Introduction
Chronic kidney disease (CKD) is the final stage of a
number of renal diseases. In recent years, it has been reported
that tubule interstitial fibrosis (TIF) is associated with the
occurrence and development of CKD (1). Tubular epithelial cells (TECs) serve
important roles in maintaining renal function and the progression
of TIF (2). Kidney injury may lead
to the phenotypic transformation of TECs (3), which is similar to the process of
epithelial mesenchymal transition (EMT). Under such conditions,
TECs acquire some properties of fibroblasts, producing excessive
extracellular matrix (ECM) and related proteins, including collagen
and fibronectin, leading to fibrosis of the kidney cells and
eventual nephropathy (4,5).
The pathogenesis of TIF remains unclear. It has been
reported that transforming growth factor (TGF)-β serves a central
role in the onset and progression of kidney fibrosis (6–8).
MicroRNAs (miRNAs or miRs) are a family of small non-coding RNAs
18–25 nucleotides in length (9).
miRNAs are able to bind to the 3′-untranslated regions (3′-UTR) of
their paired mRNAs to suppress the expression of target genes
(10). A number of miRNAs have been
reported to participate in the pathogenesis of many diseases,
including cancers, autoimmune diseases, cardiovascular diseases and
acute and chronic renal disease (10–12).
However, the underlying mechanisms of miRNAs in renal diseases
require further investigation.
Quercetin is a widely available flavonoid that can
be isolated from many plants, vegetables and fruits (13). Increasing evidence has indicated that
quercetin exerts various biological activities, such as
antioxidant, anti-tumor, anti-angiogenesis and anti-inflammatory
functions (14,15). Quercetin has also been reported to
have a therapeutic effect on renal function and tissue damage
(16,17). To the best of our knowledge, the
effect of quercetin on kidney fibrosis and whether miRNAs are
serving a role in quercetin-induced renal protective effects has
not been reported.
A previous study reported that the TGF-β1 signaling
pathway is vital for the induction of renal fibrosis (18). In the present study, in order to
simulate renal fibrosis in vitro, the human tubular
epithelial cell line HK-2 was treated with TGF-β according to
previous reports (19,20). The role of miR-21 in mediating the
renal protective effect of quercetin was assessed using
TGF-β-induced fibrosis in HK-2 cells to provide a theoretical basis
for the treatment of nephropathy using quercetin.
Materials and methods
Cell culture and treatments
The human tubular epithelial cell line HK-2 was
purchased from the American Type Culture Collection (ATCC,
Manassas, VA, USA) and cultured according to the manufacturer's
protocols. Cells were cultured in Dulbecco's Modification of
Eagle's Medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml penicillin and
streptomycin in a humidified incubator at 37°C with 5%
CO2. Fibrosis was induced by treatment with 100 ng/ml
TGF-β for 72 h. The quercetin groups were treated with medium
containing 7.5, 15 or 30 mg/ml quercetin (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany).
Cell proliferation detection
Cell Counting Kit-8 (CCK-8; Dojindo, Kumamoto,
Japan) was used according to the manufacturer's protocol to assess
cell proliferation. Briefly, HK-2 cells were treated with 7.5, 15
or 30 mg/ml quercetin for 24, 48 and 72 h. Cells were subsequently
incubated with 10 µl/well CCK8 at 37°C for 4 h. Finally, the
optical density (OD) at 450 nm was measured using a multi-well
spectrophotometer (Thermo Fisher Scientific, Inc.).
Cell transfection
miR-21 inhibitor and miR-21 mimic oligonucleotides
(3′-AGUUGUAGUCAGACUAUUCGAU-5′) were synthesized by Shanghai
GenePharma Co., Ltd. (Shanghai, China). Cells were transfected with
miR-21 inhibitor (100 nM) or miR-21 mimics (50 nM) using
Lipofectamine® RNAi Max (Thermo Fisher Scientific,
Inc.). At 48 h post transfection, cells were trypsinized and
harvested (800 × g, 5 min, 4°C) for future analysis.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from cells using TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc.). RT-qPCR was performed
with an SYBR ExScript RT-PCR kit (Takara Bio, Inc., Dalian, China)
on an ABI 7500 Real-Time PCR System machine (Applied Biosystems;
Thermo Fisher Scientific, Inc.). Reverse transcription reaction was
performed for 1 h at 37°C followed by 5 min at 95°C. The
thermocycling conditions for the qPCR step were as follows: 95°C
for 30 sec followed by 40 cycles of 95°C for 5 sec and 60°C for 30
sec. The primers were synthesized by Sangon Biotech Co., Ltd.
