Introduction
Mesenchymal stem cells (MSCs) originate from
mesoderm, and are a type of pluripotent cells with a great
proliferation and multi-direction differentiation potential in
vitro (1). The first successful
isolation was that of bone marrow mesenchymal stem cells (BMSCs)
and later from various other tissue types, including subcutaneous
adipose tissue (2–5). Numerous independent studies have
demonstrated that BMSCs are multipotent in an artificial
environment, with the ability to produce cartilage cells,
osteoblasts, supporting hematopoietic cells, adipocytes and
hepatocyte-like cells (6–9). Subsequent studies indicated that
adipose-derived stem cells (ADSCs) have similar biological
properties compared with BMSCs (10). Subcutaneous adipose tissue displays
certain advantages over bone marrow, e.g., adipose tissue is easier
to collect and enrich, and is located at sites from which patients
are more likely to accept biopsies (11,12).
Therefore, ADSCs have become a better option for medicinal
applications including tissue engineering and replacement
treatment.
To date, ADSCs have been mostly isolated from
mammals, including humans, rhesus monkeys, mice and rabbits
(13–16), but rarely from avian species.
Plentiful adipose tissues are present in broilers (Gallus gallus
domesticus). In the present study, ADSCs were obtained from
20-day-old broiler embryos and cultured in vitro, their
abilities to proliferate and differentiate into multiple lineages
were demonstrated, and they were characterized regarding the
expression of specific surface markers. The present study offers a
novel source for in vitro culturing of ADSCs and lays a
foundation for tissue engineering and regenerative medicinal
applications.
Materials and methods
Experimental animals
A total of 60 20-day-old male broiler embryos
(weight, ~40 g) were provided by the Poultry Experimental Base of
the Institute of Animal Sciences (Chinese Academy of Agricultural
Sciences, Beijing, China). All procedures conformed to the
guidelines established by the Institutional Animal Care and Use
Committee at Chinese Academy of Agriculture Sciences (Beijing,
China).
Reagents
The following reagents were used in the present
study: Dulbecco's modified Eagle's medium (DMEM)/F12, fetal bovine
serum (FBS), L-glutamine, and penicillin and streptomycin (all from
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) trypsin
(dilution, 1:250) and 0.02% (w/v) EDTA (Amresco, Inc., Framingham,
MA, USA), rabbit anti-chicken CD29 (bs-0486R), CD44 (bs-2507R),
CD71 (bs-1782R) and CD73 (bs-4834R) polyclonal primary antibody,
goat serum, EDTA, fluorescein isothiocyanate (FITC)-conjugated
goat-anti-rabbit immunoglobulin (Ig)G (all from Bioss, Beijing,
China), ascorbic acid sodium salt, dexamethasone, hepatocyte growth
factor, indometacin, insulin transferrin-selenium,
3-isobutyl-1-methylxanthine (IBMX), β-glycerophosphate, oil red O,
collagenase I (all from Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany), basic fibroblast growth factor (bFGF), fibroblast growth
factor-4 (FGF-4; both from Peprotech, Inc., Rocky Hill, NJ, USA),
alizarin red (Boster, Wuhan, China) and TRIzol (Invitrogen; Thermo
Fisher Scientific, Inc.).
Isolation and culture of ADSCs
Adipose tissue samples were collected from
20-day-old broiler embryos under aseptic conditions. The adipose
pads were washed with PBS supplemented with 100 IU/ml penicillin
and 100 µg/ml streptomycin to remove any other cells, including
hemocytes and endothelial cells. The adipose pats were fully
chopped into small pieces and subsequently incubated with 0.2%
(m/v) collagenase I in PBS at 37°C for 40 min. The enzymatic
activity was neutralized by adding an equal volume of DMEM/F12
containing 5% (v/v) FBS. The cell suspension was filtered through a
74-mm-mesh sieve and then centrifuged at 200 × g for 8 min at room
temperature. The precipitate was resuspended in complete growth
medium composed of DMEM/F12, 10% (v/v) FBS, 2 mM L-glutamine, 100
IU/ml penicillin, 100 µg/ml streptomycin and 10 ng/ml bFGF. Cells
were seeded into petri dishes at a density of
1.0×105/ml, and cultured at 37°C with 5% CO2.
After 24 h, the dishes were washed with PBS to clean out any
non-adherent cells, including pericytes, blood cells, endothelial
cells and preadipocytes (14,17–19).
The cells were referred to as ‘passage 0’ (P0) when
their confluence reached 80%. Subsequently, the cells were
sub-cultured at the ratio of 1:2 after standard trypsin digestion.
This generation was referred to as P1. The cells became purified
with increasing passages, and were then harvested for other
relevant trials.
Morphological observation
The morphology and adhesion of ADSCs prior to and
after culture was observed under an inverted microscope.
Reverse transcription polymerase chain
reaction (RT-PCR) analysis of cell surface markers
Total RNA was extracted from ADSCs at P5 by using
TRIzol and then subjected to RT using an RNA PCR kit (version 3.0;
Takara Biotechnology, Co., Ltd., Dalian, China) according to the
manufacturer's instructions. Primers were designed in accordance
with the sequences of GAPDH (internal control) CD71, CD29, CD31,
CD73 and CD44 from GenBank. The template complementary DNA was
amplified by PCR using the gene-specific primers listed in Table I. The PCR reaction was performed
using the PCR Master Mix kit (Takara Biotechnology, Co., Ltd.),
according to the manufacturer's instructions. PCR analyses were
performed in 50 µl reactions, containing 25 µl 2X PCR mix, 15 µl
ddH2O, 5 µl template cDNA and 2.5 µl forward and reverse
primers. Cycling conditions were as follows: Initialization at 94°C
for 5 min, then 35 cycles of a denaturation at 94°C for 30 sec,
annealing at 50–60°C for 30 sec, elongation at 72°C for 30 sec and
final elongation at 72°C for 5 min. PCR products were assessed by
2% agarose gel electrophoresis and bands were visualized with an
ultraviolet transilluminator.
 | Table I.Sequences of primers used for
polymerase chain reaction. |
Table I.
