Introduction
Alzheimer's disease (AD) is a neurodegenerative
disease characterized by progressive cognitive impairment. The
disease was officially named in 1906 and the pathological changes
are mainly manifested as the loss of neurons and the formation of
senile plaques and neurofibrillary tangles. In recent years,
epidemiological and molecular pathology studies have shown that
there is a close correlation between diabetes and AD. Both diabetes
and AD are aging degenerative diseases with similar behavioral and
pathological changes (1,2). The correlation between diabetes and AD
provides a new idea for the establishment of AD animal model.
Streptozotocin (STZ), a methylnitrosourea product,
has antibacterial and antitumor properties and is capable of
causing diabetes. It is a commonly used chemical compound for the
study of the association between diabetes and AD (3). Peripheral blood vessel or
intraperitoneal injection of STZ can selectively destroy pancreatic
B cells, causing diabetes, which in turn induces symptoms of AD in
mice. Intracerebroventricular injection of STZ can destroy the
central insulin receptor phosphorylation, causing disorders of
insulin signaling pathway (4).
Consequently, it can cause brain glucose utilization and energy
metabolism disorder, leading to learning and memory disorders and
other AD symptoms (5).
Although both methods can induce the development of
AD in mice (6), no comparison has
been made yet between the behavior and the changes of synaptic PSD
in the cortical and hippocampal neurons between intraperitoneal and
intracerebroventricular injection models. Therefore, in this
experiment, AD model was established by intraperitoneal and
intracerebroventricular injection of STZ, respectively. Morris
water maze (MWM) and fear-conditioning tests were used to determine
the behavioral changes in the two models (7,8). PSD95
and shank3 in cerebral cortex and hippocampus were also measured in
order to identify the differences between the two models and
discuss the potential underlying mechanisms that cause the
differences.
Materials and methods
Animals
ICR (n=110), db/db (n=10) and app/ps1 male mice
(n=10) (weighing 18–20 g, aged 8–12 weeks) were purchased from the
Research Institute of Model Animals at Nanjing University (Nanjing,
China). Mice were housed in microisolator cages under specific
pathogen-free conditions, fed with autoclaved food. Mice were
handled under a protocol approved by the Institutional Animal Care
and Use Committee of Nanjing University of Chinese Medicine
(Nanjing, China).
Experimental protocol
The ICR newborn male mice were randomly selected for
intraperitoneal injection of STZ (Beyotime Institute of
Biotechnology, Shanghai, China). The mice were given a single
subperitoneal injection of STZ 300 µl for routine feeding with the
mother mice. The control group was injected with the same amount of
PBS buffer. The changes of body weight and blood glucose were
detected at the 8th week. After the model was established
successfully, the behavior test was carried out and the mice were
sacrificed to detect the PSD protein in the cortex. In the
intracerebroventricular injection of STZ model group, the adult ICR
male mice were randomly selected at 7 weeks of age, and the lateral
ventricle was injected with STZ 2 µl once at body surface location.
The control group was injected with the same amount of PBS buffer.
After 1 week, the changes of body weight and blood glucose were
measured. After the model was established successfully, the
behavioral test was carried out, and then the mice were sacrificed
to detect the PSD protein in the cortex. The db/db male mice and
the app/ps1 male mice had the behavioral test at the age of 8 weeks
with routine feeding.
MWM test
Briefly, the mice were tested in a plastic water
pool (120 cm in diameter) containing water at 22–24°C with
distinctive spatial cues on all four walls. A platform (8 cm in
diameter) was submerged 1 cm under the water surface. The water
pool was divided into four quadrants: northeast (NE); southeast
(SE); northwest (NW); wouthwest (SW). The platform was placed in
the SW quadrant as a target quadrant (TQ). The mice were dropped
from the edge of the pool from four different locations (NW, SW,
SE, NE) on the training day in a counter balanced order for 4 days.
For each trial, the animal was allowed to swim until reaching the
platform, where it would remain for 20 sec. If a mouse did not find
the platform within 60 sec, we gently directed the animal to the
platform, where it would remain for 20 sec. Finally, the testing
trial from random quadrant was video tracked using camera to record
the escape latency (sec) to the platform.
Fear-conditioning test
Mice were placed into the chambers individually.