(Shanghai, China) with sequences as follows: Collagen I forward,
5′-TGGCCAAGAAGACATCCCTGAAGT-3′ and reverse,
5′-ACATCAGGTTTCCACGTCTCACCA-3′; fibronectin forward,
5′-CCATCGCAAACCGCTGCCAT-3′ and reverse,
5′-AACACTTCTCAGCTATGGGCTT-3′; GAPDH forward,
5′-AAGAAGGTGGTGAAGCAGGC-3′ and reverse, 5′-TCCACCACCCTGTTGCTGTA-3′;
α-smooth muscle actin (α-SMA) forward,
5′-ACTGAGCGTGGCTATTCCTCCGTT-3′ and reverse,
5′-GCAGTGGCCATCTCATTTTCA-3′; epithelial E-cadherin forward,
5′-TACACTGCCCAGGAGCCAGA-3′ and reverse; 5′-TGGCACCAGTGTCCGGATTA-3′;
phosphatase and tensin homolog (PTEN) forward,
5′-CAAGATGATGTTTGAAACTATTCCAATG-3′ and reverse,
5′-CCTTTAGCTGGCAGACCACAA-3′; TIMP metallopeptidase inhibitor 3
(TIMP3) forward, 5′-CTGCAAGGGCTGGGCATC-3′ and reverse,
5′-TCCATGGCCCGGTTGGCAGTGTGGAG-3′; U6 forward,
5′-ATAAGGATCCGGTCTCGCTATGAGGGCCTATTTCCCATG-3′ and reverse,
5′-ATAATGTACAGGTCTCCCATGTAACTTGCTATTTCTAGCTC-3′. The relative
expression of collagen I, fibronectin, SMA, (E)-cadherin, PTEN and
TIMP3 in each sample was normalized to the expression of GAPDH
using the 2−ΔΔCq method (21). miR-21 expression was examined using a
Hairpin-it™ miRNAs qPCR Quantitation kit (Shanghai
GenePharma Co., Ltd.) and U6 was used for normalization. Primers,
forward 5′-GCCGCTAGCTTATCAGACTGATGT-3′ and reverse
5′-GTGCAGGGTCCGAGGT-3′, were supplied by Sangon Biotech Co.,
Ltd.
Western blotting
Cells were lysed using radioimmunoprecipitation
assay buffer (Beyotime Institute of Biotechnology, Haimen, China).
Bradford assays were used to determine the concentration of total
extracted protein. Total proteins (40 µg) were loaded and separated
on 10% SDS-PAGE gels. Proteins were transferred onto polyvinylidene
fluoride membranes, blocked in 5% non-fat milk for 2 h at room
temperature and incubated with the following primary antibodies:
Anti-collagen I (1:1,000; cat. no. 84336; Cell Signaling
Technology, Inc., Danvers, MA, USA), anti-fibronectin (1:200; cat.
no. ab2413; Abcam, Cambridge, MA, USA), anti-α-SMA (1:1,000; cat.
no. 19245; Cell Signaling Technology, Inc.), anti-E-cadherin
(1:1,000; cat. no. 3195; Cell Signaling Technology, Inc.),
anti-PTEN (1:1,000; cat. no. 9188; Cell Signaling Technology,
Inc.), anti-TIMP3 (1:1,000; cat. no. 5673; Cell Signaling
Technology, Inc.) and anti-β-actin (1:1,000; cat. no. 4970; Cell
Signaling Technology, Inc.) overnight at 4°C. The membranes were
subsequently washed three times with TBST buffer and incubated with
the secondary horseradish peroxidase-linked antibody goat
anti-rabbit IgG (1:1,000; cat. no. A0208; Beyotime Institute of
Biotechnology) for 1 h at room temperature, following which they
were washed three times with TBST buffer, incubated with BeyoECL
Plus (Beyotime Institute of Biotechnology) and visualized using
ChemiDoc™ XRS+ System (Bio-Rad Laboratories, Inc., Hercules, CA,
USA). β-actin was used as the internal control.
Immunocytochemistry (IHC)
For IHC, cells were fixed in 4% paraformaldehyde
overnight at room temperature and permeabilized with 0.1% Triton
X-100 in 0.5% bovine serum albumin (Sigma-Aldrich; Merck KGaA) for
30 min at room temperature. Cells were incubated with the rabbit
anti-human PTEN (1:125; cat. no. 9188; Cell Signaling Technology,
Inc.) primary antibody overnight at 4°C. Following washing for
three times with PBS, cells were incubated with Alexa
Fluor® 488-conjugated rabbit mAb (1:800; cat. no. 9854;
Cell Signaling Technology, Inc.) secondary antibody for 30 min at
37°C. Cells were visualized using a fluorescent microscope
(magnification, ×400).
Statistical analysis
Statistical analysis was performed using SPSS 19.0
(IBM Corp., Armonk, NY, USA). Data are presented as the mean ±
standard deviation and a two-independent sample t-test was
performed used to make comparisons between two groups. One-way
analysis of variance followed by Dunnett's test was used for
multiple group comparisons. P<0.05 was considered to indicate a
significant difference.
