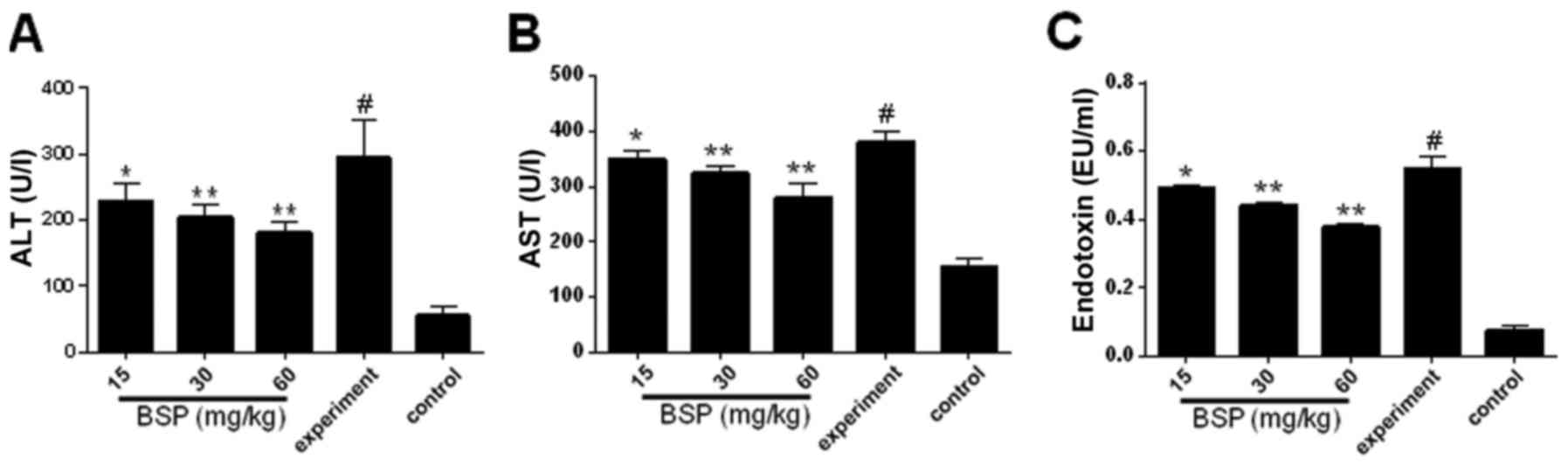
Activity of transaminases
The expression of serum AST and ALT was assessed
using ELISA kits and was demonstrated to be significantly increased
in the experiment group compared with the control (Fig. 2A and B). However, treatment with BSP
at all concentrations resulted in a significant reduction in serum
ALT and AST expression compared with the experiment group (Fig. 2A and B).
BSP reduces TAA-induced plasma
endotoxin level
As shown in Fig. 2C,
plasma endotoxin levels were significantly increased in the
experiment group compared with the control group. Treatment with
BSP at all dosages obviously decreased endotoxin levels compared
with the experiment group.
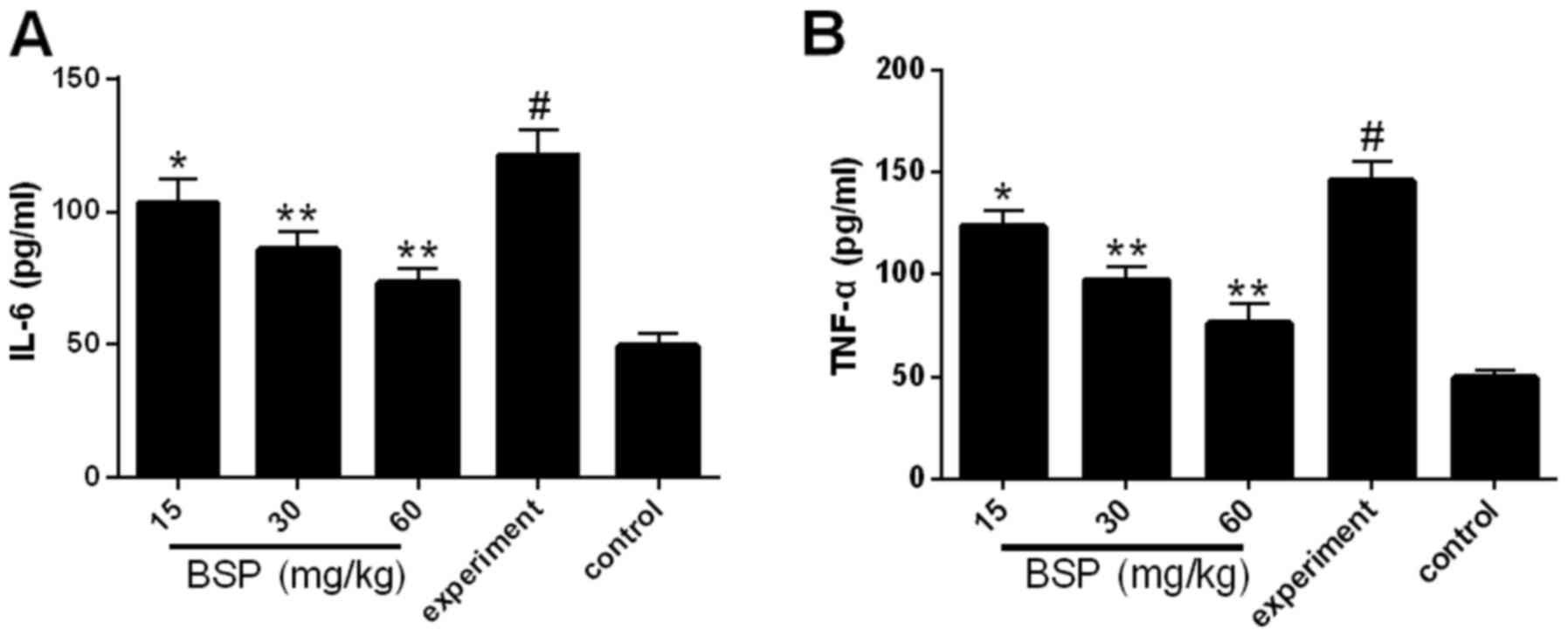
Effects of BSP on inflammatory
cytokines expression in ileal tissue