After 3 min, mice were subjected to electric foot-shocks (0.75 mA,
2 sec) 7 times with tone (2.5 kHz, 85 dB, 30 sec). The mice were
then returned to their home cages. The next day, the mice were
placed in the same chamber for 30 min without foot-shock. At the
3rd day, the mice were placed in the same chamber for 30 min with
tone but no foot-shock. The behavior of the mice was recorded by a
digital video camera. Continuous immobility for >1 sec was
defined as freezing behavior. The duration of freezing was
calculated as the ratio of freezing time to total recorded time
(%).
Western blot analysis
Western blot analysis of lysates of mouse cerebral
cortex homogenates was performed as previously described (9). Briefly, fresh tissues were lysed on ice
in the buffer containing 1 mM PMSF, 0.2 U/ml aprotinin, and 1 mM
sodium orthovanadate. Protein concentrations of the samples were
determined by use of the bicinchoninic acid protein assay kit.
Total tissue lysate proteins (30 µg, 5 µg protein loaded per lane)
were loaded for 14% SDS-PAGE, and the separated proteins were
transferred onto nitrocellulose membranes by electroblotting. The
membranes were incubated with primary antibodies overnight, washed,
and then incubated with goat anti-rabbit or anti-mouse IgG
conjugated to horseradish peroxidase (1:3,000-5,000) for 60 min at
20°C. Protein bands were detected with the ECL SuperSignal reagent
(EMD Millipore, Billerica, MA, USA). Relative band densities of the
various proteins were measured from scanned films with ImageJ
software.
Drugs, reagents, and antibodies
Mouse monoclonal Shank3 antibody (dilution, 1:500;
cat. no. ab93607); mouse monoclonal PSD95 antibody (dilution,
1:500; cat. no. ab2723); rabbit polyclonal β-actin antibody
(dilution, 1:1,000; cat. no. ab8227); secondary goat anti-rabbit
(HRP) IgG antibody (dilution, 1:2,000; cat. no. ab6721) and
secondary rabbit anti-mouse (HRP) IgG antibody (dilution, 1:2,000;
cat. no. ab6728) were all purchased from Abcam (Cambridge, MA,
USA).
Statistical analysis
One-way ANOVA and Student-Newman- Keuls test for