Results
TGF-β treatment induces fibrosis in
HK-2 cells
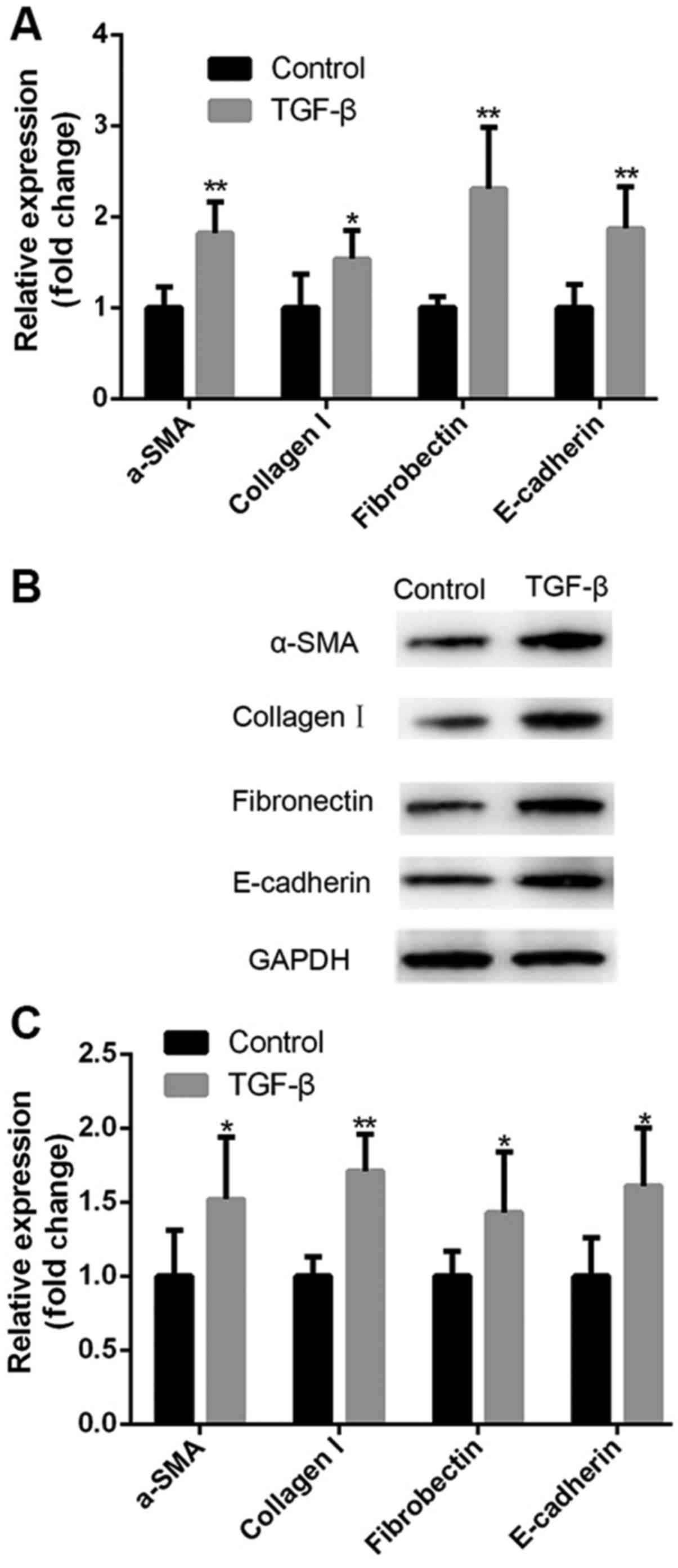
HK-2 cells were treated with TGF-β and the
expression of fibrotic markers was evaluated using RT-qPCR and
western blotting. Compared with untreated cells, TGF-β induced a
significant increase in the expression of fibrotic markers collagen
I, fibronectin, α-SMA and E-cadherin at the mRNA (Fig. 1A) and protein levels (Fig. 1B and C), suggesting that TGF-β
treatment induces a fibrosis-like state in HK-2 cells in
vitro.