Levels of the inflammatory cytokines IL-6 and TNF-α
were measured in the ileal tissues. Compared with the control
group, the expression of IL-6 and TNF-α was significantly increased
in the experiment group (Fig. 3).
However, following treatment with BSP at all concentrations, IL-6
and TNF-α expression was significantly decreased compared with the
experiment group (Fig. 3).
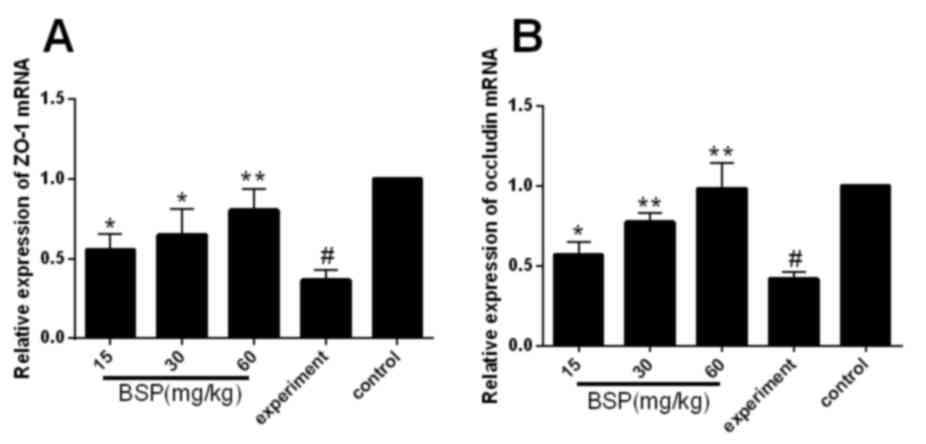
Effects of BSP on the expression of
ZO-1 and occludin mRNA in ileal tissues
RT-qPCR was used to assess the effect of BSP on ZO-1
and occludin mRNA expression. It was demonstrated that ZO-1 and
occludin mRNA was downregulated in the experiment group compared
with the control group (Fig. 4),
while treatment with BSP significantly upregulated ZO-1 and
occludin mRNA compared with the experiment group (Fig. 4).