Sequences of primers used for
polymerase chain reaction.
| Gene | Primer sequence | Tm (°C) | Cycles (n) | Product size
(bp) |
|---|
| CD29 | F,
5′-GAACGGACAGATATGCAACG-3′ | 60 | 30 | 300 |
|
| R,
5′-TAGAACCAGCAGTCACCAACG-3′ |
|
|
|
| CD31 | F,
5′-CAGGCAAAGGAGACGCACGAT-3′ | 60 | 30 | 221 |
|
| R,
5′-CTTCTGGCAGCTCACAACGT-3′ |
|
|
|
| CD44 | F,
5′-CATCGTTGCTGCCCTCCT-3′ | 56 | 30 | 290 |
|
| R,
5′-ACCGCTACACTCCACTCTTCAT-3′ |
|
|
|
| CD71 | F,
5′-CTCCTTTGAGGCTGGTGAGG-3′ | 56 | 30 | 293 |
|
| R,
5′-TCAGTGAAGCCACGACCTTC-3′ |
|
|
|
| CD73 | F,
5′-AGTGCAAACATTAAGGGAAAA-3′ | 58 | 30 | 310 |
|
| R,
5′-ACGCTCCTGGAAGATAGTGAT-3′ |
|
|
|
| GAPDH | F,
5′-TAAAGGCGAGATGGTGAAAG-3′ | 60 | 30 | 244 |
|
| R,
5′-ACGCTCCTGGAAGATAGTGAT-3′ |
|
|
|
| PPAR-γ | F,
5′-CTGTCTGCGATGGATGAT-3′ | 48 | 30 | 199 |
|
| R,
5′-AATAGGGAGGAGAAGGAG-3′ |
|
|
|
| LPL | F,
5′-AGTGAAGTCAGGCGAAAC-3′ | 49 | 30 | 477 |
|
| R,
5′-ACAAGGCACCACGATT-3′ |
|
|
|
| Collagen I | F,
5′-AAGGATGGTCGCAATG-3′ | 49 | 30 | 310 |
|
| R,
5′-GGTGGCTAAGTCTGAGGT-3′ |
|
|
|
| Osteopontin | F,
5′-CAGAACAGCCGGACTTTC-3′ | 51 | 30 | 227 |
|
| R,
5′-CTTGCTCGCCTTCACCAC-3′ |
|
|
|
| ALB | F,
5′-AGACAGACGCATGGCTTGTT-3′ | 60 | 30 | 287 |
|
| R,
5′-GGGGCTTGCGTTTAATGAGG-3′ |
|
|
|
| AFP | F,
5′-TCGGGCACGCTTGATCTTTA-3′ | 62 | 30 | 499 |
|
| R,
5′-AGCTGTTGCCTTCAACTGGA-3′ |
|
|
|
Immunofluorescent detection
ADSCs of P5 were fixed in 4% (m/v) paraformaldehyde
for 15 min at room temperature, subsequently washed with PBS. The
cells were permeabilized using 0.2% (v/v) Triton X-100 for 20 min
at room temperature and washed with PBS. The samples were blocked
with 10% (v/v) goat serum for 30 min at room temperature, and then
incubated in 1% bovine serum albumin (BSA) in PBS containing the
polyclonal rabbit anti-chicken antibodies to CD29, CD44, CD71 and
CD73 (all at 1:100 dilution) for 12 h at 4°C. Primary antibody was
replaced with PBS for the negative control. The samples were washed
with PBS and incubated in PBS containing FITC-conjugated goat
anti-rabbit IgG as the secondary antibody for 1 h at room
temperature. The samples were washed with PBS after the incubation.
They were counterstained with DAPI and were visualized by using a
Nikon TE-2000-E confocal microscope with a digital camera system
(Nikon, Tokyo, Japan).
Flow cytometry
The expression of cell-associated surface markers
was measured by flow cytometric analysis. In brief, ADSCs of P5
were collected by standard trypsin digestion, and the cells were
fixed and permeabilized in 70% (v/v) ice-cold ethanol for 12 h. The
samples were blocked with 10% (v/v) goat serum for 30 min at room
temperature and were incubated in 1% BSA containing rabbit
anti-chicken polyclonal antibodies to CD29, CD44, CD71 and CD73
(all at 1:100 dilution) for 1 h at room temperature. Primary
antibody was replaced with PBS for the negative control.
Subsequently, the samples were washed with PBS and incubated in PBS
containing FITC-conjugated goat anti-rabbit IgG as the secondary
antibody for 1 h at room temperature. After washing with PBS, the
labeled cells were detected with a flow cytometer. Flow cytometric
data were analyzed using CXP software version 2.0 (Beckman Coulter,
Brea, CA, USA).
Growth kinetics
ADSCs at P3, P16 and P30 were collected and seeded
in triplicate in 24-well plates at 1×104 cells/well.
Cell viability assessment was performed following trypan blue
staining at room temperature for 5 min and cells were counted
following detachment over 9 successive days. The population
doubling time (PDT) was calculated using the following formula:
PDT=(t-t0)xlg2/(lgNt-lgN0), with t0 representing the starting time
of culture, t the termination time of culture, N0 the initial
number of cultured cells and Nt the final number of cells.
Adipogenic differentiation of
ADSCs
ADSCs were divided into a control group and an
induction group. When the confluence of cells reached 70–80%, the
experimental group was treated with adipogenic medium consisting of
DMEM/F12 containing 10% FBS, 200 µM indomethacin, 1 mM
dexamethasone, 10 µM insulin transferrin and 0.5 mM IBMX. The
control group was cultured in complete growth medium without the
factors mentioned above. The medium was replaced every 2 days.
After 1 week, the two groups were evaluated for accumulation of
intracellular lipid using oil red O staining for 40 min at room
temperature and expression of adipogenic cell-specific genes by
RT-PCR.
Osteogenic differentiation of
ADSCs
The cells were divided into two groups as above.
Upon reaching 60–70% confluence, the cells in the experimental
group were cultured in osteogenic medium containing DMEM/F12
supplemented with 10% FBS, 0.1 mM dexamethasone, 10 mM
β-glycerophosphate and 50 µg/ml ascorbate. In the control group,
cells were cultured with normal growth medium. The medium was
replaced every 2 days. After 3 weeks, the formation of calcium
nodes was assessed by alizarin red staining for 25 min at room
temperature and osteoblast-specific genes were detected by
RT-PCR.
Hepatocyte differentiation of
ADSCs
ADSCs were divided into two groups as described
above. When the confluence reached 60–70%, the induction group was
cultured in hepatocyte medium containing DMEM/F12 supplemented with
10% FBS, 20 ng/ml FGF-4, 20 ng/ml hepatocyte growth factor and 1%
insulin transferrin. The control group was cultured in normal
growth medium. The medium was exchanged every 2 days. After 2
weeks, the capacity of the cells to secrete glycogen was detected
by periodic acid Schiff staining for 30 min at 37°C, and
hepatocyte-like cell-specific genes were detected by RT-PCR.
Statistical analysis
Values are expressed as the mean ± standard error of
the mean. Comparisons between multiple groups were evaluated by
Dunnett's post-hoc test after one-way analysis of variance. SPSS
version 19 (IBM Corp., Armonk, NY, USA) was used for statistical
analysis. P<0.05 was considered to indicate a statistically
significant difference.