Quercetin alleviates TGF-β-induced
fibrosis in HK-2 cells
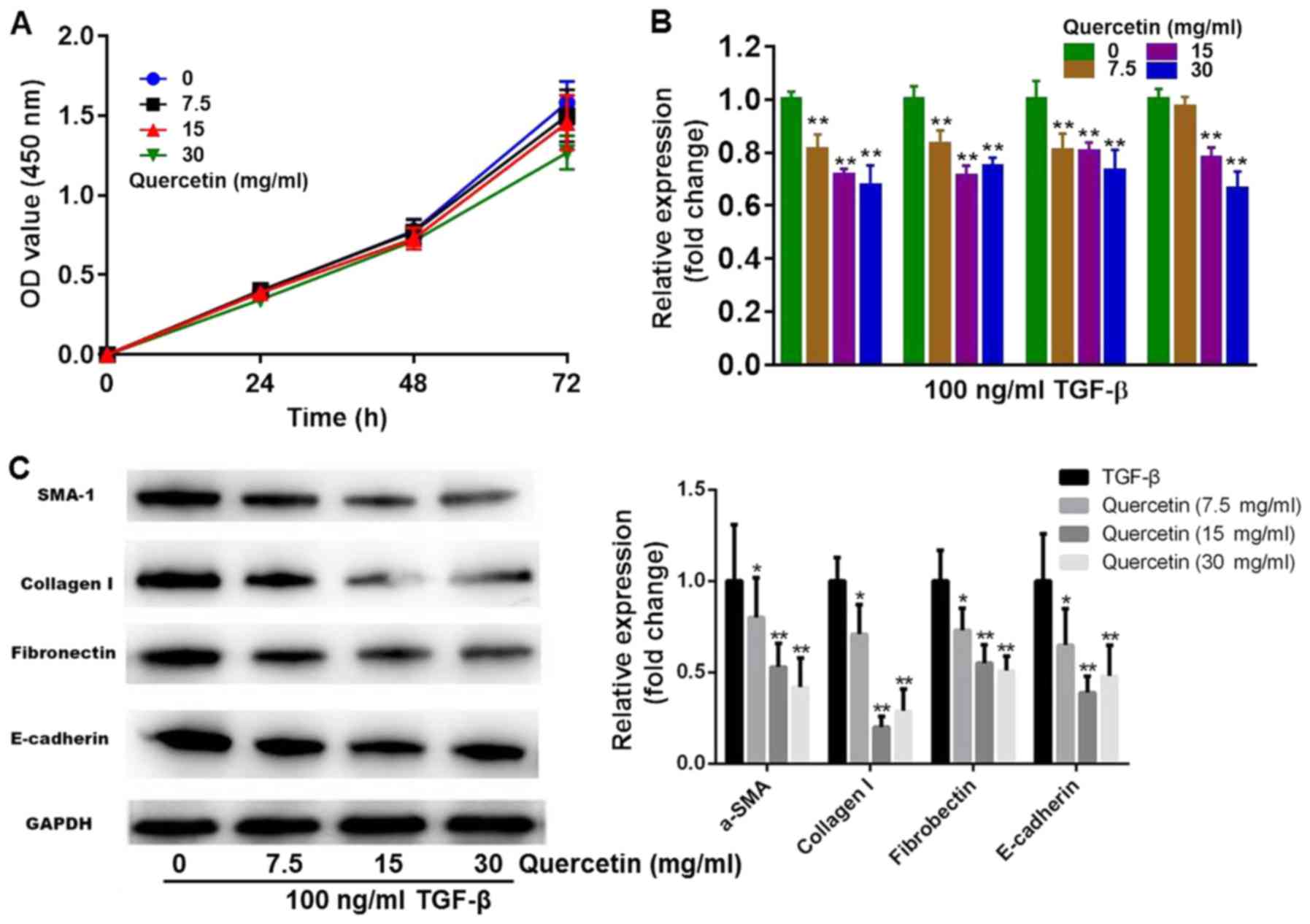
To assess whether quercetin is able to alleviate
TGF-β-induced fibrosis in HK-2 cells, TGF-β-stimulated HK-2 cells
were treated with different concentrations of quercetin and the
expression of fibrotic markers was evaluated using RT-qPCR and
western blotting. CCK-8 results revealed that treatment with 7.5
and 15 mg/ml quercetin had a limited effect on cell growth, while
30 mg/ml quercetin had a slight cytotoxic effect (Fig. 2A). Compared with the TGF-β group,
TGF-β + 7.5 mg/ml quercetin induced a significant decrease in the
expression of collagen I, fibronectin and α-SMA. In the TGF-β + 15
mg/ml and TGF-β + 30 mg/ml quercetin groups, a significant decrease
in collagen I, fibronectin, α-SMA and E-cadherin mRNA expression
was observed (Fig. 2B). Furthermore,
quercetin induced a significant decrease in the protein expression
of fibrotic markers (Fig. 2C). The
effect of quercetin on α-SMA and fibronectin expression was
dose-dependent (Fig. 2C). Based on
these results, 15 mg/ml quercetin had been selected for further
investigations.