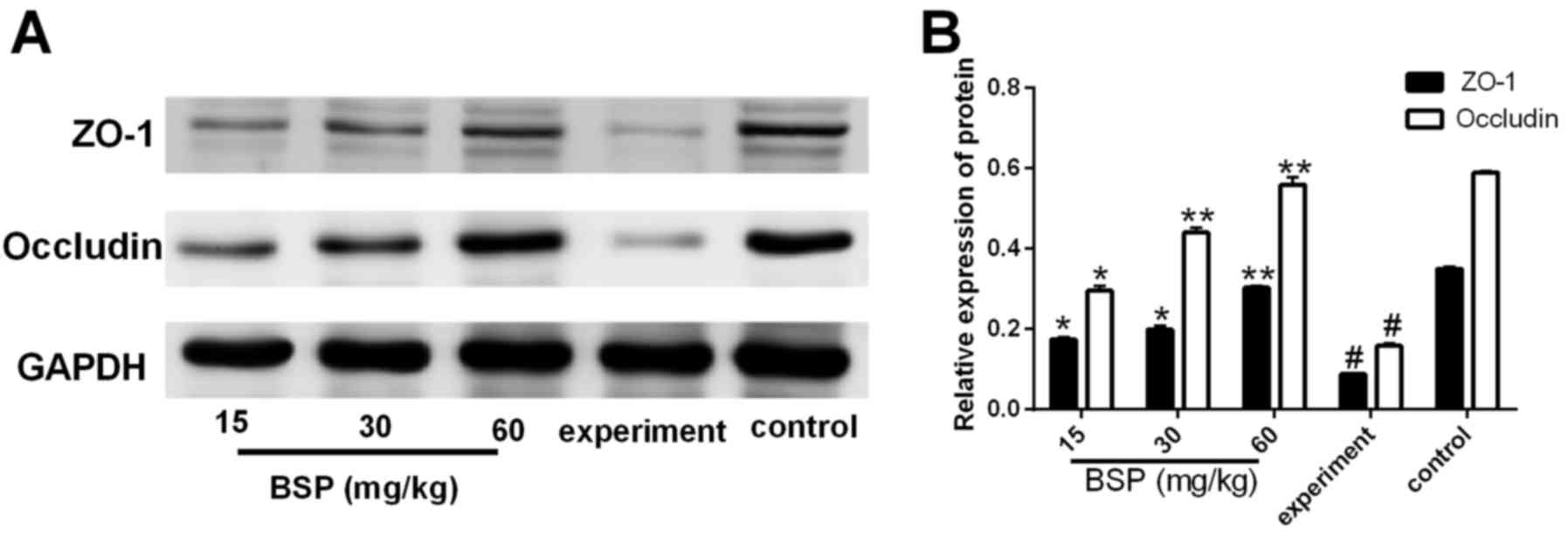
Effects of BSP on the expression of
ZO-1 and occludin protein in ileal tissues
Compared with the control group, the expression of
ZO-1 and occludin proteins in the experiment group was
significantly decreased (Fig. 5).
However, following treatment with BSP at different concentrations,
the expression of ZO-1 and occludin proteins was significantly
increased compared with the experiment group (Fig. 5).
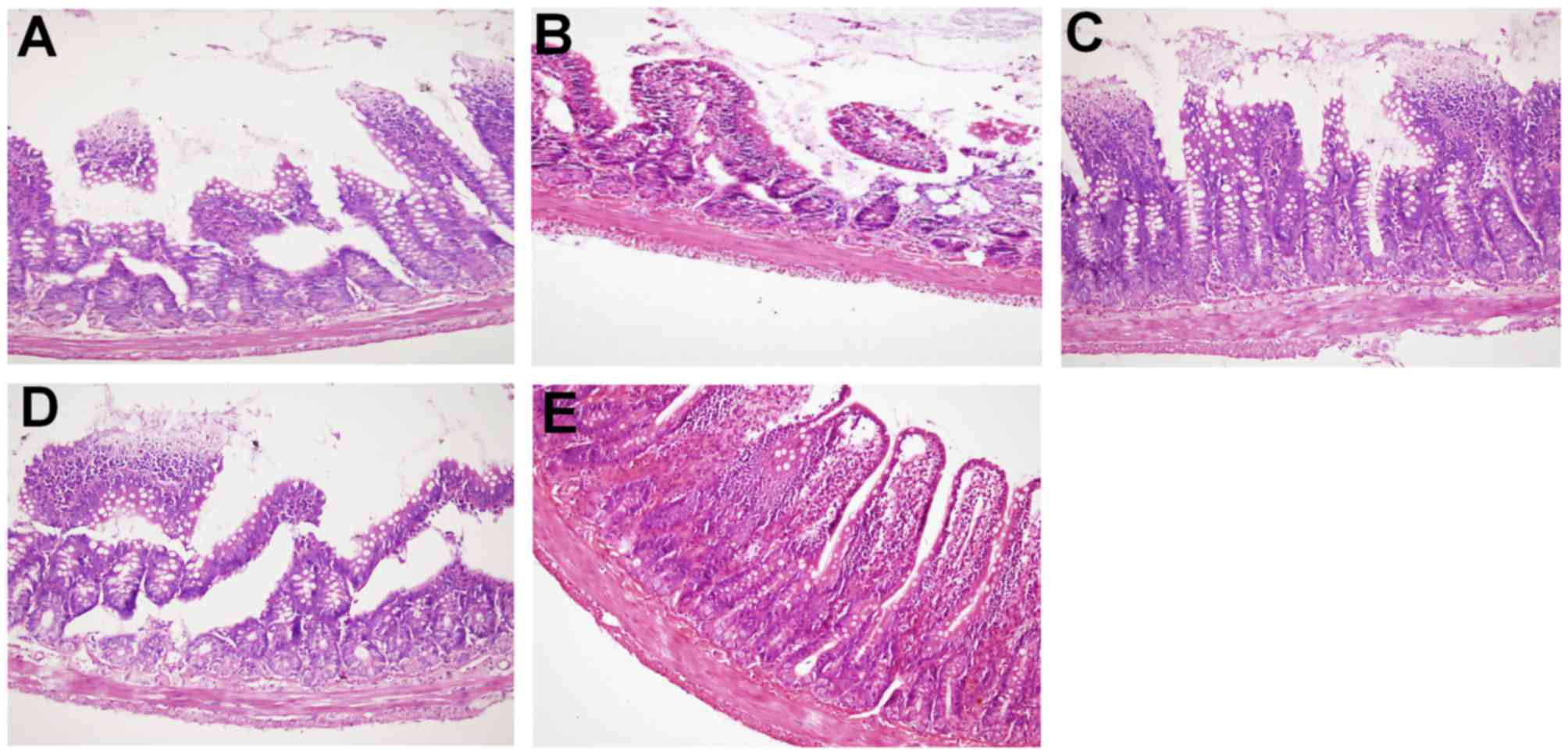
Histological changes in ileal
tissues
Intestinal mucosal damage was ameliorated in the BSP
treatment groups, with less intestinal wall structure destruction
and more villi observed (Fig. 6A-C).
These effects were most notable in the 60 mg/kg BSP group (Fig. 6C). In the experiment group (Fig. 6D), the structure of the intestinal
wall was damaged and the intestinal villus was fractured, shortened
and atrophic. Histopathology of the intestinal mucosa in the
control group revealed that the intestinal layer had a clear
structure with intact surface epithelium and neatly arranged
intestinal villus (Fig. 6E).
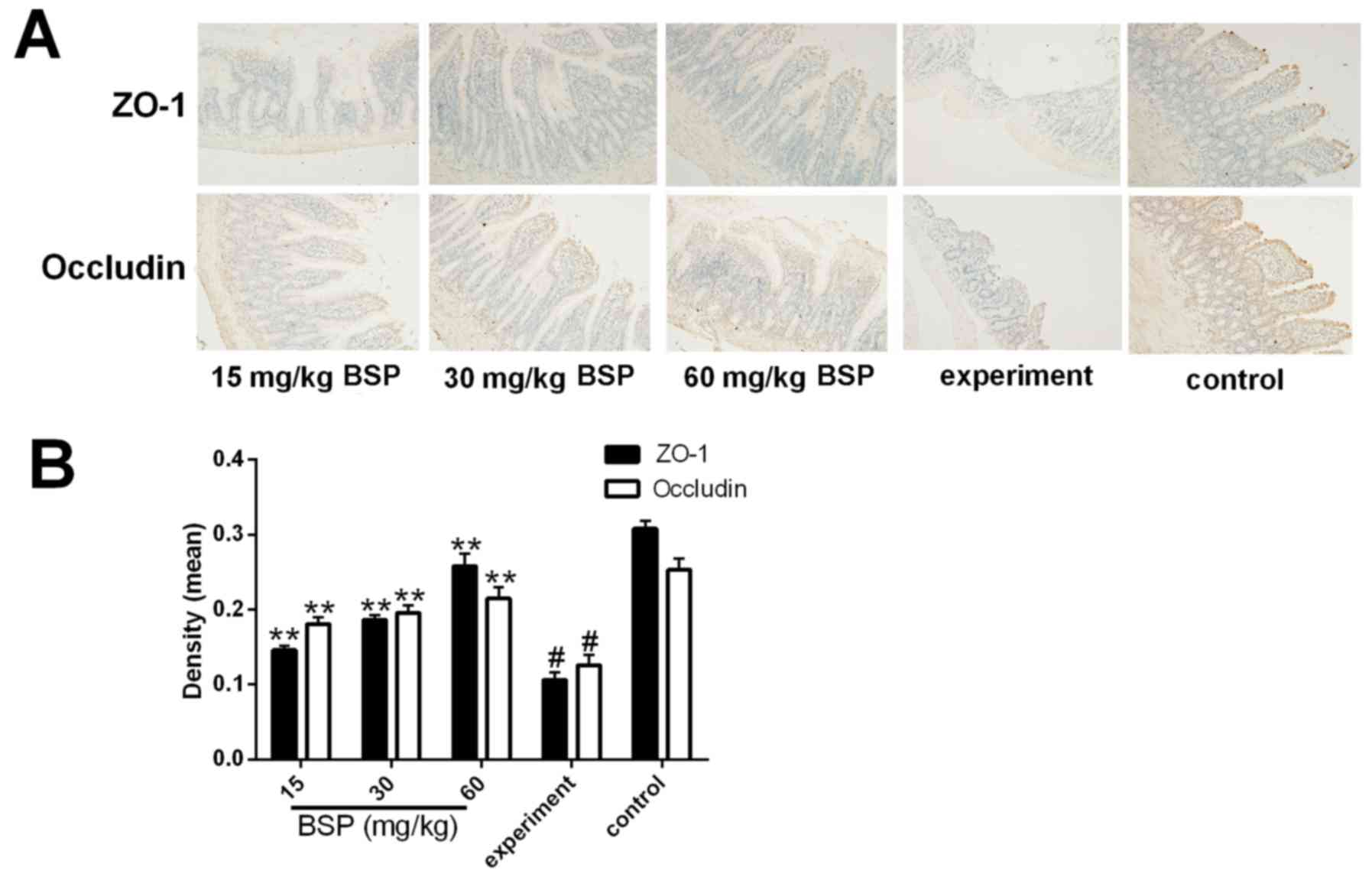
Effect of BSP on ZO-1 and occludin
protein expression in intestinal tissues
IHC was performed and the results demonstrated that
the control group was positively stained for ZO-1 and occludin,
with dark brown coloration (Fig.
7A). In the experiment group, however, positive ZO-1 and
occludin staining was significantly decreased (Fig. 7A and B). The degree of positive