post-hoc comparisons were used to determine differences between the
control and the experimental groups. Student's t-test was performed
for paired samples. Parameter changes between different groups over
time were evaluated by a two-way ANOVA with repeated measurements.
Data are expressed as means ± SE, and the differences between
groups were considered significant if P≤0.05.
Results
Body weight and blood sugar
alteration
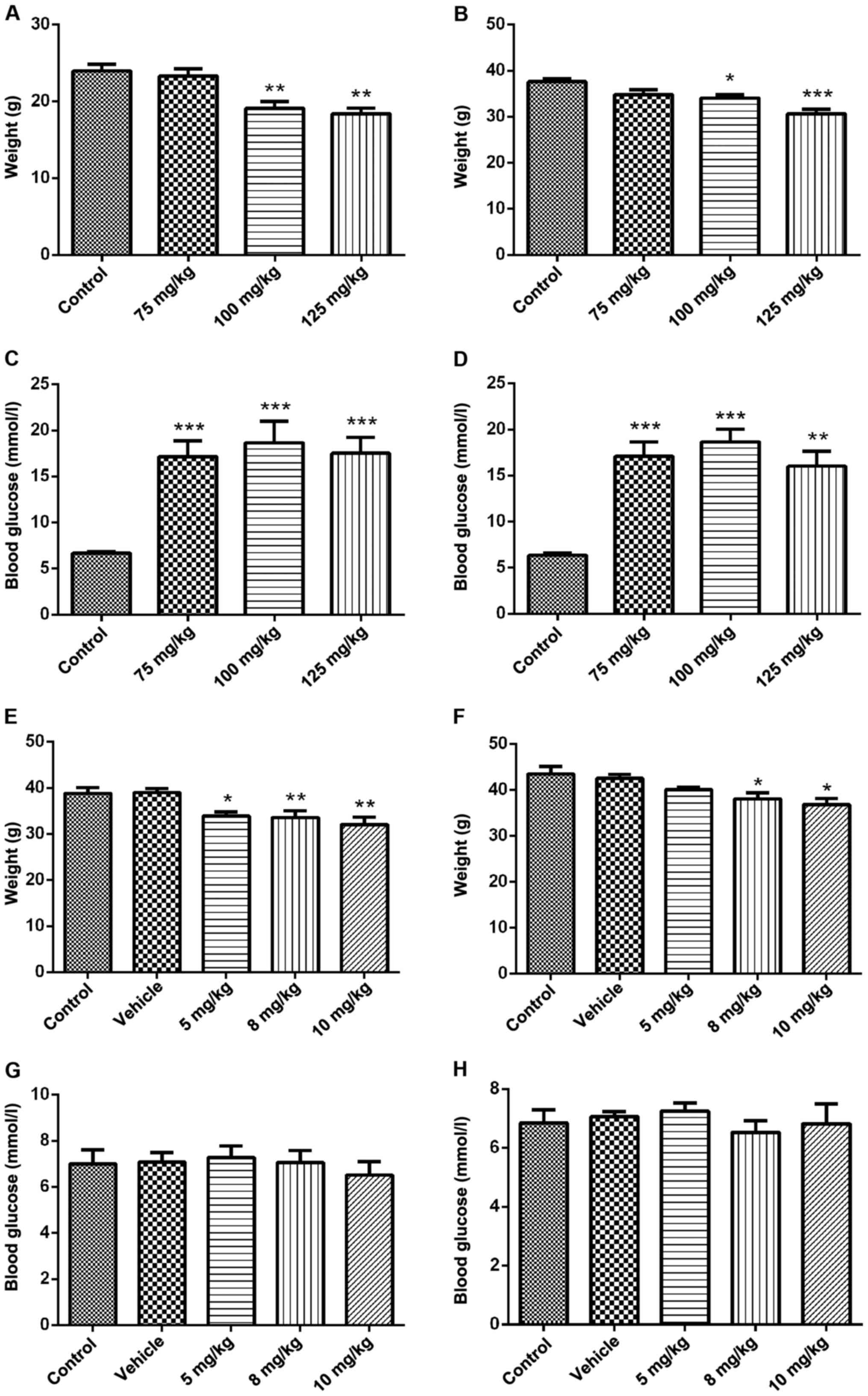
Neonatal male mice were injected with STZ (100
mg/kg) intraperitoneally and the body weight was reduced at 8 and
12 weeks of age compared with the control group (Fig. 1A and B). Blood glucose was increased
compared with the control group and showed significant signs of
diabetes (Fig. 1C and D). However,
after the intracerebroventricular injection of STZ (5 mg/kg) in
adult mice at the age of 7 weeks, body weight was decreased at the
age of 8 and 12 weeks (Fig. 1E and
F), while the blood glucose did not change significantly
(Fig. 1G and H).
Behavior alteration
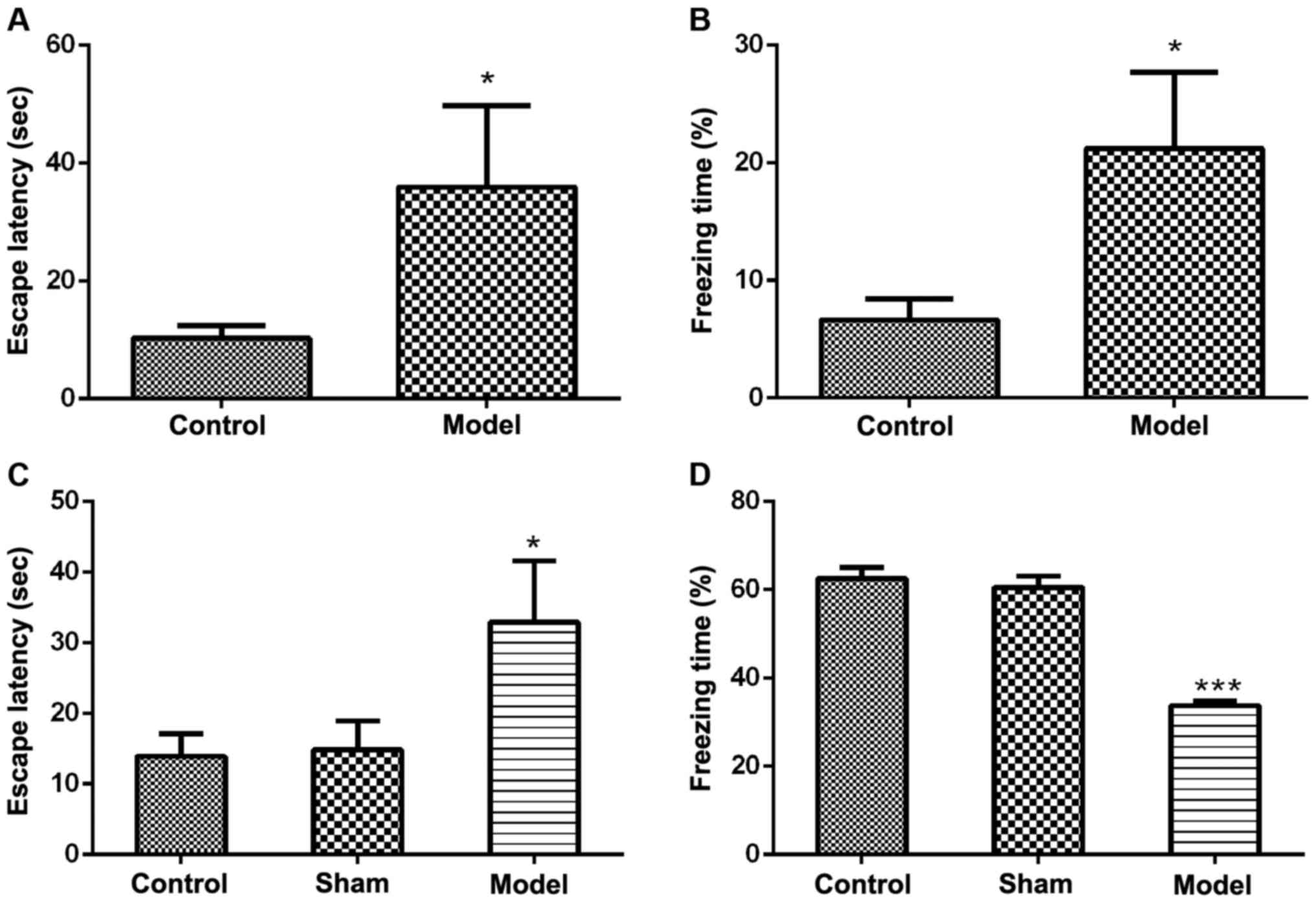