Results
Culture and morphological observation
of ADSCs
The primary ADSCs obtained from subcutaneous adipose
tissue were seeded into 60-mm Petri dishes, and after 24 h cells
had begun to stretch into various shapes. After 48 h of culturing,
a small number of polygonal cells were also observed under the
inverted microscope, and a minority of cells had irregular shapes
(Fig. 1A). The cells expanded
rapidly and reached 80–90% confluence after 5 days of seeding,
while being arranged in a whirl pattern (Fig. 1B). In primary culture, numerous
hemocytes were admixed with the ADSCs (Fig. 1B). However, from P3, the ADSCs were
fully purified, and the shape was also homogenous; the ADSCs
exhibited a fibroblast-like morphology (Fig. 1C). No difference in morphology was
apparent among the cells of different passages, and the biological
properties had remained constant after several cycles of
sub-culture (Fig. 1D-F). The cells
were cultured to P36 and P37, at which they had a typical senescent
appearance, including vacuoles, tabular shapes and karyopyknosis in
the majority of cells (Fig. 1G and
H). With increasing time, they even started to detach from the
plates.
Identification of the ADSCs
The specific surface markers of ADSCs were detected
by RT-PCR and immunofluorescence. RT-PCR revealed that the ADSCs
were positive for CD29, CD44, CD71 and CD73, and negative for CD31,
while GAPDH served as an internal control (Fig. 2A). Immunofluorescence microscopy
indicated that the ADSC surface antigens CD29, CD44, CD71 and CD73
were expressed by the cells (Fig.
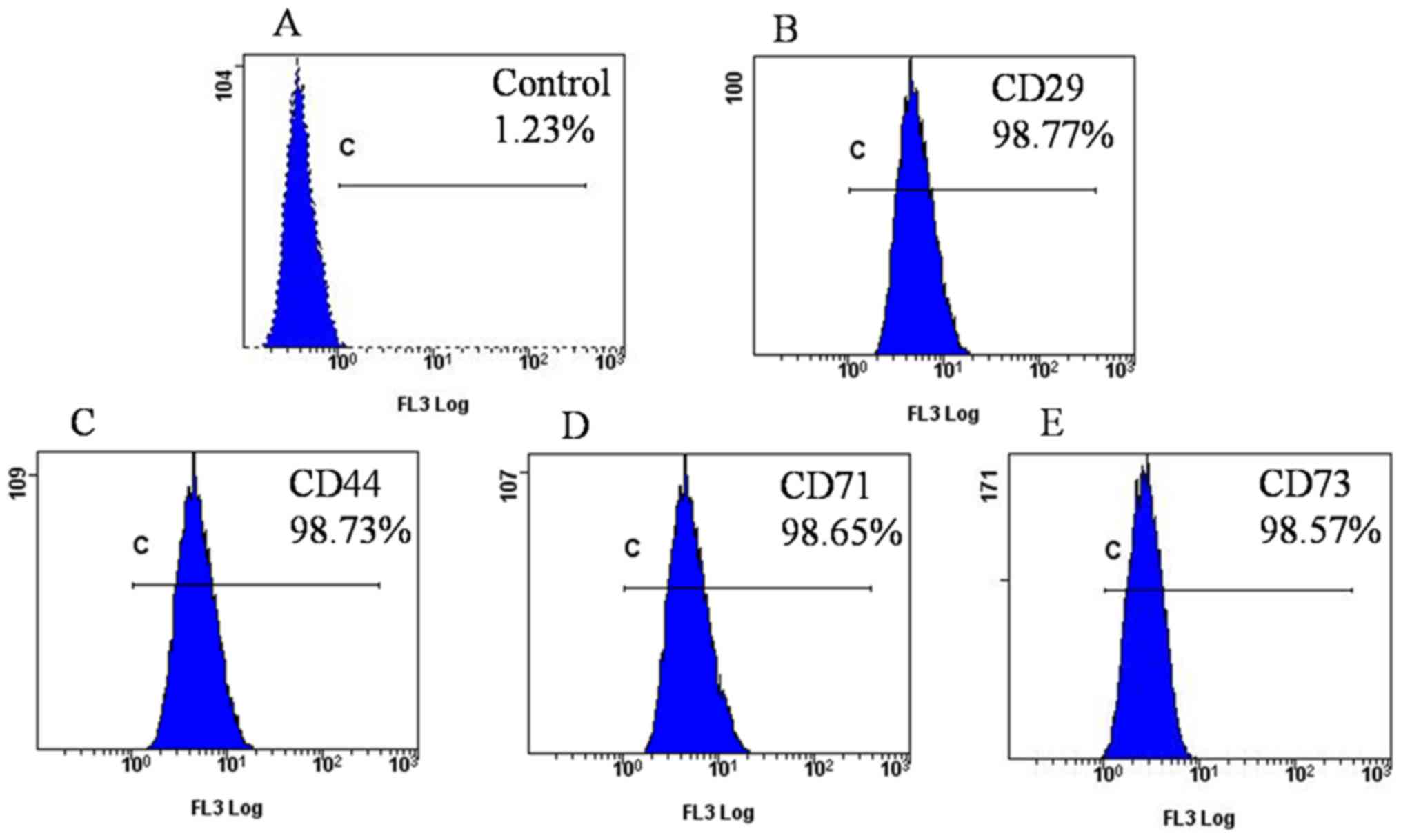
2B). Flow cytometry further confirmed that ADSCs at P5
exhibited high expression of CD29, CD44, CD71 and CD73, as the
positive rates were 98.77, 98.73, 98.65 and 98.57%, respectively
(Fig. 3).