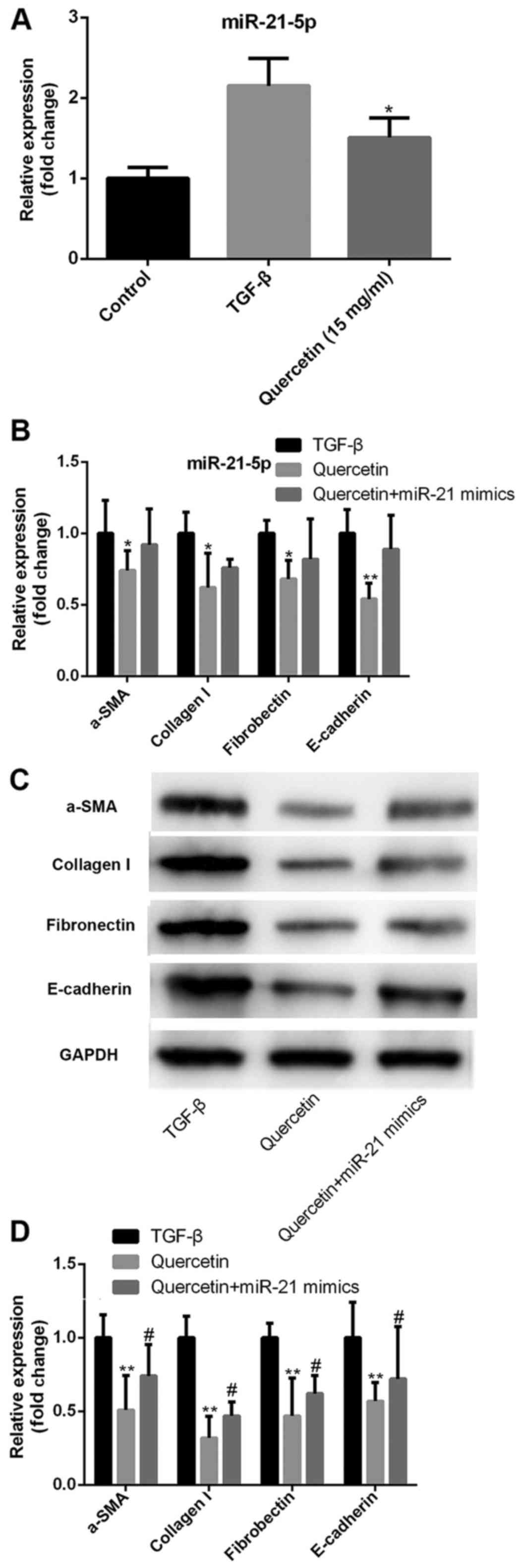
Quercetin alleviates TGF-β induced
fibrosis via regulating miR-21 expression
The role of miR-21 in quercetin-induced
anti-fibrosis was investigated. TGF-β treatment induced a
significant increase in miR-21 expression, while quercetin
treatment decreased the TGF-β-induced miR-21 upregulation in HK-2
cells (Fig. 3A). Transfection with
miR-21 mimics inhibited the quercetin-induced anti-fibrotic
effects, increasing the expression of collagen I, fibronectin,
α-SMA and E-cadherin at the mRNA (Fig.
3B) and protein levels (Fig. 3C and
D).
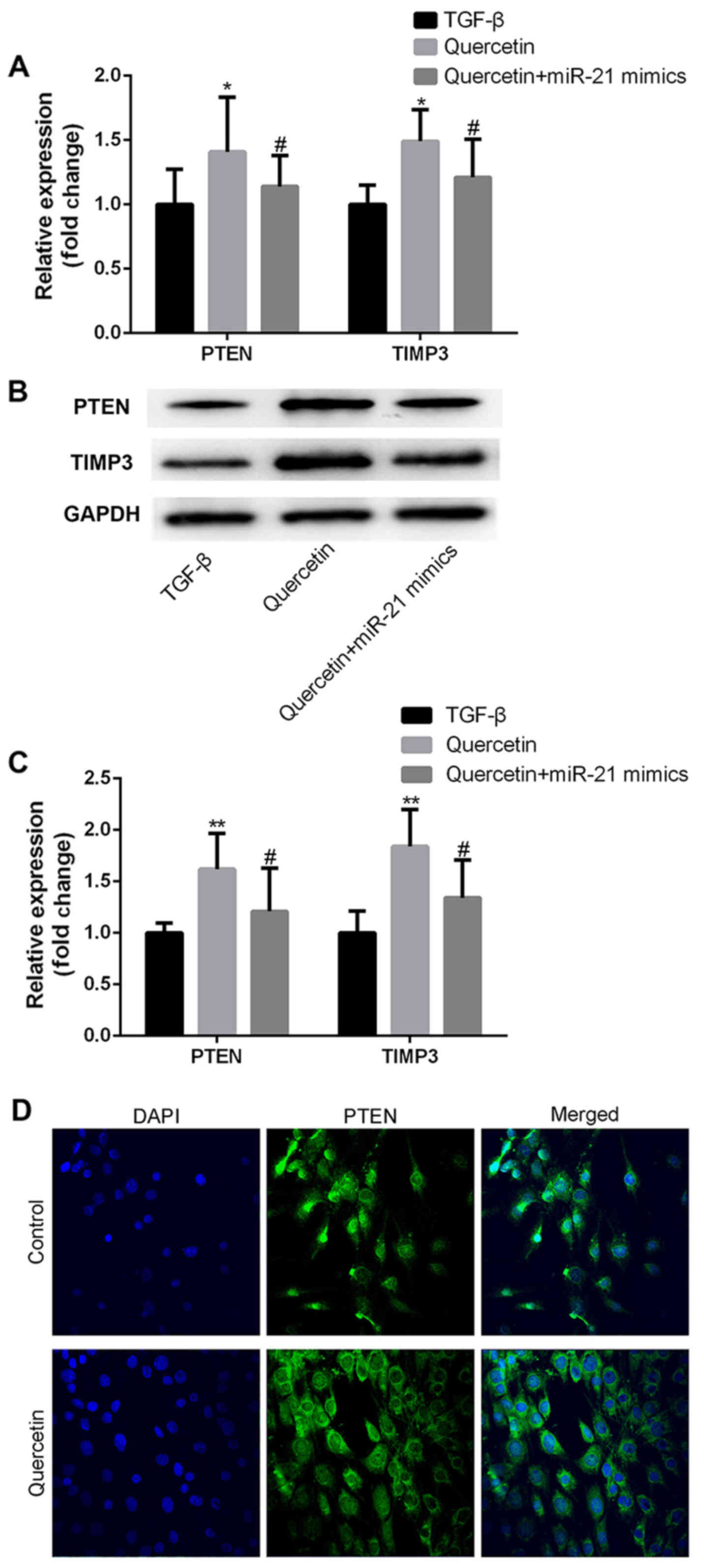
Quercetin increases the expression of
miR-21 target genes in TGF-β treated HK-2 cells
PTEN and TIMP3 have previously been confirmed as
direct targets of miR-21 (22,23). To
further explore the mechanism of miR-21 in quercetin-induced
anti-fibrotic effects, the expression of PTEN and TIMP3 in
untreated fibrotic and quercetin-treated fibrotic HK-2 cells was
assessed. Quercetin treatment induced a significant increase in the
expression of PTEN and TIMP3 at the mRNA (Fig. 4A) and protein levels (Fig. 4B-D). Furthermore, transfection with
miR-21 mimics partially inhibited the quercetin-induced increase in
PTEN and TIMP3 expression (Fig.
4A-C).
Discussion
The renal protective effect of quercetin has been
discussed in a number of studies. Gelen et al (24) reported that quercetin protected renal
function in a rat model of obesity, while Yuksel et al
(25) demonstrated that quercetin
was able to reduce methotrexate-induced oxidative stress in rat
models, suggesting that quercetin can alleviate
methotrexate-induced renal damage and restore renal function. Elbe
et al (26) revealed that
quercetin is able to reduce ciprofloxacin-induced oxidative stress
in rats, suggesting that quercetin may be an effective treatment
for injuries. In the present study, fibrosis was induced in HK-2
cells using TGF-β and it was demonstrated that treatment with 7.5,
15 or 30 mg/ml quercetin induced a significant decrease in the
expression of fibrotic markers. These results suggest that the
anti-fibrotic effect of quercetin may not be dose-dependent, as no
significant difference was observed in the expression of fibrotic
markers between the 15 and 30 mg/ml groups. Taken together, these