MWM test showed that the escape latency (the time it
takes to find the platform) in both intraperitoneal and
intracerebroventricular groups was extended compared with the
control group (Fig. 2A and C).
Fear-conditioning test showed that the freezing time in the
intraperitoneal group was increased (Fig. 2B), while in the
intracerebroventricular group, the freezing time was reduced
(Fig. 2D).
Alteration of PSD95 and shank3
proteins
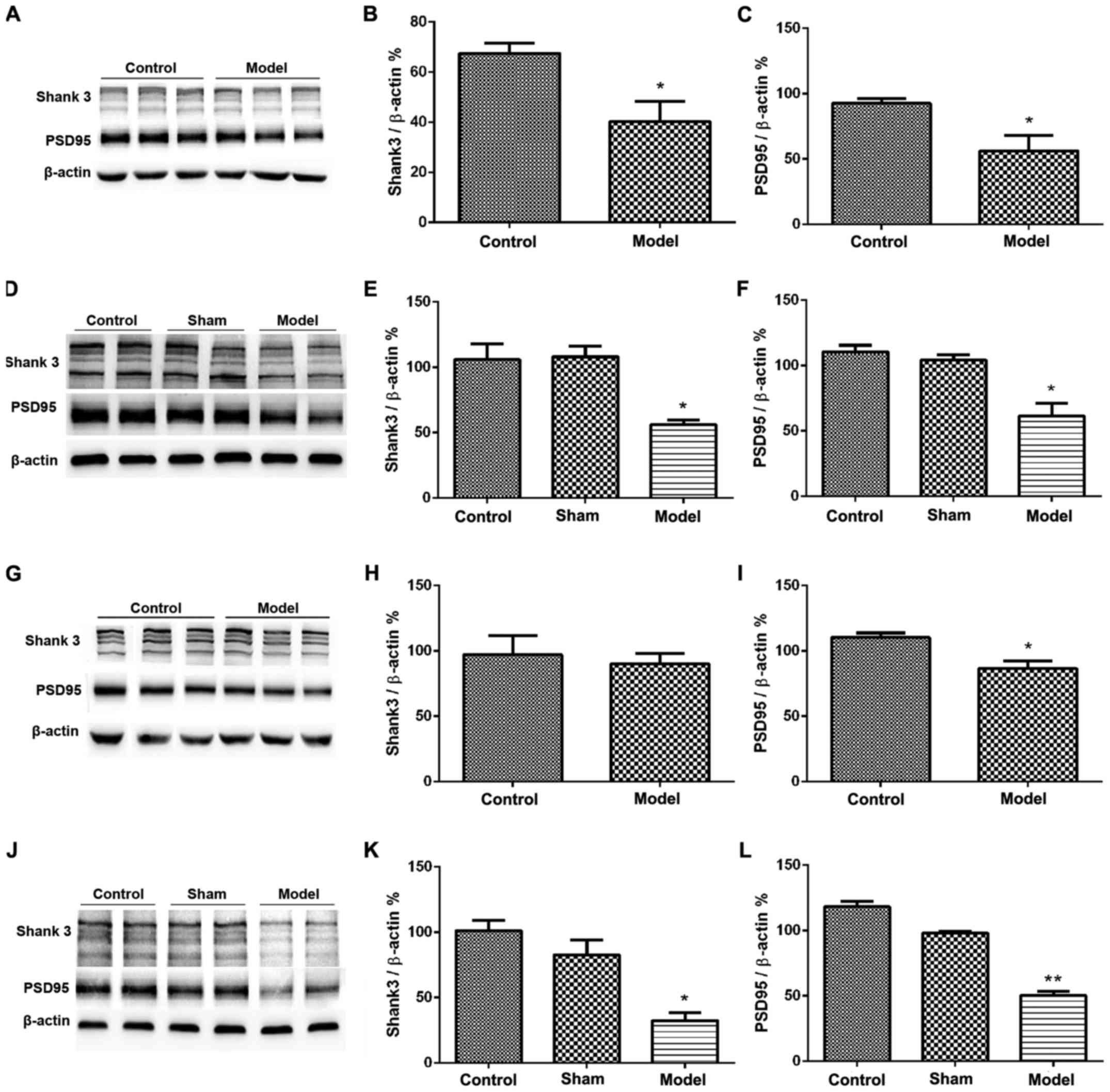
PSD95 and shank3 proteins in the cerebral cortex in
both intraperitoneal and intracerebroventricular groups were
decreased compared with the control group (Fig. 3A-F). PSD95 in the hippocampus was
reduced in both intraperitoneal and intracerebroventricular groups
compared with the control group (Fig.
3G, I, J and L). Shank3 in hippocampus in the intraperitoneal
group was not significantly different from that in the control
group (Fig. 3G and H). Shank3 in the
hippocampus in the intracerebroventricular group was significantly
reduced compared with the control group (Fig. 3J and K).