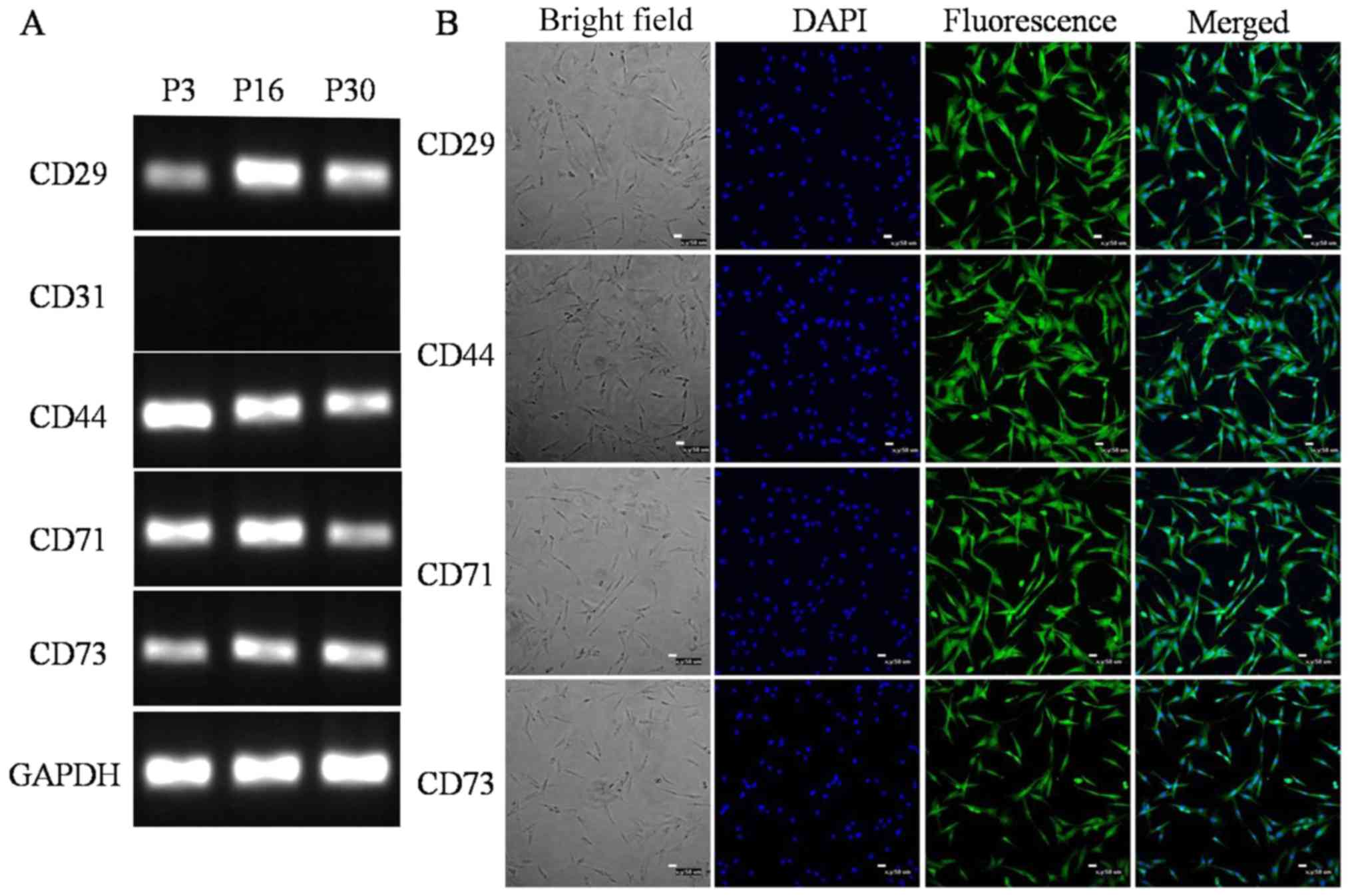 | Figure 2.Surface markers of the ADSCs. Surface
markers of the ADSCs were similar to those of bone marrow
mesenchymal stem cells. The expression of CD29, CD31, CD44, CD71
and CD73 was detected by RT-PCR and immunofluorescence. (A) RT-PCR
analysis indicated that the ADSCs expressed CD29, CD44, CD71 and
CD73, but not CD31. GADPH was used as an internal control. (B)
Immunofluorescence demonstrated immunopositivity for CD29, CD44,
CD71 and CD73 (scale bars, 50 µm). ADSCs, adipose-derived stem
cells; RT-PCR, reverse-transcription polymerase chain reaction
analysis; P3, passage 3. |
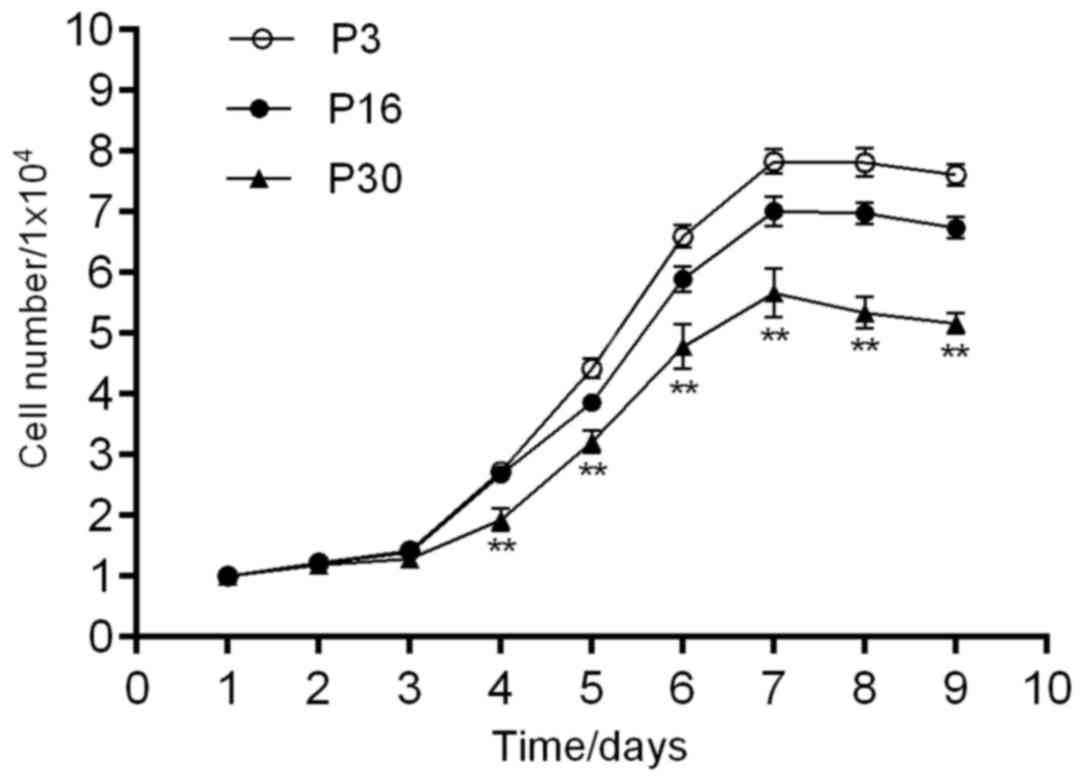
Growth kinetics
The proliferation of ADSCs at P3, P16 and P30 was
detected by determining the cell number with a hemocytometer, and
the data were used to generate a growth curve (Fig. 4). The growth curves suggested that
there was a latency phase of ~24 h, followed by rapid proliferation
of the cells after entering the logarithmic phase. As the density
of the ADSCs increased, the proliferation was inhibited and the
cells reached a growth plateau phase at 7–8 days, after which they
began to degenerate. The growth curves were all typically sigmoidal
and significant differences in the cell growth/kinetics were
identified between the different groups (P<0.05). The PDT was
38.95, 41.27 and 45.05 h for ADSCs at P3, P16 and P30,
respectively.
Adipogenic differentiation of the
ADSCs
Oil Red O staining was used to demonstrate the
adipogenic differentiation of the ADSCs (20). After culture in adipogenic
differentiation medium for 7 days, the morphology of the ADSCs
changed from fibroblast-like to oblate-like and numerous lipid
droplets appeared in the cells/dishes (Fig. 5A). With prolonged induction, the
number of lipid droplets increased gradually and they assembled
into larger droplets. The adipogenic differentiation was confirmed
by oil red O staining, as the induced cells were positive (Fig. 5B), while cells cultured in normal
growth medium were negative on oil red O staining (Fig. 5C).