staining observed in the BSP treatment groups was significantly
increased in compared with the experiment group (Fig. 7A and B).
Discussion
Cirrhosis is a common chronic progressive liver
disease, which is caused by one or more long-term or repeated
effects, including chronic viral infection (hepatitis B and C),
immunologic attack and parasitic diseases (24). The significance of intestinal
epithelial barrier dysfunction as a critical cofactor in liver
cirrhosis has been reported over the past decades (25,26).
Intestinal epithelial cells prevent the invasion of unwelcome
microorganisms, while LPS-induced intestinal barrier dysfunction
leads to an increase in intestinal mucosal permeability, which
accelerates the release of inflammatory factors (27) and intestinal cytokines, resulting in
expedited intestinal damage and inflammation (28). IL-6 and TNF-α are the critical
cytokines associated with inflammation and are common treatment
targets for inflammatory diseases, including liver cirrhosis
(29,30). TNF-α serves an important role in the
progression of intestinal epithelial barrier injury (31) and is able to initiate the production
of cytokines, including IL-6, which further exacerbates intestinal
epithelial barrier damage (32). A
clinical study reported that TNF-α and IL-6 were significantly
upregulated in patients with cirrhosis (33). Taken together, these results suggest
that inhibiting the expression of inflammatory factors may be an
effective method for protecting against intestinal endotoxemia.
In the present study, intestinal IL-6 and TNF-α were
upregulated in a TAA-induced cirrhosis rat model and this effect
was inhibited by BSP treatment. These results suggest that
cirrhosis is associated with the overexpression of inflammatory
cytokines and that BSP is able to effectively ameliorate this.
TJs are localized at the apical end of lateral
membranes and are some of the most important structures responsible
for preventing toxic substances from the intestine penetrating the
surrounding tissue through the epithelial cell layer (34,35). TJ
proteins are located between epithelial cells and comprise a
variety of transmembrane and membrane-associated proteins (36), including ZO-1 and occludin. The
primary function of these molecules is to act as a selective
intercellular barrier to regulate the diffusion of macromolecules
(37). Occludin is a
four-transmembrane protein, comprising four putative
membrane-spanning segments (38),
which is exclusively localized at TJs in epithelial and endothelial
cells (39). It has been reported
that high occludin expression is correlated with low endothelium
permeability (40). Another study
demonstrated that downregulating the expression of occludin results
in increased intestinal permeability, allowing bacteria, endotoxins
and other macromolecules to enter the body (41). As such, measuring occludin levels may
reflect the integrity of the intestinal barrier (42–44).