results indicate that treatment with an appropriate dosage of
quercetin may effectively alleviate TGF-β-induced fibrosis in HK-2
cells in vitro.
miR-21 has previously been reported to be a
profibrotic microRNA. miR-21-5p expression is associated with the
incidence of renal fibrosis in patients with IgA nephropathy
(27), while an in vitro
study revealed that miR-21 may serve a role in the pathogenesis of
TIF by targeting DDAH1 (28).
Furthermore, Wang et al (29)
demonstrated that miR-21 overexpression can cause renal fibrosis
via the MMP9/TIMP1 signaling pathway in mice, suggesting that
miR-21 has potential as a novel therapeutic target for the
management of diabetic nephropathy. In the present study, it was
demonstrated that miR-21 was upregulated in fibrosis in HK-2 cells,
which is consistent with previous findings. It was also observed
that quercetin treatment induced a significant decrease in the
expression of miR-21, suggesting that miR-21 may serve a role in
the mechanism by which quercetin induces anti-fibrotic effects.
Furthermore, transfection with miR-21 mimics partially inhibited
quercetin-induced anti-fibrotic effects. These results suggest that
the anti-fibrotic effects of quercetin may be achieved in part by
downregulating miR-21, a profibrotic factor.
PTEN and TIMP3 are known to be anti-fibrotic factors
(30–32) and are downregulated in fibrotic renal
tissues. Downregulation of either PTEN (29) or TIMP3 (31) may enhance the degree of TIF.
Bioinformatics has been used to predict PTEN and TIMP3 as direct
targets of miR-21, which has been confirmed using dual-luciferase
reporter assays (32,33). To further explore the mechanism of
miR-21 in quercetin-induced anti-fibrotic effects, the expression
of PTEN and TIMP3 was compared between quercetin-treated and
untreated fibrotic HK-2 cells. The results indicated that quercetin
treatment induced a significant increase in the expression of PTEN
and TIMP3 at the mRNA and protein levels, while transfection with
miR-21 mimics partially inhibited the quercetin-induced PTEN and
TIMP3 upregulation. These results suggest that quercetin
downregulates miR-21 and upregulates anti-fibrotic gene expression,
thereby inhibiting the progression of kidney fibrosis and
protecting renal function.