Fear-conditioning test in transgenic
mice
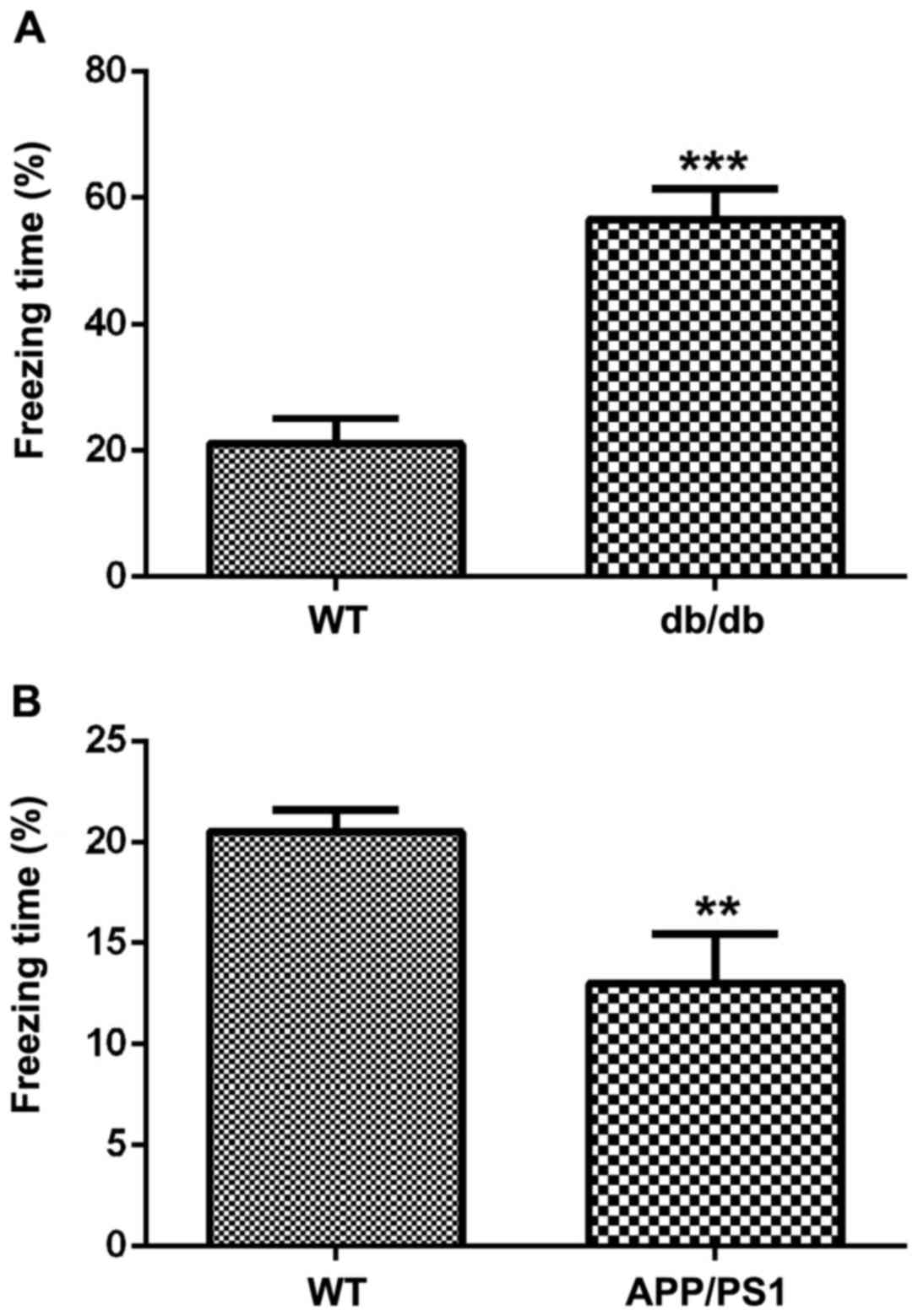
Fear-conditioning test showed that the freezing time
in db/db mice (transgenic type 2 diabetic mice) was longer than
mice in the control group (Fig. 4A).
The freezing time in the app/ps1 mice (transgenic AD mice) was
reduced compared with mice in the control group (Fig. 4B).
Discussion
The main clinical manifestations of AD patients are
cognitive decline and other mental and emotional abnormalities
(1). They were classified into
forgetting, mobility disorders, aggression and feeling upset,
depression, anxiety and fear. Animal experiments usually showed a
decrease in learning and memory ability, including spatial memory
and emotional memory loss (1,2).
Streptozotocin (STZ) can selectively destroy cells
that have the function of synthesizing and secreting insulin. In
the peripheral system (such as intraperitoneal injection), STZ can
directly destroy pancreatic B cells and cause insulin resistance,
thus inducing diabetes (3). STZ
inducing AD-like pathological changes in rats or mice has also been
considered to be a good AD experimental animal model. In the
central nervous system, STZ can reduce the expression of cortical
and hippocampal insulin receptor, thus undermining the insulin
synthesis in the central nervous system. Consequently, the
sensitivity of the central nervous system to peripheral insulin is
reduced, leading to central nervous system dysfunction, and
triggering AD symptoms (4). In
addition, intraventricular injection with different STZ doses can
induce emotional behavior changes in mice. Therefore, different STZ
injection route can lead to different changes in blood glucose and
behavior, including changes in spatial memory and emotions.