ZO-1 was the first TJ protein to be discovered (45) and is directly associated with
occludin, serving an important role in regulating TJ barrier
function (46). It has been reported
that LPS is able to decrease the expression of ZO-1 and occludin
(47), which is consistent with the
results of the present study.
In the present study, it was demonstrated that BSP
is able to upregulate the expression of ZO-1 and occludin in a
TAA-induced cirrhosis rat model. BSP is also able to reduce
endotoxin levels and decrease the degree of intestinal mucosa
damage. Taken together, these results suggest that BSP is able to
preserve the barrier function of the epithelium by keeping TJs
intact.
In summary, the results of the present study
indicate that BSP is able to protect against intestinal epithelial
barrier disruption in a TAA-induced liver cirrhosis rat model via
reducing endotoxin levels, inhibiting inflammatory cytokine
expression and improving TJs, suggesting that BSP may have
potential as a treatment for intestinal endotoxemia. However, it is
important to elucidate the underlying mechanism of BSP in
intestinal endotoxemia. Future studies on the intestinal
endotoxemia mechanisms of BSP may give a more complete
understanding and further the development of novel methods to
prevent and treat intestinal endotoxemia.
Acknowledgements
Not applicable.
Funding
The present study was supported by The Second Batch
of Young Middle-aged Talents in Wuhan.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FY and YW designed and directed the experiment. LL,
XC and YL performed the experiments. ZZ, JX and JZ performed the
statistical analysis. LL wrote the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by The Institutional Animal
Care and Use Committee of Tongji Medical College, Huazhong
University of Science and Technology (Wuhan, China). All animal
experiment protocols used in the present study followed
internationally accepted principles.
Patient consent for publication
Not applicable.
Competing interests
The authors have no potential competing interests to
disclose.
References
|
1
|
Gogoi-Tiwari J, Köhn-Gaone J, Giles C,
Schmidt-Arras D, Gratte ED, Elsegood CL, McCaughan GW, Ramm GA,
Olynyk JK and Tirnitz-Parker JEE: The murine choline-deficient,
ethionine-supplemented (CDE) diet model of chronic liver injury. J
Vis Exp. Oct 21–2017. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang H, Zhang W, Zuo L, Zhu W, Wang B, Li
Q and Li J: Bifidobacteria may be beneficial to intestinal
microbiota and reduction of bacterial translocation in mice
following ischaemia and reperfusion injury. Br J Nutr.
109:1990–1998. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Arrieta MC, Bistritz L and Meddings JB:
Alterations in intestinal permeability. Gut. 55:1512–1520. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Groschwitz KR and Hogan SP: Intestinal
barrier function: Molecular regulation and disease pathogenesis. J
Allergy Clin Immunol. 124:3–20. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Turner JR: Intestinal mucosal barrier
function in health and disease. Nat Rev Immunol. 9:799–809. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Clark JA, Doelle SM, Halpern MD, Saunders
TA, Holubec H, Dvorak K, Boitano SA and Dvorak B: Intestinal
barrier failure during experimental necrotizing enterocolitis:
Protective effect of EGF treatment. Am J Physiol Gastrointest Liver
Physiol. 291:G938–G949. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pijls KE, Koek GH, Elamin EE, de Vries H,
Masclee AA and Jonkers DM: Large intestine permeability is
increased in patients with compensated liver cirrhosis. Am J
Physiol Gastrointest Liver Physiol. 306:G147–G153. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Andreasen AS, Krabbe KS, Krogh-Madsen R,
Taudorf S, Pedersen BK and Moller K: Human endotoxemia as a model
of systemic inflammation. Curr Med Chem. 15:1697–1705. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Marshall JC, Walker PM, Foster DM, Harris
D, Ribeiro M, Paice J, Romaschin AD and Derzko AN: Measurement of
endotoxin activity in critically ill patients using whole blood
neutrophil dependent chemiluminescence. Crit Care. 6:342–348. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yoshioka N, Taniguchi Y, Yoshida A, Nakata
K, Nishizawa T, Inagawa H, Kohchi C and Soma G: Intestinal
macrophages involved in the homeostasis of the intestine have the
potential for responding to LPS. Anticancer Res. 29:4861–4865.