The present study is not without limitations. Only
cellular experiments were performed; in future studies, the roles
of quercetin and miR-21 should be further explored using animal
models. In conclusion, the results of the present study suggest
that quercetin is able to alleviate TGF-β-induced fibrosis in
tubular epithelial cells by suppressing miR-21. These findings
provide a novel insight into the effects of quercetin as an
anti-fibrotic drug for the treatment of renal fibrosis.
Acknowledgements
Not applicable.
Funding
No funding received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YCao and JH contributed to conception and design of
the study. YCao, JH and JS performed the experiments. LJ conducted
data acquisition, analysis and interpretation. YCong was involved
in data analysis, interpretation and manuscript development. GR
guided the experimental design, reviewed the manuscript and
supervised the study. All authors read and approved the final
version of this manuscript and agreed to be accountable for all
aspects of this study.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhou D, Fu H, Zhang L, Zhang K, Min Y,
Xiao L, Lin L, Bastacky SI and Liu Y: Tubule-derived wnts are
required for fibroblast activation and kidney fibrosis. J Am Soc
Nephrol. 28:2322–2336. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Omata M, Doke Y, Yamada C, Kawashima K,
Sho R, Enomoto K, Furuya M and Inomata N: Hepatocyte nuclear
factor-1β induces redifferentiation of dedifferentiated tubular
epithelial cells. PLoS One. 11:e01549122016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lu H, Chen B, Hong W, Liang Y and Bai Y:
transforming growth factor-β1 stimulates hedgehog signaling to
promote epithelial-mesenchymal transition after kidney injury. FEBS
J. 283:3771–3790. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen SJ, Wu P, Sun LJ, Zhou B, Niu W, Liu
S, Lin FJ and Jiang GR: miR-204 regulates epithelial-mesenchymal
transition by targeting SP1 in the tubular epithelial cells after
acute kidney injury induced by ischemia-reperfusion. Oncol Rep.
37:1148–1158. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jara P, Calyeca J, Romero Y, Plácido L, Yu
G, Kaminski N, Maldonado V, Cisneros J, Selman M and Pardo A:
Matrix metalloproteinase (MMP)-19-deficient fibroblasts display a
profibrotic phenotype. Am J Physiol Lung Cell Mol Physiol.
308:L511–L522. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Meng XM, Zhang Y, Huang XR, Ren GL, Li J
and Lan HY: Treatment of renal fibrosis by rebalancing TGF-β/Smad
signaling with the combination of asiatic acid and naringenin.
Oncotarget. 6:36984–36997. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Muñoz-Félix JM, González-Núñez M,
Martínez-Salgado C and López-Novoa JM: TGF-U/BMP proteins as
therapeutic targets in renal fibrosis. Where have we arrived after
25 years of trials and tribulations? Pharmacol Ther. 156:44–58.
2015.PubMed/NCBI
|
|
8
|
Vega G, Alarcón S and San Martin R: The
cellular and signalling alterations conducted by TGF-β contributing
to renal fibrosis. Cytokine. 88:115–125. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Garzon R, Calin GA and Groce CM: MicroRNAs
in cancer. Annu Rev Med. 60:167–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Han F, Konkalmatt P, Chen J, Gildea J,
Felder RA, Jose PA and Armando I: miR-217 mediates the protective
effects of the dopamine D2 receptor on fibrosis in human renal
proximal tubule cells. Hypertension. 65:1118–1125. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He F, Peng F, Xia X, Zhao C, Luo Q, Guan
W, Li Z, Yu X and Huang F: miR-135a promotes renal fibrosis in
diabetic nephropathy by regulating TRPC1. Diabetologia.
57:1726–1736. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu D, Zhang N, Zhang J, Zhao H and Wang
X: miR-410 suppresses the expression of interleukin-6 as well as
renal fibrosis in the pathogenesis of lupus nephritis. Clin Exp
Pharmacol Physiol. 43:616–625. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Han M, Song Y and Zhang X: Quercetin
suppresses the migration and invasion in human colon cancer caco-2
cells through regulating toll-like receptor 4/nuclear factor-kappa
B pathway. Pharmacogn Mag. 12 Suppl 2:S237–S244. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chan CY, Lien CH, Lee MF and Huang CY:
Quercetin suppresses cellular migration and invasion in human head
and neck squamous cell carcinoma (HNSCC). Biomedicine (Taipei).
6:152016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Song NR, Chung MY, Kang NJ, Seo SG, Jang
TS, Lee HJ and Lee KW: Quercetin suppresses invasion and migration
of H-Ras-transformed MCF10A human epithelial cells by inhibiting
phosphatidylinositol 3-kinase. Food Chem. 142:66–71. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen BL, Wang LT, Huang KH, Wang CC,
Chiang CK and Liu SH: Quercetin attenuates renal
ischemia/reperfusion injury via an activation of AMP-activated
protein kinase-regulated autophagy pathway. J Nutr Biochem.