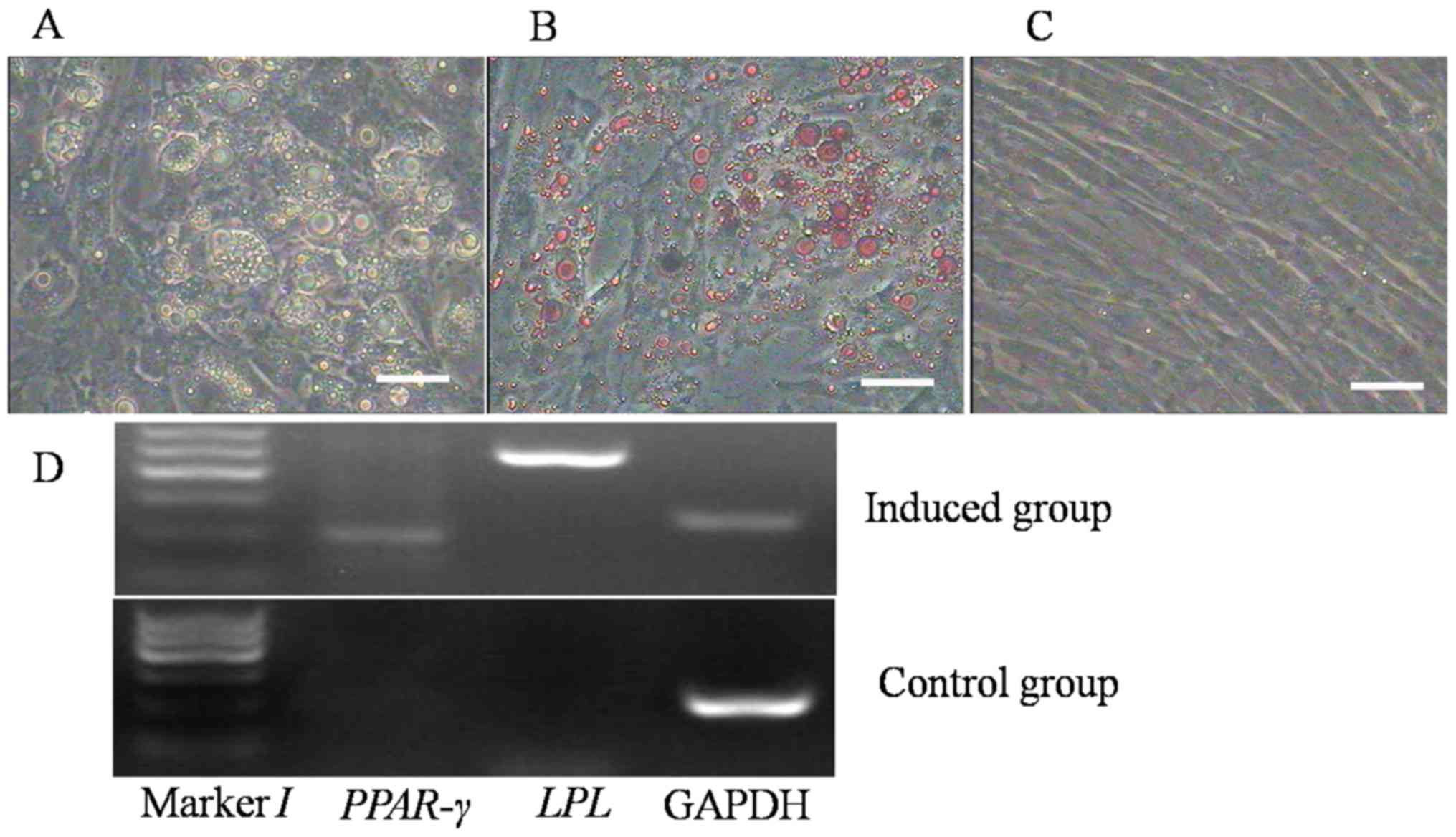 | Figure 5.Adipogenic differentiation of the
ADSCs. (A) After 1 week of induction, the morphology of the ADSCs
began to change from shuttle- to oblate-like, and numerous lipid
droplets had formed in the cells. Along with the extension of
induction time, the amount of droplets increased and they assembled
to form larger ones. (B) Oil red O staining of the differentiated
cells visualized the lipid droplets, indicating that they had
become adipocytes. (C) The cells cultured in complete growth medium
(control group) exhibited no change in morphology and phenotype,
and they were negative on oil red O staining (scale bars, 50 µm).
(D) Reverse-transcription polymerase chain reaction analysis of the
expression of the adipogenic markers LPL and PPAR-γ in the induced
group and the control group. The induced cells expressed LPL and
PPAR-γ, but the control cells did not. PPAR, peroxisome
proliferator-activated receptor; LPL, lipoprotein lipase; ADSCs,
adipose-derived stem cells. |
To further verify the adipogenic differentiation of
the ADSCs, the expression of adipogenic markers was evaluated in
the two groups by RT-PCR. In the ADSCs incubated in differentiation
medium, the adipocyte-specific genes PPAR-γ and LPL were expressed
(Fig. 5D), which did not occur in
the control group.