2009.PubMed/NCBI
|
|
11
|
Cong X, Zhang Y, Yang NY, Li J, Ding C,
Ding QW, Su YC, Mei M, Guo XH, Wu LL and Yu GY: Occludin is
required for TRPV1-modulated paracellular permeability in the
submandibular gland. J Cell Sci. 126:1109–1121. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nolan JP: The role of intestinal endotoxin
in liver injury: A long and evolving history. Hepatology.
52:1829–1835. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Trebicka J, Krag A, Gansweid S, Appenrodt
B, Schiedermaier P, Sauerbruch T and Spengler U: Endotoxin and
tumor necrosis factor-receptor levels in portal and hepatic vein of
patients with alcoholic liver cirrhosis receiving elective
transjugular intrahepatic portosystemic shunt. Eur J Gastroenterol
Hepatol. 23:1218–1225. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Adachi Y, Moore LE, Bradford BU, Gao W and
Thurman RG: Antibiotics prevent liver injury in rats following
long-term exposure to ethanol. Gastroenterology. 108:218–224. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Diao H, Li X, Chen J, Luo Y, Chen X, Dong
L, Wang C, Zhang C and Zhang J: Bletilla striata polysaccharide
stimulates inducilble nitric oxide synthase and proinflammatory
cytokine expression in macrophages. J Biosci Bioeng. 105:85–89.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang C, Sun J, Luo Y, Xue W, Diao H, Dong
L, Chen J and Zhang J: A polysaccaride isolated from the medicinal
herb Bletilla striata induces endothelial cells proliferation and
vascular endothelial growth factor expression in vitro. Biotechnol
Lett. 28:539–543. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Y, Liu J, Li Q, Wang Y and Wang C:
Two natural glucomannan polymers, from Konjac and Bletilla, as
bioactive materials for pharmaceutical applications. Biotechnol
Lett. 37:1–8. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Luo Y, Diao H, Xia S, Dong L, Chen J and
Zhang J: A physiologically active polysaccharide hydrogel promotes
wound healing. J Biomed Mater Res A. 94:193–204. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dong L, Xia S, Luo Y, Diao H and Zhang J,
Chen J and Zhang J: Targeting delivery oligonucleotide into
macrophages by cationic polysaccharide from Bletilla striata
successfully inhibited the expression of TNF-alpha. J Control
Release. 134:214–220. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Y, Lv T, Li M, Xue T, Liu H, Zhang
W, Ding X and Zhuang Z: Anti-aging effect of polysaccharide from
Bletilla striata on nematode Caenorhabditis elegans. Pharmacogn
Mag. 11:449–454. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang Y, Liu D, Chen S, Wang Y, Jiang H and
Yin H: A new glucomannan from Bletilla striata: Structural and
anti-fibrosis effects. Fitoterapia. 92:72–78. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Yang F, Xue J, Zhou X, Luo L, Ma
Q, Chen YF, Zhang J, Zhang SL and Zhao L: Anti-schistosomiasis
liverfibrosis effects of chlorogenic acid through
IL-13/miR-21/Smad7 signaling interactions in vivo and in vitro.
Antimicrob Agents Chemother. 61:pii: e01347. –16. 2017.
|
|
24
|
Tsochatzis EA, Bosch J and Burroughs AK:
Liver cirrhosis. Lancet. 383:1749–1761. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Malaguarnera G, Giordano M, Nunnari G,
Bertino G and Malaguarnera M: Gut microbiota in alcoholic liver
disease: Pathogenetic role and therapeutic perspectives. World J
Gastroenterol. 20:16639–16648. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Brun P, Castagliuolo I, Di Leo V, Buda A,
Pinzani M, Palù G and Martines D: Increased intestinal permeability
in obese mice: New evidence in the pathogenesis of nonalcoholic
steatohepatitis. Am J Physiol. 292:G518–G525. 2007.
|
|
27
|
Caine WR, Metzler-Zebeli BU, McFall M,
Miller B, Ward TL, Kirkwood RN and Mosenthin R: Supplementation of
diets for gestating sows with zinc amino acid complex and gastric
intubation of suckling pigs with zinc-methionine on mineral status,
intestinal morphology and bacterial translocation in
lipopolysaccharide-challenged early-weaned pigs. Res Vet Sci.