Conditional fear is the appearance of a neutral
stimulus (such as light or sound) followed by a dangerous
unconditional stimulus (usually a shock). After multiple times of
trainings, neutral stimuli can be transformed into conditional
stimuli. Formation of conditional fears triggers some behavioral
and physiological reactions. These reactions are typically
defensive responses (e.g., freezing or fearful jumping) that occur
when an animal is exposed to danger or fear arising (10,11).
Different from spatial learning and memory, the essence of the
formation of fear emotion memory is that some immediate stimuli
trigger the activation of hippocampus (long-term potentiation) and
amygdala, and the integration of all the three factors. This
defensive response time reflects the speed of fear memory
subsiding. Pathological mechanisms of AD are usually due to the
reduction of this defensive response time, indicating a reduced
emotional memory (11,12). However, some studies have shown that
the duration of fear and freezing was increased in diabetic mice
caused by intravenous injection of STZ. It is presumed that the
amygdala-hippocampus pathway is damaged and the sensitivity to fear
is increased, manifesting more pronounced fear (12).
Postsynaptic density (PSD) is attached to the
postsynaptic membrane and under the electron microscope this region
has high electron density. PSD is an important structure that
affects neuronal synaptic plasticity (13). As a scaffolding network of the
postsynaptic membrane, PSD has glutamate receptors, scaffold
proteins, and many signaling molecules. Shank3 is a scaffolding
protein in PSD that promotes the formation of excitatory synapses
and the development of dendritic spines (14,15).
PSD95 is a PSD scaffold protein that is associated with most
neurotransmitters and other scaffold proteins. Proteomic analysis
revealed significant changes in the PSD95 and shank3 proteins in
the postsynaptic membrane of the cerebral cortex in patients with
AD, suggesting that these proteins may directly affect synaptic
function at the molecular level (16,17).
In this study, we showed that i) if the newborn
mouse is given STZ intraperitoneally, it is not necessary to
combine a high-fat and high-sugar diet to establish a diabetic
model and induce AD symptoms with impaired learning and memory; ⅱ)
injection of STZ into the lateral ventricle of adult mice can
directly induce the learning and memory impairment of AD model
without causing changes in blood glucose; ⅲ) both methods can cause
spatial memory impairment in mice, but the conditional fear is
different between the two methods: this difference is consistent
with the results caused by simple transgenic animals; and ⅳ) the
difference in conditional fear between the two methods may be due
to different STZ injection routes, leading to different levels of
blood glucose and shank3 in the PSD structure of mouse hippocampal
synaptosomes.
In summary, this study compared two STZ-induced AD
models, and the results showed that intraperitoneal injection of
STZ in newborn mice led to high blood glucose, impaired spatial
memory, and increased emotional memory when they become adults.
Intracerebroventricular injection of STZ in adult mice directly
caused the loss of spatial and emotional memory without changes in
blood glucose, which may be closely related to the changes of the
shank3 protein in the hippocampal synaptosomes. The mechanism needs
to be further studied to better illustrate the similarities and
differences between the two models and provide sufficient
scientific evidence on diabetic and AD studies.
Acknowledgements
We would like to thank Haitong Zhang for her
assistance with the experiments.
Funding
This study was financially supported by the Priority
Academic Program Development (PAPD) fund of the Jiangsu Higher
Education Institution (Dr Y. Gong).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ conceived the study and drafted the manuscript.
RD and SW performed the animal behavior experiments and the data
analysis; ZR, LX and YW were responsible for the brain tissue
protein assay; XZ, JZ, YD and YG detected the body weight and blood
sugar of animals; YW and YG revised the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Institutional Animal
Care and Use Committee of Nanjing University of Chinese Medicine
(Nanjing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sorial ME and El Sayed NSED: Protective
effect of valproic acid in streptozotocin-induced sporadic
Alzheimer's disease mouse model: Possible involvement of the
cholinergic system. Naunyn Schmiedebergs Arch Pharmacol.