Osteogenic differentiation of the
ADSCs
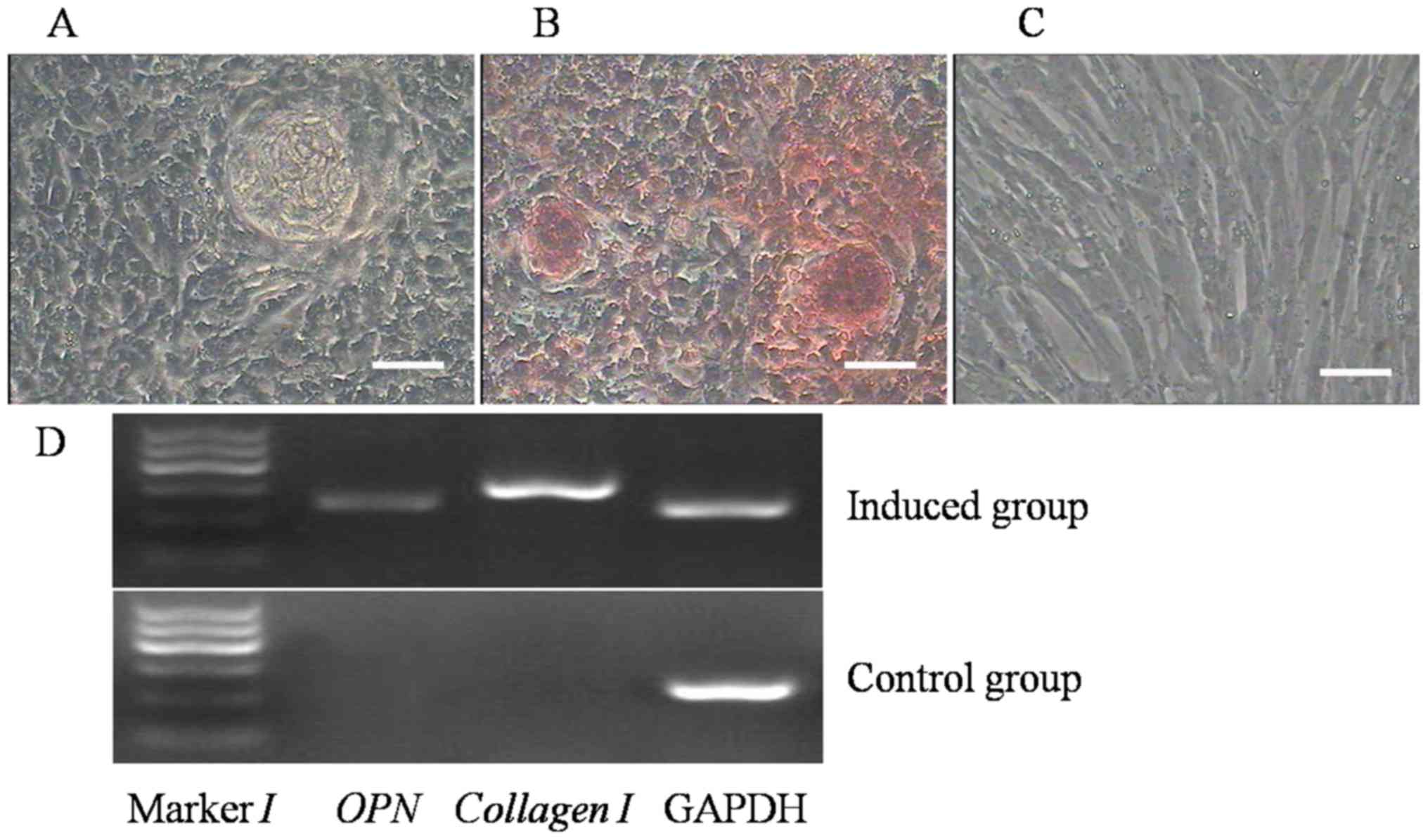
The capacity of broiler ADSCs to differentiate into
osteogenic cells was assessed, and the induced cells were subjected
to morphological and phenotypic analysis. On the 7th day of
induction, the cell shape had changed; the cells became confluent
and formed mineralized nodules, whose size increased after further
induction. After culture in osteogenic medium for 21 days,
morphological changes of the ADSCs were evident and became
polygonal (Fig. 6A). Furthermore,
the nodules were positive on alizarin red staining (Fig. 6B). However, no change in morphology
and no staining with alizarin red were observed in the control
group (Fig. 6C).
To determine that differentiation had occurred, the
expression of osteogenic markers was assessed in the two groups by
RT-PCR. The osteogenic genes OPN and collagen I were expressed in
the induced group, but not in the control group (Fig. 6D).
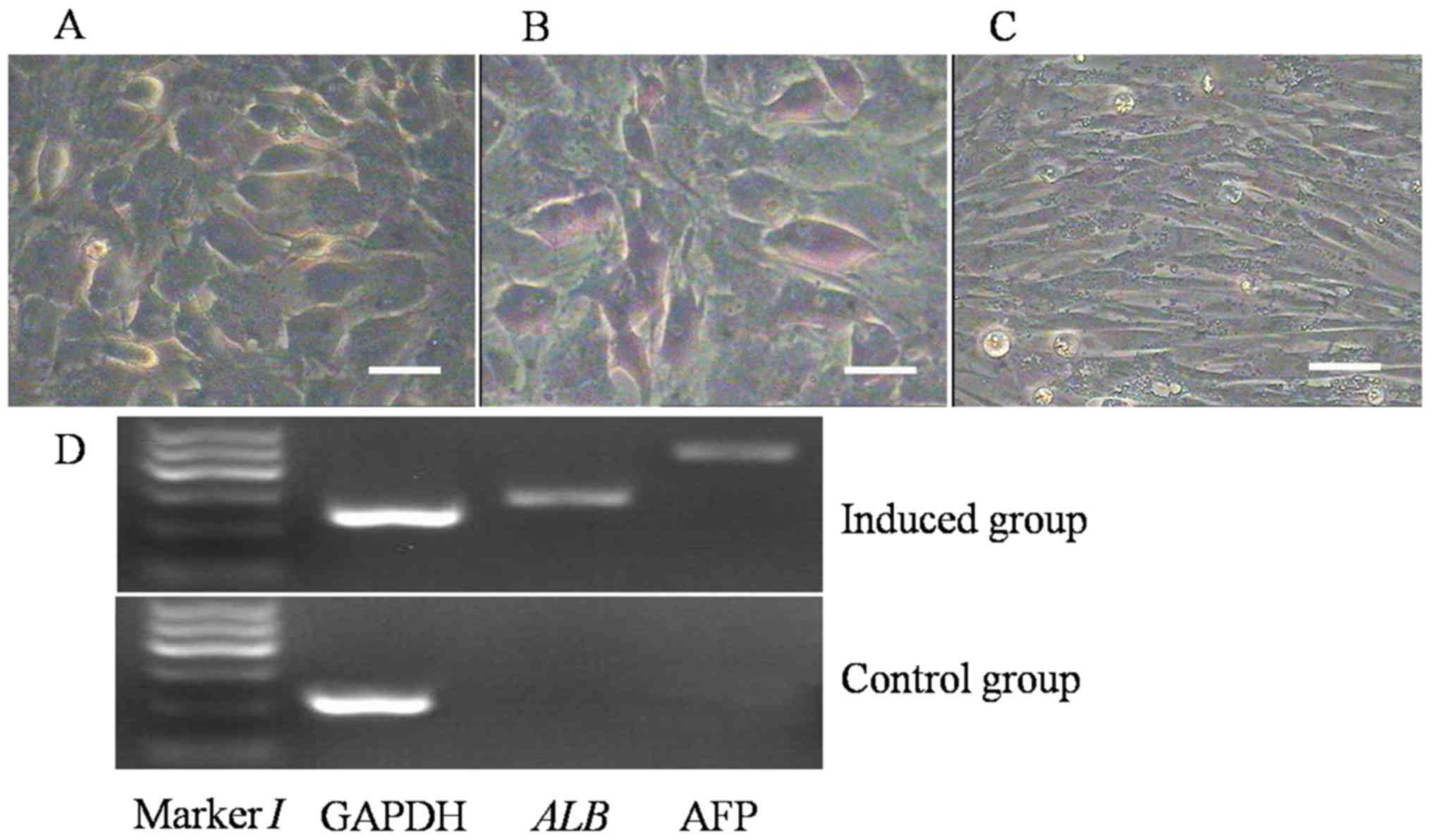
Hepatocyte differentiation
The capacity of broiler ADSCs to differentiate into
hepatocyte-like cells was proven by morphological and phenotypic
analysis of the induced cells. After culturing in hepatocyte
differentiation medium for 7 days, the morphology of certain ADSCs
in the experimental group changed to a round shape from the long
spindle type. After 14 days, numerous cells in the induction group
exhibited a cobblestone-like morphology (Fig. 7A) and were positive on glycogen
staining in the cytoplasm (purple; Fig.