86:453–462. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Song HL, Lv S and Liu P: The roles of
tumor necrosis factor-α in colon tight junction protein expression
and intestinal mucosa structure in a mouse model of acute liver
failure. BMC Gastroenterol. 9:702009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang DH, Ye ZY, Jin B, He XJ, Zhang Q,
Zhou WM, Xu WJ and Lu HX: Salvianolate inhibits cytokine gene
expression in small intestine of cirrhotic rats. World J
Gastroenterol. 17:1903–1909. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shen L, Weber CR, Raleigh DR, Yu D and
Turner JR: Tight junction pore and leak pathways: A dynamic duo.
Annu Rev Physiol. 73:283–309. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang F, Graham WV, Wang Y, Witkowski ED,
Schwarz BT and Turner JR: Interferon-gamma and tumor necrosis
factor-alpha synergize to induce intestinal epithelial barrier
dysfunction by up-regulating myosin light chain kinase expression.
Am J Pathol. 166:409–419. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Muñoz L, Borrero José M, Ubeda M, Lario M,
Díaz D, Francés R, Monserrat J, Pastor O, Aguado-Fraile E, Such J,
et al: Interaction between intestinal dendritic cells and bacteria
translocated from the gut in rats with cirrhosis. Hepatology.
56:1861–1869. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Toda K, Kumagai N, Tsuchimoto K, Inagaki
H, Suzuki T, Oishi T, Atsukawa K, Saito H, Morizane T, Hibi T and
Ishii H: Induction of hepatic stellate cell proliferation by
LPS-stimulated peripheral blood mononuclear cells from patients
with liver cirrhosis. J Gastroenterol. 35:214–220. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mitic LL, Van Itallie CM and Anderson JM:
Molecular physiology and pathophysiology of tight junctions I.
Tight junction structure and function: Lessons from mutant animals
and proteins. Am J Physiol Gastrointest Liver Physiol.
279:G250–G254. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Anderson JM and Van Itallie CM: Physiology
and function of the tight junction. Cold Spring Harb Perspect Biol.
1:a0025842009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gonzalez-Mariscal L, Betanzos A, Nava P
and Jaramillo BE: Tight junction protein. Prog Biophys Mol Biol.
81:1–44. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hu YJ, Wang YD, Tan FQ and Yang WX:
Regulation of paracellular permeability: Factors and mechanisms.
Mol Biol Rep. 40:6123–6142. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Furuse M, Hirase T, Itoh M, Nagafuchi A,
Yonemura S and Tsukita S and Tsukita S: Occludin: A novel integral
membrane protein localizing at tight junctions. J Cell Biol.
123:1777–1788. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Saitou M, Ando-Akatsuka Y, Itoh M, Furuse
M, Inazawa J, Fujimoto K and Tsukita S: Mammalian occludin in
epithelial cells: Its expression and subcellular distribution. Eur
J Cell Biol. 73:222–231. 1997.PubMed/NCBI
|
|
40
|
Li J, Ge R, Zhao C, Tang L, Li J and Li Q:
Farrerol regulates occludin expression in hydrogen peroxide-induced
EA.hy926 cells by modulating ERK1/2 activity. Eur J Pharmacol.
734:9–14. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen D, Li L, Yan J, Yang X, You Y, Zhou Y
and Ling X: The loss of αSNAP downregulates the expression of
occludin in the intestinal epithelial cell of acute pancreatitis
model. Pancreatology. 14:347–355. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lapierre LA: The molecular structure of
the tight junction. Adv Drug Deliv Rev. 41:255–264. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Berkes J, Viswanathan VK, Sarkovic SD and
Hecht G: Intestinal epithelial responses to enteric pathogens:
Effects on the tight junction barrier, ion transport, and
inflammation. Gut. 52:439–451. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Beltinger J, McKaig BC, Makh S, Stack WA,
Hawkey CJ and Mahida YR: Human colonic subepithelial myofibroblasts
modulate transepithelial resistance and sceretory response. Am J
Physiol. 277:C271–C279. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Stevenson BR, Siliciano JD, Mooseker MS
and Goodenough DA: Identification of ZO-1: A high molecular weight
polypeptide associated with the tight junction (zonula occludens)
in a variety of epithelia. J Cell Biol. 103:755–766. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cario E, Gerken G and Podolsky DK:
Toll-like receptor 2 enhances ZO-1-associated intestinal epithelial
barrier integrity via protein kinase C. Gastroenterology.
127:224–238. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chen J, Zhang R, Wang J, Yu P, Liu Q, Zeng
D, Song H and Kuang Z: Protective effects of baicalin on
LPS-induced injury in intestinal epithelial cells and intercellular
tight junctions. Can J Physiol Pharmacol. 93:233–237. 2015.
View Article : Google Scholar : PubMed/NCBI
|