25:1226–1234. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Erboga M, Aktas C, Erboga ZF, Donmez YB
and Gurel A: Quercetin ameliorates methotrexate-induced renal
damage, apoptosis and oxidative stress in rats. Ren Fail.
37:1492–1497. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kou B, Liu W, Tang X and Kou Q: HMGA2
facilitates epithelial-mesenchymal transition in renal cell
carcinoma by regulating the TGF-β/Smad2 signaling pathway. Oncol
Rep. 39:101–108. 2018.PubMed/NCBI
|
|
19
|
Kinashi H, Falke LL, Nguyen TQ, Bovenschen
N, Aten J, Leask A, Ito Y and Goldschmeding R: Connective tissue
growth factor regulates fibrosis-associated renal
lymphangiogenesis. Kidney Int. 92:850–863. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma J, Zhang L, Hao J, Li N, Tang J and Hao
L: Up-regulation of microRNA-93 inhibits TGF-β1-induced EMT and
renal fibrogenesis by down-regulation of Orai1. J Pharmacol Sci.
136:218–227. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shi B, Wang Y, Zhao R, Long X, Deng W and
Wang Z: Bone marrow mesenchymal stem cell-derived exosomal miR-21
protects C-kit+ cardiac stem cells from oxidative injury through
the PTEN/PI3K/Akt axis. PLoS One. 13:e01916162018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gutsaeva DR, Thounaojam M, Rajpurohit S,
Powell FL, Martin PM, Goei S, Duncan M and Bartoli M:
STAT3-mediated activation of miR-21 is involved in down-regulation
of TIMP3 and neovascularization in the ischemic retina. Oncotarget.
8:103568–103580. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gelen V, Şengül E, Gedikli S, Gür C and
Özkanlar S: Therapeutic effect of quercetin on renal function and
tissue damage in the obesity induced rats. Biomed Pharmacother.
89:524–528. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yuksel Y, Yuksel R, Yagmurca M, Haltas H,
Erdamar H, Toktas M and Ozcan O: Effects of quercetin on
methotrexate-induced nephrotoxicity in rats. Hum Exp Toxicol. Mar
22–2016.(Epub ahead of print). PubMed/NCBI
|
|
26
|
Elbe H, Dogan Z, Taslidere E, Cetin A and
Turkoz Y: Beneficial effects of quercetin on renal injury and
oxidative stress caused by ciprofloxacin in rats: A histological
and biochemical study. Hum Exp Toxicol. 35:276–281. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hennino MF, Buob D, Van der Hauwaert C,
Gnemmi V, Jomaa Z, Pottier N, Savary G, Drumez E, Noël C, Cauffiez
C and Glowacki F: miR-21-5p renal expression is associated with
fibrosis and renal survival in patients with IgA nephropathy. Sci
Rep. 6:272092016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu XJ, Hong Q, Wang Z, Yu YY, Zou X and
Xu LH: MicroRNA21 promotes interstitial fibrosis via targeting
DDAH1: A potential role in renal fibrosis. Mol Cell Biochem.
411:181–189. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang J, Gao Y, Ma M, Li M, Zou D, Yang J,
Zhu Z and Zhao X: Effect of miR-21 on renal fibrosis by regulating
MMP-9 and TIMP1 in kk-ay diabetic nephropathy mice. Cell Biochem
Biophys. 67:537–546. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lan R, Geng H, Polichnowski AJ, Singha PK,
Saikumar P, McEwen DG, Griffin KA, Koesters R, Weinberg JM, Bidani
AK, et al: PTEN loss defines a TGF-β-induced tubule phenotype of
failed differentiation and JNK signaling during renal fibrosis. Am
J Physiol Renal Physiol. 302:F1210–F1223. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kassiri Z, Oudit GY, Kandalam V, Awad A,
Wang X, Ziou X, Maeda N, Herzenberg AM and Scholey JW: Loss of
TIMP3 enhances interstitial nephritis and fibrosis. J Am Soc
Nephrol. 20:1223–1235. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
McClelland AD, Herman-Edelstein M, Komers
R, Jha JC, Winbanks CE, Hagiwara S, Gregorevic P, Kantharidis P and
Cooper ME: miR-21 promotes renal fibrosis in diabetic nephropathy
by targeting PTEN and SMAD7. Clin Sci (Lond). 129:1237–1249. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang N, Zhang CQ, He JH, Duan XF, Wang YY,
Ji X, Zang WQ, Li M, Ma YY, Wang T and Zhao GQ: miR-21
down-regulation suppresses cell growth, invasion and induces cell
apoptosis by targeting FASL, TIMP3, and RECK genes in esophageal
carcinoma. Dig Dis Sci. 58:1863–1870. 2013. View Article : Google Scholar : PubMed/NCBI
|