390:581–593. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li D, Huang Y, Cheng B, Su J, Zhou WX and
Zhang YX: Streptozotocin induces mild cognitive impairment at
appropriate doses in mice as determined by long-term potentiation
and the Morris water maze. J Alzheimers Dis. 54:89–98. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Singh JC, Kakalij RM, Kshirsagar RP, Kumar
BH, Komakula SS and Diwan PV: Cognitive effects of vanillic acid
against streptozotocin-induced neurodegeneration in mice. Pharm
Biol. 53:630–636. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Javed H, Vaibhav K, Ahmed ME, Khan A,
Tabassum R and Islam F, Safhi MM and Islam F: Effect of hesperidin
on neurobehavioral, neuroinflammation, oxidative stress and lipid
alteration in intracerebroventricular streptozotocin induced
cognitive impairment in mice. J Neurol Sci. 348:51–59. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pandey SP, Singh HK and Prasad S:
Alterations in hippocampal oxidative stress, expression of AMPA
Receptor GluR2 subunit and associated spatial memory loss by Bacopa
monnieri Extract (CDRI-08) in streptozotocin-induced diabetes
mellitus type 2 mice. PLoS One. 10:e01318622015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kumar A and Singh N: Calcineurin
inhibitors improve memory loss and neuropathological changes in
mouse model of dementia. Pharmacol Biochem Behav. 153:147–159.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sommer C, Roth SU and Kiessling M:
Kainate-induced epilepsy alters protein expression of AMPA receptor
subunits GluR1, GluR2 and AMPA receptor binding protein in the rat
hippocampus. Acta Neuropathol. 101:460–468. 2001.PubMed/NCBI
|
|
8
|
Stragier E, Martin V, Davenas E, Poilbout
C, Mongeau R, Corradetti R and Lanfumey L: Brain plasticity and
cognitive functions after ethanol consumption in C57BL/6J mice.
Transl Psychiatry. 5:e6962015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang Y, Liu G, Dull RO, Schwartz DE and
Hu G: Autophagy in pulmonary macrophages mediates lung inflammatory
injury via NLRP3 inflammasome activation during mechanical
ventilation. Am J Physiol Lung Cell Mol Physiol. 307:L173–L185.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Poitout V and Robertson RP: Minireview:
Secondary beta-cell failure in type 2 diabetes - a convergence of
glucotoxicity and lipotoxicity. Endocrinology. 143:339–342. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ikeda H, Ikegami M, Kai M and Kamei J:
Cannabinoid functions in the amygdala contribute to conditioned
fear memory in streptozotocin-induced diabetic mice: Interaction
with glutamatergic functions. Exp Neurol. 269:233–241. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Portha B, Giroix MH, Serradas P, Morin L,
Tormo MA and Bailbe D: Cellular basis for glucose refractoriness of
pancreatic B-cells in non insulin dependent diabetes. Frontiers of
Insulin Secretion and Pancreatic B Cell Research. Flatt PR and
Lenzen S: Smith-Gordon and Co., Ltd.; Huntingdon, Cambridgeshire:
pp. 461–472. 1994
|
|
13
|
Durand CM, Betancur C, Boeckers TM,
Bockmann J, Chaste P, Fauchereau F, Nygren G, Rastam M, Gillberg
IC, Anckarsäter H, et al: Mutations in the gene encoding the
synaptic scaffolding protein SHANK3 are associated with autism
spectrum disorders. Nat Genet. 39:25–27. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gong Y, Chang L, Viola KL, Lacor PN,
Lambert MP, Finch CE, Krafft GA and Klein WL: Alzheimer's
disease-affected brain: Presence of oligomeric A beta ligands
(ADDLs) suggests a molecular basis for reversible memory loss. Proc
Natl Acad Sci USA. 100:10417–10422. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gong Y and Lippa CF: Review: Disruption of
the postsynaptic density in Alzheimer's disease and other
neurodegenerative dementias. Am J Alzheimers Dis Other Demen.
25:547–555. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gong Y, Lippa CF, Zhu J, Lin Q and Rosso
AL: Disruption of glutamate receptors at Shank-postsynaptic
platform in Alzheimer's disease. Brain Res. 1292:191–198. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu L, Ren Z, Chow FE, Tsai R, Liu T,
Rizzolio F, Boffo S, Xu Y, Huang S, Lippa CF, et al: Pathological
role of peptidyl-prolyl isomerase Pin1 in the disruption of
synaptic plasticity in Alzheimer's disease. Neural Plast.
2017:32707252017. View Article : Google Scholar : PubMed/NCBI
|