7B), indicating that the induced cells had acquired the
function of synthesis and storage of liver glycogen. In the control
group, no morphological changes and no glycogen staining were
observed (Fig. 7C).
In addition, RT-PCR indicated that in the induction
group, the genes ALB and AFP, which are specific to hepatocyte-like
cells, were expressed, while they were not expressed in the control
group.
Discussion
In the past, adipose tissue was considered to be
merely a passive energy storage modality (21). However, from the opposite viewpoint,
adipose tissue may be regarded as a vital endocrine organ that
controls metabolism, immunity and satiety (22). In stem cell research, a step forward
was made in 2001 when a novel type of adult stem cell, ADSCs, was
first described (1). Later studies
proved that ADSCs have similar immunophenotypic properties and
multilineage differentiation abilities to those of BMSCs (23). Various factors are responsible for
the current lack of studies on avian ADSCs, including insufficient
genetic information and progress compared to other fields and
limited awareness of the scientific community of their therapeutic
potential.
In the present study, ADSCs from the adipose tissues
of 20-day-old broiler chick embryos were successfully obtained
using type I collagenase digestion. All of the results indicated
that the biological characteristics of the newly isolated stem
cells were stable. The ADSCs became homogeneous through
purification over 3–5 passages. The biological properties of the
ADSCs were assessed by cytochemistry, determination of growth
dynamics, detection of specific markers and multi-lineage
differentiation potency experiments.
Almost all of ADSCs were shuttle-shaped with two
ends elongated and proliferated rapidly to form a whirl pattern.
Their growth dynamics were assessed by generating growth curves,
which had typical sigmoidal and indicated a normal population
doubling time.
At present, specific surface markers for ADSCs are
lacking. The identity of ADSCs is generally confirmed by the
expression of certain MSC-specific surface markers, together with
the cell shape and differentiation potential in vitro
(8). In the present study, the
broiler ADSCs expressed CD29, CD44, CD71 and CD73, but not CD31.
CD29 is an integrin subunit correlated to very late antigen
receptor, and forms a heterodimer binding to the surface and
extracellular proteins of MSCs, including CD49 and CD51. It
mediates cell-to-cell and cell-to-matrix adhesion. CD44 is a
cell-surface glycoprotein, and it mediates cell-cell interactions,
cell adhesion and migration (24).
It is a receptor for hyaluronic acid. CD44 is involved in numerous
cellular activities, including recirculation, homing, lymphocyte
activation, metastasis and hematopoiesis (25). CD71, known as a member of the
transferrin receptor family, is a carrier for transferrin. It is
important for cellular iron uptake by the process of
receptor-mediated endocytosis. A low iron concentration promotes
increases in the level of transferrin receptor to elevate cellular
iron uptake, thereby mediating the iron concentration in MSCs.
Thus, the transferrin receptor maintains cellular iron homeostasis
(26). CD73 catalyzes the
transformation of extracellular nucleotides to membrane-permeable
nucleosides (27). As a significant
signaling molecule, the protein has been demonstrated to
participate in purine salvage and the purinergic cascade that
triggers cell metabolism. The results of the present study indicate
that broiler ADSCs are a group of undifferentiated stem/progenitor
cells different from mesenchymal cells.
The pluripotency of stem cells is their most useful
characteristic for cell transplantation therapy. In vitro,
under the influence of certain induction factors, the expression of
certain key genes in the signaling pathways relevant to stem cell
differentiation may change. Consequently, differentiation in
specific directions may be achieved. In the present study, broiler
ADSCs were induced to differentiate into osteoblasts, adipocytes
and hepatocyte-like cells and the expression of genes
characteristic for the corresponding cell types was assessed. The
results demonstrated that various factors were able to induce the
ADSCs to differentiate into different directions, and that the
ADSCs derived from mesoderm were able to differentiate into
endodermal and ectodermal cells. The autologous features of these
stem cells, in combination with their distinct pluripotency and
easy acquirement, make ADSCs an attractive choice for future tissue
engineering and cell-based therapies (28–30).
The above results suggested that broiler ADSCs have
a strong growth ability and the potential to differentiate towards
mesodermal and endodermal lineages. Although the multilineage
differentiation of ADSCs was successful in vitro, there are
certain technical difficulties regarding the utilization of ADSCs
in clinical applications for therapeutic purposes, including a high
rate of rejection and instability after cell transplantation. These
aspects require being taken into account in future studies and
clinical research.
In conclusion, in the present study, ADSCs were
obtained from the adipose tissue of 20-day-old broiler chick
embryos, and their proliferation and differentiation potential was
tested in vitro. Considering that male chicks are a waste
product of the meat and egg industry, the present study offers an
important potential use allowing the sourcing of stem cells and the
potential application of ADSCs as a stem cell material for
regenerative medicine.
Acknowledgements
Not applicable.
Funding
The present study was funded by the National Natural
Science Foundation of China (grant no. 31472099), the Agricultural
Science and Technology Innovation Program (grant no. cxgc-ias-01)
and the project National Infrastructure of Animal Germplasm
Resources (2016).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TL, WP and KW analyzed data and drafted the
manuscript; WP, FC and YW performed cell culturing and PCR
experiments; TL, SZ and YW performed immunofluorescence and flow
cytometry experiments; WG participated in the studies design and
coordination. All authors have read and approved the final version
of the manuscript.
Ethics approval and consent to
participate
The protocol of the present study was approved by
the Ethics Committee of the Institute of Animal Sciences, Chinese
Academy of Agricultural Sciences (Beijing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zuk PA, Zhu M, Mizuno H, Huang J, Futrell
JW, Katz AJ, Benhaim P, Lorenz HP and Hedrick MH: Multilineage
cells from human adipose tissue: Implications for cell-based
therapies. Tissue Eng. 7:211–228. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rangwala SM and Lazar MA: Transcriptional
control of adipogenesis. Ann Rev Nutri. 20:535–559. 2000.
View Article : Google Scholar
|
|
3
|
Deslex S, Negrel R, Vannier C, Etienne J
and Ailhaud G: Differentiation of human adipocyte precursors in a
chemically defined serum-free medium. Int J Obes. 11:19–27.
1987.PubMed/NCBI
|
|
4
|
Hauner H, Entenmann G, Wabitsch M,
Gaillard D, Ailhaud G, Negrel R and Pfeiffer EF: Promoting effect
of glucocorticoids on the differentiation of human adipocyte
precursor cells cultured in a chemically defined medium. J Clin
Invest. 84:1663–1670. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Halvorsen YD, Bond A, Sen A, Franklin DM,
Lea-Currie YR, Sujkowski D, Ellis PN, Wilkison WO and Gimble JM:
Thiazolidinediones and glucocorticoids synergistically induce
differentiation of human adipose tissue stromal cells: Biochemical,
cellular, and molecular analysis. Metabolism. 50:407–413. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Deliloglu-Gurhan SI, Vatansever HS,
Ozdal-Kurt F and Tuglu I: Characterization of osteoblasts derived
from bone marrow stromal cells in a modified cell culture system.
Acta Histochem. 108:49–57. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tashiro K, Kondo A, Kawabata K, Sakurai H,
Sakurai F, Yamanishi K, Hayakawa T and Mizuguchi H: Efficient
osteoblast differentiation from mouse bone marrow stromal cells
with polylysin-modified adenovirus vectors. Biochem Bioph Res
Commun. 379:127–132. 2009. View Article : Google Scholar
|
|
8
|
Guilak F, Lott KE, Awad HA, Cao Q, Hicok
KC, Fermor B and Gimble JM: Clonal analysis of the differentiation
potential of human adipose-derived adult stem cells. J Cell
Physiol. 206:229–237. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gimble JM, Katz AJ and Bunnell BA:
Adipose-derived stem cells for regenerative medicine. Circ Res.
100:1249–1260. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Izadpanah R, Trygg C, Patel B, Kriedt C,
Dufour J, Gimble JM and Bunnell BA: Biologic properties of
mesenchymal stem cells derived from bone marrow and adipose tissue.
J Cell Biochem. 99:1285–1297. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Naghdi M, Tiraihi T, Namin SA and
Arabkheradmand J: Transdifferentiation of bone marrow stromal cells
into cholinergic neuronal phenotype: A potential source for cell
therapy in spinal cord injury. Cytotherapy. 11:137–152. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhu Y, Liu T, Song K, Fan X, Ma X and Cu
Z: Adipose-derived stem cell: A better stem cell than BMSC. Cell
Biochem Funct. 26:664–675. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vachkova E, Bosnakovski D, Yonkova P,
Grigorova N, Ivanova Zh, Todorov P, Penchev G, Milanova A,
Simeonova G, Stanilova S and Georgiev IP: Adipogenic potential of
stem cells derived from rabbit subcutaneous and visceral adipose
tissue in vitro. In Vitro Cell Dev Biol Anim. 52:829–837. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jiang Z, Harrison DE, Parsons ME,
McClatchy S, Jacobs L and Pazdro R: Heritability of in vitro
phenotypes exhibited by murine adipose-derived stromal cells. Mamm
Genome. 27:460–468. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jung HG, Ahn EK, Lee JH, Kim YH, Leem SH,
Heo J and Kim H: Effects of harvesting sites and ages on adipose
tissue-derived stem cells in rat. Tissue Eng Regen Med. 11:137–142.
2014. View Article : Google Scholar
|
|
16
|
Madonna R, Geng YJ and De Caterina R:
Adipose tissue-derived stem cells characterization and potential
for cardiovascular repair. Arterioscl Throm Vasc Biol.
29:1723–1729. 2009. View Article : Google Scholar
|
|
17
|
Rodbell M: The metabolism of isolated fat
cells. IV. Regulation of release of protein by lipolytic hormones
and insulin. J Biol Chem. 241:3909–3917. 1966.PubMed/NCBI
|
|
18
|
Rodbell M and Jones AB: Metabolism of
isolated fat cells. 3. The similar inhibitory action of
phospholipase C (Clostridium perfringens alpha toxin) and of
insulin on lipolysis stimulated by lipolytic hormones and
theophylline. J Biol Chem. 241:140–142. 1966.PubMed/NCBI
|
|
19
|
Yoshimura H, Muneta T, Nimura A, Yokoyama
A, Koga H and Sekiya I: Comparison of rat mesenchymal stem cells
derived from bone marrow, synovium, periosteum, adipose tissue, and
muscle. Cell Tissue Res. 327:449–462. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jing W, Lin Y, Wu L, Li X, Nie X, Liu L,
Tang W, Zheng X and Tian W: Ectopic adipogenesis of preconditioned
adipose-derived stromal cells in an alginate system. Cell Tissue
Res. 330:567–572. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Unger RH, Scherer PE and Holland WL:
Dichotomous roles of leptin and adiponectin as enforcers against
lipotoxicity during feast and famine. Mol Biol Cell. 24:3011–3015.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Seo BM, Miura M, Gronthos S, Bartold PM,
Batouli S, Brahim J, Young M, Robey PG, Wang CY and Shi S:
Investigation of multipotent postnatal stem cells from human
periodontal ligament. Lancet. 364:149–155. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jang HJ, Cho KS, Park HY and Roh HJ:
Adipose tissue-derived stem cells for cell therapy of airway
allergic diseases in mouse. Acta Histochem. 113:501–507. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Conget PA and Minguell JJ: Phenotypical
and functional properties of human bone marrow mesenchymal
progenitor cells. J Cell Physiol. 181:67–73. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Haynesworth SE, Baber MA and Caplan AI:
Cytokine expression by human marrow-derived mesenchymal progenitor
cells in vitro: Effects of dexamethasone and IL-1 alpha. J Cell
Physiol. 166:585–592. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Levy JE, Jin O, Fujiwara Y, Kuo F and
Andrews NC: Transferrin receptor is necessary for development of
erythrocytes and the nervous system. Nat Genet. 21:396–399. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gong X, Hou L, Bai C, Jin D, He X, Guan W
and Ma Y: Isolation and biological characteristics of chicken
adipose-derived progenitor Cells. DNA Cell Biol. 30:453–460. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nogami M, Tsuno H, Koike C, Okabe M,
Yoshida T, Seki S, Matsui Y, Kimura T and Nikaido T: Isolation and
characterization of human amniotic mesenchymal stem cells and their
chondrogenic differentiation. Transplantation. 93:1221–1228. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nakagami H, Morishita R, Maeda K, Kikuchi
Y, Ogihara T and Kaneda Y: Adipose tissue-derived stromal cells as
a novel option for regenerative cell therapy. J Atheroscler Thromb.
13:77–81. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Uzbas F, May ID, Parisi AM, Thompson SK,
Kaya A, Perkins AD and Memili E: Molecular physiognomies and
applications of adipose-derived stem cells. Stem Cell Rev Rep.
11:298–308. 2015. View Article : Google Scholar
|